Bay leaf extract is a chemotaxis repellent for C. elegans
Samuel H. Wu, Anna Amine, Kaya Ben-Efraim, Nicole J. Dye, Massiel Melian, Keira C. Nakamura, Ruhee Nemawarkar, Keshav Saigal, Harmony M. Sosa, Linh T. Vo, Baraa J. Abdelghne, Gwendolyne K. Aguilar, Riley E. Carolan, Ashley N. Carter, Daniela A. Castro-Martinez, Melody Chang

TL;DR
Bay leaf extract repels the nematode C. elegans, with cadinene and eugenol identified as the key compounds causing this repulsion.
Contribution
The study identifies specific compounds in bay leaf extract that repel C. elegans and demonstrates their use in an educational setting.
Findings
C. elegans avoids bay leaf extract in chemotaxis assays.
Cadinene and eugenol are key compounds responsible for the repulsion effect.
The experiments were conducted by undergraduate students in a research course.
Abstract
Plants synthesize compounds that modulate animal nervous systems through various mechanisms, but the key interactions remain understudied. We used chemotaxis assays with the nematode Caenorhabditis elegans to test if plant extracts can be detected by the worm nervous system and which compounds induce behavioral responses. We found that C. elegans avoid the extract of bay leaves ( Laurus nobilis ). Subsequent testing of known bay leaf compounds identified cadinene and eugenol as key molecules that may mediate the repulsion effect. These experiments were conducted by undergraduate students in an upper-division laboratory course, providing practical research experiences and new insights into plant-animal interactions.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
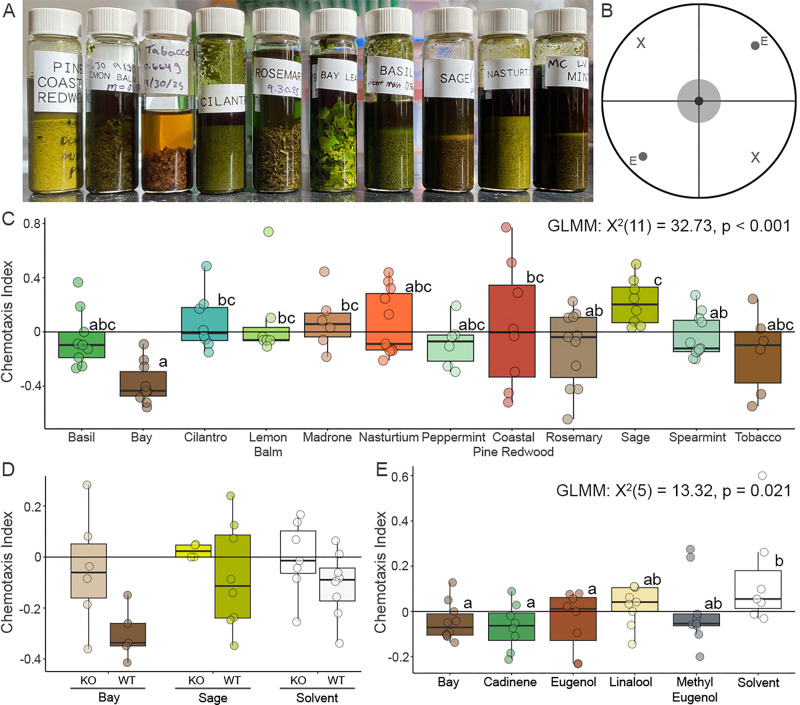 Figure 1
Figure 1|
Strain |
Genotype |
Source |
|
|
wild type reference strain, formally known as
|
Caenorhabditis Genetics Center (CGC) at the University of Minnesota |
|
|
|
Miriam Goodman Lab at Stanford University |
- —National Science Foundation (United States)https://ror.org/021nxhr62
- —Camille and Henry Dreyfus Foundation (United States)https://ror.org/019p8h045
- —National Science Foundation (United States)https://ror.org/021nxhr62
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · Allelopathy and phytotoxic interactions · Chemical synthesis and alkaloids
Description
Plants have many secondary metabolites that influence the nervous system of animals (Nishida, 2014), both to attract pollinators and to repel herbivores (Metcalf and Kogan, 1987; Nerio et al., 2010). The neurogenetic mechanisms of these effects are relatively unclear due to the complexity of chemical extracts and the limited genetic tools available for non-traditional research organisms. Here, we used chemotaxis assays in * Caenorhabditis elegans * to identify plant-derived products that act as neuronal actuators. This nematode has tractable genetics and well-described chemosensory behavior (Bargmann, 2006), which enables incorporation into undergraduate course research experiences. We hypothesized that edible plants commonly used by humans would contain compounds that interact with heterospecific nervous systems. The ability to detect plant compounds occurs in * C. elegans * (Fryer et al., 2024) , who thrive in compost. We tested our hypothesis using (1) * C. elegans * chemotaxis assays to identify plant extracts that interact with animal nervous systems and (2) carried out these experiments in an undergraduate laboratory classroom to enable hands-on research experiences in chemical ecology.
We asked if commonly available plants have natural products that influence * C. elegans * chemotaxis behavior. We found that wild-type worms ( CGC1 , wild type reference strain formally known as PD1074 ) responded differently to plant compounds (GLMM: χ²(11) = 32.731, p < 0.001). Bay leaf extract was more repulsive than cilantro (t(88) = -3.696, p adj = 0.013), lemon balm (t(88) = -3.374, p adj = 0.024), madrone (t(88) = -3.193, p adj = 0.026), coastal pine redwood (t(88) = -3.056, p adj = 0.033), and sage (t(88) = -4.709, p adj = 0.001). In addition to being more attractive than bay, sage extract is more attractive to wild-type worms compared to spearmint (t(88) = 3.200, p adj = 0.026) and rosemary (t(88) = -2.889, p adj = 0.046). All other plant extract pairwise comparisons were not significantly different (p adj
0.05). These results are consistent with other studies demonstrating repellent activity of bay leaves to mosquitoes (Oktiansyah et al., 2022), beetles (Saim and Meloan, 1986), and ants (Ikechi-Nwogu et al., 2025). On the other hand, sage is a repellent to mites (Laborda et al., 2013), which is inconsistent with our results. However, this plant extract comparison experiment lacked a solvent control to determine whether sage is overall attractive, and this control was incorporated into subsequent experiments.
Chemosensation in * C. elegans * can be ablated by removing specific ion channels, including TAX-4 (Kumatsu et al., 1996) and OSM-9 (Colbert et al., 1997) *, * as chemosensory neurons express one or both of these genes. We tested the chemotaxis responses of * tax-4 ( p678 ); osm-9 ( ky10 ) * double mutants (Fryer et al., 2024) and wild-type worms ( CGC1 ) to bay and sage extracts. The worm strains differed in their response to the extracts (GLMM: strain: χ²(1) = 9.609, p = 0.002; compound: χ²(2) = 8.782, p = 0.012), although the interaction effect was not significant, likely due to low sample size. Regardless, bay extract continued to repel worms compared to solvent (t(31) = -2.312, p adj = 0.041) and sage (t(31) = -2.768, p adj = 0.028), whereas sage and solvent did not differ (p adj
0.05). Our data suggests that bay leaf extract is consistently repulsive to
C. elegans * , whereas sage does not elicit a response significantly different from solvent. It is likely this effect is due to chemosensory function through * osm-9 * and/or * tax-4 * ion channels, although more experiments with increased sample size and single mutant strains are necessary to confirm.
As bay extract consistently produced a repellent effect in * C. elegans * , we next tested which natural products in bay extract were responsible for this effect. Previous studies on the chemical composition of bay leaves detected cadinene, eugenol, linalool, and methyl eugenol, among others (Biondi et al., 1993). Wild-type ( CGC1 ) worms responded differently to these compounds (GLMM: compound: χ²(5) = 13.319, p = 0.021). Bay leaf extract was more repulsive than solvent (t(44) = -3.042, p adj = 0.030), consistent with our previous two experiments. In addition, cadinene (t(44) = -2.779, p adj = 0.040) and eugenol (t(44) = -3.053, p adj = 0.030) are more repulsive than solvent, whereas linalool and methyl eugenol were not different than solvent (p adj
0.05). Cadinene is a sesquiterpene produced by a wide range of essential oil-producing plants and has antifungal activity (Kundu et al., 2013). Eugenol is also present in some essential oil-producing plants and, consistent with our results, is a repellent for mites (Zeringóta et al., 2013) and ants (He et al., 2023). Eugenol has also been proposed as a natural insecticide and antimicrobial agent that can be incorporated into textiles (Wen et al., 2024). Indeed, linalool and methyl eugenol have been described as insect repellents (Müller et al., 2009; Xu et al., 2015). Testing a range of concentrations in
C. elegans * chemotaxis assays would help determine if these compounds also impact nematode behavior.
In summary, our study shows that bay leaf extract is a chemotaxis repellent for * C. elegans * , and this response may be due in part to cadinene and eugenol. We cannot rule out that the other plants tested here can impact * C. elegans * chemotaxis behavior given different processing steps or concentrations and further research is warranted. Given our results here, coupled with the genetic tools available for * C. elegans * , these chemotaxis assays may be useful for understanding the neurogenetic mechanisms underpinning the behavioral effects of plant compounds across organisms.
Methods
Worm strains
Nematode strains ( * Caenorhabditis elegans * ) were obtained from the Caenorhabditis Genetics Center (CGC) at the University of Minnesota or from the lab of Miriam Goodman at Stanford University. Worms were maintained in 20°C incubators and synchronized by bleaching adults. The remaining eggs were grown on nematode growth media plates spread with OP50 * Escherichia coli * (Stiernagle, 2006). Hatched eggs were kept at 20°C for 2-5 days, depending on the strain, when the population reached a young adult stage and were used for chemotaxis assays.
Plant extracts and compounds
Students selected a range of plants from either grocery stores or local gardens. The leaves were dried for 24-72 hours at 60°C. The plants were then crushed with a mortar and pestle and then placed into a pre-weighed glass vial. Then, methanol was added to equal the volume of the plant material (1:1 methanol:plant volume) and incubated at room temperature for 3 days ( Figure 1A ). The methanol was then filtered to remove plant debris, moved to a new glass vial, and then evaporated under nitrogen gas. The remaining plant extract residue was resuspended in dimethyl sulfoxide (DMSO) at a concentration of 10 mg/mL for use in chemotaxis assays.
We also used commercially available phytochemicals in this study, including cadinene (Millipore Sigma Cat # CH6H8C2C47CC-5G), eugenol (Millipore Sigma Cat # E51791-5G), linalool (Millipore Sigma Cat # L2602-5G), and methyl eugenol (Millipore Sigma Cat # W247502-SAMPLE-K). All compounds were dissolved in DMSO and used at 20 mM final concentration.
Chemotaxis assays
Undergraduate students conducted chemotaxis assays in a laboratory classroom as previously described (Alfonso et al., 2023; Lopez et al., 2024; Gaerlan et al., 2025) following standard procedures (Bradon et al., 2024). Students were unaware of the chemical stimulus or worm strain being tested until class data was submitted to the instructor. Chemotaxis plates [5mM KPO 4 (pH 6), 1mM CaCl 2 , 1mM MgSO 4 , 2% agar] were divided into four quadrants ( Figure 1B ) and 5 μL plant extract (concentration unknown) or compound (20 mM) were placed on dots located in two non-adjacent quadrants (E, experimental), while 5 μL of DMSO (solvent) was placed on X marks in the other two quadrants. Solvent was also placed in the experimental quadrants of the control plates, leaving all four quadrants containing DMSO. Plates were incubated for 30 min. During this time, worms were removed from the growth plates using chemotaxis assay buffer [5mM KPO 4 (pH 6), 1mM CaCl 2 , 1mM MgSO 4 ] and moved to a microfuge tube. The worms were washed two to three times with chemotaxis buffer. Then, 2 μL 0.5 M sodium azide solution was applied to each of the quadrant spots to paralyze the worms at their choice locations. Immediately after, roughly 100 worms were pipetted into the center of each chemotaxis plate, where they were allowed to roam for 30 min undisturbed. Worms were manually counted under a dissecting microscope using a tally counter. Worms in the center area of overlapping quadrants were not counted to avoid including dead worms in the dataset.
Data analysis
The Chemotaxis Index (CI) was calculated for each plate, where CI = (Number of worms in the two experimental quadrants – Number of worms in the two solvent quadrants) / Total number of worms. Plates were removed from the dataset if they had less than 30 or more than 300 worms to reduce large differences in worm abundance and to avoid statistical artifacts due to variability in technique among students. Plates were also removed if a student noted a technical error in the plate setup, such as mistakes in the placement of worms or compounds.
Data analysis and visualization were performed in R version 4.3.0 (R Core Team, 2024) in RStudio version 2025.09.1+401. We used the glmmTMB package version 1.1.7 (Brooks et al., 2017) for statistical analyses. We used generalized linear mixed models and followed the model with the Anova.glmmTMB function using either Type II or Type III “marginal” sums of squares for reported statistical values. Appropriate model diagnostics were confirmed with the DHARMa package version 0.4.6 (Hartig, 2024). The number of worms in the experimental quadrants divided by the total number of worms on the plate was used as the dependent variable. For the experiments involving only wild-type worms, the compound was the independent variable. For experiments involving two worm strains, compound, worm strain, and their interaction were the independent variables. Post hoc analyses were performed using emmeans version 1.8.6 (Lenth et al., 2024) with false discovery rate (fdr) adjustment of p-values to account for multiple comparisons. Boxplots were generated using the ggplot2 version 3.4.3 (Wickham, 2016) package.
Classroom pedagogy
The experiments in this study were performed over four laboratory sessions. These sessions were preceded with one training session where students learned how to conduct * C. elegans * chemotaxis assays using known attractant (isoamyl alcohol) and repulsive (carvone) compounds. One laboratory session involved grinding dried plants and extracting compounds. The other three laboratory classroom sessions were used to run chemotaxis assays. Weekly homework included reading relevant literature, analysis and visualization of data collected by the entire class, and writing individual drafts of a journal-style article, which were combined into this article by the instructors. Assignments were graded as complete/incomplete and students received detailed feedback at each stage. All student co-authors edited and approved of the final manuscript.
Reagents
Worm Strains
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alfonso SA Arango Sumano D Bhatt DA Cullen AB Hajian CM Huang W Jaeger EL Li E Maske AK Offenberg EG Ta V Whiting WW Adebogun GT Bachmann AE Callan AA Khan U Lewis AR Pollock AC Ramirez D Bradon N Fiocca K Cote LE Sallee MD Mc Kinney J O'Connell LA 2023314 Argentine ant extract induces an osm-9 dependent chemotaxis response in C. elegans.Micro Publ Biol 202310.17912/micropub.biology.00074537008729 PMC 10051032 · doi ↗ · pubmed ↗
- 2Bargmann CI 20061025 Chemosensation in C. elegans.Worm Book 12910.1895/wormbook.1.123.118050433 PMC 4781564 · doi ↗ · pubmed ↗
- 3Biondi Daniela Cianci Paola Geraci Corrada Ruberto Giuseppe Piattelli Mario 1993111 Antimicrobial activity and chemical composition of essential oils from sicilian aromatic plants Flavour and Fragrance Journal 860882-573433133710.1002/ffj.2730080608 · doi ↗
- 4Bradon Nicole Fiocca Katherine D. Sallee Maria Cote Lauren A. Shaykevich Daniel A O'Connell Lauren 2023922 Chemical ecology in the classroom: chemotaxis assay using C. elegans v 110.17504/protocols.io.e 6nvwde 9zlmk/v 1 · doi ↗
- 5Brooks Mollie,E. Kristensen Kasper Benthem Koen,J.,van Magnusson Arni Berg Casper,W. Nielsen Anders Skaug Hans,J. Mächler Martin Bolker Benjamin,M. 2017 glmm TMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling The R Journal 922073-485937837810.32614/rj-2017-066 · doi ↗
- 6Colbert HA Smith TL Bargmann CI 1997111 OSM-9, a novel protein with structural similarity to channels, is required for olfaction, mechanosensation, and olfactory adaptation in Caenorhabditis elegans.J Neurosci 17210270-647482598269933440110.1523/JNEUROSCI.17-21-08259.1997 PMC 6573730 · doi ↗ · pubmed ↗
- 7Fryer E Guha S Rogel-Hernandez LE Logan-Garbisch T Farah H Rezaei E Mollhoff IN Nekimken AL Xu A Seyahi LS Fechner S Druckmann S Clandinin TR Rhee SY Goodman MB 2024627 A high-throughput behavioral screening platform for measuring chemotaxis by C. elegans.P Lo S Biol 2261544-9173 e 3002672 e 300267210.1371/journal.pbio.300267238935621 PMC 11210793 · doi ↗ · pubmed ↗
- 8Gaerlan M Carrillo M Ceva S Chundi S Diallo B Fong JN Huang K Jackson J Padilla J Quintana L Santa Maria K Sarkisian SM Sequeira PR Tatlock EU Baker PR Bachmann L Park S Perez MJ Phipps ME Sharma SN Soto-Hernandez Y Juarez BH Mena C Morales G Gonzalez M Fiocca K Bradon N Madrzyk M O'Connell LA 2025531 Velvety tree ant extract is a chemotaxis repellent for C. elegans.Micro Publ Biol 202510.17912/micropub.biology.00153140535527 PMC 12175000 · doi ↗ · pubmed ↗