Genome-wide identification of the HSF gene family in Chinese chestnut and functional characterization of CmHSF4 under temperature stress
XiuRong Xu, ZiQi Wu, Xibing Jiang, Shiming Cheng

TL;DR
This paper identifies and characterizes HSF genes in Chinese chestnut, focusing on CmHSF4's role in heat stress response.
Contribution
The study provides a genome-wide analysis of HSF genes in Chinese chestnut and identifies CmHSF4 as a key gene in heat stress response.
Findings
Eighteen CmHSF genes were identified and classified into three subfamilies, with subfamily A being the largest.
CmHSF4 showed a 324-fold increase in expression under high-temperature stress and is localized in the nucleus.
Transgenic tobacco experiments showed that CmHSF4 is involved in oxidative damage under heat stress.
Abstract
Heat shock transcription factors (HSFs) are key regulatory factors involved in plant responses to abiotic stress. To explore the HSF gene family characteristics in the Chinese chestnut (Castanea mollissima) (Cm) and its role in abiotic stress responses, we conducted a systematic analysis at the whole-genome level. Using bioinformatic methods, 18 CmHSF genes in the Chinese chestnut genome were identified, and these were unevenly distributed across 12 chromosomes. Phylogenetic analysis classified them into three subfamilies, A, B, and C, with the largest number of members (12) in A. Protein-conserved motif and gene structure analyses indicated that the internal structures of each subfamily were conserved, whereas significant differences existed among subfamilies, suggesting functional differentiation. Synteny analysis revealed that segmental duplication was the main driving force for the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
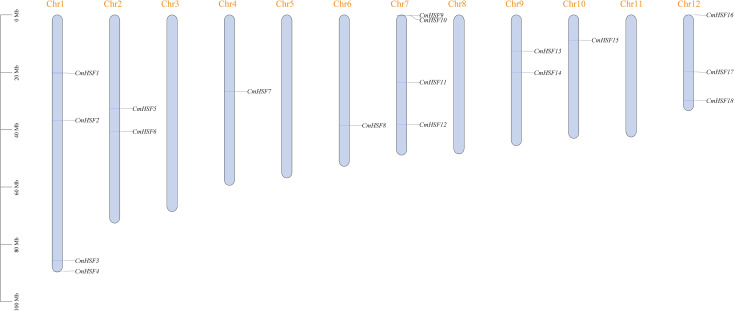 Figure 1
Figure 1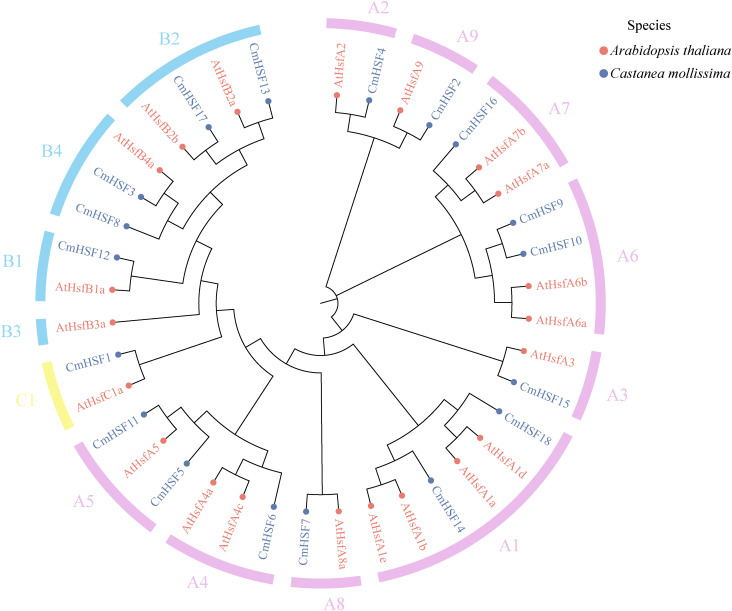 Figure 2
Figure 2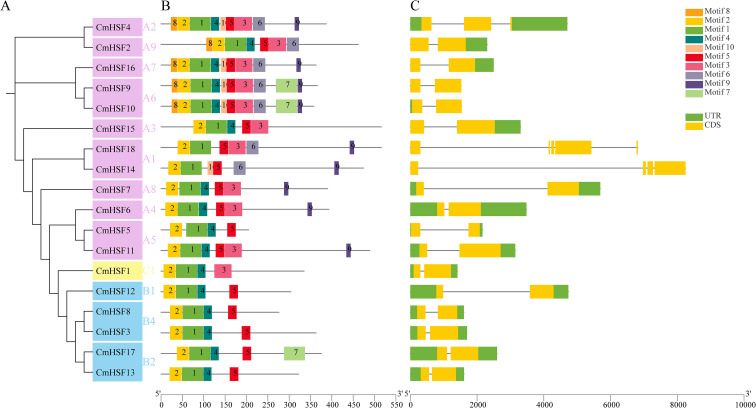 Figure 3
Figure 3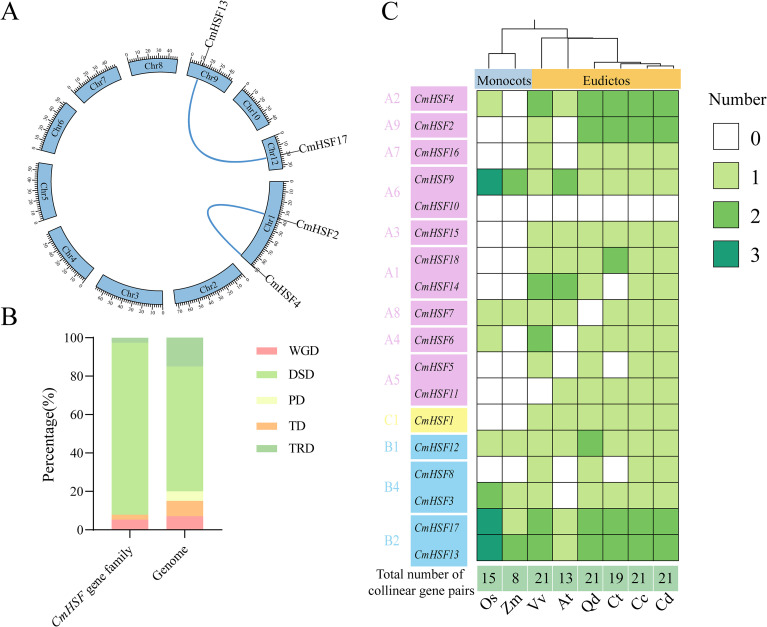 Figure 4
Figure 4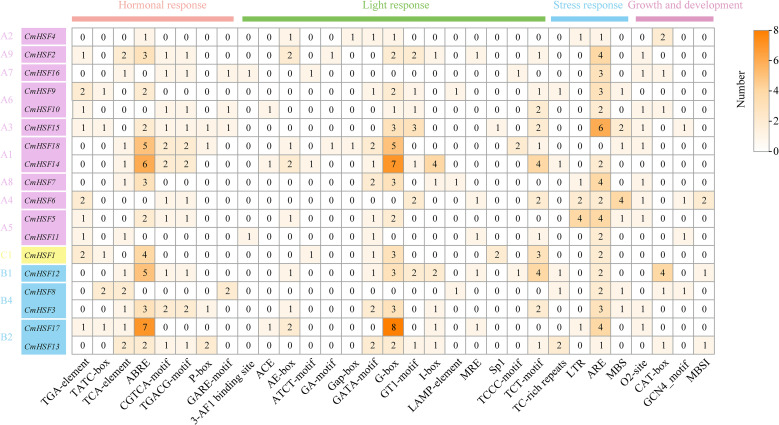 Figure 5
Figure 5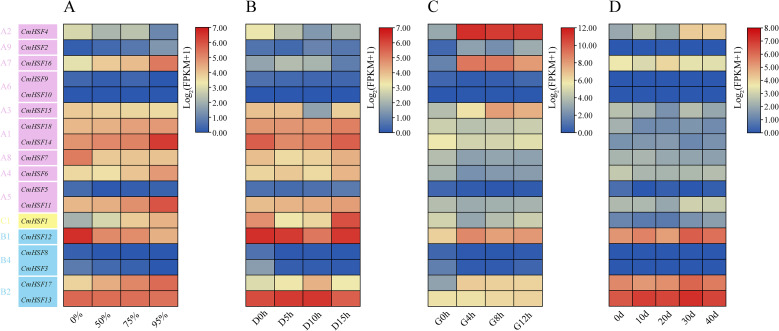 Figure 6
Figure 6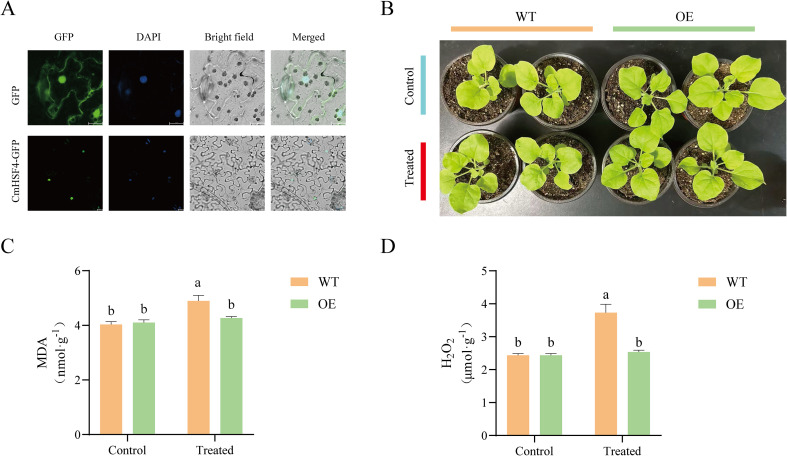 Figure 7
Figure 7| Gene ID | Chromosome ID | Gene name | Number of amino acid | Molecular weight | Theoretical pI | Instability index | Aliphatic index | Grand average of hydropathicity | Subcellular localization |
|---|---|---|---|---|---|---|---|---|---|
| EVM0015595 | CmHSF1 | Chr1 | 334 | 37271.16 | 5.59 | 55.30 | 68.29 | -0.455 | nucleus |
| EVM0029317 | CmHSF2 | Chr1 | 461 | 52774.3 | 4.63 | 66.15 | 80.69 | -0.644 | nucleus |
| EVM0031998 | CmHSF3 | Chr1 | 362 | 40750.02 | 8.08 | 47.90 | 69.20 | -0.561 | nucleus |
| EVM0015246 | CmHSF4 | Chr1 | 386 | 43240.12 | 4.80 | 51.93 | 75.26 | -0.562 | nucleus |
| EVM0025110 | CmHSF5 | Chr2 | 204 | 23648.91 | 7.60 | 54.47 | 69.36 | -0.68 | nucleus |
| EVM0007654 | CmHSF6 | Chr2 | 392 | 44801.05 | 5.08 | 55.47 | 71.33 | -0.696 | nucleus |
| EVM0007790 | CmHSF7 | Chr4 | 389 | 45139.93 | 4.71 | 51.30 | 69.90 | -0.748 | nucleus |
| EVM0018836 | CmHSF8 | Chr6 | 275 | 32032.17 | 6.95 | 43.27 | 70.51 | -0.645 | nucleus |
| EVM0028816 | CmHSF9 | Chr7 | 365 | 41898.06 | 5.39 | 56.39 | 69.40 | -0.768 | nucleus |
| EVM0010861 | CmHSF10 | Chr7 | 357 | 41531.42 | 5.32 | 58.72 | 65.21 | -0.899 | nucleus |
| EVM0018972 | CmHSF11 | Chr7 | 488 | 54821.75 | 5.53 | 64.29 | 58.18 | -0.849 | nucleus |
| EVM0022912 | CmHSF12 | Chr7 | 303 | 32867.23 | 5.54 | 31.56 | 61.75 | -0.78 | nucleus |
| EVM0003816 | CmHSF13 | Chr9 | 321 | 36026.15 | 5.74 | 53.43 | 69.88 | -0.725 | nucleus |
| EVM0013238 | CmHSF14 | Chr9 | 473 | 52210.06 | 4.81 | 46.04 | 75.48 | -0.525 | nucleus |
| EVM0008927 | CmHSF15 | Chr10 | 515 | 57846.48 | 4.73 | 66.16 | 69.81 | -0.639 | nucleus |
| EVM0019959 | CmHSF16 | Chr12 | 362 | 41218.54 | 5.56 | 58.85 | 63.26 | -0.794 | nucleus |
| EVM0007153 | CmHSF17 | Chr12 | 374 | 40517.68 | 4.73 | 51.32 | 69.79 | -0.645 | nucleus |
| EVM0015459 | CmHSF18 | Chr12 | 515 | 56971.41 | 5.02 | 62.70 | 64.16 | -0.701 | nucleus |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHeat shock proteins research · Plant biochemistry and biosynthesis · Plant and Fungal Interactions Research
Introduction
1
Chinese chestnut (Castanea mollissima Blume) is an important woody tree species in China. The fruit of the tree (Chinese chestnuts) are rich in nutrients and have significant economic and ecological value. However, Chinese chestnuts are often threatened by various abiotic stresses during their growth and development, such as high temperature, drought, and salinity. Of these stresses, extreme high-temperature weather caused by global warming is becoming increasingly frequent and severe, and it is one of the key environmental factors restricting the improvement of Chinese chestnut yield and quality. To breed superior stress-resistant Chinese chestnut varieties through the use of molecular breeding methods and ensuring the sustainable development of the industry, it is therefore necessary to determine stress-resistant genetic resources and identify the key stress-resistance genes.
Adverse environmental stresses such as cold, high temperature, drought, and salt can severely affect seed germination, growth, development, yield, and plant quality (Kowalczewski et al., 2020). Heat-shock transcription factors (HSFs) play a core regulatory role in the complex molecular networks of plant responses to high-temperature stress. When plants are exposed to heat stress, denatured proteins accumulate in the cytoplasm, activating HSFs, and causing them to form trimers and translocate to the nucleus. HSFs contain five special structures: the N-terminal DNA-binding domain, oligomerization domain, nuclear localization signal, C-terminal transcriptional activation domain, AHA, and nuclear export signal, which are also known as HSF domains. The first HSF gene in plants was cloned from tomato (Lycopersicon esculentum) (Scharf et al., 1990). In the nucleus, HSF specifically recognizes and binds to heat shock elements (HSE, with the conserved sequence 5’-AGAAnnTTCT-3’) in the promoters of downstream target genes through its highly conserved N-terminal DNA-binding domain (DBD) (Jiang et al., 2021). This initiates the transcription of numerous stress response genes, including heat shock proteins, which jointly assist in the repair of damaged proteins and maintaining intracellular homeostasis, thus establishing heat tolerance (Baniwal et al., 2004).
Typical plant HSF proteins have a modular structure, including a DBD, an HR-A/B region responsible for protein oligomerization (Oligomerization Domain, OD), a nuclear localization signal, a nuclear export signal, and a C-terminal transcriptional activation domain, where the short peptide motif specific to class A HSF is called the AHA motif (Guo et al., 2008). Based on the structural characteristics of the HR-A/B region, plant HSFs are classified into three major classes: A, B, and C. Of these, class A is generally regarded as the main positive regulator of the high-temperature response. The AHA motif is a specific transcriptional activation motif in HSFA, whereas HSFB and HSFC lack transcriptional activation activity because of the absence of the AHA motif (Scharf et al., 1990). The heat stress elements (HSEs) of HSFs are formed by the repetitive pattern of the palindromic binding motif upstream of the HS-induced gene TATA box, and they usually require two or more HSE motifs (Nover et al., 2001).
Many reports of functional studies on members of the HSF gene family have been published. For example, the HSF family in Camellia sinensis comprises 22 members. Of these, both CsHsf15 and CsHsf16 have high induction rates under light and low-temperature stress, and contain cis-acting elements related to light and low-temperature responses, which indicates that they may play a role in light resistance and low-temperature stress (Li et al., 2025). Seventeen members have been identified in the carnation (Dianthus caryophyllus), and each member is upregulated by high temperatures and drought. Among them, four members, DCAhSCF-A2A, DCAhSCF-A5, DCAhSCF-B2B, and DCAhSCF-C1 are slightly upregulated at low temperatures, which indicates that DcaHsf plays a key role in different stress response pathways (Li et al., 2019). Twenty-five HSF members have been identified in the Rosaceae fruit tree Malus domestica. At high temperatures, 12 were significantly higher than the reference sample, whereas only MdHsfA9b and Mdhsfb4A-b were strongly downregulated in response to an increase in temperature (Giorno et al., 2012). Therefore, members of the HSF gene family respond to various stressors through the activation or inhibition of plant growth. Many studies have confirmed that HSF proteins play important roles in plant responses to high-temperature stress. For example, the silencing of HSFA1a in tomatoes reduces the synthesis of chaperone proteins and HsfA1a proteins induced by heat stress, thereby increasing the sensitivity of HSFA1A-silenced tomato plants to heat stress. Furthermore, compared to wild-type plants, HSFA2 mutant plants of Arabidopsis thaliana are more sensitive to heat stress at 37°C. In addition to heat stress, HSFs are involved in plant growth and other abiotic stresses (high temperature, low temperature, salt, and drought). For example, HSFA9 is involved in the embryonic development and seed maturation of Arabidopsis thaliana and sunflower (Almoguera et al., 2002), and the four HSF genes (HSFA1e, HSFA3, HSFA4a, HSFB2a) of Arabidopsis thaliana are strongly induced by salt, low temperature and osmotic stress (Kilian et al., 2007).
With the extensive development of plant genome sequencing, the HSF gene family has been systematically identified at the whole-genome level in various plants, such as rice (Oryza sativa L.) (Baniwal et al., 2004), Arabidopsis (Guo et al., 2008), maize (Zea mays) (Lin et al., 2011), apple (Malus domestica) (Giorno et al., 2012), soybean (Glycine max), and cucumber (Cucumis sativus L.) (Chung et al., 2013). These studies have shown that the number of HSF family members varies significantly among different species and that they play diverse roles in response to high temperature and other abiotic stresses (such as low temperature, drought, and salt), presenting the coexistence of sub-functionalization and functional redundancy in the HSF family. These studies have provided an important foundation for understanding the roles and biological functions of the HSF gene family in the molecular mechanisms of high-temperature stress responses.
Based on this, this study employed bioinformatics methods to conduct the first genome-wide identification of the HSF gene family in Chinese chestnuts based on whole-genome data of Chinese chestnut. We systematically analyzed the physicochemical properties, evolutionary relationships, gene structures, and conserved motifs of the HSF gene family, and used transcriptome data to explore the expression patterns under various abiotic stresses. The results of this study provide a theoretical basis for understanding the biological functions of HSF genes in Chinese chestnuts and clarifying the molecular mechanisms of heat tolerance in Chinese chestnuts, with the aim offering guidance for obtaining valuable genetic resources to improve stress resistance in Chinese chestnuts and other forest crops.
Materials and methods
2
Identification, chromosome mapping and protein physicochemical property analysis of the Castanea mollissima HSF gene family
2.1
The preliminary data preparation involved downloading the genome data file and annotation file of the ‘N11-1’ chestnut variety from the Chinese National Genome Database (https://ngdc.cncb.ac.cn/gwh), the HSF domain file with the identifier PF00447 from the Pfam database (http://pfam-legacy.xfam.org/), and all 21 AtHSF protein sequences from the TAIR database (https://www.arabidopsis.org/) (Wang et al., 2021). Subsequently, the HSF domain and all AtHSF sequences were compared using the Simple HMM Search and Blast Several Sequences in the large database program of the TBtools software (Su et al., 2023), and the intersection of the comparison results of the two programs was uploaded to the InterProScan database (https://www.ebi.ac.uk/interpro/result/interprosca/). Finally, all members of the CmHSF gene family were identified (Chen et al., 2023).
After identifying all members, chromosome mapping and drawing of chromosome mapping diagrams for each CmHSF gene family member were conducted using the Gene Location Visualization (Advanced) program in TBtools software. They were named according to their chromosomal distributions.
To better understand the physicochemical properties of these CmHSF members, the amino acid quantity, molecular weight, isoelectric point, instability coefficient, and other indicators of CmHSF gene family member proteins were analyzed using the ExPASy ProtParam online tool (https://web.expasy.org/protparam/). The specific location of each family member protein in the cell, where it plays a specific role, was predicted using the BUSCA online tool (https://busca.biocomp.unibo.it/).
Construction of the evolutionary tree of CmHSF gene family
2.2
To better understand the grouping of the family members, we constructed phylogenetic trees for AtHSF and CmHSF. We first aligned all protein sequences using the MAFFT v7.471 program and employed MEGA7 software to construct a phylogenetic tree, which is displayed on the Chiplot website (https://www.chiplot.online/) (Xie et al., 2023). CmHSF was grouped based on the grouping of 21 AtHSF protein sequences in the TAIR database (https://www.arabidopsis.org/) and the phylogenetic tree.
Motif analysis and gene structure analysis of CmHSF gene family
2.3
The Simple MEME Wrapper program of TBtools software was used to analyze the motifs in the CmHSF protein sequences. Ten conserved motif sequences were obtained and uploaded to the InterProScan database (https://www.ebi.ac.uk/interpro/result/interprosca/) for functional annotation. Finally, the Gene Structure View of the TBtools software was used to visualize the motifs and gene structures (Chen et al., 2023; Xu et al., 2026).
Analysis of cis-regulatory elements in CmHSF gene family
2.4
Using the Fasta Extract program of the TBtools software, the 2000 bp promoter sequences preceding the ATG start codon of all CmHSF genes were extracted and uploaded to the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/) for cis-regulatory element prediction. The number of such elements was counted, and the final results were presented as a heatmap using the HeatMap program in TBtools (Chen et al., 2023; Wang et al., 2012; Qiao et al., 2019).
Genomic collinearity analysis of CmHSF gene family
2.5
The collinearity of CmHSF gene family members within the Chinese chestnut genome was analyzed using MCScanX software, and a collinearity map was created using the Advanced Circos program in TBtools. The non-synonymous substitution rate/synonymous substitution rate (Ka/Ks) of the collinear gene pairs was calculated using the Simple Ka/Ks Calculator (NG) in TBtools. The replication types of the CmHSF gene family were analyzed using the DupGenfinder program and classified into four categories: whole-genome duplication (WGD), transposed duplication (TRD), dispersed duplication (DSD), and tandem duplication (TD) (Chen et al., 2023). The proximal duplication (PD) pattern was not observed in the CmHSF gene family members.
Genome-wide co-linearity analysis of CmHSF gene family
2.6
The genome and gff annotation files of Chinese chestnut, grape, and corn were downloaded from the National Genome Database of China (https://ngdc.cncb.ac.cn/gwh), and the genome and gff annotation files of Arabidopsis thaliana and rice were downloaded from the Ensembl Plants database (https://plants.ensembl.org/). The genome and annotation files of Castanopsis tibetana were downloaded from the CNGB database (https://db.cngb.org/); the genome and gff annotation files of Japanese chestnut were downloaded from the PlantGenomePortal database (https://plantgarden.jp/en/index); and the genome and gff annotation files of American chestnut were downloaded from the Phytozome13 database (https://phytozome-next.jgi.doe.gov/info/cdentata_v1_1.). MCScanX software was used to conduct a collinearity analysis between the Chinese chestnut genome and those of these species, and to determine the number of collinearity gene pairs. The formation time of each species was analyzed using the Timetree database (https://timetree.org/) to infer the time point at which the members of the CmHSF gene family were generated.
Expression patterns of CmHSF gene family members under abiotic stresses
2.7
The transcriptome FASTQ file with the accession number GSA: CRA022911 was obtained from the Chinese National Genome Database (https://ngdc.cncb.ac.cn/gsa), and the transcriptome was analyzed to explore transcriptome data of Yanbao under low light stress (0%, 50%, 75%, and 90%, which represents the gene expression status of two-year-old Yanbao chestnut seedlings after 10 d under shading intensities of 0%, 50%, 75%, and 90%, respectively).
The transcriptome FASTQ file with the accession number PRJNA1166987 was obtained from the NCBI database (https://www.ncbi.nlm.nih.gov/). The transcriptome was analyzed to explore the gene expression status of Yanshan Zao Feng chestnuts under low-temperature (D0h, D5h, D10h, and D15h) and high-temperature (G0h, G4h, G8h, and G12h) stress. D0h, D5h, D10h, and D15h represent the transcriptome data of Chinese chestnut seedlings at -15°C after 0, 5, 10, and 15 h, and G0h, G4h, G8h, and G12h represent the transcriptome data of Chinese chestnut seedlings at 45°C after 0, 4, 8, and 12 h.
The transcriptome FASTQ file (accession number: PRJNA731244) was obtained from the NCBI for Biotechnology Information database (https://www.ncbi.nlm.nih.gov/). The transcriptome was analyzed to explore the gene expression status of Yanshan Zao Feng chestnuts under drought (0d, 10d, 20d, 30d, 40d) stress, where 0d, 10d, 20d, 30d, and 40d represent the transcriptome data of Chinese chestnut seedlings under continuous water-free conditions for 0, 10, 20, 30, and 40 days.
All transcriptome data were sequenced using the Illumina platform in the paired-end mode. After obtaining the reads, they were aligned to the reference genome of Chinese chestnut N11-1, and the fragments per kilobase of transcript per million fragments mapped (FPKM) was used as the gene expression level.
Verification of relative expression level of CmHSF4 under high-temperature stress using RT-qPCR
2.8
Leaves of one-year-old Chinese chestnut seedlings grown in a 45°C high-temperature stress environment for 0 h (G0h), 4 h (G4h), 8 h (G8h), and 12 h (G12h) were rapidly collected, frozen with liquid nitrogen, and stored at -80°C for subsequent gene cloning and qRT-PCR experiments. RNA was extracted from the tender leaves using the Plant RNA Extraction Kit (Takara, Beijing, China), and RNA purity was determined using a NanoPhotometer^®^ spectrophotometer (IMPLEN, CA, USA). RNA was reverse-transcribed into cDNA using the PrimeScriptTM RT Reagent Kit with gDNA Eraser (Takara, Beijing, China). RT-qPCR was performed on 12 samples (G0h, G4h, G8h, and G12h, four periods, with three replicates for each treatment, totaling 12 samples) using a SYBR PrimeScript RT-PCR Kit (Takara, Beijing, China). An ABI 7500 Real-Time PCR system (ABI 7500; Thermo Fisher Scientific, Singapore) was employed. The relative expression levels of the genes were calculated using the 2-ΔΔCT method. Fluorescent quantitative PCR primers were designed using the Batch qPCR Primer Design of the TBtools software. Primer information is provided in Supplementary Table 1, where CmActin is the internal reference gene for Chinese chestnuts. Bar graphs were plotted using GraphPad Prism (version 10).
Subcellular localization and tobacco transgenic verification
2.9
Using the seamless cloning technique, the ORF sequence of CmHSF4 with the stop codon removed was ligated into the pAN580 vector, and a GFP tag was attached to the N-terminus. These recombinant vectors were then introduced into Arabidopsis thaliana protoplasts using polyethylene glycol-mediated transformation. Under 470 nm excitation, the GFP fluorescence signal was observed using a laser confocal microscope to analyze the localization of CmHSF4 in the cells.
Using the seamless cloning method, the ORF sequence of CmHSF4 was ligated into the pBWA(V)HS vector containing the CaMV35S promoter, and the recombinant plasmid was transformed into the root-knot bacterium strain GV3101. Transgenic tobacco plants were obtained by Agrobacterium infection of the tobacco leaves. Wild-type (WT) tobacco and overexpression (OE) tobacco were incubated at 45°C for 5 min, and the differences between WT tobacco and OE tobacco under high and normal temperatures were observed and photographed. The determination of H_2_O_2_ and MDA contents using the kits provided by Wuhan ProNets Biotechnology Co., Ltd (Wuhan, China).At the same time, spectrophotometers were used to measure the MDA (malondialdehyde) and H_2_O_2_ (hydrogen peroxide) values of WT tobacco and OE tobacco under high and normal temperatures (Kaseb et al., 2021; Deng et al., 2025).
Data analyses were performed using Excel and SPSS v21.0(International Business Machines Corporation, Amonk, New York, USA). Statistical significance was determined by the difference levels using a paired Student’s t-test. The mean standard deviation (SD) from the mean of at least three biological replicates are presented.
Results
3
Chromosome localization and protein physicochemical properties analysis of the CmHSF gene family
3.1
After searching the Chinese chestnut genome data and comparing them using MAGA7.0, 18 CmHSF genes were identified, and these were named CmHSF1–CmHSF18, based on the positions of the genes on the chromosomes (Table 1). The physicochemical properties were predicted. The sequence analysis results showed that the protein sequences encoded by CmHSFs varied greatly, ranging from 204 (CmHSF5) to 515 (CmHSF18) amino acids, with corresponding molecular weights of 23.64–56.97 kD, and theoretical isoelectric points (pI) of 4.63 (CmHSF2) to 8.08 (CmHSF3). The instability index ranged from 31.56 (CmHSF12) to 66.16 (CmHSF15), with an average value of 54.18. Protein hydrophobicity analysis revealed that the average hydrophobicity ranged from -0.899 (CmHSF10) to -0.455 (CmHSF1), indicating that all CmHSFs were hydrophilic proteins, but there were certain differences in hydrophilicity. The most and least hydrophilic were CmHSF10 and CmHSF1, respectively, but most CmHSFs were both unstable and hydrophilic. Prediction of the subcellular localization of the proteins showed that all CmHSFs were located in the nucleus.
Chromosome localization analysis revealed that the 18 CmHSF genes were unevenly distributed on the 12 Chinese chestnut chromosomes (Figure 1). Among them, members on chromosomes 1 and 7 were the most abundant, with four members each, followed by chromosome 12, with three members each; chromosomes 2 and 9, with two members each; and chromosomes 4, 6, and 10, with only one CmHSF gene each. No CmHSF genes were detected on chromosomes 3, 5, 8, or 11.
The istribution of the CmHSF genes on the Chinese chestnut chromosomes. Vertical colored bars represent the chromosomes of Chinese chestnut. The gene name and number are shown at the right/left of each chromosome. The scale bar on the left represents the length of the chromosomes.
Phylogenetic analysis of CmHSF gene family proteins in Chinese chestnut and Arabidopsis
3.2
Phylogenetic analysis of the HSF family members in Chinese chestnut and Arabidopsis was conducted using MEGA software, and the results are shown in Figure 2. Based on the topological structure of the phylogenetic tree, the HSF gene families in Chinese chestnut and Arabidopsis were divided into three subfamilies (Class A, Class B, and Class C). Class C contained the fewest number of genes, with only one CmHSF1 gene; Class B contained five CmHSF genes, namely CmHSF3, CmHSF8, CmHSF12, CmHSF13, and CmHSF17; and Class A contained the largest number of members, with 12 CmHSF genes, accounting for 70.59% of the total. The distribution of the number of genes in Chinese chestnut was similar to that observed in Arabidopsis. In addition, multiple CmHSFs were clustered with Arabidopsis AtHSF genes in the same branch, including CmHSF13 in Class B with ATHsfB2a, CmHSF17 with ATHsfB2b, and CmHSF12 with ATHsfB1a. This indicated a high degree of relatedness and similar evolutionary trends, suggesting that they may have similar biological functions.
Phylogenetic tree based on the amino acid sequences of different species. Different branch colors in the figure indicate different groups; different colors of gene names indicate different orthogroups.
Prediction of CmHSF protein domain in chinese chestnut and analysis of gene structure
3.3
The Chinese chestnut CmHSF domain was predicted using TB tools. The results showed that the number of motifs in Chinese chestnut CmHSFs ranged from four to ten. Among these, the Class C subgroup had the fewest motifs (four). Except for CmHSF17, which contained five motifs, all other members of the Class C subgroup contained only four motifs. Notably, all the CmHSFs contained motifs 1 and 2, which indicated that motifs 1 and 2 are relatively conserved in CmHSFs and may play important roles. Additionally, motifs 6–10 only appeared in the Class A subgroup. The motif structures of CmHSFs in the same evolutionary branch were similar, suggesting that CmHSF proteins in the same branch have similar functions. The different numbers and distributions of motifs among the family members suggest that different CmHSFs may have different biological functions (Figures 3A, B). Analysis of the CmHSF gene structure using TBtools (Figure 3C) revealed that the number of introns in CmHSFs ranged from one to four, and the number of exons ranged from two to five. Among these, CmHSF14 and CmHSF18 in the Class A subgroup contained the most introns. The most closely related CmHSF genes contained the same number of introns and intron and exon lengths, and the gene structures were similar.
(A) The ten conserved motifs in CmHSFs. Conserved motifs of the CmDofs were identified using the online MEME program based on 25 full-length amino acid sequences with the following parameters: maximum number of motifs, 10; maximum width, 100. The lengths and positions of different motifs in the protein sequences are identified by the lengths and positions of the different color blocks. (B) Gene structure of CmHSFs. Exons, introns, and untranslated regions (UTRs) are indicated by yellow rectangles, black lines, and green rectangles, respectively.
Correlation analysis of CmHSF genes in Chinese chestnut
3.4
To elucidate the potential evolutionary mechanisms of the Chinese chestnut CmHSF gene family, we studied the replication of these genes. The results (Figure 4A) indicated that among the 18 CmHSF genes, two fragments were duplicated, located on three different chromosomes, and two homologous gene pairs were formed. These paired correlated genes may have similar functions. Based on the correlation results, the analysis of gene replication types revealed that, compared to the entire Chinese chestnut genome, the PD replication type was completely lost in the CmHSF gene family, the proportions of TD and TRD were decreased, and the proportion of the DSD replication type were significantly increased (Figure 4B). Further Ka/Ks calculations were conducted, and the results showed that the Ka/Ks values of the two gene pairs were all less than 0.3 (Supplementary Table S1). Further correlation analyses were performed between Chinese chestnuts and other species, and a heat map was drawn. A further analysis of the correlation between Chinese chestnut CmHSF genes and those of Arabidopsis thaliana, rice, grape, and corn (Figure 4C) showed that there were different numbers of correlated gene pairs between Chinese chestnut and the different species. The number of gene pairs between Chinese chestnuts and corn was the lowest, with only eight genes. The number of gene pairs between Chinese chestnuts and grapes was the highest at 21. The heatmap also showed that the majority of Class A members were only correlated with dicotyledonous plants, suggesting that these Chinese chestnut CmHSF members formed after the differentiation of monocot and dicotyledonous plants. The genes CmHSF12, CmHSF13, and CmHSF17 were correlated in all species, suggesting that they may play an important role in plant adaptation.
Analysis of collinearity and duplication types in the CmHSFgene family. (A) Intra-genomic collinearity of the CmHSFgenes. (B) Duplication types of the CmHSFgenes. TRD (Tandem Repeat Duplication), DSD (Dispersed Duplication), PD (Proximal Duplication), TD (Transposed Duplication) and WGD (Whole Genome Duplication) indicate transposon duplication, dispersed duplication, proximal duplication, tandem duplication and whole genome duplication in the figure. The same below (C) The number of genetic combinations of Chinese chestnut with other species.
Analysis of promoter anterior regulatory elements of CmHSF gene family in Chinese chestnut
3.5
The upstream promoter cis-regulatory elements, located at the beginning of the gene, serve as a reference for the biological function of the gene. This study analyzed the cis-regulatory elements of the 2,000 bp upstream promoter of CmHSF. As shown in Figure 5, the number of elements involved in biological and non-biological stress responses was the highest, including the cold response element (LTR) and anaerobic response element (ARE). Among them, ARE contained the highest number, which were present in almost all CmHSFs. The other elements were related to hormone responses, including (but not limited to) jasmonic acid methyl response elements (CGTCA-motif, TGACG motif) and salicylic acid response elements (TCA-element). The results of cis-regulatory element prediction further indicate that CmHSFs may have multiple biological functions and may be induced and regulated by hormones in response to non-biological stress.
Cis- elements in the promoter regions of CmHSFs. The colored block with a number represents the cis-element number of CmHSFs.
Expression pattern analysis of CmHSF genes in different stress conditions of Chinese chestnut plants
3.6
During the growth of a plant, it encounters various abiotic stresses that negatively affects its growth and development. To analyze the expression patterns of the CmHSF gene family under different abiotic stress conditions, a heat map was created based on the transcriptome data. The results showed that the expression levels of five genes (CmHSF3, CmHSF5, CmHSF8, CmHSF9, and CmHSF10) were nearly zero under different stress treatments. In contrast, the expression levels of the other 12 genes were relatively high under the different stress conditions, although their expression patterns differed. Under shading stress, the expression levels of six genes gradually decreased with an increase in the shading intensity, while the expression levels of seven genes gradually increased (Figure 6A). Under low-temperature stress, in addition to the above-mentioned five genes, the expression level of CmHSF2 was very low and was not considered. With an increase in the low-temperature treatment time, the expression levels of two genes gradually decreased, three genes gradually increased, three genes initially increased and then decreased, and four genes initially decreased and then increased (Figure 6B). As the intensity and duration of high-temperature stress increased, the expression levels of three genes gradually decreased, six genes gradually increased, one gene first increased and then decreased, and three genes first decreased and then increased (Figure 6C). Under drought stress, in addition to the above five genes, the expression of CmHSF2 was very low and was not considered (Figure 6D). With increasing drought duration, the expression levels of four genes gradually decreased, four genes gradually increased, two genes first increased and then decreased, and two genes first decreased and then increased. Notably, the expression of CmHSF4 increased by 324 times at 4 h of high temperature treatment. This gene exhibited the most drastic changes, and the qRT-PCR results confirmed the correctness of the transcriptome data (Supplementary Figure S1).
The expression pattern of CmHSF genes. (A) 0%, 50%, 75%, and 90% respectively represent different shading conditions. (B) D0h, D5h, D10h, D15h respectively represent the results after low-temperature treatment for 0 hours,5 hours, 10 hours, and 15 hours. (C) G0h,G4h, G8h, G12h respectively represent the results obtained after high-temperature treatment for 0 hours,4 hours, 8 hours, and 12 hours.The Roman numerals along the right-hand side of the figure indicate log2FPKM.
Subcellular localization of CmHSF4 and transgenic validation
3.7
To explore the subcellular localization of CmHSF4, we built a 35 s promoter CmHSF4–GFP fusion expression vector and converted Arabidopsis protoplasts. Confocal microscopy revealed that the GFP fluorescent signal exhibited a clear intracellular distribution pattern. More importantly, the GFP signal was completely colocalized with the staining region of the specific nuclear dye DAPI, confirming that CmHSF4 is a nuclear protein (Figure 7A). This study also analyzed the effects of 42°C high-temperature stress for 5 min on tobacco seedlings (Figure 7B), by measuring the MDA and hydrogen peroxide (H_2_O_2_) contents of tobacco leaves. The results showed that after high-temperature treatment, the leaves of tobacco seedlings exhibited extensive water loss and chlorosis, with the leaves drying up and curling, and the contents of MDA and H_2_O_2_ showed an upward trend (Figure 7C), indicating that the tobacco seedlings suffered significant high-temperature damage following high-temperature stress.
Functional characterization of the CmHSF gene. (A) Subcellular localization of CmHSF4. (B) The phenotype of tobacco after CmHSF4 overexpression. (C, D) Determination of MDA and H2O2 contents in tobacco leaves.
Discussion
4
In the present study, we successfully classified 18 CmHSF members into three classes: A, B, and C. The distribution pattern of their numbers (class-A > class-B > class-C) was consistent with that of model plants, such as Arabidopsis thaliana, rice, and corn. Previous studies have also found that each type of HSF family in various plants is highly similar, and that the distribution of members is relatively similar. All were classified into three classes, A, B, and C, of which class A was the most common. This indicates that the HSF gene family has reached a certain degree of conservation in the plant kingdom. It is speculated that with the evolution of plants, members of the class-B and class-C may have lost some amino acids, whereas the amino acids between the bilateral heptavalent configuration (HR-A/B) composed of hydrophobic amino acid residues in members of the A HSF family are relatively complete, suggesting that members of the class-A HSF family may be more conserved than those of the B and C classes.
Phylogenetic analysis showed that multiple CmHSF members (such as CmHSF12/13/17) were clustered in the same evolutionary branch as the directly homologous genes in Arabidopsis thaliana, strongly suggesting that they may have similar functions. This inference was supported by gene structure analysis, which showed that members with close genetic relationships had similar motif compositions and gene structures (such as the number of introns/exons), whereas there were significant differences among different subfamilies, especially motif 6-10, which are specific to the Class-A subfamily and may determine its unique transcriptional regulatory function.
The expansion of gene families and functional differentiation are important driving forces for species adaptation to the environment. Collinearity analysis indicated that fragment replication was the key mechanism driving expansion of the CmHSF gene family. The Ka/Ks values of the two pairs of replicated genes were much lower than 1, indicating that this family has been subjected to intense purification selection pressure during evolution and that its protein sequences and functions have been highly conserved, which is crucial for maintaining its core biological functions. Further cross-species collinearity analysis provided clues for the functional differentiation of different subfamilies. Most Class-A members only had collinearity with dicotyledonous plants, suggesting that they might have emerged after the differentiation of monocotyledonous and dicotyledonous plants, and functionally might have been more specialized in the adaptability of woody dicotyledonous plants. Genes such as CmHSF12, CmHSF13, and CmHSF17 were directly homologous in all analyzed species, including monocotyledonous plants, indicating that they are ancient and highly conserved members that may play an indispensable role in the core pathways of the plant kingdom in response to environmental stress.
Research on CmHSF upstream of the cis-elements of the 2000 bp sequence component revealed that the CmHSF promoter contains many light response elements, hormone response components, and adversity stress response elements. Considering that transcription factors can activate or inhibit transcription by binding to cis-elements in the promoters of related genes, it is speculated that some functional genes may regulate gene expression by binding to certain cis-elements in the CmHSF promoter, thereby participating in the regulation of adverse stress and affecting plant growth and development. However, related gene mining and specific regulatory methods require further investigation. In plants, HSF gene expression is mediated by abscisic acid (ABA) signaling. Upstream transcription factors can bind to ABRE to regulate the expression of HSF (Ohama et al., 2017), salicylic acid (SA), ethylene (Eth), and methyl jasmonate (MeJA), etc. stimulate the expression of the HSF gene in plants through hormone signaling pathways (Snyman) (Wang et al., 2022; Nie et al., 2022), thereby enhancing the tolerance of plants to high temperatures, indicating that plant hormones are widely involved in regulating the heat shock response (Clarke et al., 2009). Through our analysis of the HSF promoter region in Chinese chestnuts, numerous hormone response elements were identified, further indicating that plant hormones play a significant role in regulating the responses of plant HSF genes to heat shock. It is also known from the common cationic action elements of higher plant promoters, that the ABRE element is not only an abscisic acid signal response element, but also a high-temperature signal response element. In this study, ABRE elements were found in all 13 Chinese chestnut HSF genes, further indicating that Chinese chestnut HSF genes are widely involved in their responses to thermal shock.
This study also reports on the functions of some HSF transcription factors in the model plant Arabidopsis thaliana. By building a system for Eucommia ulmoides and members of the family of Arabidopsis, and according to the Chinese chestnut and Arabidopsis thaliana HSF clustering relationship, some gene functions can be predicted from the HSF evolutionary tree analysis; for example, AtHsfA2 and CmHSF4. In this respect, CmHSF4 has a relatively close genetic relationship and a similar evolutionary trend. Moreover, the over-expression of AtHsfA2 in Arabidopsis thaliana enhances its tolerance to high-temperature stress, suggesting that CmHSF4 may respond to high-temperature stress. An analysis of the expression patterns strongly verified the above prediction and revealed a diversity of functions among the family members. Under different abiotic stresses, CmHSF genes exhibit rich and complex expression profiles, indicating that although they belong to the same family, there may be a functional division of labor in stress responses. Notably, the expression level of CmHSF4 was sharply upregulated (324 times) after 4 h of high-temperature stress. This phenomenon made it a crucial candidate gene for the response to heat stress in Chinese chestnuts. To verify its function, we conducted subcellular localization experiments and confirmed that CmHSF4 was located in the nucleus, which is consistent with its expected function as a transcription factor. Subsequent functional verification experiments on transgenic tobacco demonstrated that high-temperature stress caused significant oxidative damage (increased MDA and H_2_O_2_ content) to tobacco seedlings; as such, further in-depth research should focus on the downstream target gene CmHSF4 regulation to enhance heat tolerance in Chinese chestnuts.
Our findings, in conjunction with recent studies, collectively delineate the HSF-mediated heat stress response mechanism as a multi-level, dynamic and coordinated complex network. Firstly, the core function of HSF lies in directly activating the transcription of downstream protective genes. For instance, MdHSFA2 in apples has been confirmed to directly bind to and activate the promoter of MdGolS4, thereby enhancing heat tolerance (Li et al., 2025). This study predicts that CmHSF4 may have a similar function, and its nuclear localization feature is consistent with the role model of a transcription factor. Secondly, there is a fine functional division of labor and collaboration within the HSF family. Traditionally less studied C-class HSFs, such as LlHSFC2 in lilies, have been proven to act as co-activators of transcription, significantly enhancing the transactivation ability of A-class HSFs (such as HSFA1, HSFA2, HSFA3) through heterologous interactions, and this interaction is specifically promoted after heat stress.
More importantly, the HSF pathway does not operate in isolation but extensively interacts with other signaling pathways, forming a larger regulatory network. Promoter analysis revealed that all CmHSF genes contain the abscisic acid (ABA) response element ABRE, which is not only a marker of ABA signaling but also a high-temperature response element, indicating that ABA signaling is deeply involved in the transcriptional regulation of Chinese chestnut HSF genes. This regulatory level may be very complex: on the one hand, transcription factors from other families can directly regulate HSF expression. For example, the HD-Zip I transcription factor LlHB16 in lilies can directly bind to and activate the promoters of LlHSFA2 and LlMBF1C, thereby positively regulating heat tolerance and linking the basic heat response pathway with ABA signaling. On the other hand, some transcription factors may also exert negative regulation on the HSF pathway. For instance, LlERF110, an AP2/ERF member in lilies, although induced by heat, its overexpression disrupts ROS homeostasis and inhibits the expression of heat protection genes (such as AtHSFA2), ultimately leading to reduced heat tolerance. Combining previous research, we speculate that CmHSF4 may directly regulate downstream heat protection genes (such as HSPs or GolS, etc.) through a mechanism similar to that of MdHSFA2; at the same time, it may also act as a network node, interacting with other members within the family (such as C-class HSFs) or transcription factors from other families to jointly integrate hormone signals such as ABA, precisely regulating the heat stress adaptability and growth and development balance of Chinese chestnuts. These findings lay a theoretical foundation for understanding the molecular mechanism of heat tolerance in woody plants and provide excellent candidate genes such as CmHSF4 for molecular breeding of Chinese chestnut stress resistance. Future work should verify the downstream target genes of CmHSF4 through EMSA and dual-luciferase reporter systems, and clarify its function at the plant level using transgenic technology, while further exploring its interaction with proteins such as CmHSFC to comprehensively map the transcriptional regulatory network of Chinese chestnut heat stress response.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Almoguera C. Rojas A. Díaz-Martín J. Prieto-Dapena P. Carranco R. Jordano J. (2002). A seed−specific heat−shock transcription factor involved in developmental regulation during embryogenesis in sunflower. J. Biol. Chem. 277, 43866−43872. doi: 10.1074/jbc.M 207330200, PMID: 12228226 · doi ↗ · pubmed ↗
- 2Baniwal S. K. Bharti K. Chan K. Y. Fauth M. Ganguli A. Kotak S. . (2004). Heat stress response in plants: a complex game with chaperones and more than twenty heat stress transcription factors. J. Biosciences 29, 471–487. doi: 10.1007/BF 02712120, PMID: 15625403 · doi ↗ · pubmed ↗
- 3Chen C. Wu Y. Li J. Wang X. Zeng Z. Xu J. . (2023). T Btools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 16, 1733–1742. doi: 10.1016/j.molp.2023.09.010, PMID: 37740491 · doi ↗ · pubmed ↗
- 4Chung E. Kim K. M. Lee J. H. (2013). Genome-wide analysis and molecular characterization of heat shock transcription factor family in Glycine max. J. Genet. Genomics 40, 127–135. doi: 10.1016/j.jgg.2012.12.002, PMID: 23522385 · doi ↗ · pubmed ↗
- 5Clarke S. M. Cristescu S. M. Miersch O. Harren F. J. M. Wasternack C. Mur L. A. J. (2009). Jasmonates act with salicylic acid to confer basal thermotolerance in. Arabidopsis thaliana New Phytologist 182, 175187. doi: 10.1111/j.1469-8137.2008.02735.x, PMID: 19140948 · doi ↗ · pubmed ↗
- 6Deng H. Y. He J. M. Huang R. Wei R. P. Wang R. H. Yan S. . (2025). Clonal variation of the basic resistance factors (POD, SOD, CAT, MDA, PRO) in Michelia chapensis. Biol. Plantarum 69, 88–97. doi: 10.32615/bp.2025.008 · doi ↗
- 7Giorno F. Guerriero G. Baric S. Mariani C. (2012). Heat shock transcriptional factors in Malus domestica: identification, classification and expression analysis. BMC Genomics 13, 639. doi: 10.1186/1471-2164-13-639, PMID: 23167251 PMC 3575323 · doi ↗ · pubmed ↗
- 8Guo J. Wu J. Ji Q. Wang C. Luo L. Yuan Y. . (2008). Genome-wide analysis of heat shock transcription factor families in rice and Arabidopsis. J. Genet. Genomics 35, 105–118. doi: 10.1016/S 1673-8527(08)60016-8, PMID: 18407058 · doi ↗ · pubmed ↗