Candida albicans promotes TSST-1 production by Staphylococcus aureus through glucose depletion and relief of CcpA-mediated repression
Mathias Carriou, Cédric Badiou, Alexandre Soulard, Christophe d'Enfert, Karine Dufresne, Gérard Lina

TL;DR
Candida albicans increases TSST-1 production by Staphylococcus aureus by depleting glucose, which may raise the risk of menstrual toxic shock syndrome.
Contribution
This study reveals a novel metabolic interaction between C. albicans and S. aureus that enhances bacterial virulence.
Findings
C. albicans depletes glucose, which relieves CcpA repression of the tst gene in S. aureus.
Glucose depletion allows SaeRS to activate TSST-1 production in S. aureus.
C. albicans may act as a biomarker for increased mTSS risk in individuals with TSST-1-producing S. aureus.
Abstract
Menstrual toxic shock syndrome (mTSS) is a rare yet life-threatening disease that is caused when the opportunistic pathogenic bacterium Staphylococcus aureus releases the toxic shock syndrome toxin 1 (TSST-1) superantigen, which triggers systemic inflammation. The main risk factor for mTSS is prolonged use of intravaginal products, which facilitates S. aureus growth and TSST-1 production in menstrual blood. However, some studies suggest that the vaginal microbiota may also play a role in mTSS occurrence. A previous study reported that the presence of S. aureus in menstrual fluids was correlated with the simultaneous presence of Candida species, particularly C. albicans. Here, we assessed the potential involvement of C. albicans in the stimulation of TSST-1 production by incubating S. aureus strains with C. albicans culture supernatants and measuring TSST-1 production. We found that the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
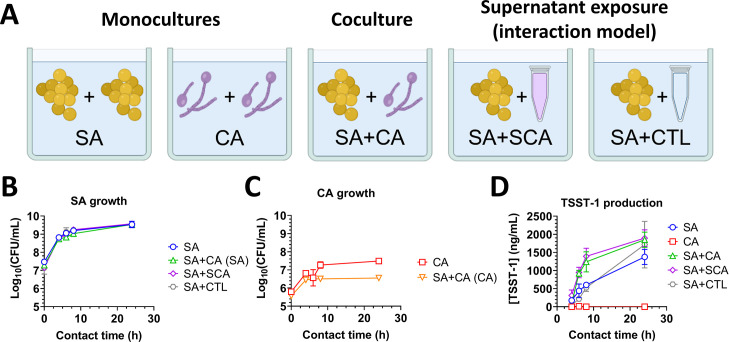 Fig 1
Fig 1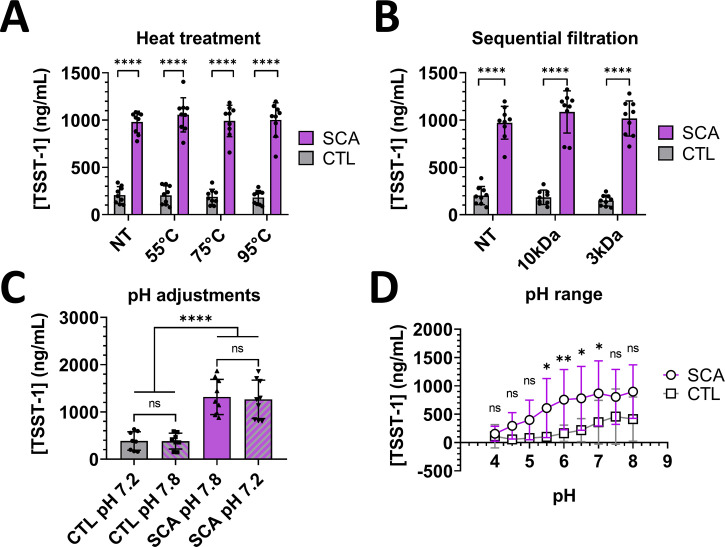 Fig 2
Fig 2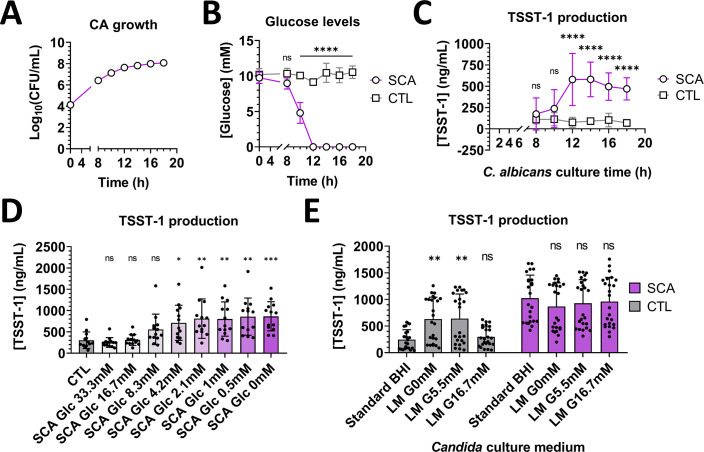 Fig 3
Fig 3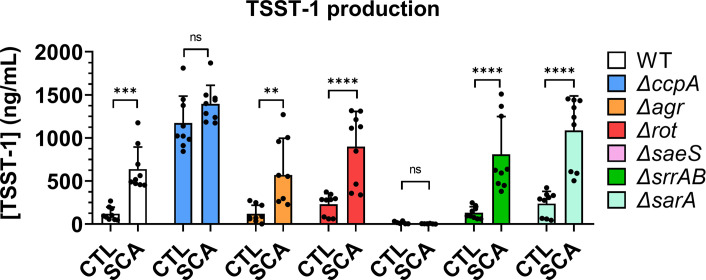 Fig 4
Fig 4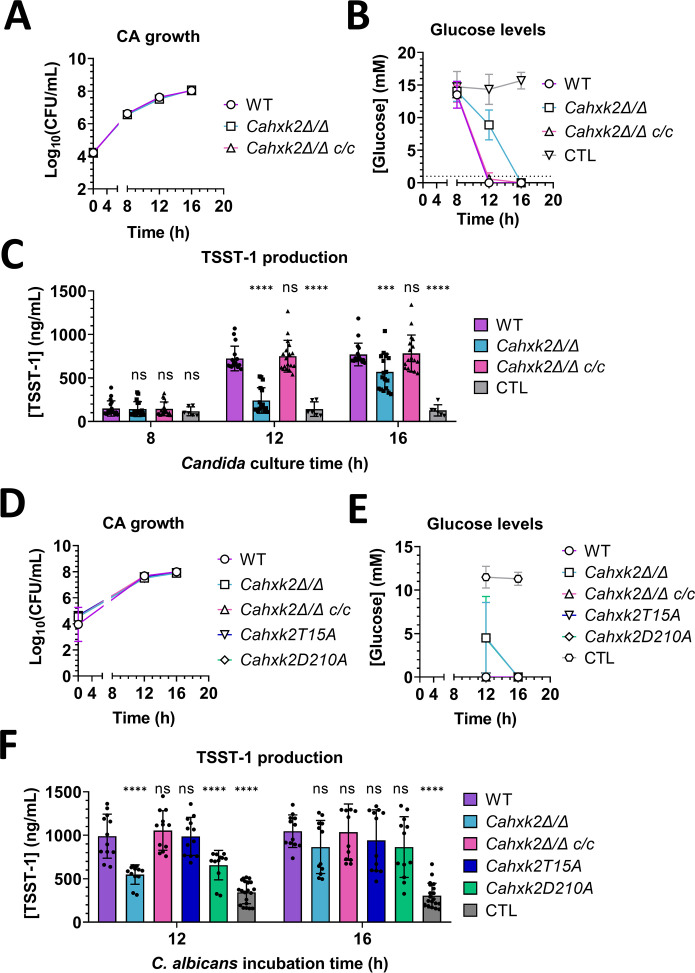 Fig 5
Fig 5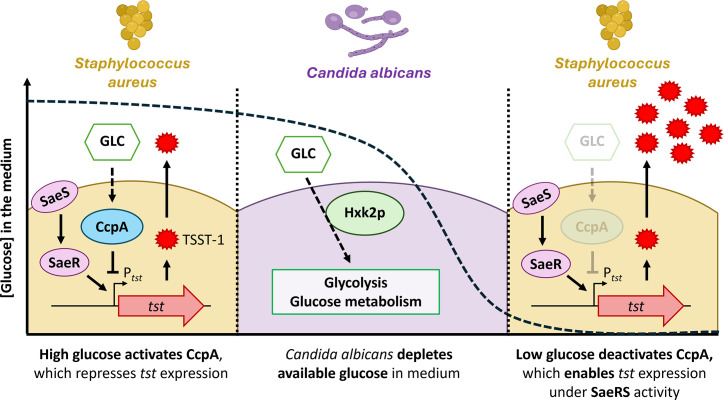 Fig 6
Fig 6| Strain name | Description | References |
|---|---|---|
|
| ||
| SC5314 WT | Wild-type reference strain |
|
| SC5314 | SC5314 with deletion of both | |
| SC5314 | SC5314 | |
| SC5314 | SC5314 | |
| SC5314 | SC5314 | |
|
| ||
| MN8 | TSST-1+ reference strain, isolated from a patient with TSS in the USA in 1980; USA200, CC30 |
|
| MN8 | MN8 with deletion of |
|
| MN8 | MN8 with the |
|
| MN8 | MN8 with deletion of |
|
| MN8 | MN8 with deletion of |
|
| MN8 | MN8 with deletion of |
|
| MN8 | MN8 with deletion of |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive tract infections research · Antimicrobial Resistance in Staphylococcus · Preterm Birth and Chorioamnionitis
INTRODUCTION
Staphylococcus aureus (S. aureus, SA) is a commensal bacterium found in 30% of the human population, which mainly colonizes the skin and nares and is detectable in the vaginal tract of 10% of healthy women (1, 2). Some S. aureus strains produce superantigen toxic shock syndrome toxin 1 (TSST-1), and their presence in the vulvovaginal environment is a major risk factor for menstrual toxic shock syndrome (mTSS). This rare yet severe condition occurs with an estimated incidence of 0.7 per 100,000 menstruating individuals and is characterized by symptoms including high fever, hypotension, macular erythroderma, and multisystem dysfunction (3–5). During menses, blood-soaked intravaginal menstrual products—such as tampons and cups—concentrate nutrients and oxygen in the vaginal niche, potentially promoting colonization and growth of S. aureus and enabling TSST-1 production (6, 7). TSST-1 can reach the bloodstream and force polyclonal activation of T cells, leading to massive proinflammatory cytokine release and causing severe mTSS symptoms (8).
TSST-1 is encoded by the tst gene, and its production is controlled by various regulators. Catabolite control protein A (CcpA) is a dominant glucose-dependent transcription repressor of the tst gene and is associated with carbon catabolite repression (CCR) (9, 10). Another major tst regulator is the SaeRS two-component system, encoded by the S. aureus exotoxin expression (sae) operon, which is reportedly required for tst expression (11). Additionally, the quorum-sensing accessory gene regulator (Agr) system positively regulates tst by activating transcription of the regulatory RNA RNAIII, which inhibits translation of the repressor of toxins (Rot) messenger RNA, thereby relieving tst repression (12, 13). Furthermore, the staphylococcal respiratory response (Srr) system can either enhance or inhibit tst transcription under high or low oxygen conditions, respectively (14, 15). Finally, the staphylococcal accessory regulator SarA exhibits nuanced control—inhibiting tst expression through direct binding to the tst promoter and activating RNAIII transcription (16).
To better prevent mTSS development, it is important to understand the associated risk factors, which may include the vaginal microbiota composition. Numerous studies report that Lactobacillus species—mainly L. crispatus, L. gasseri, and L. jensenii—play a protective role in healthy vaginal environments, notably by releasing lactic acid, which lowers the vaginal pH and thereby prevents pathogen proliferation (17–19). Studies have also examined how species within the vaginal microbiota can influence TSST-1 production by S. aureus, revealing that Lactobacillus iners and Gardnerella vaginalis may exacerbate toxin production and immune activation (20, 21). Despite results suggesting the potential importance of fungi in shaping the vaginal microbiota, little is presently known about molecular interactions between fungi and S. aureus in the vaginal niche (22–24). Jacquemond et al. previously demonstrated that the presence of S. aureus during menstruation showed strong positive associations with the presence of Globicatella, Prevotella, Gardnerella, and Candida species (25). Notably, Candida albicans (C. albicans, CA) comprises an average of 60% of Candida species and overall fungal populations found in the vaginal tract (26).
In the present study, we explored the role of C. albicans in the molecular regulation of TSST-1 production by S. aureus and, consequently, in mTSS development. We used C. albicans with mutations affecting glucose utilization, media with varying glucose levels, and mutant strains of S. aureus to assess the importance of glucose availability with regard to microbial interactions and TSST-1 production. Our results demonstrated that the growth of C. albicans depleted glucose levels in the medium, which lifted the CcpA-mediated repression of the tst gene and thereby enabled a two- to fivefold increase of TSST-1 production by S. aureus, mediated by SaeRS activity.
RESULTS
C. albicans culture supernatant stimulates TSST-1 production by S. aureus
To establish the experimental model for studying interactions between C. albicans and S. aureus, each organism was cultured separately (SA and CA), or co-cultured (SA + CA). S. aureus was also incubated in contact with either brain heart infusion (BHI) control medium (SA + CTL) or C. albicans culture supernatant obtained after 18 h of culture at 37°C (SA + SCA; Fig. 1A). At t = 0 h, bacterial counts were between 10^7^ and 2 × 10^7^ CFU/mL, while yeast counts were between 3 × 10^5^ and 4 × 10^5^ CFU/mL. This difference in cell counts is likely due to the different cell volumes. Yeast cells are 4–5 times larger than cocci; thus, achieving an equivalent biomass (initial OD_600nm_ of 0.1) will require more bacterial cells than yeast cells (27).
C. albicans stimulates TSST-1 production by S. aureus during coculture or through supernatant exposure. (A) Diagram showing the tested conditions for the interaction model, with S. aureus (SA), C. albicans (CA), C. albicans culture supernatant (SCA), and BHI control medium (CTL). CFU counts of S. aureus MN8 (B) and C. albicans S5314 (C) during monocultures, cocultures, or supernatant exposure, after incubation at 37°C for 4 to 24 h. (D) TSST-1 production by monocultures of S. aureus MN8 and C. albicans SC5314 cells, their coculture, and SA with CA supernatant or control BHI medium. The cumulative data were obtained from three independent experiments.
Under all conditions, C. albicans and S. aureus showed typical growth curves, reaching the stationary phase at 8 h post-inoculation (Fig. 1B). S. aureus growth did not significantly differ when incubated alone (SA), versus in coculture with C. albicans (SA+CA), incubated with C. albicans supernatant (SA+SCA), or with control medium (SA+CTL). C. albicans growth was slightly impaired by 4 h in coculture with S. aureus (SA+CA), compared to in monoculture (CA), which might result from earlier nutritional depletion of the medium (Fig. 1C). Interestingly, TSST-1 production by S. aureus was significantly increased after 6 and 8 h of contact with C. albicans (SA+CA) or with its supernatant (SA+SCA), compared to S. aureus incubated alone (SA) or with control medium (SA+CTL). TSST-1 levels produced by S. aureus did not significantly differ between coculture with C. albicans (SA+CA) or exposure to fungal supernatants (SA+SCA) for 6, 8, and 24 h (Fig. 1D). These results indicated that, in this interaction model, C. albicans did not interfere with S. aureus growth. Moreover, both C. albicans itself and its culture supernatant significantly and similarly stimulated TSST-1 production by S. aureus. This effect of C. albicans on TSST-1 production was likely mediated through an alteration of the medium by the yeast, rather than by direct interaction between the yeast and the bacterium.
Stimulation of TSST-1 production is mediated by heat-resistant low-molecular-weight molecules and does not depend on pH variations
To assess whether the stimulatory activity of C. albicans supernatant was mediated by heat-sensitive substances, we collected fungal supernatant after 18 h of culture at 37°C and heat treated these samples, along with control BHI medium, at various temperatures for 30 min. We then exposed S. aureus suspensions to the heat-treated C. albicans supernatant or control BHI medium and evaluated TSST-1 production. None of the heat treatments significantly altered the ability of C. albicans supernatant to stimulate TSST-1 production, and heat treatment of BHI medium did not affect basal TSST-1 production (Fig. 2A).
*Characterization of the stimulatory activity of C. albicans culture supernatant on TSST-1 production by S. aureus. TSST-1 production by S. aureus MN8 after 6 h of incubation with either C. albicans supernatant (SCA, purple bars, circles) or control BHI medium (CTL, gray bars, squares) following heat treatment (A), sequential filtration (B), or pH buffering (C, D). Statistical significance was tested by two-way analysis of variance and post-hoc Dunnett’s test (A, B, D), and Kruskal-Wallis test with post-hoc Dunnett’s test (C). *P < 0.05; **P < 0.001; ***P < 0.0001. The cumulative data were collected from three independent experiments. ns, not significant.
Next, we aimed to identify the molecular weight of the molecular mediators of TSST-1 production stimulation. C. albicans supernatant obtained from 18 h cultures at 37°C, and control BHI medium, was sequentially filtered to cut-offs of 10 kDa and 3 kDa, and then incubated with S. aureus and TSST1 production was quantified. The molecular weight cut-off showed no significant effect, either in comparisons of each supernatant with the corresponding control, or in comparisons among the supernatants or controls (Fig. 2B). Overall, these results demonstrated that the stimulation of TSST-1 production likely occurred through the accumulation or depletion of low-molecular-weight, heat-resistant peptides, compounds, or ions.
Low vaginal pH prevents pathogen proliferation; an increase in vaginal pH can drastically alter vaginal health (18). To assess the role of pH variations in the modulation of TSST-1 production, we adjusted the pH of supernatants from 18 h C. albicans cultures to that of the base BHI medium (pH = 7.2; 7.16 ± 0.05), and the pH of base BHI medium was buffered to that of 18 h culture supernatant of C. albicans (pH = 7.8; 7.63 ± 0.27). Fungal supernatant and control BHI medium were also adjusted from pH 4.0 (a healthy vaginal pH) to pH 8.0. Basal TSST-1 production did not significantly differ between control BHI media at pH 7.2 versus pH 7.8. Similarly, stimulation of TSST-1 production was not significantly affected by lowering the fungal supernatant pH to 7.2, compared to untreated supernatant at pH 7.8 (Fig. 2C). C. albicans supernatants significantly stimulated TSST-1 production only when buffered to pH values between 5.5 and 7. Additionally, TSST-1 production may have been stimulated, although to a non-significant degree, by supernatants of pH ranging from 4.5 to 8.0. TSST-1 production from S. aureus was higher when incubated with C. albicans supernatants at pH 6 to 8, compared to supernatants at pH 4 and 4.5. No significant difference in TSST-1 production was observed when comparing control medium at all tested pH values (Fig. 2D). Despite the high variability, these results indicated that pH itself was likely not directly responsible for stimulation of TSST-1 production by the yeast but did interfere with basal TSST-1 production. To summarize these first results, TSST-1 stimulation of S. aureus by C. albicans supernatants appeared to be mediated through the accumulation or depletion of low-molecular-weight, heat-resistant molecules, independently of pH modulation by C. albicans.
Stimulation of TSST-1 production correlates with glucose depletion in medium
Since TSST-1 production is controlled by glucose levels, we next investigated whether the stimulation of TSST-1 production was driven by C. albicans glucose consumption. C. albicans strain SC5314 WT was cultivated in BHI medium at 37°C for up to 18 h, and samples were collected at different time points for cell counting. Fungal supernatants were harvested and incubated for 6 h with S. aureus, followed by titration of TSST-1 production. Starting from an inoculum of 10^4^ CFU/mL, C. albicans showed standard growth curves for both biomass and viable cell counts, reaching stationary phase after around 12 h at 37°C (Fig. 3A). Concomitantly, the glucose levels in the fungal supernatant significantly decreased from 10 h of culture and fell to undetectable levels after 12 h of culture (Fig. 3B). TSST-1 production by S. aureus significantly increased after incubation with the 12 h culture supernatant of C. albicans (Fig. 3C). This suggested that the C. albicans-mediated stimulation of TSST-1 production by S. aureus was likely correlated with fungal growth phase and the glucose levels in the medium.
*Glucose levels control TSST-1 production in media. (A) Concentration of CFUs in cultures of C. albicans SC5314 (CA) incubated at 37°C from 0 to 18 h. (B) Glucose levels measured in control BHI medium (CTL) and C. albicans culture supernatant (SCA) obtained from cultures shown in (A). (C) TSST-1 production by S. aureus MN8 after 6 h of incubation with control BHI medium and C. albicans culture supernatant obtained from (A). (D) TSST-1 production by S. aureus MN8 after 6 h of incubation with glucose-supplemented C. albicans supernatants and control standard BHI (CTL). (E) TSST-1 production by S. aureus MN8 after 6 h of incubation with C. albicans supernatants obtained from 18 h cultures of C. albicans SC5314 in standard commercial BHI or laboratory-made BHI with either 0 mM (LM-G0), 5.5 mM (LM-G5.5mM), or 16.7 mM (LM-G16.7mM) of glucose. Statistical significance was tested using Kruskal-Wallis tests, followed by Dunn’s post-hoc test (D) and two-way analysis of variance with post-hoc Tukey’s tests (B, C, and E). *P < 0.05; **P < 0.001; **P < 0.0005. Cumulative data were obtained from three independent experiments. ns, not significant.
To confirm the effect of glucose on TSST-1 production in our interaction model, supernatants obtained from 18 h cultures of C. albicans were gradually supplemented with glucose from 0 mM to 33.3 mM (twice the initial glucose concentration of standard BHI medium) and filter-sterilized before the TSST-1 production stimulation assay. TSST-1 production was significantly modulated after contact with supernatants with glucose supplementation ranging from 0 mM to 4.2 mM. Supernatants with higher glucose supplementation were associated with low TSST-1 production, which did not significantly differ from the TSST-1 production obtained after incubation with control BHI medium (Fig. 3D).
*TSST-1 production by S. aureus strains lacking regulators of tst expression. TSST-1 production by S. aureus MN8 WT (white bars), ΔccpA (blue bars), Δagr (orange bars), Δrot (red bars), ΔsaeS (pink bars), ΔsrrAB (green bars), and ΔsarA (teal bars) after 6 h of incubation with 18 h C. albicans culture supernatant (SCA) or control BHI medium (CTL). Statistical significance was tested by two-way analysis of variance with post-hoc Tukey’s tests. *P < 0.05; **P < 0.001; ***P < 0.0005; ***P < 0.0001. Cumulative data were obtained from three independent experiments. ns, not significant.
Although glucose levels in C. albicans supernatants clearly affected TSST-1 production by S. aureus, it could not be excluded that other molecules accumulated or depleted in supernatants might be involved in TSST-1 production. Therefore, we conducted further tests to confirm whether glucose was the sole regulator of the stimulation of TSST-1 production in our interaction model. Laboratory-made (LM) BHI with glucose supplementation at 0 mM (LM-G0mM), 5.5 mM (LM-G5.5mM), and 16.7 mM (LM-G16.7mM) was used for C. albicans growth for 18 h, and the supernatant effects on S. aureus were investigated in comparison to standard commercial BHI (base glucose concentration of 16.7 mM). The basal TSST-1 production with control LM-G0 and LM-G5.5mM BHI media was significantly higher compared to standard and LM-G16.7mM BHI. Stimulation of TSST-1 production by S. aureus was significantly higher than controls only for supernatants from C. albicans grown in standard, LM-G5.5mM, and LM-G16.7mM BHI. Supernatants of yeast grown in glucose-free LM BHI exhibited no significant effect on TSST-1 production compared to the controls (Fig. 3E). Overall, these results indicated that, in the present interaction model between C. albicans and S. aureus, C. albicans supernatant stimulated TSST-1 production by S. aureus mainly due to glucose levels.
CcpA and SaeRS control TSST-1 production by S. aureus during exposure to C. albicans culture supernatant
In S. aureus, glucose-mediated repression of tst expression is controlled by CcpA (9, 10). To confirm the CcpA-controlled glucose-mediated repression of TSST-1 production, and to further study the regulation of the tst gene in this interaction model, S. aureus strains lacking the ccpA gene or other known regulators of tst expression—namely agr, rot, saeS, srr, and sarA—were incubated with C. albicans supernatant or control standard BHI medium. Only the ΔccpA and ΔsaeRS mutants displayed TSST-1 production profiles different from the WT profile. Specifically, ΔccpA showed high basal TSST-1 production, both with C. albicans supernatants and control BHI medium, whereas the ΔsaeRS strain showed low to no TSST-1 production. The Δagr, Δrot, and Δsrr strains displayed basal TSST-1 production similar to WT. Additionally, ΔsarA produced a profile similar to the WT strain, although TSST-1 production after contact with SCA was significantly higher for the ΔsarA strain compared to the WT strain (Fig. 4).
Stimulation of TSST-1 production by C. albicans involves hexokinase activity of fungal Hxk2p
WT and slow-glucose-depleting mutant strains of C. albicans were used to confirm that glucose depletion was due to C. albicans growth and metabolism. The HXK2 gene encodes the hexokinase Hxk2p, which is one of the three glucose-phosphorylating enzymes in C. albicans that ensures glucose import and phosphorylation, for the first step of glycolysis. Deletion of both HXK2 alleles leads to slower glucose depletion in culture medium (28, 29). WT, Cahxk2Δ/Δ, and complemented (Cahxk2Δ/Δ c/c) strains were cultivated, their CFU counts determined, and their supernatants collected for a TSST-1 production stimulation assay. The three strains displayed similar growth over time (Fig. 5A). After 12 h, the remaining glucose concentration was significantly higher with the Cahxk2Δ/Δ strain, compared to the WT and complemented strains. At 16 h of culture, the glucose concentrations were undetectable for all three strains (Fig. 5B). TSST-1 production by S. aureus was negatively correlated with the glucose levels in supernatants. Indeed, S. aureus produced significantly less TSST-1 during exposure to 12 h Cahxk2Δ/Δ supernatants, compared to exposure to 12 h supernatants of WT and complemented strains. However, the 16 h culture supernatants of all three strains increased TSST-1 production by S. aureus to the same levels (Fig. 5C).
*Glucose depletion and fungal hexokinase Hxk2p involvement in the C. albicans (CA)-mediated stimulation of TSST-1 production by S. aureus. (A and D) Concentration of CFUs and (B and E) glucose levels measured in cultures of C. albicans SC5314 WT (purple, circles), Cahxk2Δ/Δ (light blue, squares), Cahxk2Δ/Δ c/c (pink, right-side-up triangles), Cahxk2Δ/Δ T15A (dark blue, upside-down triangles), and Cahxk2Δ/Δ D210A (green, rhombus) at 37°C from 8 to 16 h, and control BHI medium (gray, hexagon). (C and F) TSST-1 production by S. aureus MN8 after 6 h of incubation with supernatants from WT and mutant strains of C. albicans, and control BHI medium. Statistical significance was tested using two-way analysis of variance and post-hoc Tukey’s tests. *P < 0.05; **P < 0.001; ***P < 0.0001. Cumulative data were collected from at least two independent experiments for each graph. ns, not significant.
In addition to its enzymatic activity, Hxk2p is a transcriptional regulator controlling the expression of glucose-repressed genes, associated with the CCR of C. albicans (29, 30). To assess which function was affecting TSST-1 production stimulation, yeast mutant strains lacking either regulatory (Cahxk2T15A) or catalytic (Cahxk2D210A) activity of Hxp2p were cultivated, followed by CFU counts, supernatant recovery, and use in a TSST-1 stimulation assay. At 12 and 16 h, the Cahxk2D210A strain showed growth and glucose depletion profiles similar to those of the Cahxk2Δ/Δ strain. In contrast, the Cahxk2T15A strain displayed 12 h and 16 h growth and glucose depletion profiles similar to those of the WT and Cahxk2Δ/Δ c/c strains (Fig. 5D and E). At 12 h of culture, culture supernatants of the Cahxk2Δ/Δ and Cahxk2D210A strains failed to stimulate TSST-1 production to the same degree as supernatants obtained from the WT, complemented, and Cahxk2T15A strains (Fig. 5F). This difference between strains was lost at 16 h of cultures, at which all five strains stimulated TSST-1 production to similar levels. These findings confirmed that C. albicans glucose metabolism led to the depletion of glucose in medium, and this process partially relied on the enzymatic function of Hxk2p. Moreover, it was this depletion that elevated TSST-1 production by S. aureus, according to our interaction model.
Overall, based on our interaction model of S. aureus with C. albicans culture supernatants, we showed that TSST-1 production was increased two- to fivefold, and this stimulation was mainly dependent on glucose levels in the medium. C. albicans depletes glucose levels during its growth, enabling early release of CcpA-mediated glucose repression of the tst gene, which then allows SaeRS-driven gene expression, thereby promoting TSST-1 production.
DISCUSSION
mTSS is a rare severe disease. While awareness about safe use of intravaginal menstrual products has improved, other less controllable factors, such as vaginal microbiota, may affect mTSS risk. In the present study, we demonstrated that the opportunistic fungal pathogen C. albicans may promote virulence in S. aureus. Specifically, C. albicans depletes glucose levels and thereby lifts CCR of the tst gene, which is mediated by CcpA, consequently enabling SaeRS-driven transcription of tst (Fig. 6).
Hypothetical model of the stimulation of TSST-1 production by S. aureus through interaction with C. albicans. SaeS, S. aureus exotoxin expression sensor histidine kinase; SaeR, SaeS-associated response element; Glc, glucose; tst, TSST-1-encoding gene; Ptst, tst promoter; Hxk2p, hexokinase 2. Solid arrows and dotted arrows indicate direct and indirect action, respectively.
In our study, glucose depletion was identified as the main signal stimulating TSST-1 production in the interaction between C. albicans and S. aureus. Glucose levels in cervicovaginal mucus vary over the menstrual cycle, peaking at around 4.4–8.3 mM during the post-ovulatory phase (31). The average bloodstream glucose levels are reportedly around 6 mM (32). Exact glucose concentrations from menstrual fluid samples have not yet been reported. Alterations of glucose availability might result in drastic variations of TSST-1 production and higher mTSS risk during menstruation. Vaginal glucose also originates from glycogen, a large and ramified glucose polymer produced by vaginal and cervical epithelial cells (33). Glycogen can be broken down into glucose by the local microbiota, especially Lactobacillus species (34–36). Glycogen is crucial for lactic acid production and Lactobacillus colonization, but disruptions in Lactobacillus abundance can lead to vaginal dysbiosis and pathogen proliferation (37). Glycogen can also be degraded by vaginal pathogens such as Gardnerella species and C. albicans (38, 39). No reports have described glycogen transport and degradation by S. aureus and its potential effects on virulence and metabolism. Thus, there remains a need for additional studies on how the vaginal microbiota metabolizes vaginal nutrients and their effects on the virulence of pathogens, such as S. aureus.
Although our present analyses demonstrated that TSST-1 production was mainly affected by glucose levels, we cannot exclude the potential effects of other metabolites, which could include naturally occurring amino acids, amino-acid-derived metabolites, and heat-stable peptides (40, 41). Further studies are needed to explore the whole exometabolome of C. albicans in its interaction with S. aureus and their effect on TSST-1 production and the overall virulence of S. aureus. Furthermore, the molecular composition of menstrual fluids differs from that of BHI medium, despite their similar neutral pH and various nutrient sources (42). While previous studies have used media simulating vaginal and cervical secretions, no defined medium representing menstrual fluid has yet been published (43).
In our interaction model, only CcpA seemed to be significantly involved in the stimulation of TSST-1 production by C. albicans, as SaeRS likely activates tst expression independently of C. albicans and glucose depletion. Deletions of ccpA and saeRS yielded results consistent with previously published findings that CcpA is a major tst repressor, and SaeRS is a dominant activator of tst expression (9–11). On the other hand, deletions of agr and rot did not lead to altered TSST-1 production under our experimental conditions. Agr and Rot are well-described regulators of exotoxin production, including TSST-1. Tuffs et al*.* (13) demonstrated that Rot overexpression led to low levels of tst expression, whereas wild-type levels of Rot seemed to enable relatively high tst expression in the MN8 strain, compared to a Δrot strain (13, 44). Corroborating these results, Dufresne et al*.* showed that rot deletion also led to lower tst transcription in a medium simulating vaginal fluids (vaginally defined medium, VDM); they also reported no tst expression in a Δagr strain grown in VDM (45).
Among other described tst regulators, the redox-sensitive SrrAB system reportedly regulates tst expression according to oxygen levels. SrrAB seems to both inhibit tst expression under microaerobic conditions and enhance tst expression under aerobic conditions (14, 15). Dufresne et al*.* reported near-complete extinction of tst expression under aerobic conditions in the MN8 ΔsrrAB strain (45). Intravaginal menstrual products enable oxygen to enter the microaerophilic vaginal environment, such that SrrAB may be deactivated, enabling TSST-1 production (6, 46). Our interaction model was maintained under aerobic conditions, relevant to mTSS; therefore, the absence of SrrAB-mediated repression of tst expression is consistent with previous observations related to TSST-1 inhibition under microaerobic conditions. Finally, although previous reports have demonstrated the role of SarA in the direct and indirect regulation of TSST-1 production (16, 44), in our present study, sarA deletion did not alter the stimulation of TSST-1 production by C. albicans culture supernatants.
Furthermore, we used variants of fungal Hxk2p to confirm that glucose depletion was indeed caused by C. albicans glucose consumption and metabolism. Our results show that this process required the enzymatic activity of Hxk2p but not its regulatory activity. In C. albicans, Hxk2p has been reported as a co-regulator, controlling the expression of various genes associated with CCR, along with Mig1p and Mig2p, two transcription factors that are activated when glucose levels are high (29, 47). It is possible that the impairment of Hxk2p regulatory activity in our study might not have been sufficient to alter the CCR. Additional studies using C. albicans strains harboring deletions of MIG1, MIG2, and HXK2 could be useful for better deciphering the possible impact of CCR disturbance on the interaction between C. albicans and S. aureus.
Previous studies of C. albicans–S. aureus interactions have described the stimulation of alpha-toxin (or alpha-hemolysin) production by S. aureus during coculture with C. albicans (48, 49). Another study revealed the importance of ribose during this interaction, which is depleted by C. albicans during coculture and enables Agr activation through unclear molecular mechanisms (50). The available literature does not include any report of the ribose concentration within the vaginal mucosa, further highlighting the importance of additional studies on the precise metabolite composition of vaginal and menstrual fluids. Notably, in our interaction model, pH alkalinization, agr, and rot were not necessary for the stimulation of TSST-1 production, which may hint at the involvement of different molecular mechanisms.
In the present study, we only examined the effect of C. albicans on S. aureus, and not the other way around, since the investigation was focused on TSST-1 production and mTSS. Moreover, this study was specifically designed to focus on the effects of C. albicans culture supernatants on TSST-1 production by S. aureus. Although TSST-1 production did not differ between this model and cocultures, the interaction between the two organisms might have involved other mechanisms based on molecular crosstalk. Furthermore, we only studied the interaction between C. albicans and S. aureus since C. albicans comprises most of Candida species and the overall fungal population of the vaginal microbiota (26, 51). However, the decrease in glucose levels in menstrual secretions is probably not specific to C. albicans, and it is highly likely that other microorganisms in the vaginal microbiota can also decrease glucose levels in the presence of S. aureus and induce stimulation of TSST-1 production. MacPhee et al*.* (20) demonstrated that supernatants of Streptococcus agalactiae culture in BHI significantly increased tst expression, while supernatants of Gardnerella vaginalis and Lactobacillus iners cultures decreased tst expression (20). Maduta et al. (2024) showed that supernatants of Lactobacillus iners and Gardnerella vaginalis cultures in VDM were able to increase TSST-1 production by the MN8 ΔccpA strain and enhance T-cell activation in both WT and MN8 ΔccpA strains (21). These studies did not investigate glucose levels in the microbial culture supernatants. Despite the inconsistencies, which are likely due to the use of different media, both studies underscore the complexity of microbial interactions in the vaginal ecosystem, particularly related to TSST-1 production, metabolite availability, including glucose, and mTSS development. Additionally, no published report describes an association between mTSS development and the presence of Candida species within the vaginal microbiota. Given the results of previous studies, it is likely that mTSS onset and development may be associated with the specific composition of vaginal microbiota that can be isolated from menstrual fluids, including C. albicans. Since C. albicans represents around 65% of the total fungal vaginal population, and due to its efficient uptake of nutrients, especially glucose, it might quickly colonize and proliferate within menstrual blood, alongside S. aureus (26, 52, 53). This could mean that C. albicans could expedite glucose depletion, leading to early relief of CcpA-mediated repression of tst expression in S. aureus and thereby enabling the production of more TSST-1 and triggering mTSS.
In conclusion, here we demonstrated that supernatants from C. albicans cultures could stimulate TSST-1 production by S. aureus. This occurred because C. albicans growth depleted the glucose levels in media, which relieved repression of tst gene expression by CcpA and enabled SaeRS-mediated transcription of tst. These findings highlight the significance of metabolic interplay within the vaginal microbiota in disease development. Given the central role of TSST-1 in mTSS, our present findings underscore the likely importance of metabolic interactions within the vaginal microbiota in the development of this disease. This study also highlights the broader conceptual intersection of microbial metabolism and virulence, suggesting that metabolic interactions could be a pivotal parameter affecting the onset and severity of polymicrobial infections.
MATERIALS AND METHODS
Strains and media
Table 1 lists all strains used in the study. Strains were streaked onto plates of Columbia Agar containing 5% sheep blood (bioMérieux, France) and incubated for 24 h at 37°C for S. aureus and 48 h at 30°C for C. albicans. To standardize the physiological state before each experiment, the strains were pre-cultivated in BHI liquid medium (BBL BHI, BD 211059), under agitation at 200 rotations per minute (rpm), for 18 h at 37°C for S. aureus, and 24 h at 30°C for C. albicans. Dilution-plating counts were realized by plating mono-microbial suspensions on trypticase soy agar (TSA; BD). Polymicrobial suspensions were plated on Sabouraud agar (Bacto tryptone 5 g/L, Bacto proteose peptone 5 g/L, glucose 40 g/L, and agar 15 g/L) for C. albicans isolation or on mannitol salt agar (#236950, BD) for S. aureus isolation. LM-BHI was prepared based on the composition of commercial standard BBL BHI (BD, #211059), including BHI solids 6 g/L, peptone from animal tissue (Peptone A) 6 g/L, peptone from pancreatic digest of gelatin (Peptone G) 14.5 g/L, NaCl 5 g/L, and Na_2_HPO_4_ 2.5 g/L. After dissolution of powders and salts by stirring, LM-BHI was filter sterilized using Nalgene Rapid-Flow 75-mm filter units (Thermo Scientific). Next, D-glucose was added into LM-BHI up to 16.7 mM (LM-G16.7mM), the solution was filter sterilized again, and lower glucose concentrations were prepared by diluting LM-G16.7mM with LM-G0mM.
C. albicans culture supernatant, treatments, and glucose titration
Overnight pre-cultures of C. albicans strains were grown in 5 mL BHI, with incubation at 37°C under agitation in DeepWell24 plates (Axygen), to a final OD_600 nm_ of 0.001 (10^4^ CFU/mL). For experiments using LM-BHI, 2 mL samples were collected from the precultures in standard commercial BHI and centrifuged at 15,000 × g for 5 min. Then the supernatants were discarded, and the pellets were resuspended in sterile DPBS 1× (Thermo Fisher). The samples were washed in this manner twice and then adjusted to an OD_600 nm_ of 0.001 in LM-BHI.
Cultures were collected at each time point and for each condition, and 1 mL aliquots were used for cell counting by OD_600 nm_ measurement and dilution plating. Culture supernatants were obtained after centrifugation for 30 min at 2,600 × g at 4°C and were stored at −20°C until use. Unless otherwise stated, supernatants were obtained from cultures of the SC5314 WT strain after 18 h of growth at 37°C under shaking (OD_600 nm_ ≈ 4–6; 10^8^ CFU/mL). C. albicans supernatants were supplemented with glucose by adding 6 g of D-glucose per liter of supernatant (16.7 mM), followed by filter sterilization. Next, these supplemented supernatants were twofold serially diluted to a final concentration of 0.5 mM (0.09 g/L). For heat treatment, 2 mL aliquots of 18 h culture supernatants were heated to 55°C, 75°C, and 95°C in a dry block for 30 min. These temperatures were chosen to establish the thermal stability of regulatory compounds in the media and supernatants and to help with their identification. For sequential filtration, supernatants were first filtered using Vivaspin centrifugal concentrators (Sartorius) with a 10 kDa molecular weight cut-off, then refiltered with a 3 kDa cut-off. The final filtrates for each cut-off were filter sterilized using 0.22 µm syringe filters (Clearline, Dutscher). For pH adjustments, supernatants and control media were buffered to final pH values using HCl 1 M or NaOH 1 M and then filter sterilized. In media and supernatants, the glucose concentration was measured using a Tyson Bio TB100 glucometer and its compatible bands, according to the user manual. The supplied control solution was used to calibrate the glucometer before each measurement session and before using each new test strip batch. Then, 10 µL drops of each sample were placed onto the strip reactive zone to measure glucose levels within the range 1.1–33.3 mmol/L. Concentrations lower than 1.1 mmol/L were reported as undetectable and were considered equal to 0.
S. aureus exposure to microbial supernatants and cocultures
Overnight pre-cultures of S. aureus and C. albicans were diluted to an OD_600 nm_ of 0.1 in BHI. Cocultures were established by mixing 100 µL of S. aureus suspension with 100 µL of C. albicans suspension in 96-well microplates (#351172, Falcon). For supernatant exposure, 100 µL of S. aureus suspension was mixed with 100 µL of C. albicans supernatants or control medium. For monocultures, S. aureus or C. albicans suspensions were prepared at a starting OD_600 nm_ of 0.1 in 200 µL. All cultures were incubated at 37°C for 4–24 h, under agitation at 200 rpm. Supernatants from cocultures and fungal supernatant exposures were collected after 30 min of centrifugation at 2,600 × g and at 4°C and were stored at −20°C until use.
Toxin titration by enzyme-linked immunosorbent assay
Toxins produced in contact with supernatants were quantified by sandwich enzyme-linked immunosorbent assay (57). Anti-TSST-1 capture antibodies (monoclonal) and anti-TSST-1 secondary antibodies (F[ab’]2 fractions, polyclonal, biotin-coupled) were obtained from AgroBio. Detection was performed by adding horseradish-peroxidase-coupled streptavidin, 3,3',5,5'-tetramethylbenzidine substrate (Seracare), and then H_2_SO_4_ 1 M was added to stop the reaction. Colorimetric signals were read at λ = 450 nm using a Biorad microplate reader.
Statistical analysis and figure creation
Statistical analyses were performed, and graphs were constructed using GraphPad PRISM. In the graphs, each dot represents one biological replicate, each originating from one initial isolated colony. Graphs show the mean values for each group, with error bars representing the standard deviation, using cumulative data from several independent experiments. Data were tested for significance using appropriate tests, including the Kruskal-Wallis test and two-way analysis of variance with appropriate post-hoc tests for between-group comparisons. For all tests, a P value of <0.05 was considered to indicate statistical significance. Diagrams were created using BioRender and Microsoft PowerPoint.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wertheim HFL, Melles DC, Vos MC, van Leeuwen W, van Belkum A, Verbrugh HA, Nouwen JL. 2005. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect Dis 5:751–762. doi:10.1016/S 1473-3099(05)70295-416310147 · doi ↗ · pubmed ↗
- 2Guinan ME, Dan BB, Guidotti RJ, Reingold AL, Schmid GP, Bettoli EJ, Lossick JG, Shands KN, Kramer MA, Hargrett NT, Anderson RL, Broome CV. 1982. Vaginal colonization with Staphylococcus aureus in healthy women: a review of four studies. Ann Intern Med 96:944–947. doi:10.7326/0003-4819-96-6-9447046548 · doi ↗ · pubmed ↗
- 3Tong SYC, Davis JS, Eichenberger E, Holland TL, Fowler VG. 2015. Staphylococcus aureus infections: epidemiology, pathophysiology. Clinical Manifestations, and Management. Clin Microbiol Rev 28:603–661. doi:10.1128/CMR.00134-1426016486 PMC 4451395 · doi ↗ · pubmed ↗
- 4Berger S, Kunerl A, Wasmuth S, Tierno P, Wagner K, Brügger J. 2019. Menstrual toxic shock syndrome: case report and systematic review of the literature. Lancet Infect Dis 19:e 313–e 321. doi:10.1016/S 1473-3099(19)30041-631151811 · doi ↗ · pubmed ↗
- 5Schlievert PM, Davis CC. 2020. Device-associated menstrual toxic shock syndrome. Clin Microbiol Rev 33:e 00032-19. doi:10.1128/CMR.00032-1932461307 PMC 7254860 · doi ↗ · pubmed ↗
- 6Wagner G, Bohr L, Wagner P, Petersen LN. 1984. Tampon-induced changes in vaginal oxygen and carbon dioxide tensions. Am J Obstet Gynecol 148:147–150. doi:10.1016/s 0002-9378(84)80165-96419612 · doi ↗ · pubmed ↗
- 7Schlievert PM. 1985. Effect of magnesium on production of toxic-shock-syndrome toxin-l by Staphylococcus aureus. Journal of Infectious Diseases 152:618–620. doi:10.1093/infdis/152.3.6184031559 · doi ↗ · pubmed ↗
- 8Tuffs SW, Haeryfar SMM, Mc Cormick JK. 2018. Manipulation of innate and adaptive immunity by staphylococcal superantigens. Pathogens 7:53. doi:10.3390/pathogens 702005329843476 PMC 6027230 · doi ↗ · pubmed ↗