Comparative analysis of placental transmission mechanisms for Dengue and Zika viruses: outcomes and future directions
Indrani Mukherjee, Neil Ferland, Katelyn Tran Nguyen, Sara Moghaddam Adames, Helena Solo-Gabriele, Joshua Anzinger, Ivan A. Gonzalez, Ruby Dhar, Subhradip Karmakar, Rana Chakraborty

TL;DR
This paper compares how Dengue and Zika viruses cross the placenta during pregnancy, highlighting their transmission mechanisms and effects on maternal and fetal health.
Contribution
The paper provides a comparative analysis of placental transmission mechanisms and clinical outcomes of Dengue and Zika viruses in pregnancy.
Findings
Dengue virus can lead to pre-term birth and thrombocytopenia in infants.
Zika virus infects placental cells and causes Congenital Zika Syndrome with microcephaly.
Both viruses disrupt placental signaling pathways like JAK/STAT and mTOR.
Abstract
Zika and Dengue viruses are arboviral pathogens capable of crossing the placental barrier, representing major global health risks for maternal and fetal outcomes. In this narrative review, we compare their epidemiology, clinical consequences in pregnancy, and underlying mechanisms of vertical transmission. Emerging molecular insights are highlighted, including disruptions to placental signaling pathways such as JAK/STAT and mTOR, and strategies to evade Hofbauer cells and the immune system. A comparative analysis of these processes underscores a critical need for improved understanding of placental pathophysiology, immune regulation, and molecular pathways of transmission. Identifying such mechanisms may promote vaccine development, improved diagnostics, and therapies to reduce adverse outcomes in the mother-infant dyad during maternal infection. Vertical transmission of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
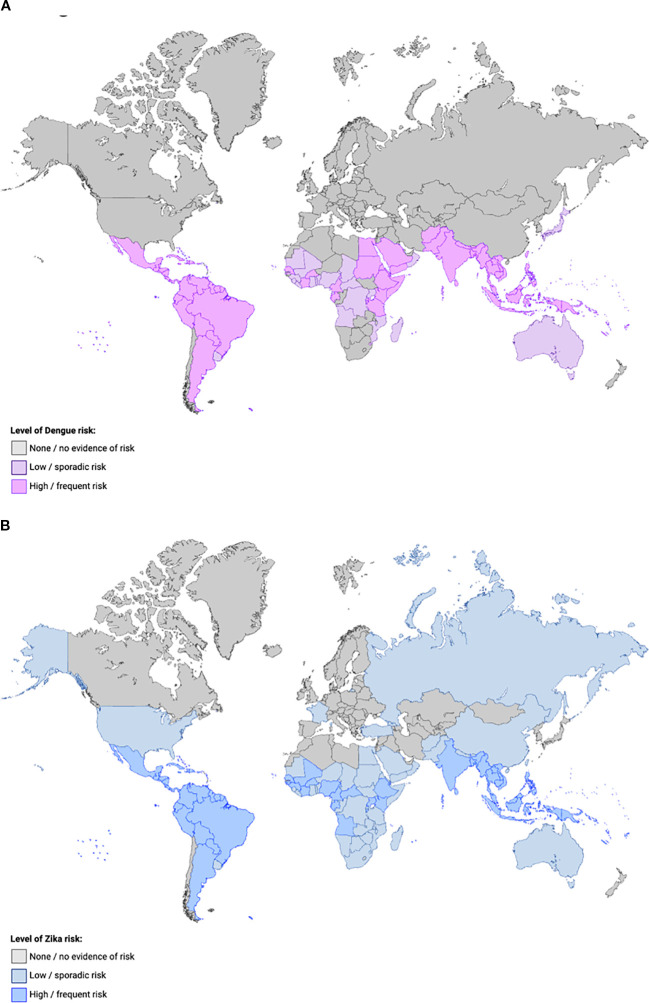 Figure 1
Figure 1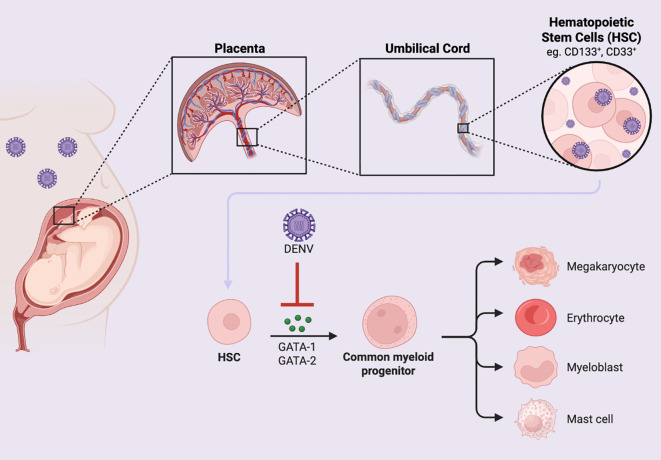 Figure 2
Figure 2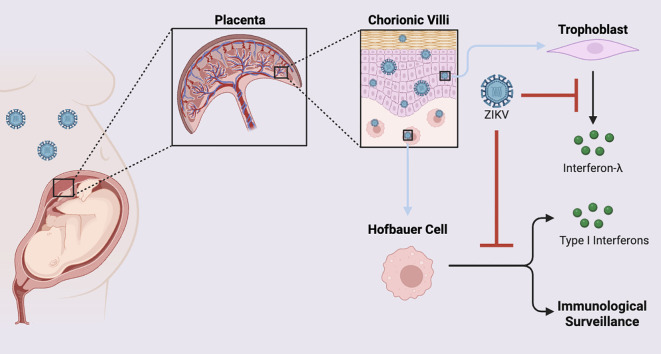 Figure 3
Figure 3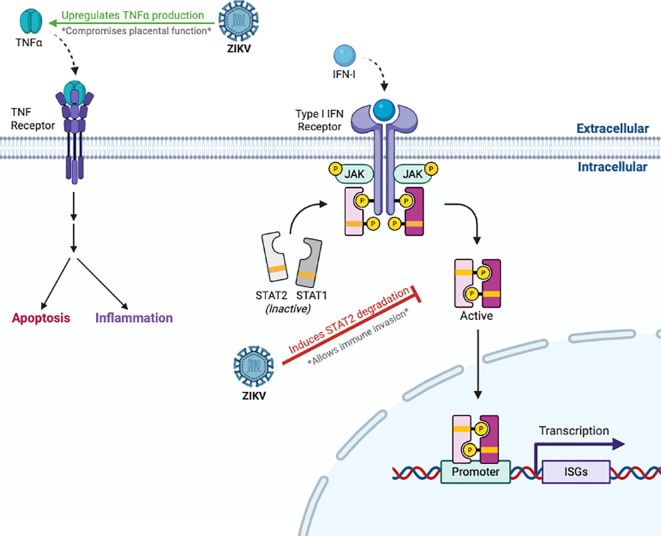 Figure 4
Figure 4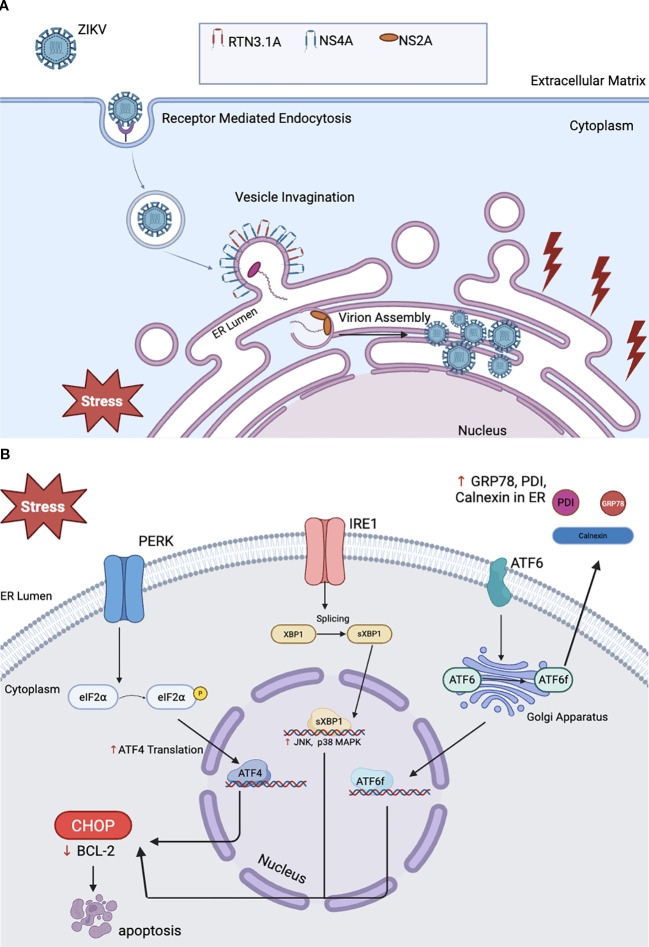 Figure 5
Figure 5| Factors | DENV | ZIKV |
|---|---|---|
| Innate Immunity | Major immune cells involved in the pro-inflammatory phase of pregnancy include dendritic cells, decidual macrophages, and NK cells. Later shifts to anti-inflammatory with Hofbauer cells and Tregs. Hofbauer cells are highly permissive ( | Trophoblasts generate type III interferons (IFN-λ); Hofbauer cells act as reservoirs for viral loads. Endothelial cells and placental fibroblast cells are also susceptible ( |
| Cytokine Response | Decidual macrophages secrete TNF-α and IL-6 in a pro-inflammatory environment, while producing IL-10 in the anti-inflammatory phase ( | Pro-inflammatory cytokines (TNF-α, IL-6) disrupt placental function ( |
| Maternal Antibodies | Maternal neutralizing IgG antibodies protects infants, but declining levels can lead to neonatal death from DENV ( | Maternal neutralizing antibodies may protect, but cross-reactive/non-neutralizing antibodies enhance ZIKV infection (ADE). Antibodies may also form immune complexes in fetus ( |
| Immune Evasion Mechanisms | FcyR-mediated pathway to silently enter and replicate in immune cells. Hofbauer and cord blood cells serve as a passage allowing DENV to bypass the immune system ( | Autophagy and exosome-mediated pathways for viral spread across placenta. Antiviral pathways are downregulated, and immunological tolerance genes are upregulated in the virus’s modulation of placental immunity gene expression ( |
| Key Outcomes/Risks | Infant DENV due to decrease in maternal antibodies; vertical transmission through cord blood stem cells ( | Congenital ZIKV Syndrome (CZS), microcephaly, neurological damage, impaired neurodevelopment ( |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMosquito-borne diseases and control · COVID-19 Impact on Reproduction · Viral Infections and Outbreaks Research
Introduction
1
The placenta functions as an immunological barrier protecting the developing fetus from pathogenic infections while simultaneously facilitating essential maternal-fetal exchange of nutrients, gases, and metabolic substrates through specialized syncytiotrophoblast (STB) and cytotrophoblast (CTB) populations, supported by a complex array of maternal decidual immune cells (1). However, certain viral pathogens have evolved sophisticated immune evasion mechanisms that enable placental cell infection and vertical transmission to the developing fetus (2). Trophoblast invasion occurs within a highly immunologically active maternal-fetal interface (MFI) characterized by a specific immune cell composition, which includes: 70% decidual natural killer (dNK) cells, 20-25% villous macrophages (Hofbauer cells), 3-10% T lymphocytes, and 1.7% dendritic cells, with decidual cytotoxic T cell and macrophage populations exhibiting dynamic fluctuations throughout gestation (3–6). The persistent presence of NK cells in decidual tissue during the first trimester and throughout pregnancy reflects their essential role in establishing immunotolerant conditions, while maintaining antimicrobial surveillance capabilities (7). Despite these robust immune defenses, Zika (ZIKV) and Dengue (DENV) and additional viruses such as human cytomegalovirus (HCMV) and HIV, have evolved to circumvent placental immune barriers, resulting in vertical transmission with fetal infection (8).
Both ZIKV and DENV are public health threats in tropical and subtropical regions and transmitted primarily through Aedes species mosquitos. While Aedes aegypti and Aedes albopictus are the most common vectors, other Aedes species such as Aedes scutellaris, Aedes polynesiensis, Aedes malayensis, and Aedes mediovittatus are known to be competent viral vectors for both ZIKV and DENV (9–12). Both viruses exhibit varying capacities for transplacental transmission associated with adverse pregnancy outcomes (13, 14). ZIKV exhibits well-documented tropism for placental cell populations, particularly in CTBs, STBs, and Hofbauer cells, while DENV has been detected in placental and fetal tissue (15–18).
ZIKV infection during pregnancy (19, 20), especially during the first trimester, is associated with congenital Zika syndrome (CZS), characterized by microcephaly, neurodevelopmental abnormalities, and fetal malformations (13, 14). These same clinical effects have been observed during ZIKV co-infection with HIV (21). Studies have documented transplacental transmission of ZIKV by identifying viral proteins and RNA in placental tissue at different stages of gestation, associated with an increase in placental abnormalities (22, 23). Maternal DENV infection is associated with preterm birth, low birth weight, but without an established pattern of congenital anomalies (24, 25). DENV nonstructural (NS) proteins have been detected in placental and umbilical cord cells (26). The global DENV burden increased 8-fold between 2000–2019 according to WHO data, with expanding geographic distribution into previously non-endemic regions including Europe and continental United States (27). A critical factor influencing orthoflavivirus placental transmission involves antibody-dependent enhancement (ADE), wherein pre-existing maternal antibodies from prior DENV infections paradoxically facilitate ZIKV cellular entry through an Fc receptor-mediated mechanism (28–30). Immunological cross-reactivity between DENV and ZIKV can potentially increase infectious susceptibility in pregnant women leading to adverse maternal-fetal health outcomes.
Given the escalating incidence of orthoflavivirus epidemics, comprehensive elucidation of transplacental transmission mechanisms represents a critical research priority to promote development of targeted therapies, prophylactic vaccines, and evidence-based public health interventions. This review provides a comparative evaluation of DENV and ZIKV placental cell tropism, maternal-fetal immunological interactions, clinical outcomes, and summarizes preventive approaches, while identifying substantial knowledge gaps and delineating future research priorities.
The general epidemiology of DENV and ZIKV infections
2
DENV and ZIKV are single-stranded RNA arboviruses belonging to the Flaviviridae family in the genus Orthoflavivirus and are transmitted by Aedes mosquitoes (31). DENV was first isolated in 1943 in Japan and consists of four different serotypes (DENV1-4). The first isolated serotype was DENV-1, with DENV-2 being isolated two years later in Hawaii (32). After the first isolation, viral mapping showed that DENV was solely endemic to tropical and subtropical climates. However, as global travel became more common and climates changed, DENV infections spread to areas where the virus was not endemic. While all four serotypes were endemic to South America and Southeast Asia, cases were documented in continents with similar climates, such as Africa and Australia (Figure 1A). However, endemic transmission has also been reported in North America and Europe (32). ZIKV was first isolated from a rhesus monkey in the Zika Forest in Uganda in 1947, and for the first time outside of Africa in 1966 in Malaysia. Surveys conducted in the mid 1900’s suggested that this virus was widespread in both Africa and Asia before the time of discovery (35) (Figure 1B).
World map demonstrating the epidemiology of DENV (A) and ZIKV (B). Countries are shaded according to the estimated level of transmission risk of DENV ((A) purple) or ZIKV ((B) blue) from 2025 CDC data (33, 34): grey indicates little or no evidence of risk ((A) no reported DENV cases in the past decade; (B) not known to have mosquitoes that transmit ZIKV), light colors indicate low or sporadic risk ((A) evidence of at least 1 locally acquired DENV case in the past decade; (B) known to have mosquitoes that transmit ZIKV but no ZIKV cases), and darker colors indicate high or frequent risk ((A) evidence of more than 10 locally acquired DENV cases in the past decade; (B) known to have mosquitoes that transmit ZIKV but no ZIKV cases).
The clinical impact of arboviral infection during pregnancy
3
Throughout gestation, maternal physiology undergoes extensive immunological and physiological modifications to establish immune tolerance toward the semi-allogeneic fetus, which would ordinarily trigger alloimmune rejection responses from paternal antigenic components. These immune adaptations, essential for successful fetal accommodation and development, compromise maternal immunocompetence and increase susceptibility to viral pathogens (36). This section summarizes the clinical consequences of maternal DENV, ZIKV, or coinfection in pregnancy.
Dengue viral infection in pregnancy is associated with dengue hemorrhagic fever, dengue shock syndrome, stillbirth, and preterm birth
3.1
DENV seroprevalence among pregnant women in endemic areas often surpasses 50%, although diagnostic challenges, particularly in the early phase of infection and during co-infection, hinder appropriate risk assessment and response (37). The clinical manifestations of DENV range from asymptomatic or mild flu-like symptoms to organ failure (38). In pregnant women, DENV infection can be more marked with an increased risk of stillbirth (36). The clinical manifestations of severe maternal dengue include dengue hemorrhagic fever (DHF) and dengue shock syndrome (DSS). Both conditions increase the likelihood of maternal ICU admissions and obstetric interventions (39). Recent systematic reviews and two meta-analyses projected a preterm birth risk of 18–25% and a stillbirth risk at 5–7%, with greater severity associated with high viral load (40, 41). These findings, considered together, underscore an urgent need for gestational surveillance, vector control, improved diagnosis, and maternal immunization strategies to reduce the impact of DENV during pregnancy.
Megakaryopoiesis disruption and thrombocytopenia during DENV infection
3.2
The main molecular pathway impacted by DENV infection is the PI3K/AKT/mTOR pathway, which plays a role in cell survival, maturation, and megakaryocyte development (42). The activation of PI3K and AKT regulates platelet production in megakaryocytes and promotes cell survival by inhibiting apoptosis. Several transcription factors are involved in platelet production by megakaryocytes. These include GATA-1 and GATA-2, which regulate polyploidization, cell cycle progression, and expression of genes specific to megakaryopoiesis through the downstream effector, STAT1. The other is NF-E2, which is required for megakaryocyte development and maturation, and for determining the stages of megakaryopoiesis (42). DENV infection in megakaryocyte cell lineages impairs AKT activation by first impacting PI3K, an upstream signaling molecule of AKT, reducing the expression of both regulator (p85) and catalytic units (p110β). DENV infection also results in decreased activation of the downstream effector proteins P70-S6 kinase and PKC-α, which affects the function of mTORC1 and mTORC2. The downstream impact of DENV infection on signaling and effector proteins in this pathway prevents expression of GATA-1, GATA-2, and NF-E2 transcription factors, impairing megakaryopoiesis (Figure 2). DENV infection also decreases expression of the anti-apoptotic protein Bcl-2, providing further evidence that dengue can stimulate apoptosis in megakaryotes (42). These mechanism may explain how DENV causes thrombocytopenia, DHF, and DSS in infected individuals.
DENV downregulates hematopoiesis during perinatal infection. DENV can be detected in the umbilical cord of a DENV-infected pregnant person. DENV can impair the differentiation of HSCs into myeloid lineages through inhibition of the transcription factors, GATA-1 and GATA-2.
Infant outcomes of DENV infection in pregnancy include low birth weight, and intrauterine growth restriction
3.3
Recent DENV outbreaks from Brazil, Southeast Asia, and India document that symptomatic maternal DENV in pregnancy is associated with intrauterine growth restriction (IUGR), low birth weight, and newborn thrombocytopenia (41, 43, 44). The evidence demonstrating the ability of DENV to be vertically transmitted from mother to baby during pregnancy is not as robustly supported as it is for ZIKV. However, rare cases document DENV in neonates only days after birth, in the absence of mosquito bites (45, 46). In these cases of vertical transmission when maternal DENV infection was documented peripartum, DENV RNA was detected in cord blood, placental tissue, and infant serum (47). Other studies have provided additional circumstantial evidence to support vertical transmission by detecting dengue NS proteins in the neonatal serum of newborns (48–50). Maternal anti-DENV antibodies could influence newborn immunological outcomes; in rare cases, ADE has been proposed as a mechanism for worsening maternal or infant illness (51).
Diagnostic challenges to neonatal dengue recognition in postnatal life
3.4
A comprehensive case series involving 32 neonates with confirmed DENV infection, the largest and only multicenter neonatal dengue case study reported to date, demonstrated the substantial prevalence of diagnostic errors in neonatal dengue recognition, with 25% of cases initially misdiagnosed as neonatal sepsis and 12.5% erroneously attributed to immune thrombocytopenia (52). The mean symptom onset occurred at 7 days postpartum, creating an extended temporal window for misdiagnosis and inappropriate therapeutic interventions. Although no neonates developed severe dengue manifestations, the clinical presentation included petechiae in 87.5% of cases, hepatomegaly in 75%, and pharyngeal haemorrhage in two patients. This report underscores both the frequency of diagnostic misattribution and the diagnostic challenges associated with neonatal dengue. Furthermore, serological profiles varied significantly with some neonates testing positive for both NS1 antigen and IgM antibodies while others exhibited isolated marker positivity. In addition, nearly 20% of confirmed cases were asymptomatic despite virological confirmation. A case study conducted between October-November 2021 during a DENV epidemic at a single hospital in Pakistan (53) documented five neonates presenting with clinical symptoms of neonatal sepsis. All five tested positive for NS1, three were diagnosed with DENV fever, one with DHF, and the other with DSS. These reports highlight how neonatal DENV infection cryptically mimics neonatal sepsis, underscoring the need to incorporate routine consideration of DENV infection in neonates presenting with sepsis-like symptoms in areas and timeframes when DENV is known to be endemic.
The clinical manifestations Congenital Zika Syndrome
3.5
In contrast to DENV, ZIKV infection during pregnancy can result in Congenital Zika Syndrome (CZS). The teratogenic potential of ZIKV especially in the first trimester, was documented by an outbreak in Brazil with subsequent spread throughout the Americas between 2015 and 2017 (54). Latent viral circulation and underdiagnosis in asymptomatic individuals made surveillance and prompt intervention challenging (55). Regional differences and the need for improved prenatal screening were highlighted by a major retrospective study conducted in French Guiana that reported an incidence of ZIKV infection in pregnancy as high as 188 per 1,000 (56). The impact of early detection and vector control were underscored by cohort data from investigators in Northeastern Thailand and Brazil, who noted that even mild or asymptomatic maternal infection resulted in adverse neonatal outcomes (57, 58).
ZIKV has a particular capacity among orthoflaviviruses to cross the placenta and target fetal neural progenitor cells. Cohort studies from Brazil, Colombia, and Puerto Rico revealed a spectrum of clinical manifestations ranging from fetal loss to severe neurological disability in postnatal life. Specific clinical manifestations include fetal brain disruption sequence (59) characterized by microcephaly, cerebral calcifications, structural brain abnormalities, and neurodevelopmental delay. Other manifestations of CZS include ocular abnormalities, and limb and joint contractures (60, 61). Though risks are present throughout gestation, studies show that first-trimester infection during embryogenesis is strongly related to adverse fetal outcomes. ZIKV RNA has been detected in amniotic fluid, and fetal brain tissue (62, 63). ZIKV and DENV circulate simultaneously in many regions. By utilizing ADE, maternal DENV immunity could promote ZIKV proliferation exacerbating placental damage (64–66).
Neonatal ZIKV transmission and the impact on infant neurodevelopment
3.6
Neonatal ZIKV acquisition occurs through three primary transmission routes: transplacental vertical transmission during gestation, perinatal exposure during the intrapartum and immediate postpartum periods, and postnatal acquisition including breastfeeding-mediated transfer (67). While CZS has garnered significant attention, contemporary studies demonstrate that neonates with in-utero ZIKV exposure exhibit ongoing neuroinflammatory processes and progressive synaptic dysfunction that extend beyond the classical CZS phenotype.
The persistent neuropathological manifestations of ZIKV in neonatal populations can manifest as a heterogenous spectrum of persistent neurodevelopmental and neurologic outcomes. Benazzato et al. (68) demonstrated through analysis of brain tissue specimens from infants with CZS that vertical ZIKV transmission induces sustained neuroinflammatory responses that disrupt synaptic function. By extrapolation, these data provide evidence for the long-term neurological consequences of perinatal ZIKV exposure and underscore the need for continuous neurodevelopmental and neurological monitoring for affected neonates including those who appear asymptomatic at birth. In addition, recent studies have evaluated neurological and ophthalmologic symptoms in newborns exposed to ZIKV. According to a case report from Puerto Rico, a premature baby was later discovered to have ocular defects linked to ZIKV after being initially misdiagnosed with a different congenital condition (69, 70). This report highlights the ongoing need for clinicians, particularly in endemic areas and for individuals with travel history to affected regions, to consider ZIKV in the differential diagnosis when neonates exhibit unexplained developmental or neurological abnormalities.
The association of Dengue and Zika virus infections with Guillain Barre syndrome
3.7
There is a strong association between ZIKV and DENV infection and the development of Guillain-Barré syndrome (GBS). GBS is an immune-mediated disease of the peripheral nerves (71) and the most common cause of acute neuromuscular paralysis (72). A case-control study conducted in Puerto Rico in 2016 during a ZIKV epidemic estimated the risk association between ZIKV infection and GBS development (73). There were 47 case-patients enrolled, and 39 had a confirmed diagnosis of GBS (83%). One of the risk factors identified for GBS development was acute ZIKV infection within two months prior to GBS onset, which occurred in 23% of the cases (73). Similar results were noted in a study of 68 GBS patients in Colombia in 2015–2016 during a ZIKV outbreak (74). Sixty-six patients had symptoms of ZIKV infection prior to GBS onset, with the median period between ZIKV symptoms and GBS onset being 7 days. 42 of these 66 individuals (64%) were positive for ZIKV by RT-PCR. While these studies were limited by small sample sizes, they demonstrated an association between ZIKV infection and the onset of GBS (74). An association between DENV infection and GBS has been documented but is less common than with ZIKV (75).
The molecular impact of arboviral infections on the placenta
4
To fully understand the impact that arboviruses have on the placenta during pregnancy, it is important to focus on the underlying molecular mechanisms and immune pathways impacted by these viruses. Here, we discuss key cellular pathways associated with DENV and ZIKV infection in pregnancy and how altered pathways correlate with clinical manifestations in neonates following maternal infection.
Vertical transmission represents critical windows for infectious disease acquisition in fetuses and infants (76). Maternal DENV infection induces increased vascular permeability and endothelial dysfunction, compromising placental integrity and facilitating vertical transmission at documented rates ranging from 1.6% to 22.7% (50). ZIKV vertical transmission rates is gestational age-dependent with some studies reporting mean transmission risks as high as 47% influenced by the trimester of maternal infection (77).
The impact of maternal Dengue Virus infection at the maternal fetal interface
4.1
Although the precise mechanisms by which DENV affects placental tissue at the cellular level remain poorly characterized, elucidating viral-induced alterations in cell signaling pathways that contribute to characteristic clinical manifestations is essential for understanding the impact of maternal infection on fetal and neonatal outcomes (78). In severe DENV, a life-threatening pathophysiological mechanism involves enhanced vascular permeability resulting from NS1 viral protein interactions with toll-like receptor (TLR) signaling pathways, causing endothelial tissue damage that precipitates plasma extravasation, systemic hypotension, circulatory collapse, and multi-organ dysfunction (78, 79).
Studies directly examining the impact of DENV infection at the MFI, in contrast, are limited. DENV has been detected in umbilical cord blood (UCB) cells and alters cytokine expression with increased production of IL-6 and IL-8 (80). Increased levels of these cytokines were also detected in the serum of patients diagnosed with DHF. These data suggest HUVECs may act as a target for DENV infection (80). Similar studies in human brain microvascular endothelial cells (HBMECs) demonstrate that DENV can infect and replicate in these cells, while increasing the secretion of the inflammatory cytokines IL-6 and IL-8, associated with an increase in reactive oxygen species (ROS) (81). The induction of ROS that occurs in HBMECs can be extrapolated to placental cells, providing a potential explanation for the increased rates of preeclampsia documented in pregnant women infected with DENV (82, 83). In vitro studies have noted the ability of DENV to infect CD133+ and CD34+ cells, potentially acting as replicative reservoirs (84). Murine studies have demonstrated that DENV can cross the placental barrier and be vertically transmitted under conditions that mimic ADE, providing further, albeit limited, evidence of the vertical transmission of DENV (85). An important aspect of maternal DENV infection in pregnancy is placental damage. Histopathological analyses have documented hypoxia, choriodeciduitis, deciduitis, and intervillositis in DENV-infected placentae (86). Additionally, murine studies have noted that DENV infection can lead to neutrophil infiltration, disrupting the vascular network (87) associated with adverse neonatal and maternal outcomes (88). Comparisons between DENV infection in the placenta can be drawn with SARS-CoV-2. Several studies have identified viral proteins of SARS-CoV-2 and active replication of the virus in-vivo in trophoblast, Hofbauer, and endothelial cells (89–91). The resulting inflammation can result in placental damage (92, 93). Despite placental inflammation, evidence for vertical transmission of SARS-CoV-2, like DENV, is limited (84, 94).
Molecular effects of ZIKV at the placenta that promote perinatal transmission and adversely impact synaptic development
4.2
Maternal and fetal immune responses interact intricately to affect ZIKV’s capacity to penetrate the placenta and infect the growing fetus. The placenta provides an immunological and physical barrier against viral infections. Trophoblasts generate type III interferons (IFN-λ), which aid in thwarting viral invasion (15). ZIKV can infect placental macrophages or Hofbauer cells, which provide an immunological barrier and are involved in immune surveillance. Upon infection, these cells promote viral dissemination throughout the placenta (68). In addition, ZIKV reduces the antiviral effects of type I IFN signaling by suppressing maternal IFN using the ZIKV NS5 protein, which targets the STAT2 host immune defense complex for degradation (95, 96) (Figures 3, 4) This method of immune evasion increases the risk of vertical transmission by enabling the virus to infiltrate the placenta.
ZIKV evasion of placental trophoblast- and Hofbauer cell-mediated immune defenses during perinatal infection. Perinatal ZIKV infection of the chorionic villi can prevent trophoblasts lining the placenta from secreting IFN-λ, allowing for viral invasion. Furthermore, ZIKV can dampen the ability of Hofbauer cells to provide immune surveillance and type I IFN secretion, weakening the placental innate immune defenses and facilitating vertical transmission to the fetus.
Perturbation of host immunity by ZIKV. ZIKV antagonizes IFN-I signaling by targeting STAT2 for proteosomal degradation, suppressing the transcription of ISGs and weakening antiviral defenses. ZIKV also compromises placental function by upregulating TNF-α production, leading to hyperinflammation and apoptosis.
Adverse gestational outcomes following maternal ZIKV infection are mechanistically linked to maternal immune activation, characterized by upregulation of pro-inflammatory cytokines including tumor necrosis factor-alpha (TNF-α) and IL-6. This has been shown to compromise placental physiological function and induce fetal neuroinflammatory responses (68) that adversely shape neurotranscriptional expression negatively influencing synaptic development (68, 97). This inflammatory cascade also enhances placental barrier permeability leading to cerebral edema, complicated further by placental ischemia (98). Infected placental cells release extracellular vesicles containing inflammatory mediators and viral ribonucleic acid, which can traverse to fetal organs and trigger neuroinflammatory cascades. This inflammation-mediated tissue damage may contribute to fetal brain disruption sequence and the severe neurological abnormalities characteristic of CZS (68).
Inhibition of trophoblast syncytialization by ZIKV infection
4.3
Trophoblast syncytialization is key to placental development, as these cells differentiate to form multinucleated syncytiotrophoblasts (STBs) that form the outer layer of the placenta (99). STBs are resistant to ZIKV infection, allowing them to serve as a barrier to transplacental spread (100). However, ZIKV can modulate this protective layer. Studies of ZIKV-infected placental tissue examined changes in paracellular permeability at the STB layer, normally controlled by tight junctions. The expression of tight junction proteins, such as E-cadherin, claudins, and occludin, were measured in addition to paracellular permeability (101). Tight junction protein expression was the same, apart from claudin-4, which was decreased. Additionally, the paracellular permeability of ZIKV-infected placentae was greater than in healthy control samples, suggesting ZIKV can modulate tight junction proteins, allowing for viral spread in the placenta (101).
Zika virus has been shown to prevent the differentiation of trophoblastic stem cells to syncytiotrophoblasts (102). An in-vitro study using placental organoids examined the vulnerability of human trophoblast stem cells to ZIKV infection. Trophoblasts were highly permissive to ZIKV, and resistance to infection increased as the stem cells began differentiating. Expression of AXL and TIM-1 by trophoblast stem cells contributed to the sensitivity of these cells to ZIKV infection. AXL encodes a receptor tyrosine kinase of the TAM receptor family, while TIM-1 produces a phosphatidylserine receptor, both functioning to facilitate ZIKV entry (102–105). The study concluded that ZIKV infection in trophoblast stem cells disrupts differentiation to cytotrophoblasts and syncytialization, providing an explanation for the placental damage often observed during maternal ZIKV infection (102).
ZIKV-mediated teratogenesis through endoplasmic reticulum stress induction
4.4
ZIKV can disrupt embryogenesis by triggering endoplasmic reticulum (ER) stress in both placental trophoblast populations and developing neural tissue (106, 107). Following receptor-mediated endocytosis, ZIKV gains entry into host cells and localizes to the ER, where viral genome replication and polyprotein translation occur (108). Upon cellular invasion, ZIKV exploits viral non-structural proteins NS4A and NS2A to commandeer ER membrane architecture, establishing specialized replication organelles including convoluted membrane structures (CM) and zippered ER formations (zER), that provide protected microenvironments for viral particle assembly and maturation (109, 110). The NS4A protein specifically binds host reticulon 3.1A (RTN3.1A) to orchestrate ER membrane remodeling, while NS2A facilitates viral ribonucleoprotein complex formation and recruits structural proteins to assembly sites through homotypic protein-protein interactions (109, 111) (Figure 5A). This extensive membrane reorganization exceeds the ER’s homeostatic protein folding capacity, precipitating ER stress conditions that activate the unfolded protein response (UPR) pathway (108, 109). The UPR activates C/EBP Homologous Protein (CHOP), leading to cell apoptosis. ZIKV achieves this by activating one of 3 UPR pathways: PERK, IRE1, and ATF6. Through PERK, ZIKV facilitates the phosphorylation of eIF2α to suppress protein synthesis; however, mRNAs like ATF4 become translated, activating CHOP (109–111) (Figure 5B). The IRE1 pathway splices XBP1 mRNA into its active form, XBP1s, and activates death proteins JNK and p38 MAPK, further promoting CHOP expression in the nucleus (106, 111, 112). ZIKV also triggers the cleavage of ATF6 to its active form (ATF6f) in the Golgi apparatus, allowing fragments to enter the nucleus and upregulate GRP78, PDI, and calnexin. These proteins reduce ER stress and, if unsuccessful, ultimately activate CHOP (108). After upregulation, CHOP functions as an apoptotic switch by suppressing the anti-apoptotic protein BCL-2. In fetal tissue, CHOP expression can result in apoptosis to neural progenitor cells, further contributing to fetal brain disruption sequence, when infection occurs in the first or second trimester (110). ZIKV persistence in trophoblast cells increases vertical transmission to the fetus (106, 107).
ZIKV-mediated teratogenesis through endoplasmic reticulum stress induction. (A) ZIKV enters the host cell via receptor-mediated endocytosis, using proteins NS4A and NS2A to remodel the ER membrane and create replication organelles for virion assembly. The reorganization of the ER membrane disrupts normal ER protein-folding capacity, triggering ER stress and activation of UPR. (B) Within the UPR, the PERK, IRE1, and ATF6 branches activate stress effectors, including increased translation of ATF4, splicing of XBP1 with subsequent activation of death proteins JNK and p38 MAPK, and expression of ATF6f. This culminates in CHOP upregulation, suppression of BCL-2, and apoptosis of placental and neural progenitor cells.
Systemic innate and adaptive host immune factors to arboviral infection
5
Acquired immunity to DENV and antibody-dependent enhancement
5.1
The innate immune response to DENV infection involves dendritic cells (DCs), macrophages, and monocytes as the initial responders to viral pathogens. The innate immune response to DENV is typical for any foreign pathogen: Pathogen recognition receptors (PRRs) on innate immune cells recognize pathogen-associated molecular patterns (PAMPs), and toll-like receptor (TLR) signaling triggers the production of inflammatory cytokines and IFNs (113). During pregnancy, the innate immune response to DENV changes. In-vitro studies in HTR8 cells, a human extravillous trophoblast cell line, found that DENV infection documented increased production of IL-6, IL-8, TNF-α, and CCL2 compared to non-infected cells (114). This increase in inflammatory cytokines is consistent with the characteristic cytokine storm of DENV.
Acquired immunity to DENV is mediated through the DENV neutralizing antibody responses to multiple serotypes in which infection with one results in long term protection but not against other serotypes. Neutralizing antibodies are commonly detected in individuals from countries with endemic dengue. These maternally derived neutralizing anti-DENV IgG antibodies efficiently cross the placental barrier may confer dengue immunity to newborns during the first few months of life (84, 115). Antibody titers for all four different DENV serotypes are known to be high initially after birth but steadily decay until they are absent after about one year (116). However, these antibodies can trigger an inflammatory response to DENV infection, where pathogens or infected cells bound by IgG form immune complexes and help neutralize DENV (117). One of the primary causes of fetal and neonatal death from DENV infection during pregnancy is insufficient maternally-derived IgG neutralizing antibodies (84). Most cases of DHF in infants occur when there are sub-optimal levels of maternally-derived neutralizing antibodies (117, 118).
However, non-neutralizing IgG can also be acquired after recurrent infections in mothers and in primary infection in infants (117). Unlike neutralizing antibodies, these are reactive and can lead to ADE. During ADE, non-neutralizing antibodies enhance pathogenesis through Fc gamma receptor (FcγR) mediated entry of the virus into leukocytes (119). These surface receptors recognize the Fc domain on antibodies (119), which triggers effector cells to respond during viral infection once activated. The normal pathway triggered by FcγR’s starts when the IgG immune complex binds, which triggers the phosphorylation of immunoreceptor tyrosine activating motifs (ITAM), further activating kinases and the protein kinase C pathway, opening Ca^2+^ channels and increasing the intracellular concentration of Ca^2+^. Kinase activation leads to actin remodeling, allowing for phagocytosis and cellular intake of the bound IgG immune complex and activating transcription factors that drive the expression and release of pro-inflammatory cytokines, leading to an antiviral immune response. ADE occurs through the exploitation of FcγR mediated uptake by viral complexes bound by non-neutralizing IgG antibodies, which allows for viral entry and DENV replication in immune cells (119). In neonates born to DENV infected mothers, ADE can occur during a specific window of opportunity. This is when maternally-derived IgG antibodies have decayed to a level where the infant has sub neutralizing levels of antibodies that are still capable of enhancing DENV infection in cells expressing Fc receptors (118).
Antibody-mediated ZIKV control and enhancement mechanisms – a dichotomous relationship
5.2
Maternal antibodies serve dual roles in ZIKV pathogenesis, with neutralizing antibodies generated from prior orthoflavivirus infections, particularly DENV, capable of either protective neutralization or pathogenic enhancement through ADE (120). As previously discussed, in ADE, non-neutralizing or sub-neutralizing antibodies facilitate viral cellular entry rather than clearance, increasing infectious severity and raising significant concerns regarding vaccine development strategies that might inadvertently elevate ZIKV vertical transmission risk through cross-reactive humoral responses (121).
Maternal antibodies against ZIKV survive in fetal circulation after crossing the placenta. According to Megli and Coyne (122), some of these antibodies can contribute to immune complexes, exacerbating placental inflammation and injury to the fetus, while others offer passive immunity. To promote transmission, ZIKV has been shown to take advantage of the autophagy pathway by producing exosomes that contain viral proteins and RNA. A different method of viral transmission to the fetus is possible because of the ability of these exosomes to cross the placenta (123). Blocking autophagy has been shown to lower ZIKV transmission, suggesting this mechanism may be important for viral dissemination in pregnancy (124). Apart from inhibiting the interferon response, ZIKV utilizes supplementary immune evasion strategies to improve its survival. Mechanisms include downregulation of antiviral pathways and upregulation of immunological tolerance by viral modulation of placental immunity gene expression (68). This modification may make it possible for ZIKV to survive in the placenta without inducing a strong maternal immune response (125).
Host innate and adaptive immunity towards ZIKV infection
5.3
ZIKV reduces the antiviral effects of type I IFN signaling by suppressing the maternal interferon response through the degradation of STAT2 (95). While maternal antibodies can help control the spread of ZIKV, the potential for ADE can worsen the infection (126) (Table 1).
Recent studies have identified IL-27 as an important host innate immune factor against viral infection in the placenta. Trophoblast organoids express both IL-27 and IL27 receptor constitutively, which can restrict ZIKV infection in the placenta (131). Additionally, decidual natural killer cells act as the front line of defense at the MFI and have shown to kill ZIKV-infected trophoblasts (132). However, the virus also has the capability of escaping detection by NK cells by upregulating MHC class 1 proteins, inhibiting NK cell receptors, demonstrating the ability of the virus to evade innate immunity (133). As previously discussed, ZIKV exacerbates placental damage by inducing a strong inflammatory response at the MFI. High concentrations of pro-inflammatory cytokines, such as IL-6, IL-1β, and TNF-α, are produced by infected trophoblasts and immune cells in the placenta (134). The body’s physiologic antiviral response includes these protective inflammatory mediators. Studies on STAT2-deficient mice, showed heightened vulnerability to ZIKV infection and provide additional evidence of the significance of STAT2 in ZIKV defense (29). Type III interferon (IFN-λ1), which offers paracrine and autocrine protection against viral propagation, has been demonstrated in term placentae to exhibit resistance to ZIKV infection constitutively (15). Duramycin administration, which reduces ZIKV infection rates in both primary placental cells and tissue explant cultures, may be a potential therapeutic approach for limiting viral placental invasion and fetal transmission (16).
The placenta contains specific micro RNAs (miRNAs) that act as genetic silencers by binding the 3’ untranslated region (UTR) of mRNA (135). A selection of these placenta-specific miRNA come from the chromosome 19 miRNA cluster (C19MC). It was recently discovered that this gene cluster protects trophoblast cells by hindering the excessive activation of innate immunity. So, C19MC may not only be important in maintaining immunity at the MFI, but also in protecting the placenta against potentially harmful autoimmunity (136). During ZIKV infection, C19MC attenuates the pathogenesis of the virus limiting its ability to infect and reproduce in the placenta. However, the exact mechanism by which this gene cluster acts is still unknown, as it doesn’t influence the IFN-III pathway or synergize with IFN-λ1 (137).
Concurrent pathogen infection during gestation
6
Coinfection represents the simultaneous establishment of multiple infectious agents within a single host organism. This phenomenon presents diagnostic and therapeutic challenges due to overlapping clinical manifestations and potential inter-pathogen interactions that may facilitate enhanced viral replication kinetics (138). This discussion examines gestational complications arising from concurrent infection with DENV or ZIKV alongside other viral pathogens, as well as dual DENV-ZIKV infection. DENV and ZIKV constitute phylogenetically related arboviruses that demonstrate geographic co-circulation in endemic regions (139). Although documented cases of dual infection exist, these pathogens exhibit competitive viral interference, whereby one agent predominates and suppresses replication of the competing virus (138). In areas with established co-circulation of both orthoflaviviruses, concurrent DENV-ZIKV infection in pregnancy is a significant clinical concern.
These viruses share antigenic cross-reactivity and seasonal transmission cycles, creating complex diagnostic challenges. Clinical presentations vary, with some cases demonstrating predominant ZIKV symptomatology (139) while others exhibit more pronounced DENV manifestations (140). DENV-ZIKV dual infections typically lack demonstrable viral synergism (140). The gestational impact of coinfection remains inadequately characterized. Recent data suggests pre-existing dengue immunity can modulate ZIKV pathogenesis during pregnancy. A murine study by Rathore et al. (126) demonstrated that pre-existing DENV antibodies may enhance ZIKV replication through ADE, resulting in elevated viral loads within placental and fetal tissue. Consequent outcomes included increased fetal mortality, enhanced inflammatory responses, and exacerbated placental pathology. These findings corroborate clinical observations suggesting that dual viral exposure may correlate with more severe gestational complications.
In humans, differentiation between the clinical manifestations of co- versus mono-infection with DENV and ZIKV in pregnancy may not be possible (141, 142). Congenital Zika syndrome (CZS) is known to result from isolated ZIKV infection, although co-infection may also be associated with microcephaly, intrauterine growth restriction (IUGR), premature birth, neurological, and neurodevelopmental aftereffects. Because of their structural similarities, especially in envelope proteins, ZIKV and DENV may cause cross-reactivity in serological tests complicating diagnostic testing in pregnancy. During the acute stage of illness, RT-PCR is still the gold standard for detecting viral RNA; however, once viremia goes away, the usefulness of this test decreases.
A study by João et al. (143) examined outcomes in pregnant women co-infected with HIV and ZIKV. The investigators noted that co-infection increased fetal risks. Additional studies have been conducted on coinfection between these arboviruses, including one on a cohort of pregnant women in the south of Mexico (142) and a case report from Colombia (142). These studies did not find that infection with multiple viruses enhanced or synergized clinical outcomes during coinfection. In a recent study Nolan et al. (144) investigated seroprevalence rates of co-infection among laboring mothers in El Salvador and documented elevated antibody titers against DENV and ZIKV. The need for more sophisticated and virus-specific diagnostic algorithms is suggested by the difficulty in attributing infant outcomes to a single pathogen due to the overlap in maternal immune responses (145) Strong maternal screening, molecular diagnostics, and integrated arbovirus surveillance are therefore urgently needed especially in areas endemic to both viruses.
Future directions
7
The comparative placental transmission mechanisms of DENV and ZIKV require future multidisciplinary research that connects immunology, virology, and maternal-fetal medicine. ZIKV can directly infect cytotrophoblasts and Hofbauer cells, while DENV infection relies on immune-mediated disruption of placental integrity (64). Most severe cases of dengue may occur because of ADE during secondary infection (146). However, a recent study that examined 619 DENV-infected infants in India questioned this dogma. In the study, primary and secondary DENV infection was determined by finding the ratio of IgM to IgG antibodies using ELISA. During secondary infection, severe cases were associated with the presence of maternal IgG resulting in ADE, while primary infections were associated with higher concentrations of IgM antibodies. Of the 619 total cases, primary infection was noted in 344 infants and children. Of those 344, 202 were severe, and 112 of those severe cases were due to primary infection. There were also 7 fatalities, and 5 of those 7 involved primary infection. The cohort of individuals analyzed were between newborn to 16 years of age, and 60% of children younger than 1 year old. All were positive for IgM, indicating primary infection. The results from this study show that primary infections can cause severe disease in pediatric subjects (146). Such studies open new avenues for improved DENV management in infants and children. However, it is important to note the lack of direct studies on the MFI during DENV infection as this field lacks both in-vitro models and true clinical or in-vivo studies. Given the dangers that DENV and ZIKV infection present to fetuses and infants, this area of study is crucial for the health of the mother-infant dyad when exposed to arboviral infection during pregnancy (13, 147, 148). Studies highlighting recent outbreaks of arboviral infections demonstrate differences between infections. One study explored an outbreak of ZIKV, DENV, and CHKV in Brazil during 2014-2016. While this outbreak resulted in increased cases of microcephaly, this association was documented with ZIKV and CHKV, not DENV (149). Another examined an increase of microcephaly in Brazil in 2015. This ZIKV outbreak was correlated with more children being born with microcephaly, but previous DENV outbreaks did not have the same association, leading to the conclusion that microcephaly as a result of CZS is unique to ZIKV (150).
The use of placental organoids, single-cell transcriptomics, and in vivo imaging, are recommended to identify differential pathways in real time, as well as in situations when there is co-infection or previous exposure to orthoflavivirus. The role of maternal immunity, particularly ADE, where non-neutralizing antibodies, especially from previous DENV exposure, can exacerbate ZIKV infection and result in worse placental and fetal outcomes, requires further investigation (63, 64). Vaccine design is directly affected by the interaction between antibodies and antigens. Due diligence must be taken in the development of vaccines against both viruses to prevent cross-reactive antibodies from exacerbating ADE, especially in pregnant or childbearing women. Despite being in early-phase trials, the current live-attenuated and DNA-based ZIKV vaccines are not yet authorized for general maternal use because of safety concerns regarding fetal exposure and the potential of ADE. The use of DENV vaccinations, such Dengvaxia, has also been restricted in some areas because of ADE-related effects in DENV-naïve people (65). Future research must therefore focus on developing vaccinations (151, 152) that elicit strong, widespread, and non-enhancing neutralizing antibody responses while also incorporating placental safety assessments in preclinical research. To lessen the effects of both viruses on perinatal outcomes, a comprehensive public health response incorporating vaccinations, diagnostics, surveillance, and treatments specific to the pregnancy will be necessary as the climate changes and arbovirus range spreads.
Conclusion
8
Both DENV and ZIK demonstrate maternal-fetal transmission capability during gestation, potentially resulting in severe morbidity for both. Elucidating the mechanistic pathways of transplacental viral dissemination is fundamental to developing comprehensive maternal-fetal protective strategies. While these orthoflaviviruses share common immunopathological features and epidemiological characteristics necessitating coordinated diagnostic and prophylactic approaches, their distinct placental invasion mechanisms and pathogenic impacts require pathogen-specific interventions. Future research priorities should emphasize placental barrier integrity maintenance, maternal-fetal immune response modulation, and vaccine safety profiles during pregnancy. Only through such targeted investigative efforts can we effectively mitigate the long-term sequelae associated with congenital orthoflavivirus infections. The compiled evidence presented in this comprehensive review reinforces the critical importance of early detection and therapeutic intervention, which has been demonstrated as the most efficacious strategy for limiting severe maternal and pediatric outcomes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arora N Sadovsky Y Dermody TS Coyne CB . Microbial vertical transmission during human pregnancy. Cell Host Microbe. (2017) 21:561–7. doi: 10.1016/j.chom.2017.04.007, PMID: 28494237 PMC 6148370 · doi ↗ · pubmed ↗
- 2Moffett A Chazara O Colucci F . Maternal allo-recognition of the fetus. Fertil Steril. (2017) 107:1269–72. doi: 10.1016/j.fertnstert.2017.05.001, PMID: 28577615 · doi ↗ · pubmed ↗
- 3Mor G Cardenas I Abrahams V Guller S . Inflammation and pregnancy: The role of the immune system at the implantation site. Ann N Y Acad Sci. (2011) 1221:80–7. doi: 10.1111/j.1749-6632.2010.05938.x, PMID: 21401634 PMC 3078586 · doi ↗ · pubmed ↗
- 4Whitelaw PF Croy BA . CURRENT TOPIC granulated lymphocytes of pregnancy. Placenta (1996) 17:533–43. doi: 10.1016/S 0143-4004(96)80070-1, PMID: 8916201 · doi ↗ · pubmed ↗
- 5Manaster I Mandelboim O . The unique properties of uterine NK cells. Am J Reprod Immunol. (2010) 63:434–44. doi: 10.1111/j.1600-0897.2009.00794.x, PMID: 20055791 · doi ↗ · pubmed ↗
- 6Williams PJ Searle RF Robson SC Innes BA Bulmer JN . Decidual leucocyte populations in early to late gestation normal human pregnancy. J Reprod Immunol. (2009) 82:24–31. doi: 10.1016/j.jri.2009.08.001, PMID: 19732959 · doi ↗ · pubmed ↗
- 7Parker EL Silverstein RB Verma S Mysorekar IU . Viral-immune cell interactions at the maternal-fetal interface in human pregnancy. Front Immunol. (2020) 11:522047. doi: 10.3389/fimmu.2020.522047, PMID: 33117336 PMC 7576479 · doi ↗ · pubmed ↗
- 8Pereira L . Congenital viral infection: traversing the uterine-placental interface. Annu. Rev. Virol. (2025) 5:273–99:. doi: 10.1146/annurev-virology, PMID: 30048217 · doi ↗ · pubmed ↗