Altitudinal and household breeding patterns of the medically important mosquitoes Aedes aegypti, Aedes albopictus and Culex quinquefasciatus in Nepal
Sunita Baral, Pramod Joshi, Bishnu P. Marasini, Ishan Gautam, Meghnath Dhimal, Ruth Muller

TL;DR
This study maps the breeding patterns of disease-carrying mosquitoes in Nepal, showing they thrive at various altitudes and in household containers.
Contribution
The study provides new insights into the altitudinal distribution and household breeding preferences of medically important mosquitoes in Nepal.
Findings
Aedes albopictus was the most abundant immature mosquito species found.
Cx. quinquefasciatus dominated adult mosquito collections.
Mosquitoes were found breeding across a wide altitude range from 140 to 2066 meters.
Abstract
Medically relevant mosquitoes have established populations across Nepal, adapting to diverse environmental conditions and altitudes. In addition to arbovirus vectors, the presence of Culex quinquefasciatus, a primary vector of lymphatic filariasis, underscores the need for comprehensive mosquito surveillance. This study investigated the presence, abundance, container preferences, and altitudinal distribution of Aedes aegypti, Aedes albopictus, and Cx. quinquefasciatus across selected districts in Nepal. We also examined the co-occurrence of Aedes species and their association with container type and location. An entomological survey was conducted along an altitudinal transect from 140 meters above sea level (m asl) (Chitwan) to 2340 m asl (Dolakha). Immatures were collected from 1,157 water-holding containers across 636 households using dipper and dropper techniques, while adults were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
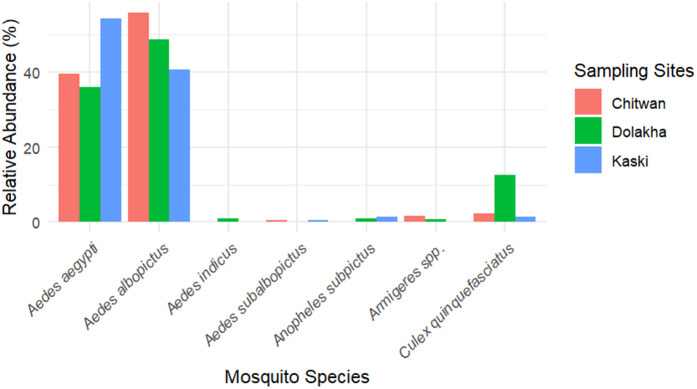 Fig 1
Fig 1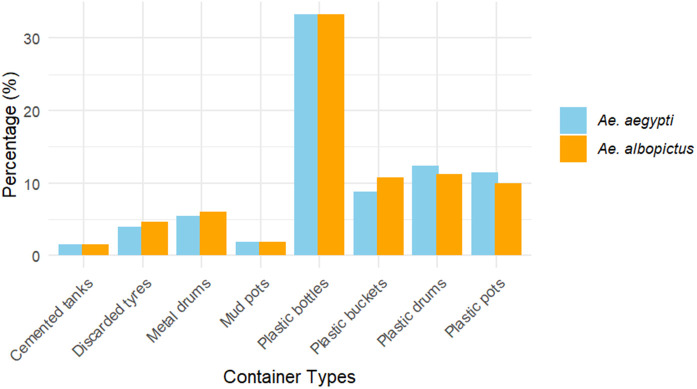 Fig 2
Fig 2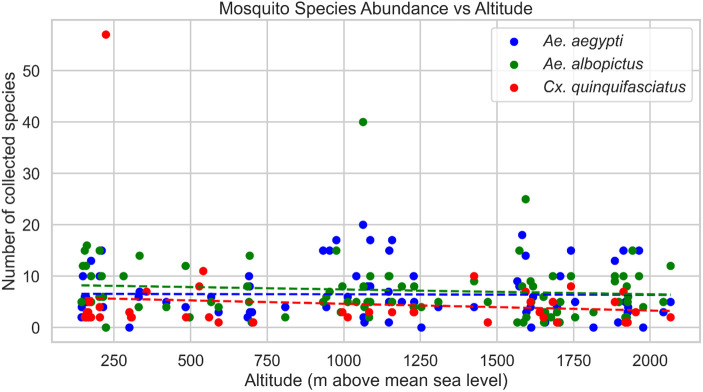 Fig 3
Fig 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMosquito-borne diseases and control · Malaria Research and Control · Parasitic Diseases Research and Treatment
Introduction
Vector-borne diseases accounted for more than 80% of global population risk, with mosquito-borne diseases (MBDs) being the major contributor [1]. MBDs are considered a major health threat in Southeast Asian countries, including Nepal. They account for more than 700 million infections with 1 million deaths worldwide [2]. Nepal is now endemic to various MBDs such as malaria, lymphatic filariasis, Japanese encephalitis, chikungunya and dengue posing a significant public health burden [3]. Though the first case of dengue in Nepal was reported in a Japanese volunteer in 2004 [4], the very first outbreak in Nepal and the detection of the primary dengue vector Ae. aegypti in low-altitude regions of Terai bordering India were confirmed in 2006 [5]. Since then, sporadic dengue cases and dengue outbreaks have occurred from tropical lowlands to subtropical hilly regions year by year, with the so far largest dengue outbreak affecting all districts previously considered as non-endemic in 2022 [6–8]. Chikungunya virus (CHIKV) was first reported in Nepal in 2013, and subsequent studies confirmed its presence in different areas, including the Terai regions during the 2014–2015 outbreak [9,10].
Aedes aegypti and Ae. albopictus are not only the primary and secondary vectors capable of transmitting the dengue virus [11], but are also the principal vector species capable of transmitting the CHIKV [12]. In the case of Nepal, Ae. albopictus was reported in the 1950s [13] and Ae. aegypti in 2006 [14]. Genomic data indicate a non-gradual expansion of Ae. aegypti in Central Nepal [15]. Now both vectors are recorded at altitudes up to 2438 m above sea level [16] and Ae. albopictus was reported up to 2520 m [17]. Between the 1950s and the detection of Ae. aegypti in Kathmandu in 2009, immature and adult stages of DENV and CHIKV vectors were recorded in the lowland and middle mountain regions [14,18,19]. However, the expansion of these vectors to the high mountainous regions up to 2438m asl [16,20], highlights the increasing adaptability of these vectors to higher altitudes and the growing risk of vector-borne diseases in previously unaffected areas. This is attributed to the fact that the warming in higher mountain regions is more prominent than in the lowland areas such as Terai and Siwalik [21–23].
Besides dengue vectors, other vectors have already established their distribution in Nepal. Lymphatic filariasis (LF), a mosquito-borne parasitic disease, is transmitted by the bite of different species of mosquitoes (Culex, Anopheles & Aedes) and is caused by thread like filarial worms; Wuchereria bancrofti and Brugia species. In Nepal, Culex quinquefasciatus is the principal vector of LF. Sixty-three out of 77 districts of Nepal are considered endemic to LF on the basis of Immunochromatography Test (ICT) card surveys, morbidity reporting, vector density, sanitation status of the districts, and geo-ecological comparability [24]. In the absence of a vaccine and effective treatment for several of these diseases, symptomatic treatment and vector control programs are at present the only possible approaches for tackling these infections.
The principal vector of LF, Cx. quinquefasciatus, has been recorded at elevations up to 2100 meters above mean sea level (m asl) in central Nepal [20]. Similarly, Cx. tritaeniorhynchus, the principal vector of Japanese encephalitis in Nepal, has been observed at elevations up to 2000 m asl in Eastern Nepal [25]. Additionally, malaria vectors such as Anopheles fluviatilis, Anopheles annularis, and the Anopheles maculatus complex have been recorded in eastern Nepal up to 1820 m asl, with their larvae found at elevations as high as 2310 m asl [25]. Similarly, Aedes japonicus, Anopheles farauti, and Culex sasai were reported for the first time from Nepal from various landscapes [26]. However, their function as vectors in the spread of human pathogens in the area is still unknown and needs more research.
Our study aimed to examine the ecological pattern of three medically important mosquito species at varying elevations: Ae. aegypti, Ae. albopictus, and Cx. quinquifasciatus. In particular, we addressed the following research questions: 1. What is the relative abundance of Ae. aegypti, Ae. albopictus, and Cx. quinquefasciatus across different altitudes in Nepal? 2. Which container types are most frequently used as breeding habitats for Aedes mosquitoes? 3. How does altitude influence the distribution of Ae. aegypti, Ae. albopictus, and Cx. quinquefasciatus? This study also aimed to record the presence of other mosquito vectors found during the collection process to gain a more comprehensive understanding of the overall vector ecology in the area.
Methods
Study design
The sampling was carried out during the post-monsoon season from 30^th^ October 2021–30^th^ November 2021. Three districts (Chitwan, Kaski, and Dolakha) were selected purposely based on the highest number of dengue cases determined by the reports published by EDCD (Epidemiology and Disease Control Division) Nepal in 2019 and also in order to reflect various altitudinal and ecological settings. Altitude and temperature were chosen as the main environmental factors affecting vector ecology in order to lessen the influence of covariates like direct road connectivity, wind corridors, and urbanization. Within three districts, 35% of the governmental health facilities were randomly selected by generating a random number generator in Microsoft Excel resulting in 15 health facilities from Chitwan, 18 health facilities from Kaski, and 20 health facilities from Dolakha districts, respectively. From within those health facilities, 636 households (12 per health facility) were selected conveniently along an altitudinal gradient ranging from lowland to highland, from 140 m asl (Chitwan) to 2339 m asl (Dolakha).
The elevation of collection sites in Chitwan district ranged from 140 m to 1066 m asl, with temperatures ranging from 23°C to 30°C with minimal rainfall. The population density is approximately 325 people per km². The collection sites in Kaski were located from 680 m to 1957 m asl and average temperature ranges from 10 °C to 20 °C. The collection month is among the driest months, with minimal to no rainfall. The population density is approximately 297 people per km². The elevation of sampling sites in Dolakha ranges from 977 m to 2339 m asl. The district’s annual average temperature is approximately 31.06 °C. The population density is about 79 people per km².
Entomological survey
In the study areas, all types of possible breeding habitats, like cemented tanks, plastic drums, metal drums, mud pots, discarded tyres etc., were searched in and around the houses for Aedes immatures. A total of 180 households from Chitwan, 240 from Dolakha, and 216 from Kaski were chosen. The immature collections were done using the dropper and dipper technique, whereas adults were collected using the BG-Sentinel traps equipped with a BG-Lure (Biogents AG, Regensburg, Germany) without carbon dioxide for 24 hours. BG-Sentinel traps were placed in the residential areas for safety purposes. The collected larval and adult mosquito samples were transported to the laboratory of the Natural History Museum, Tribhuvan University, Swayambhu, Kathmandu, Nepal. Larvae and pupae were reared to adulthood in plastic cups containing water from breeding habitats at room temperature covered with a net for adult emergence. Emerged adults were morphologically identified using the standard identification key by Darsie and Pradhan [13]. Permanent slides of larvae unable to emerge were made and identified using a stereomicroscope based on larval characteristics.
Data analysis
Data obtained was entered into the Microsoft Excel spreadsheet and analysed using SPSS version 23 and R version 4.3.3. Binary logistic regression analysis was used to assess the associations between the presence of Ae. aegypti, Ae. albopictus and co-occurrence with container type, location (indoor/outdoor) and district. However, due to sparse data and instability of model estimates across some containers categories, multivariate logistic regression analysis could not be performed. The statistical significance of associations was evaluated using p-values, with a threshold of p < 0.05 considered significant. In accordance with World Health Organization Southeast Asia Regional Office (WHO SEARO) guidelines [27], following entomological indices have been calculated: House Index (HI, percentage of houses positive for Aedes larvae), Container Index (CI, percentage of containers positive for Aedes larvae), and Breteau Index (BI, number of containers positive for Aedes larvae per 100 houses). The container preference of Aedes larval breeding was additionally assessed by calculation of the breeding preference ratio (BPR = % of positive containers/% of wet containers) [16]. The mean ratio between Ae. aegypti and Ae. albopictus was calculated as the percentage of Ae. aegypti positive containers divided by the percentage of Ae. albopictus positive containers for each container type. Similarly, the effect of elevation on mosquito abundance was evaluated using a generalized linear model (GLM) with a Poisson distribution.
Ethical statement
Ethical clearance for the entomological survey was obtained from the Ethical Review Board of Nepal Health Research Council under the Research Proposal ID: 206/2021. Since the study did not involve human participants, informed consent was not required. However, permission from household members was granted prior to sampling.
Results
Mosquito distribution and abundance across study sites
A total of 1,157 (Chitwan = 197, Kaski = 644 and Dolakha = 316) water-holding containers from 636 households were surveyed, yielding 1,323 immature mosquitoes (Chitwan = 311, Kaski = 467 Dolakha = 545). Additionally, 158 (Chitwan = 121, Kaski = 28, Dolakha = 9) adult mosquitoes were collected from both inside and around the houses. Both dengue vectors, Ae. aegypti and Ae. albopictus, as well as the lymphatic filariasis vector, Cx. quinquefasciatus, were recorded throughout the study transect. Other species showed more restricted distributions, with Armigeres spp. recorded up to 1,912 m asl, Aedes indicus up to 1,657 m asl, Pallidus triatus up to 1,555 m asl, Anopheles subpictus up to 1,081 m asl, and Aedes subalbopictus up to 591 m asl.
Among the immatures, 91% are known as dengue vector species (Ae. aegypti: 43.4%, Ae. albopictus: 47.6%). Other immature mosquitoes belong to Cx. quinquefasciatus (6.1%), Anopheles subpictus (0.8%), Armigeres spp. (0.7%), Ae. subalbopictus (0.4%) and Ae. indicus (0.4%). Immature Ae. aegypti, Ae. albopictus, and Cx. quinquefasciatus were recorded from all study areas (Fig 1), indicating their widespread presence and adaptability to varying environmental conditions. Notably, all three species were observed at altitudes as high as 2067 m, highlighting their ability to thrive across a broad altitudinal range, including high-altitude environments. In contrast, Ae. subalbopictus was recorded exclusively in Chitwan and Kaski, Armigeres spp. in Chitwan and Dolakha, Anopheles subpictus in Kaski and Dolakha and Ae. indicus in Dolakha. The relative abundance of immature mosquitoes varies between three districts. The relative abundance of immature Ae. albopictus was found to be highest in the lowlands (Chitwan: 55.9%) whereas immature Ae. aegypti were found to be highest in the hilly region (Kaski: 54.4%). Likewise, the relative abundance of Cx. quinquefasciatus was found to be highest in Dolakha (12.5%). House index (HI) for Ae. albopictus (16.67) and Ae. aegypti (15.42) were found to be highest in Dolakha whereas container index (CI) and Breteau index (BI) for both Ae. albopictus (CI: 9.14; BI: 10) and Ae. aegypti (CI: 9.64; BI: 10.56) were found to be highest in Chitwan (Table 1).
Table 1: Prevalence of Ae. aegypti and Ae. albopictus larvae across different districts in Nepal.
The relative abundance of mosquito immatures across Chitwan, Kaski, and Dolakha.
Among the adults, the majority of the species belong to the principal vector of lymphatic filariasis, Cx. quinquefasciatus (82.9%). The relative abundance of adult mosquitoes differs between districts. Adult Cx. quinquefasciatus was recorded across all study sites, while adult Ae. albopictus was found exclusively in Dolakha. In contrast, adult Ae. aegypti, Ae. indicus, Armigeres spp., Anopheles subpictus, and Pallidus triatus were detected only in Kaski. Moreover, the altitude did not have a statistically significant effect on the number of collected mosquitoes (GLM with Poisson distribution, p = 0.563).
Breeding habitats associated with the presence and co-occurrence of Ae. aegypti and Ae. albopictus
Overall, about 7.6% of wet containers were infested with Ae. albopictus, 7.5% with Ae. aegypti and 6.3% contained larvae of both species, Ae. aegypti and Ae. albopictus. The location of containers (574 indoor versus 583 outdoor containers) significantly influenced the presence of Ae. aegypti, Ae. albopictus, and their co-occurrence, with p-values of <0.001 for all comparisons. In indoor containers, solely Ae. aegypti larval infestation was found in 3% and Ae. albopictus in 2.6%, respectively, whereas larvae of both species co-occurred in 2.4% of the inspected indoor containers. In comparision to indoor container, the odds of Ae. aegypti infestation were 4.47 times higher in outdoor container (95% CI: 2.60–7.70, p < 0.001), Similarly, the odds of Ae. albopictus presence were 5.53 times higher outdoor (95% CI: 3.02–9.42, p < 0.001). Likewise, co-occurrence of both species was significantly greater outdoor (OR = 4.50, 95% CI: 2.49–8.16, p < 0.001).
Statistical analysis showed significant associations between all wet container types (P < 0.001) for both mosquito species and their co-occurrence. Using cemented tanks as the reference group, plastic containers showed markedly higher odds of Aedes occurrence. The most notable findings were observed in plastic bottles, where 33.3% tested positive for both Ae. aegypti and Ae. albopictus, with an odds ratio (OR) of 33.50 (95% CI: 2.47–452.76), indicating a strong association for both species. Similarly, plastic drums showed a high prevalence, with 12.4% testing positive for Ae. aegypti (OR = 9.47, 95% CI: 1.28–70.09) and 11.2% for Ae. albopictus (OR = 8.43, 95% CI: 1.13–62.54), both with significant associations. Plastic buckets also exhibited considerable positivity, with 8.8% positive for Ae. aegypti (OR = 6.43, 95% CI: 0.84–49.30) and 10.8% for Ae. albopictus (OR = 8.1, 95% CI: 1.07–61.66). Co-occurrence of both species was found in 7.7% of plastic bucket containers (OR = 5.61, 95% CI: 0.727–43.33). On the other hand, cemented tanks, discarded tyres, metal drums, and mud pots showed lower positivity, with no significant association with Ae. aegypti or Ae. albopictus (Fig 2). Rock pools, temporary ditches, and wooden water containers had no recorded positivity for any species. The overall findings suggest that plastic containers, particularly plastic bottles and plastic drums, are strongly associated with the presence of both mosquito species. The mean ratio of Ae. aegypti to Ae. albopictus breeding preference across all container types is 1.23. This indicates that, on average, Ae. aegypti has a slightly higher breeding preference than Ae. albopictus in the observed container types (Table 2).
Table 2: Distribution of Ae. aegypti and Ae. albopictus larvae in different container types.
Percentage of Ae. aegypti and Ae. albopictus positive containers.
Bivariate logistic regression analysis of the district-wise occurrence of Ae. aegypti and Ae. albopictus revealed significant differences across the study districts. In Chitwan, 197 wet containers were searched, with 4.6% testing positive for Ae. aegypti and 4.5% for Ae. albopictus. In Kaski, a total of 644 wet containers were examined, with 75.9% testing positive for Ae. aegypti (OR = 5.408, 95% CI: 1.943–15.049, p = 0.001) and 73.9% for Ae. albopictus (OR = 5.316, 95% CI: 1.909–14.80, p = 0.001), showing a statistically higher occurrence of both species compared to Chitwan. In Dolakha, 316 containers were searched, with 19.5% testing positive for Ae. aegypti (OR = 2.095, 95% CI: 0.695–6.320, p = 0.189) and 21.6% for Ae. albopictus (OR = 2.335, 95% CI: 0.789–7.027, p = 0.125), but neither species showed a statistically significant association. These findings highlight a stronger occurrence and association of both mosquito species in Kaski compared to Dolakha and Chitwan.
Discussion
During the study period, the dengue vectors, Ae. aegypti and Ae. albopictus, and the lymphatic filariasis vector, Cx. quinquefasciatus, were found throughout the study transect from 140 m asl up to 2066 m asl (Fig 3). Consistent with our findings, the same vectors were found up to 2100 m asl in high mountain regions, Dhunche and Rasuwa of Nepal, and immatures of Ae. aegypti up to 2130 m asl in Darjeeling [28] and in Puebla, Mexico [29]. The broad distribution of these vectors may be attributed to anthropogenic factors such as habitat modification, water storage practices and climate change as well as environmental factors including rainfall patterns, seasonality, and geographical adaptability [30]. Thus, understanding these dynamics is crucial to managing the risk of mosquito-borne diseases and forecasting mosquito behavior. Our study documented a slightly higher abundance of Ae. albopictus than Ae. aegypti, which is in accordance with the studies carried out in India [31,32], Malaysia [33], Pakistan [34] and Nepal [26]. By contrast, findings from the study carried out by Tuladhar et al, 2019 [35] and Kawada et al, 2020 [36] showed the higher abundance of Ae. aegypti. Plastic containers, particularly drums and buckets, have been identified as the most productive breeding habitats for Ae. aegypti and Ae. albopictus, consistent with previous studies that highlight plastic containers as favorable breeding sites for these mosquito species [37,38]. The preference of Ae. aegypti and Ae. albopictus for plastic containers may be due to their natural ability to easily warm up the contained water, which stimulates the hatching of mosquito eggs and encourages larval development. This is also because long-lasting containers that retain water for longer periods, particularly artificial ones, provide ideal breeding conditions [39,40]. However, other studies reported discarded tyres as the preferred breeding habitats for Ae. aegypti and Ae. albopictus [33,41–44]. The decreased productivity of tyres in our study probably indicates their limited availability, regular disruption, direct sunlight exposure or variation in rainfall retention across ecological settings.
The altitudinal distribution of immature and adult life stages of predominant and medically relevant mosquitoes.
In contrast to our findings, natural breeding containers did not harbor Aedes immatures, aligning with studies conducted in Nepal and Thailand, which reported a preference for artificial breeding containers over natural ones by Aedes immatures [20,44]. The shift towards a preference for artificial breeding containers by Aedes immatures could be attributed to the ongoing evolution of the anthropophilic behavior of Aedes mosquitoes, strongly associated with the density of the human population and environmental factors [45]. Several factors may contribute to this preference, including container type, microclimatic conditions, and species-specific behaviors [46]. Ae. aegypti, typically considered a domestic mosquito, has demonstrated adaptability to outdoor breeding habitats, particularly in artificial containers such as plastic buckets, drums, and discarded tires [47,48]. Aedes albopictus, on the other hand, is well-documented for its ability to exploit a wide range of outdoor breeding sites, including both artificial and natural containers, making it highly adaptable to diverse environmental conditions [16]. Breeding in artificial containers close to human dwellings provides convenient access to potential hosts for blood-feeding. Both vector mosquito species breed in containers and have a close association with humans, displaying a strong preference for human blood [49,50]. Although binary logistic regression revealed significant association between container types and mosquito presence, some odds ratio showed wide confidence interval indicating low precision of estimates. This likely reflects the small number of positive containers and limited sample size within certain container categories.
Despite their habitat preferences, in areas where both species coexist, their larvae are frequently discovered together in the same breeding sites [50]. The presence of Ae. aegypti, Ae. albopictus, and their co-occurrence was significantly higher in outdoor containers, consistent with previous studies conducted in Nepal, India, and Peru [20,33,51,52]. This trend suggests that outdoor environments provide optimal conditions for mosquito breeding, likely due to greater exposure to rainfall, increased availability of water-holding containers, and minimal human intervention in emptying or treating these sites. Co-occurrence of both species in outdoor containers may increase the risk of interspecific competition, which could influence larval survival and adult emergence. Moreover, under high larval densities, Ae. albopictus often outcompetes Ae. aegypti, leading to reduced survivorship and altered adult traits, particularly in outdoor plastic containers as observed in this study [52]. However, the persistence of both species in shared breeding sites suggests that resource partitioning or differential habitat selection may play a role in mitigating competitive exclusion [50]. The presence of both species in outdoor settings is particularly concerning for vector control efforts, as it suggests increased potential for arbovirus transmission in peri-domestic environments.
In countries endemic to dengue fever, entomological indices are crucial tools for guiding vector control programs [53]. Among the commonly used entomological indices, the most widely used indices in monitoring dengue vector populations are HI and BI [54]. The Pan American Health Organization (PAHO) has divided the risk factors for dengue transmission into three levels: low (HI < 0.1%), medium (0.1% < HI < 5%), and high (HI > 5%). A House Index (HI) threshold of 1% or less or a Breteau Index (BI) threshold of five or less was considered to prevent dengue transmission [55]. In our study, three studied districts revealed HI and BI of greater than 5% indicating a high risk of dengue infection. These elevated indices imply an increased likelihood of mosquito-human contact, significantly raising the potential for a dengue outbreak. However, because sampling was conducted over a limited post-monsoon period, seasonal variation in infestation indices could not be assessed, which should be considered when interpreting transmission risk. Similar indices in other regions have been associated with dengue outbreaks, reinforcing the necessity of proactive preventive strategies in these districts [56]. Though some researchers have suggested that BI alone is not the most reliable indicator of predicting dengue transmission [54,57,58], another study demonstrated that BI outperforms other indices as the most effective predictor of dengue infections [58]. Moreover, the striking dominance of dengue vectors in both lowland and highland districts, highlights the growing threat of arboviral transmission extending beyond Nepal’s traditionally endemic low altitude zones. Furthermore, the higher infestation levels observed in Kaski are likely due to rapid urban development, extensive water storage practice and accumulation of waste linked to tourism.
In our study, altitude was found not to have a significant effect on mosquito abundance (p = 0.563). The number of mosquitoes collected did not clearly show a trend with increasing altitude, despite variations in elevations across study sites. These findings suggest that mosquito distribution may be influenced by variables other than altitude, such as local environmental conditions, habitat availability, and microclimatic elements. In contrast, it has usually been shown that mosquito populations typically decrease as elevation rises, indicating the impact of elevation on mosquito abundance [29]. Moreover, environmental variables like temperature and humidity have a significant impact on this pattern. At lower elevations, mosquitoes can breed and survive better because of the warmer temperatures and higher humidity levels [59].
The dominance of Cx. quinquefasciatus, along with Ae. aegypti and Ae. albopictus, indicates the species’ ability to successfully colonize and thrive across a wide range of environmental conditions and altitudes, including both lowland and highland areas. Because of its remarkable ecological adaptability, Cx. quinquefasciatus can thrive in a variety of settings, including both urban and rural ones. These features are highlighted by its capacity to reproduce in contaminated waters and endure in environments that are unsuitable for other species [60]. When it comes to competitive interactions, Ae. aegypti and Ae. albopictus are frequently regarded as better competitors than Cx. quinquefasciatus. It has been found that, Cx. quinquefasciatus can take advantage of habitats when conditions become limited, especially during dry seasons, whereas Ae. albopictus tends to dominate during wet seasons [61]. This dominance suggests their ecological adaptability, competitive advantage, and potential role as key vectors in the transmission of mosquito-borne diseases within the study region. Moreover, our study showed the absence of adult Aedes despite the presence of their larvae. This may be due to temporal mismatch between larval and adult sampling, low adult resting densities or limited effectiveness of traps under local environmental conditions.
Overall, our study provided comprehensive data on the abundance of two dengue vector species and the principal vector of lymphatic filariasis along three altitudinally differing districts of Nepal. The main limitation of this study is that we could not relate our study in terms of seasons and climatic determinants, which are regarded as the important factors in determining the abundance and presence of those vectors [16,20,27]. Moreover, we couldn’t link our data with the number of DF cases because the mosquito population sampling was not conducted during dengue’s peak transmission period, which could lead to insufficient data to correlate between vector species and dengue fever incidence. The co-occurrence of Ae. aegypti and Ae. albopictus in breeding habitats definitely merits further research, as it represents a complex and so far underreported ecological phenomenon in Nepal with significant implications for dengue transmission dynamics and vector control efforts.
Supporting information
S1 TableDistrict wise abundance of mosquito immatures.(DOCX)
S2 TableDistrict-wise abundance of adult mosquitoes.(DOCX)
S3 TableContainer type associated with presence of Ae. aegypti, Ae. albopictus and their co-occurrence.(DOCX)
S4 TableBivariate logistic regression analysis for the district-wise occurrence of Ae. aegypti and Ae. albopictus.(DOCX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Franklinos LHV, Jones KE, Redding DW, Abubakar I. The effect of global change on mosquito-borne disease. Lancet Infect Dis. 2019;19(9):e 302–12. doi: 10.1016/S 1473-3099(19)30161-6 31227327 · doi ↗ · pubmed ↗
- 2Huang W, Wang S, Jacobs-Lorena M. Use of Microbiota to Fight Mosquito-Borne Disease. Front Genet. 2020;11:196. doi: 10.3389/fgene.2020.00196 32211030 PMC 7076131 · doi ↗ · pubmed ↗
- 3Dhimal M, Ahrens B, Kuch U. Climate Change and Spatiotemporal Distributions of Vector-Borne Diseases in Nepal--A Systematic Synthesis of Literature. P Lo S One. 2015;10(6):e 0129869. doi: 10.1371/journal.pone.0129869 26086887 PMC 4472520 · doi ↗ · pubmed ↗
- 4Takasaki T, Kotaki A, Nishimura K, Sato Y, Tokuda A, Lim CK, et al. Dengue virus type 2 isolated from an imported dengue patient in Japan: first isolation of dengue virus from Nepal. J Travel Med. 2008;15(1):46–9. doi: 10.1111/j.1708-8305.2007.00165.x 18217869 · doi ↗ · pubmed ↗
- 5Pandey BD, Morita K, Khanal SR, Takasaki T, Miyazaki I, Ogawa T, et al. Dengue virus, Nepal. Emerg Infect Dis. 2008;14(3):514–5.18325280 10.3201/eid 1403.070473 PMC 2570825 · doi ↗ · pubmed ↗
- 6Adhikari N, Subedi D. The alarming outbreaks of dengue in Nepal. Trop Med Health. 2020;48:5. doi: 10.1186/s 41182-020-0194-1 32055230 PMC 7007638 · doi ↗ · pubmed ↗
- 7Gyawali N, Johnson BJ, Dixit SM, Devine GJ. Patterns of dengue in Nepal from 2010-2019 in relation to elevation and climate. Trans R Soc Trop Med Hyg. 2021;115(7):741–9. doi: 10.1093/trstmh/traa 131 33197254 · doi ↗ · pubmed ↗
- 8Bijukchhe SM, Hill M, Adhikari B, Shrestha A, Shrestha S. Nepal’s worst dengue outbreak is a wake-up call for action. J Travel Med. 2023;30(7):taad 112. doi: 10.1093/jtm/taad 112 37584474 PMC 10658652 · doi ↗ · pubmed ↗