DtxR is a global iron-dependent regulatory protein with multiple roles in the control of gene expression in Corynebacterium diphtheriae
Lindsey R. Lyman, Eric D. Peng, Michael P. Schmitt

TL;DR
This study explores how the DtxR protein controls gene expression in the bacteria that causes diphtheria, revealing new genes influenced by iron and DtxR.
Contribution
The study identifies novel iron- and DtxR-regulated genes in Corynebacterium diphtheriae through a comprehensive transcriptomic analysis.
Findings
Iron and DtxR regulate multiple genes in C. diphtheriae, including repression and induction of transcription.
The RipA repressor is regulated by iron and DtxR, with its gene transcription being repressed under these conditions.
DtxR binds to sequences upstream of newly identified genes, and ferritin gene expression requires multiple DtxR binding sites.
Abstract
Corynebacterium diphtheriae is the causative agent of diphtheria, a severe respiratory disease in humans. Diphtheria toxin, encoded by the tox gene, is a potent exotoxin secreted by C. diphtheriae that is responsible for much of the morbidity and mortality of diphtheria. Expression of the tox gene is regulated by iron and the diphtheria toxin repressor (DtxR). In addition to controlling toxin expression, DtxR functions as a global iron-dependent regulatory factor that mediates iron homeostasis in C. diphtheriae. While numerous genes regulated by DtxR and iron are known, a genome-wide study of both the iron and DtxR regulons is lacking in C. diphtheriae. Here, we report novel iron- and DtxR-regulated genes revealed by a comprehensive transcriptomic analysis in C. diphtheriae. The findings from the study show that iron and DtxR not only result in the repression of transcription of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
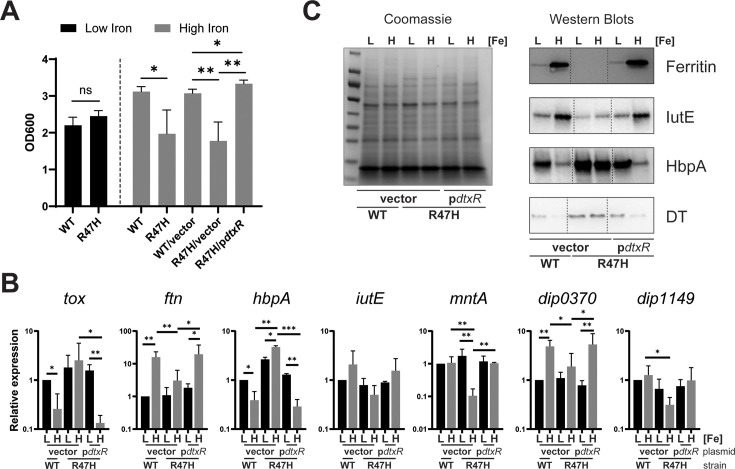 Fig 1
Fig 1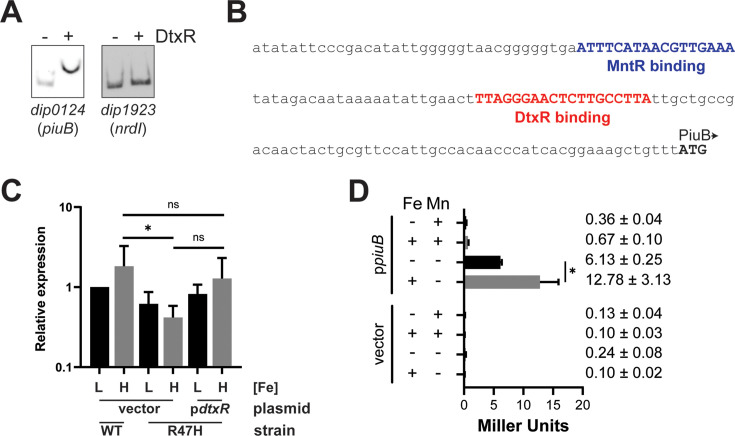 Fig 2
Fig 2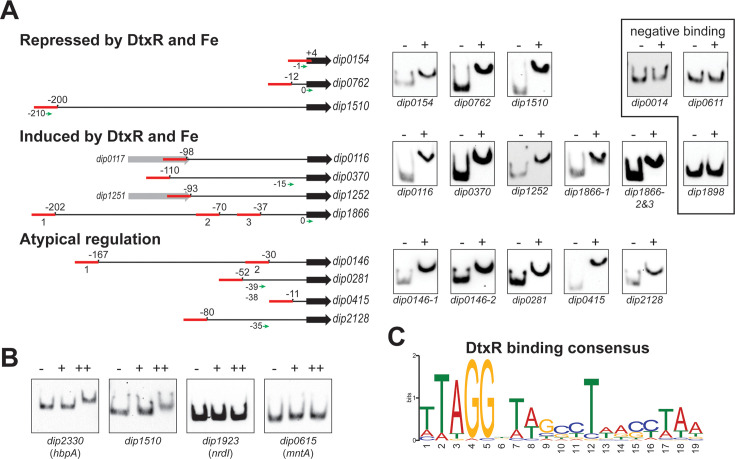 Fig 3
Fig 3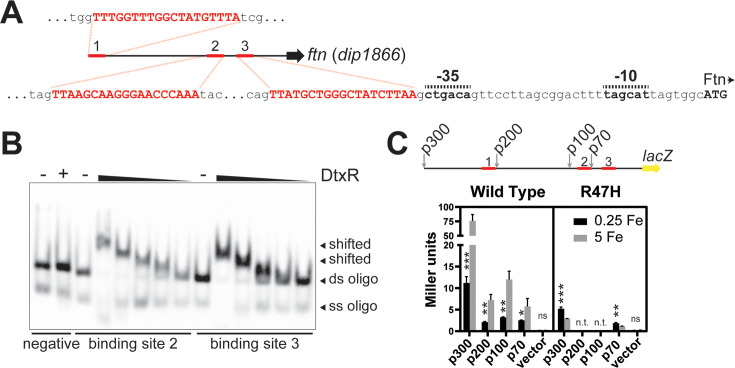 Fig 4
Fig 4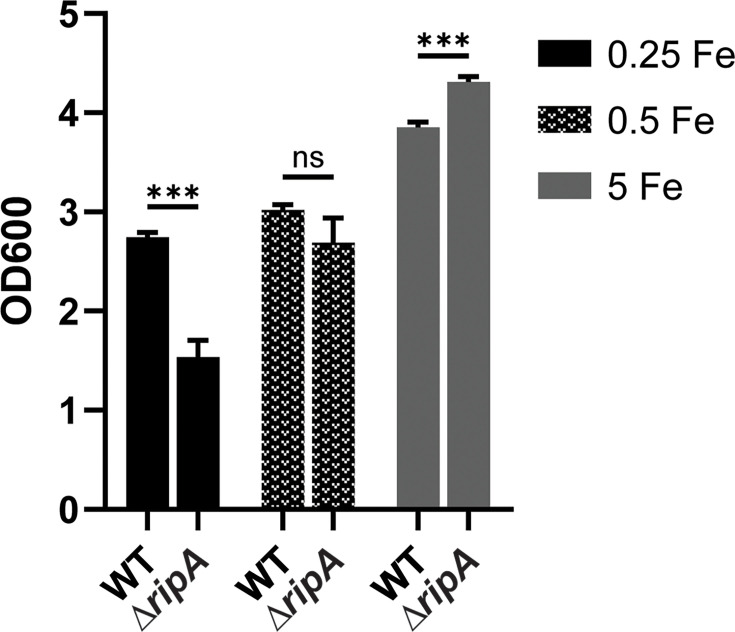 Fig 5
Fig 5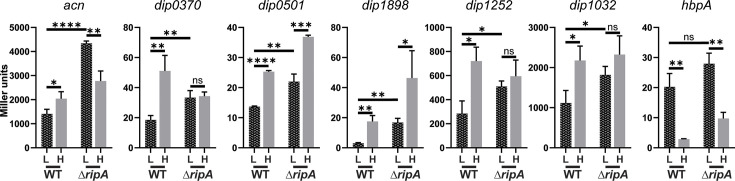 Fig 6
Fig 6| Locus | Gene description | Log2 fold change | ||
|---|---|---|---|---|
| wt | wt:R47H | R47H | ||
|
| ABC transporter | −5.06 | −4.76 | |
|
| Transport system, ATP-binding protein | −4.37 | −3.94 | |
| DIP0108 ↓ | −4.31 | −4.19 | ||
| DIP0109 | −3.48 | −3.78 | ||
|
| −3.53 | −3.25 | ||
|
| Putative lipoprotein | −1.27 | −1.39 | |
|
| Putative endopeptidase | −3.00 | −2.61 | |
| Putative secreted protein | −1.15 | −1.27 | ||
|
| Putative membrane protein | −1.42 | −1.30 | |
|
| Putative lipoprotein-ABC transporter | −1.45 | −1.63 | |
| DIP0169 ↓ | −5.89 | −5.38 | ||
| DIP0170 | −4.30 | −5.12 | 1.27 | |
| DIP0171 | −3.86 | −4.65 | 1.62 | |
|
| −2.16 | −2.86 | 1.51 | |
| DIP0222 | −2.32 | −1.33 | ||
| ABC transport system ATP-binding protein; (dip0321-0325) | −1.21 | −1.58 | ||
|
| ABC transport system ATP-binding protein | −1.38 | −1.51 | |
|
| ABC transport system membrane protein | −1.34 | −1.38 | |
|
| Putative membrane protein; DUF418 | −2.22 | −2.28 | |
|
| −6.78 | −7.28 | ||
| DIP0523 ↑ | −6.88 | −7.34 | ||
|
| Putative membrane protein | −1.37 | −2.83 | 2.22 |
|
| DUF5319 domain-containing protein | −2.63 | −2.91 | |
| −1.00 | −1.71 | |||
| DIP0582 ↓ | −4.95 | −5.04 | ||
| DIP0583 | −4.30 | −4.79 | ||
| DIP0584 | −3.69 | −4.67 | 1.46 | |
|
| −4.43 | −3.54 | ||
| DIP0586 ↓ | −6.71 | −6.57 | ||
|
| −4.13 | −3.43 | ||
|
| Putative ABC transport system lipoprotein | −1.38 | −4.39 | 2.85 |
|
| Putative membrane protein | −1.44 | −2.50 | 1.40 |
|
| Putative membrane protein | −2.05 | −3.02 | 1.20 |
|
| −4.46 | −5.20 | ||
| DIP0624 ↑ | −8.94 | −9.39 | ||
| DIP0625 ↓ | −7.17 | −6.83 | ||
| DIP0626 | −6.88 | −6.28 | ||
| DIP0627 | −5.43 | −5.13 | ||
|
| −3.80 | −3.38 | ||
| DIP0629 | −4.90 | −5.02 | ||
|
| Transposase | −1.19 | −2.74 | 1.29 |
|
| IS element transposase (partial) | −1.16 | −2.95 | |
|
| Cupin type-2 domain-containing protein | −3.11 | −2.28 | |
| Methyltransferase type 11 domain-containing protein | −5.98 | −4.96 | ||
|
| Peptidase M23 domain-containing protein | −1.04 | −1.08 | |
| DIP0922 | −7.30 | −6.56 | ||
|
| −2.56 | −2.69 | ||
| DIP1060 | −3.00 | −2.95 | ||
| DIP1061 ↑ | −3.12 | −2.91 | ||
| DIP1062 | −4.10 | −4.15 | ||
|
| HD domain-containing protein | −1.60 | −1.28 | |
|
| −2.00 | −2.56 | ||
| DIP1518 | Insertion element DNA-binding protein | −2.71 | −2.37 | |
|
| −6.46 | −6.05 | ||
| DIP1520 ↑ | −4.75 | −3.46 | −1.32 | |
|
| Putative membrane protein | −1.22 | −2.36 | |
|
| GntR-family transcriptional regulator | −1.52 | −1.06 | |
| DIP1669 | −2.72 | −5.16 | 1.82 | |
|
| Putative DNA-binding protein | −1.70 | −1.65 | |
|
| Putative amino acid export carrier protein | −1.88 | −2.28 | 1.14 |
|
| Pyruvate dehydrogenase | −3.20 | −2.66 | 1.98 |
|
| Na:dicarboxylate symporter family | −1.85 | −2.43 | 1.71 |
|
| Putative membrane protein | −1.25 | −2.22 | |
|
| −2.28 | −1.37 | −1.41 | |
| DIP2161 ↑ | −2.44 | −1.20 | −1.20 | |
| DIP2162 ↓ | −1.46 | −1.56 | ||
| DIP2163 | peptide/Ni transport system permease protein | −1.57 | −1.66 | |
|
| peptide/Ni transport system permease protein | −1.52 | −1.35 | |
|
| Conserved hypothetical protein (possible ATP/GTP-binding) | −1.68 | −2.02 | |
|
| Cu-Zn Superoxide dismutase | −1.63 | −1.54 | |
|
| Uncharacterized protein | −1.08 | −1.13 | |
| DIP2330 | −8.71 | −9.45 | ||
| DIP2356 | Conserved membrane protein | −2.56 | −2.70 | |
| Locus | Gene description | Log2 fold Change | ||
|---|---|---|---|---|
| wt | wt:R47H | R47H | ||
| DIP0033 ↓ | 1.13 | |||
| DIP0034 | 1.10 | 1.30 | ||
|
| 1.08 | |||
| DIP0116 | VIT1 family protein; iron transport and | 5.06 | 5.18 | |
| DIP0124 | 2.48 | 4.14 | −3.89 | |
| DIP0125 | TIGR00730 family Rossman fold protein | 1.21 | 1.52 | |
| DIP0173 | 2.51 | 2.14 | ||
| DIP0299 | WhiB family transcriptional regulator | 1.40 | 2.74 | −1.73 |
| DIP0303 | CRP-like cAMP-activated global | 1.46 | 1.42 | −1.14 |
| DIP0357 | Alpha-1,6-glucosidase domain-containing protein | 1.49 | 1.15 | −1.09 |
| DIP0370 ↓ | 3.73 | 3.14 | ||
| DIP0371 | Fumarate reductase/succinate dehydrogenase flavoprotein subunit | 3.72 | 2.87 | |
|
| Succinate dehydrogenase/fumarate reductase iron-sulfur subunit | 3.51 | 2.49 | |
| DIP0373 | Hypothetical protein | 1.78 | 1.49 | |
| 1.35 | 1.85 | |||
| DIP0489 ↓ | 1.65 | 3.05 | −2.92 | |
|
| sdaA | L-serine ammonia-lyase | 1.78 | 2.95 | −3.04 |
|
| 1.29 | 1.48 | ||
| DIP0498 | 2.73 | 2.44 | ||
| DIP0499 | 2.77 | 2.55 | ||
| DIP0500 | 2.88 | 2.35 | ||
| DIP0501 ↑ | MFS transporter | 2.98 | 2.45 | |
| DIP0672 ↓ | Hydrogenase small subunit | 2.96 | 3.00 | |
| DIP0673 | Ni-dependent hydrogenase large subunit | 2.43 | 2.50 | |
| DIP0674 | Ni/Fe-hydrogenase, b-type cytochrome subunit | 1.41 | 1.57 | |
|
| HyaD/HybD family hydrogenase maturation endopeptidase | 1.00 | 1.00 | |
| DIP0739 ↓ | Methylmalonyl-CoA carboxytransferase subunit 5S | 1.55 | 2.97 | −2.40 |
| DIP0740 | Acyl-CoA carboxylase subunit beta | 1.58 | 2.99 | −2.72 |
| DIP0741 | Hypothetical protein | 2.10 | 2.32 | −1.67 |
|
| Biotin/lipoyl-containing protein | 1.89 | 2.19 | −1.75 |
| DIP0956 ↓ | Peptide ABC transporter substrate-binding protein | 1.41 | 2.47 | −1.83 |
| DIP0957 | ABC transporter permease | 1.15 | 2.16 | −1.51 |
| DIP0958 | ABC transporter permease | 1.56 | 1.60 | −1.29 |
|
| dipeptide ABC transporter ATP-binding protein | 1.87 | 2.38 | −1.10 |
| DIP0976 | GtrA family protein | 1.02 | 1.29 | |
| DIP0980 | Amino acid permease | 1.26 | 1.26 | |
| DIP1030 ↓ | Lactate dehydrogenase family protein | 4.18 | 3.53 | |
| DIP1031 | LutB/LldF family L-lactate oxidation iron-sulfur protein | 4.29 | 3.57 | |
|
| (Fe-S)-binding protein | 4.35 | 3.74 | |
| DIP1033 | L-lactate permease | 1.62 | 1.59 | |
|
| Zinc ribbon domain-containing protein YjdM | 1.95 | 1.83 | |
| DIP1124 ↑ | Biotin synthase BioB | 1.96 | 3.00 | −1.50 |
|
| Amino acid ABC transporter ATP-binding protein | 1.70 | 1.86 | |
| DIP1143 | Amino acid ABC transporter permease | 1.94 | 2.47 | |
| DIP1144 ↑ | ABC transporter substrate-binding protein | 2.06 | 2.13 | |
| DIP1149 | C4-dicarboxylate transporter DcuC | 1.47 | 4.86 | −4.36 |
| DIP1198 | Anaerobic C4-dicarboxylate transporter | 1.45 | 3.11 | −1.36 |
|
| Precorrin-3B synthase (dip1231-1233) | 3.49 | 3.12 | |
| DIP1252 | HigA family addiction module antitoxin | 3.81 | 4.71 | |
| DIP1255 | Anaerobic C4-dicarboxylate transporter | 2.01 | 2.68 | |
| DIP1283 | 3.48 | 1.79 | ||
| DIP1284 | TetR/AcrR family transcriptional regulator | 1.50 | ||
| DIP1389 | Dyp-type peroxidase | 1.38 | 1.70 | |
| DIP1390 ↓ | Copper chaperone PCu(A)C | 2.20 | 2.18 | |
|
| Copper resistance CopC family protein | 1.65 | 2.22 | |
| DIP1500 | 1-deoxy-D-xylulose-5-phosphate reductoisomerase | 1.08 | 1.27 | |
| DIP1502 | 23S rRNA (adenine(2503)-C ( | 2.30 | 1.49 | |
| DIP1542 | Alanine/glycine:cation symporter family protein | 1.28 | 1.36 | |
| DIP1555 | Indole-3-glycerol phosphate synthase TrpC | 1.24 | 1.02 | |
| DIP1686 | DUF3052 domain-containing protein | 1.25 | 1.61 | |
| DIP1866 | F | 5.83 | 6.09 | |
| DIP1898 ↓ | Cytochrome ubiquinol oxidase subunit I | 5.00 | 4.03 | |
| DIP1899 | Cytochrome d ubiquinol oxidase subunit II | 4.11 | 3.15 | |
|
| ABC transporter ATP-binding protein/permease | 3.91 | 2.84 | |
|
| Acetate kinase | 1.19 | 1.31 | −1.03 |
| DIP2055 ↑ | 1.16 | 1.49 | −1.53 | |
| Locus | Gene description | Log2 fold change | ||
|---|---|---|---|---|
| wt | wt:R47H | R47H | ||
| DIP0124 | 2.48 | 4.14 | −3.89 | |
| DIP0615 ↓ | 6.01 | −6.02 | ||
| DIP0616 | 5.29 | −5.36 | ||
| DIP0617 | 4.41 | −4.13 | ||
|
| 3.70 | −3.08 | ||
| DIP1923 ↓ | 5.48 | −6.37 | ||
|
| 5.40 | −6.44 | ||
| DIP2261 | −2.49 | 2.45 | ||
| DIP2262 | Oxidoreductase | 4.21 | −4.43 | |
- —U.S. Food and Drug Administrationhttp://dx.doi.org/10.13039/100000038
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDiphtheria, Corynebacterium, and Tetanus · Toxin Mechanisms and Immunotoxins · Botulinum Toxin and Related Neurological Disorders
INTRODUCTION
Corynebacterium diphtheriae is the etiological agent of diphtheria, a disease associated with respiratory and cutaneous infections in humans. The exotoxin, diphtheria toxin (DT), induces cell death by inhibiting protein synthesis and is responsible for the severe symptoms associated with respiratory diphtheria. Although the gene that encodes DT, tox, is found on a prophage locus, the regulation of DT production is dependent upon the availability of iron and diphtheria toxin repressor (DtxR), which is encoded by the dtxR gene that resides on the bacterial chromosome (1–4). DT expression is repressed in iron-replete conditions but derepressed under iron limitation. When intracellular iron levels are sufficient, iron binds to DtxR, resulting in conformational changes in the protein that allow DtxR to interact with DNA at a 19-base pair motif that is typically found near promoter elements (5). C. diphtheriae DtxR has a broad role in regulating numerous genes associated with iron acquisition and metabolism (6).
Iron is critical for essential processes in bacteria, including nucleotide synthesis (7), oxidative stress tolerance (8), and general metabolism (9). Iron in excess, however, may cause oxidative stress through the generation of reactive free radicals by the Haber-Weiss reaction (10). Organisms must maintain appropriate intracellular iron levels to enable cellular processes to continue while minimizing toxicity. Within the human host, available iron is limited by numerous host factors that sequester iron (11). The host proteins transferrin and lactoferrin bind free iron with high affinity in serum and at mucosal interfaces, restricting iron availability. Bacteria produce siderophores, small molecules with high affinity for iron, to compete against transferrin and lactoferrin iron sequestration (12). However, certain siderophores can be bound by the host protein siderocalin (13, 14), which prevents uptake by bacteria and further limits iron availability. Within eukaryotic cells, iron can be associated with proteins, such as those with iron-sulfur clusters, or complexed to protoporphyrin in the form of heme. Most of the heme in the human host is sequestered by hemoglobin, which serves as a critical source of iron for numerous bacterial pathogens (15).
The mechanisms of iron restriction encountered by bacterial pathogens present challenges that must be overcome to successfully colonize the host and cause disease. C. diphtheriae employs numerous mechanisms to acquire heme-iron from the human host, and the genes encoding these heme-iron utilization systems represent a significant portion of the known iron- and DtxR-regulated genes (16–24). C. diphtheriae also produces the iron- and DtxR-regulated siderophore corynebactin (25) and its cognate ABC-type transporter (26). Additionally, C. diphtheriae encodes several other putative iron or metal ABC-type transporters for which substrates have not been identified (18, 27). While many genes of the DtxR and iron regulons have been described (6, 28), a genome-wide transcriptomic analysis comparing both the iron and DtxR regulons has not been done for C. diphtheriae.
In this study, we present a global analysis of differential gene expression in response to iron and DtxR using an iron-limited, semi-defined minimal medium for the growth of a C. diphtheriae wild-type (wt) strain and an isogenic dtxR mutant. In addition to known iron- and DtxR-repressed genes, we found numerous targets that have not been previously reported. By demonstrating DtxR binding at putative promoter regions for new targets, we show that DtxR not only functions to repress gene transcription but can also directly and indirectly induce gene expression. For example, expression of the gene (ftn) encoding the ferritin protein is activated directly through DtxR binding upstream of the ftn promoter region under high-iron conditions. We further demonstrate that the RipA protein, whose gene is repressed by DtxR in elevated iron conditions, functions to repress the expression of several genes encoding metabolic proteins in low-iron environments. The findings from this study expand our knowledge of the iron and DtxR regulon in C. diphtheriae and reveal that DtxR acts through diverse mechanisms to control gene expression.
RESULTS
Identification of differentially expressed genes
We used transcriptome sequencing (RNA sequencing [RNA-seq]) to assess global changes in gene expression in response to iron availability in the C. diphtheriae wt strain 1737. Additionally, we examined differential gene expression between the wt 1737 strain and an isogenic dtxR point mutant, R47H. The mutation in R47H is a single-nucleotide substitution in the 1737 dtxR gene that results in a change from Arg to His at position 47 in the DtxR protein (29). Previous studies with the C7 strain of C. diphtheriae carrying the dtxR R47H mutation showed that the DtxR-R47H protein was produced by the mutant strain but had significantly reduced binding to the DtxR consensus binding site when compared to wt DtxR (29). The R47H mutation has been extensively examined in previous studies using the C7 strain of C. diphtheriae to assess iron- and DtxR-dependent gene expression (16, 19, 29). These earlier studies demonstrated that the R47H amino acid substitution in the DtxR protein resulted in a significant reduction in the ability of DtxR to repress gene expression of several DtxR-repressed genes.
For the transcriptomic analysis, RNA was isolated from C. diphtheriae grown in iron-depleted mPGT medium that was supplemented with either 0.25 µM (low-iron) or 5 µM (high-iron) FeCl_3_. The mPGT medium used throughout this study was slightly altered from the mPGT medium used in previous studies (30) (see Materials and Methods). The R47H mutant showed similar growth to that of the wt strain in the low-iron mPGT medium but exhibited reduced growth under high-iron conditions, which could be restored to wt growth levels in the presence of the cloned dtxR gene (Fig. 1A). To identify genes differentially expressed in response to iron, we compared expression levels in wt C. diphtheriae following growth in low- and high-iron mPGT medium. Genes regulated by DtxR were identified by comparing the wt and the R47H mutant following growth in high-iron medium. Genes with a log_2_ fold change greater than 1 and P < 0.05 were considered significantly regulated.
*(A) Comparison of the growth of wt C. diphtheriae strain 1737 and the 1737 dtxR mutant, R47H, was assessed in low- (0.25 µM FeCl3) and high-iron (5 µM FeCl3) mPGT media; plasmids indicated: vector (pKN2.6Z) and pdtxR (pKN2.6Z carrying the wt dtxR gene). Results show the mean and standard deviation from at least three experiments. **P <0.01; *P <0.05; and ns for not significant by unpaired t test. (B) qPCR analysis of indicated target genes from indicated strain/plasmids grown in either low- (L, 0.25 µM) or high- (H, 5 µM) iron conditions. Relative expression is compared to wt under low-iron conditions for each respective gene. Graphs show means with standard deviations of at least three replicates. Unpaired t tests were performed on ΔCq values for the indicated comparisons: ***P <0.001; **P <0.01; and P <0.05. (C) Coomassie-stained SDS gel of cell lysates (left) and a Western blot (right) performed on cell lysates were probed with antibodies specific to the proteins Ferritin, IutE, and HbpA. Culture supernatants containing DT were also tested. Strains carrying either the vector or pdtxR were grown with either low (L, 0.25 µM) or high (H, 5 µM) iron. A representative experiment is shown.
In the transcriptomic analysis, we identified 139 genes in which transcription was both iron- and DtxR-regulated. The expression of 73 genes was repressed by iron in a DtxR-dependent manner (Table 1), while 66 genes were induced by iron and DtxR (Table 2). To assess the validity of the RNA-seq analysis, we examined the expression of a representative group of genes using qPCR and Western blots (Fig. 1B and C). Expression of all of the genes examined in Fig. 1B and C showed regulation that was consistent with what was observed in the RNA-seq analysis, and the presence of the cloned wt dtxR gene on plasmid p2.6dtxR restored wt gene expression in the R47H mutant (Fig. 1B and C).
DtxR- and iron-repressed genes
Forty-two of the genes in Table 1 whose expression is repressed by iron and DtxR were previously identified and characterized (indicated in normal font). The most notable gene in Table 1 is the tox gene (dip0222), which encodes DT, the primary virulence factor of C. diphtheriae and the source for the current diphtheria vaccine (31). Many of the previously identified DtxR- and iron-repressed genes encode proteins involved in the transport of iron. Several of the iron transport proteins are required for the acquisition and utilization of heme-iron (19, 22, 23), while other transporters are involved in the uptake of non-heme iron, which includes proteins involved in the biosynthesis and transport of the C. diphtheriae siderophore corynebactin: ciuABCDE (dip0582-586) (25, 26). Another important iron-repressed gene is ripA (dip0922), which encodes a regulatory protein that is known in C. glutamicum to repress transcription of several genes that encode iron-containing metabolic enzymes (32, 33).
Among the iron- and DtxR-repressed genes in Table 1 are 31 genes that were not previously shown to be regulated by iron and DtxR (indicated in bold). They are predicted to encode a diverse group of proteins, including factors involved in transport (dip0014-15, dip0611, dip1581, dip0321-323, dip2162-64, and dip0570) and gene regulation; gntR (dip1662) and arsR (dip1879). The putative transporters encoded by dip0611 and dip0570 were also strongly iron regulated in the dtxR mutant background (Table 1; R47H, H:L), suggesting that factors in addition to or other than DtxR are involved in their iron-dependent regulation. The specific function of the gene product of many of these novel genes has not been determined.
DtxR and iron-induced genes
Of the 66 genes whose expression was increased in high-iron conditions in a DtxR-dependent manner (Table 2), only two genes were previously characterized; iutE (dip0173), which encodes the substrate-binding component of a metal-dependent ABC transporter, and the acn gene (dip1283), which encodes the enzyme aconitase (34). The C. diphtheriae acn gene is also regulated by RipA (34). Although the expression of the iutE gene is regulated by DtxR and iron, DtxR does not bind to the sequence upstream of the iutE gene, and no region with significant similarity to the consensus DtxR-binding site was found upstream of iutE. Additionally, RipA was not involved in iutE regulation, and the mechanism as to how iutE expression is regulated by DtxR and iron has not been determined. The genes identified in Table 2 represent a diversity of functions, including several genes encoding transport proteins as well as numerous metabolic enzymes. However, the most strongly iron and DtxR-induced genes were ftn (dip1866), which encodes the iron storage protein ferritin, and dip0116, which is predicted to encode a protein from the VIT1 family that includes proteins involved in iron export and intracellular iron storage (35).
Several genes that were induced in high iron in a DtxR-dependent manner were also strongly repressed in high iron in the dtxR mutant, suggesting that the iron regulation observed for these genes is not directly dependent on DtxR (Table 2; R47H, H:L). The repression observed for these genes in the dtxR mutant under high-iron conditions could be due to other regulators that are responsive to the high intracellular iron levels that would be expected in the dtxR mutant grown in high-iron conditions. One unusual gene among this group of iron-induced genes, in which expression appears to be at least partially independent of DtxR, is piuB (dip0124), which was previously shown to be repressed under high-Mn conditions by the Mn-responsive transcriptional regulator MntR (36). None of the other genes in Table 2 that exhibit iron repression in the dtxR mutant are known to be regulated by MntR or Mn, and the mechanism of regulation for these other genes is not known.
The piuB gene is regulated by iron and Mn
Table 3 shows the RNA-seq results of previously identified Mn-regulated genes in C. diphtheriae, which includes the piuB gene (24). An interesting feature of the expression of these genes is that almost all of them are strongly repressed by iron in the dtxR mutant (Table 3, R47H, H:L). Additionally, expression of these Mn-regulated genes shows significantly stronger repression under high-iron conditions in the dtxR mutant than the expression levels observed in the wt strain (Table 3, wt:R47H, H:H). A possible reason for the strong repression of the MntR-regulated genes in the dtxR mutant under high-iron conditions is likely associated with the high intracellular iron levels that are expected in the dtxR mutant under these growth conditions. These high-iron levels may result in the binding of iron to MntR and the subsequent activation of its DNA-binding function that results in repression of the MntR-regulated genes. In support of this possibility, we previously showed that at high-iron levels, MntR can bind in vitro to its native DNA-binding sites, which will inhibit transcription of MntR-regulated genes (37).
Among the Mn-regulated genes, only piuB is iron regulated in the wt strain (Table 3, wt, H:L), suggesting that DtxR may have a direct role in piuB expression. Although the function of the piuB gene product in C. diphtheriae is not known, the PiuB protein contains a PepSY domain that is known to have a diverse range of activities in bacteria, including siderophore iron utilization and peptidase regulation (38). We previously showed using an electrophoretic mobility shift assay (EMSA) that the Mn regulator, MntR, binds upstream of the start codon for the piuB gene (36). We show in Fig. 2A that DtxR also binds in the upstream region of piuB, suggesting that DtxR is directly involved in the iron-dependent regulation of piuB transcription. The putative binding sites for MntR and DtxR in the region upstream of the piuB gene are shown in Fig. 2B, and their location is based on the consensus binding sites for these transcriptional regulators. Since the transcription start site for the piuB gene could not be identified in a previous study (28), the location of the promoter elements for piuB is not known, and it is unclear how the binding of DtxR and MntR at their respective binding sites impacts gene expression. A control for the EMSA showed that DtxR does not bind in the upstream region for the MntR-regulated gene nrd1 (dip1923), as expected (Fig. 2A). To better understand the regulation of the piuB gene, we measured the expression of piuB using both qPCR and a piuB-LacZ promoter fusion. Although the qPCR results in general support the finding from the RNA-seq data that the cloned wt copy of the dtxR gene is able to complement the dtxR mutation (Fig. 2C), a statistically significant difference in the expression of piuB in high-iron conditions was not established between the mutant carrying the vector and the mutant carrying the cloned dtxR gene due to the variability of the assay. However, the data showed a consistent trend supporting restoration of the wt phenotype for the mutant. In further support of the complementation of the dtxR mutant by the cloned dtxR gene, no significant difference in expression in high-iron conditions was observed between the wt strain carrying only the vector and the dtxR-complemented mutant (Fig. 2C). The LacZ results showed that expression from the piuB promoter was repressed by high levels of Mn (5 μM) as observed previously (36), and the Mn repression of piuB was maintained even in the presence of 5 μM iron (Fig. 2D). In the absence of both Mn and iron, 6.13 units of activity was detected, which was increased two-fold with the addition of 5 μM iron (12.78 units); consistent with the iron-dependent induction of piuB gene expression that was observed in the RNA-seq results. Minimal LacZ activity was observed in the vector control. The findings suggest that the regulation of piuB expression is complex and that it is the only C. diphtheriae gene known to be regulated by both DtxR/iron and MntR/Mn.
*The piuB gene is regulated by iron and Mn. (A) EMSA results with biotinylated DNA target regions detected in the absence (−) or presence (+) of purified DtxR using the promoter region of piuB (dip0124) and negative control gene nrdI (dip1923). (B) The promoter region of piuB with putative MntR and DtxR-binding sites is indicated. (C) qPCR analysis of piuB expression from the indicated strain/plasmids grown in either low- (L, 0.25 µM) or high- (H, 5 µM) iron conditions. R47H indicates the dtxR- strain. Graphs show means with standard deviations of at least three replicates. Unpaired t tests were performed on indicated comparisons: *P <0.05 and ns for not significant. (D) LacZ activity was measured from cultures of C. diphtheriae 1737 WT carrying empty pSPZ vector (vector) or the promoter probe plasmid pSPZ-piuB (ppiuB). Strains were grown in mPGT medium with high (+; 5 µM) or low (−; 0.25 µM) iron and with (+; 5 µM) or without (−) manganese supplementation. P <0.05 by unpaired t test.
Genes that displayed atypical regulation by iron and DtxR
We also identified numerous genes from the RNA-seq analysis whose expression and regulation by iron and DtxR differed from those in Tables 1 and 2. In Table S1, we show the RNA-seq results for 17 genes observed to be only regulated by iron in the wt strain. The most notable gene in this group was dip0281 (cat), which encodes the catalase enzyme, a protein critical for protection from oxidative damage that may occur during growth in high-iron environments. The cat gene was induced in high-iron conditions in the wt strain and also in the dtxR mutant. It is possible that the iron regulation of some of the genes in Table S1, including cat, may be dependent on DtxR, but the difference in expression between wt and the dtxR mutant was below the threshold for statistical significance in the RNA-seq analysis.
In Table S2, RNA-seq results are shown for 83 genes that exhibited differential regulation between wt and the R47H dtxR mutant in high-iron conditions (R47H:wt, H:H). The genes in Table S2 did not exhibit differential regulation by iron levels in the wt strain. Interestingly, many of the genes in Table S2 (65 genes) were also regulated by iron in the dtxR mutant (R47H, H:L), and most of these genes showed elevated expression levels in the high-iron medium in the dtxR mutant, conditions that are expected to have high intracellular iron levels. These results indicate that the elevated expression of these genes in high iron in the dtxR mutant is independent of DtxR and is likely due to other regulatory proteins responding to stress conditions associated with growth in high intracellular iron environments. In support of this possibility, numerous genes encoded by bacteriophage β on the C. diphtheriae chromosome (dip0180-dip0222) are induced under these high-iron conditions (Table S2). The induction of these phage genes may be due to an iron stress response, which was observed for phage genes in other Corynebacterium species (39). Interestingly, the tox gene (dip0222) encoded on phage β is not induced by high iron (as are many of the other phage genes), but is repressed by iron and DtxR (Table 1) (3). It was also noted that numerous genes that showed elevated expression under high-iron conditions only in the dtxR mutant are predicted to express proteins that are associated with stress conditions, many of which have putative roles in DNA damage repair, including: dip0134, dip0135, dip0268, dip0589, dip0604, dip0612, dip0722-0723, dip1026, dip1182, dip01450, dip1861, dip2023, dip2025, dip2132, dip2304, and dip2372.
Only genes with the greatest differential expression are shown in Table S2, and the complete list of genes with this mode of regulation is provided in Table S3.
Identification of new DtxR-binding sites
Our analysis identified numerous genes for which DtxR and iron-dependent regulation were not previously described. We used the MEME suite algorithm (40) to assist in the identification of putative DtxR-binding sites upstream of several of the newly identified DtxR and/or iron-regulated genes (Fig. 3A). We then performed EMSAs to determine whether purified DtxR could bind to DNA fragments carrying the predicted sites indicated in Fig. 3A. All of the upstream regions that had predicted DtxR-binding sites bound DtxR, suggesting that DtxR is directly involved in the transcriptional regulation of these target promoters (Fig. 3A). DtxR binding was verified not only for genes that displayed iron-dependent expression by DtxR but also for genes that showed atypical expression by either iron or DtxR. Interestingly, the upstream region for gene dip0014, which was strongly repressed by iron in a DtxR-dependent manner (Table 1), did not bind DtxR in the EMSA (Fig. 3A). Additionally, dip0611, which is regulated by iron and DtxR, also failed to bind DtxR in its upstream region; however, the expression of dip0611 was also regulated by iron in the dtxR mutant (Table 1), which suggests that factors other than DtxR are likely involved in the regulation of dip0611. The mechanism as to how dip0014 is repressed by iron and DtxR has not been determined, but the findings suggest that DtxR has an indirect role in dip0014 regulation.
DtxR binding to the promoter region of novel iron- and DtxR-regulated genes was assessed using EMSAs. (A) Positions of putative DtxR-binding sites (red boxes) relative to the coding region for the indicated genes. The transcriptional regulation of the various genes examined is categorized as repressed by DtxR and iron, induced by DtxR and iron, or atypical regulation. Corresponding EMSA blots with biotinylated DNA target regions detected in the absence (−) or presence (+) of purified DtxR for each are shown (right). A subset of genes that fall within the gene regulation category was shown to have no binding to DtxR (negative binding inset). (B) Analysis of apo-DtxR and Fe-DtxR binding to selected promoter regions: (−) no DtxR or iron, (+) DtxR added, (++) DtxR and iron added. (C) DtxR-binding consensus sequence generated from verified DtxR-binding sites validated using the MEME suite (40).
Additionally, we tested the binding of apo-DtxR to promoter regions using purified DtxR dialyzed against metal-chelating agents (Fig. 3B). While binding was not observed under any of the conditions in the negative controls, which included two Mn- and MntR-regulated genes (dip1923 and dip0615), our positive controls, dip2330 and dip1510, showed binding to DtxR only with the addition of iron (Fig. 3B). Based on the DtxR-binding sites identified from the current study and from previous reports, a new DtxR consensus sequence was generated (Fig. 3C).
DtxR directly induces C. diphtheriae ferritin expression
The ftn gene (dip1866), which encodes a putative non-heme ferritin, was identified as one of the iron- and DtxR-activated genes (Table 2). In C. diphtheriae, direct activation of gene expression by DtxR is not well understood; however, the presence of three DtxR-binding sites upstream of the promoter region (Fig. 4A) as well as induction by iron suggests a direct role for DtxR in the regulation of ftn transcription. DtxR binding was verified upstream of the ftn gene using a DNA fragment harboring binding site 1 (dip1866-1; Fig. 3A), and a separate DNA fragment carrying both sites 2 and 3 (dip1866-2&3; Fig. 3A). Using biotinylated oligonucleotides for the two downstream sites, we confirmed that DtxR binds independently at both binding sites 2 and 3 (Fig. 4B). The −35, −10, and transcriptional start site of ftn were previously reported (28) and are indicated in Fig. 4A. Interestingly, the −35 is positioned one base downstream from DtxR-binding site 3, and the ftn transcript is predicted to lack a 5′ untranslated region (UTR) (28). The requirement for DtxR in the expression of the ftn gene was further confirmed in Western blots that showed that Ftn was strongly detected in the wt strain only under high-iron conditions and that Ftn was not detected in the R47H mutant (dtxR^-^) regardless of the iron levels, indicating a requirement for DtxR in the expression of ftn (Fig. 1C).
*DtxR directly induces transcription of the ftn gene in high-iron conditions. (A) Sequences of the three DtxR-binding sites found upstream of the ftn gene (dip1866) with the transcriptional start site (Ftn) and promoter elements indicated. (B). EMSAs were performed using annealed oligonucleotides spanning the indicated 19-base putative binding motif and three adjacent bases. A negative control sequence was run alongside binding sites 2 and 3. DtxR was added where indicated (+), and serial twofold dilutions of DtxR were tested with the two binding sites. (C) LacZ activity was measured from cultures of C. diphtheriae 1737 WT and R47H (dtxR-) carrying pSPZ (vector) or the promoter probe plasmids carrying various regions of the ftn promoter region. The line diagram shows promoter fragments relative to identified binding sites. Strains were grown in mPGT medium with the indicated levels of iron supplementation (µM); n.t., not tested. Unpaired t tests were used to compare results for each plasmid grown in 0.25 Fe and 5 Fe: *** P <0.001; **P <0.01; P <0.05; and ns for not significant.
To determine the role of the various DtxR-binding sites in the iron- and DtxR-dependent induction of ftn gene expression, we designed a series of promoter-LacZ fusions that contained varying amounts of the ftn upstream region (Fig. 4C). The fusions were designed such that all sequence upstream of the start codon for the lacZ gene was derived from the C. diphtheriae ftn upstream region with no intervening plasmid sequence; this construction is predicted to mimic the configuration of the native ftn gene to result in a lacZ transcript lacking a 5′ UTR. We measured ß-galactosidase activity following growth in low- and high-iron conditions and observed that all promoter fusion constructs for the wt strain exhibited increased LacZ activity following growth in high iron, consistent with increased transcripts detected in the RNA-seq analysis. The construct with the largest amount of upstream region, p300, harbors all three DtxR-binding sites and resulted in the highest levels of activity in both low- and high-iron conditions. The p200 construct, which removes DtxR-binding site 1, resulted in reduced overall activity following growth in low- and high-iron conditions, but stimulation by iron was still observed. Similar levels of activity were observed from the p100 construct, which possesses the same number of DtxR-binding sites as p200, suggesting that the removal of the region between binding sites 1 and 2 has minimal effect on promoter activity. The p70 construct, which contains only DtxR binding site 1, resulted in the weakest overall β-galactosidase activity, but activity was still increased under high-iron conditions, indicating that the DtxR-binding site closest to the coding region is sufficient for induction by iron. To test whether DtxR is required for the increased expression in response to iron, the p300 and p70 fusions were introduced into the R47H mutant (dtxR^-^). Activity from both p300 and p70 in the R47H mutant was significantly weaker than activity detected in the wt strain, with the p300 fusion showing stronger activity than the p70 fusion (Fig. 4C). Furthermore, induction by iron was abolished for both constructs in the R47H mutant. Together, our results suggest a mechanism in which DtxR binding at the ftn promoter region is required for iron-responsive induction of ftn expression, and the DtxR-binding site adjacent to the −35 element (binding site 3) is sufficient for DtxR-dependent activation.
RipA-regulated genes
The expression of genes regulated by the RipA repressor protein is predicted to be optimally transcribed under high-iron conditions, since expression of the ripA gene is repressed by DtxR in high-iron media. Based on this mode of regulation, it is predicted that RipA-regulated genes should be fully expressed regardless of the iron levels in a ripA mutant. In addition to the RipA-regulated acn gene, which encodes aconitase, it is possible that additional genes in C. diphtheriae whose expression is elevated in high-iron conditions in a DtxR-dependent manner are also regulated by RipA (Table 2). In C. glutamicum, several genes were shown to be RipA repressed, which included the acn gene as well as genes that encode proteins for the production of succinate dehydrogenase and nitrate reductase (33). In C. diphtheriae, expression of genes encoding the succinate dehydrogenase complex (dip0370-dip0372), the nitrate reductase system (dip0497-dip0501) as well as aconitase (acn, dip1283) was shown to be induced by iron and DtxR in the RNA-seq study (Table 2). To determine how the expression of these genes, as well as three other similarly iron- and DtxR-induced genes, is affected in a ripA mutant, we examined the expression of these genes using promoter-lacZ fusions in the wt strain and in a ripA deletion mutant grown under high- and low-iron conditions. The hbpA gene, in which expression is iron and DtxR repressed and is not expected to be regulated by RipA, was used as a control for this study. Prior to initiating these studies, we noted that the growth of the C. diphtheriae ripA deletion mutant was reduced under low-iron conditions compared to wt (0.25 μM FeCl_3_), but had similar growth to the wt strain in high-iron conditions (Fig. 5); similar growth characteristics were previously reported for a C. glutamicum ripA mutant (33).
*Growth of wt C. diphtheriae strain 1737 and the 1737ΔripA mutant was assessed in low (0.25 µM and 0.5 µM) and high-iron (5 µM FeCl3) mPGT media. Results show the mean and standard deviation from at least three experiments. **P <0.001 and ns for not significant by unpaired t test.
The results of the promoter-lacZ analysis showed that the expression of all of the genes that were induced in high iron in the RNA-seq analysis also exhibited significant increases in expression in high iron in the LacZ study (Fig. 6); this finding validates the use of the LacZ reporter for assessing the expression of this group of iron-regulated genes. All of the iron-induced genes also showed significantly increased levels of expression in the ripA deletion mutant in the low-iron medium compared to expression levels in the wt strain in the low-iron medium (Fig. 6). This result is consistent with the expression levels that would be expected in a ripA deletion mutant for a RipA-regulated gene. In the absence of the RipA protein, RipA-repressed genes should show elevated expression levels relative to their expression in the wt strain. As expected, expression of the hbpA control showed similar levels of iron-dependent repression in both wt and the ripA mutant, and no significant difference in expression in low-iron conditions between wt and the ripA mutant was observed for hbpA (Fig. 6). It was further observed that three of the genes (acn, dip0501, and dip1898) showed differences in expression between high- and low-iron conditions in the ripA mutant (Fig. 6). Differences in expression between high- and low-iron conditions in the ripA mutant were also observed previously for the C. glutamicum acn gene (33), which may be associated with the additional regulators that are known to control the expression of acn in C. glutamicum (41). It is also possible that additional regulators impact the expression of RipA-regulated genes in C. diphtheriae, but additional studies will be required to confirm this possibility.
*LacZ activity was measured from cultures of C. diphtheriae 1737 WT and 1737ΔripA carrying the pSPZ plasmid with the promoter region indicated above each graph. Strains were grown in mPGT medium with high (H; 5 µM) or low (L; 0.5 µM) iron. The empty vector control had <0.5 units of activity under all of the conditions examined. ****P <0.0001; ***P <0.001; **P <0.01; P <0.05; and ns for not significant by unpaired t test.
DISCUSSION
In the current study, we observed that expression of the ferritin gene, ftn, was strongly induced by iron and DtxR. Understanding how expression of the ftn gene is regulated by iron is of significant interest since ferritins are known to contribute to the virulence of various bacterial pathogens (42, 43). Because of the unusual DtxR- and iron regulation of the ftn gene, we explored the mechanism of regulation for this gene in greater detail. We found that ftn is directly induced by DtxR in the presence of iron, with DtxR binding directly to three independent sites upstream of the ftn promoter region. Transcriptional regulation of the ftn gene in C. diphtheriae shares some similarities to the regulation of the Mycobacterium tuberculosis ferritin gene, bfrB. The expression of bfrB was shown to be controlled by two separate factors, the Lsr2 repressor, an H-NS-like protein that is involved in both gene expression and chromosome structure, and IdeR, an ortholog of DtxR (44). The bfrB promoter contains two Lsr2-binding sites and four IdeR-binding sites configured in two groups of two adjacent binding sites. One of the Lsr2 sites is located between the two groups of IdeR-binding sites, while the second Lsr2 site is downstream of the bfrB promoter. Like DtxR, IdeR responds to intracellular iron levels. The authors proposed that binding of IdeR displaces Lsr2 near the IdeR-binding sites and alleviates Lsr2 repression to allow for transcription by the bfrB promoter in iron-replete conditions. C. diphtheriae encodes an Lsr2 homolog (dip2266); however, a role for dip2266 in ftn expression has not been examined. It is possible that DtxR binding at site 3, which is adjacent to the ftn −35 promoter element, may facilitate RNA polymerase recognition of the promoter region and result in enhanced transcription of the ftn gene. Additional studies are needed to determine if DtxR interacts with an Lsr2 homolog or another DNA-binding protein at the ftn promoter in C. diphtheriae.
Prior to this report, only the gene encoding aconitase (acn) was shown to be RipA regulated in C. diphtheriae (34). In this study, we found several genes induced by iron in the RNA-seq analysis to be homologous to genes that were RipA regulated in C. glutamicum (33). A transcriptomic analysis of the RipA regulon in C. glutamicum identified 13 genes that were repressed by RipA in a low-iron medium (33). We searched for C. diphtheriae orthologs to these 13 C. glutamicum RipA-regulated genes (33) and assessed their expression profile in the RNA-seq analysis. Among the 13 orthologous genes, the C. diphtheriae sdhCAB (dip0370-0372), acn (aconitase, dip1283), pta (dip2055), and the nar operon genes (dip0501-0497) were found to be iron- and DtxR-induced, whereas the leuCD (dip1127-1128) genes did not show significant iron regulation. The cat gene (dip0281) is known to be iron and DtxR induced and RipA regulated in C. glutamicum (33), but was found to be only iron regulated in C. diphtheriae. However, the upstream region for the cat gene contains a DtxR-binding site consistent with the DtxR binding observed in the EMSA (Fig. 3A), suggesting that DtxR has a role in the regulation of the cat gene. Many of the genes regulated by RipA in C. glutamicum encode iron-containing enzymes involved in basic metabolism, and it is likely that other regulators affect the expression of these genes; for example, the expression of the C. glutamicum acn gene is regulated by at least four transcriptional regulators in addition to RipA (45). Our previous study of C. diphtheriae RipA revealed that expression of the RipA-regulated aconitase gene is complex and likely controlled by factors in addition to RipA (34), which may explain some of the unexpected expression profiles observed for a few of the putative RipA-regulated genes examined in this study (Fig. 6).
While our study focused primarily on genes found to be both iron- and DtxR-regulated, we identified many genes that appear to be differentially regulated only by iron (Table S1) or DtxR (Table S2). Many of the genes regulated only by DtxR encode proteins that are typically associated with DNA repair and are often produced during periods of oxidative stress, as might be expected under high-iron conditions. Many of the proteins in Table S2 were induced in the dtxR mutant under high-iron conditions, an environment that is expected to result in high intracellular iron levels. Most of the iron uptake systems in C. diphtheriae are repressed by DtxR in high-iron conditions in the wt strain (Table 1), and are, therefore, strongly expressed in the dtxR mutant, where these uptake systems would likely transport iron into the cell under conditions where intracellular iron levels are already high. While it is not known whether any of the genes induced in the dtxR mutant in high-iron conditions, such as dip0611, are directly controlled by DtxR, a DtxR-binding site was not found in the dip0611 upstream region (Fig. 3A), which suggests that the intracellular environment in the dtxR mutant may result in the activation of specific regulatory factors that respond to these high-iron stress conditions.
Among the genes that were only differentially regulated by iron in the R47H mutant (Table S2) were numerous genes with putative roles in oxidative stress. Since DtxR mediates intracellular iron homeostasis in C. diphtheriae, the R47H mutant likely experiences some level of oxidative stress due to its inability to repress genes involved in iron uptake. The 1737 R47H mutant showed reduced growth in the high-iron medium, consistent with stress conditions likely due to elevated intracellular iron levels (Fig. 1A).
Together, the results from this study identified new roles and targets for C. diphtheriae DtxR regulation. We have characterized DtxR binding at newly identified iron-regulated promoters, with several of these promoters exhibiting non-canonical iron and DtxR regulation.
MATERIALS AND METHODS
Strains, media, and growth conditions
C. diphtheriae strain 1737 (46) and mutant derivatives (Table S4) were routinely cultured in heart infusion broth with 0.2% (vol/vol) Tween 80 (HIBTW) or on heart infusion agar (HIA; 1.5% agar) at 37°C. Strains were stored at −80°C in heart infusion broth with 20% (vol/vol) glycerol. Modified mPGT medium was prepared as described previously (47) except that increased amounts of Chelex100-treated Casamino acids were added (1.5% instead of 0.5%) to support growth of the R47H strain. Low- and high-iron conditions are defined as mPGT containing 0.5 and 5 µM FeCl_3_, respectively.
Kanamycin was used at 25 µg/mL and 50 µg/mL for C. diphtheriae and E. coli, respectively. Spectinomycin was used at 100 µg/mL for C. diphtheriae and E. coli strains. For experiments with the R47H strain, strains were grown on HIA supplemented with 5µg/mL ethylenediamine-N,N′-diacetic acid (EDDA); instead of HIBTW, strains were grown in mPGT with 0.25 and 5 µM FeCl_3_ used for low- and high-iron conditions, respectively, and stored in mPGT, 0.5 µM FeCl_3_ and 20% (vol/vol) glycerol at −80°C.
Growth assays
Colonies from HIA plates with appropriate additives were used to inoculate HIBTW or mPGT cultures. Cultures were incubated overnight at 37°C with shaking, then diluted with fresh media (1:1). The diluted cultures were grown for an additional 4–6 h for mPGT cultures or 1–2 h for HIBTW cultures. For HIBTW cultures, after this incubation, 500 µL of culture was harvested and the cells were pelleted, resuspended in mPGT supplemented with 1 µM FeCl_3_, and grown for an additional 3–5 h. The cultures were then used to inoculate 1 mL mPGT at an OD_600_ of 0.03 and grown overnight with shaking at 37°C; the growth measured at OD_600_ 16–20 h after inoculation is reported.
DNA cloning and generation of C. diphtheriae mutants
Plasmids used in this study were generated using PCR amplification from wt C. diphtheriae strain 1737 genomic DNA and are listed in Table S4. The NEBuilder HiFi DNA assembly cloning kit (New England Biolabs, Inc.) was used to insert DNA into vectors pK18mobsacB (pKdtxRR47H), pSPZ (lacZ fusions), and pKN2.6z (dtxR complementation clones). Plasmid sequences were verified by DNA sequencing (Macrogen or Plasmidsaurus).
The dtxRR47H point mutation DNA fragment was constructed by Genscript and inserted into the pK18mobsacB plasmid as above. The mutation was then integrated into the C. diphtheriae 1737 genome using the same allelic exchange method described previously (48) with the addition of 5 µg/mL EDDA to HIA plates used for the sucrose selection stage of mutant construction due to the R47H point mutant’s intolerance of high-iron conditions. The point mutation was confirmed by PCR across the gene locus, followed by sequencing (Plasmidsaurus).
RNA extraction
For RNA-seq and qPCR, C. diphtheriae strains were grown on HIA plates with 5 µg/mL EDDA and antibiotics as appropriate, at 37°C or 30°C. Single colonies were used to inoculate mPGT, 0.5 µM FeCl_3,_ and antibiotics as needed, and grown at 37°C with shaking overnight. Overnight cultures were then diluted to a final OD_600_ of 0.1 in mPGT with appropriate iron and antibiotic supplementation and grown to logarithmic phase. Iron was supplemented at 0.25 and 5 µM FeCl_3_. Cultures were harvested in logarithmic growth phase and resuspended in Zymo DNA/RNA Shield following centrifugation. Aliquots of this material were submitted for RNA-seq (SeqCenter), or RNA was isolated in-house for RT-qPCR. Cell lysis was done mechanically using Zymo ZR bashing bead lysis tubes and lysis buffer from the subsequently applied Zymo Quick RNA-miniprep kit. RNA integrity and concentration were assessed using an Agilent RNA 6000 Nano chip and Bioanalyzer.
RNA sequencing
For RNA-seq, RNA extraction was conducted at SeqCenter using the ZymoBIOMICS Quick-RNA Miniprep Kit (Zymo Research, R1055) following the recommendations in the protocol. Final RNA concentrations were quantified using a Qubit, after which samples were treated with DNAse (RNAse free) (Invitrogen). Library preparation used Illumina’s Stranded Total RNA Prep Ligation with Ribo-Zero Plus kit and 10bp unique dual indices. SeqCenter obtained a custom rRNA depletion probe pool using C. diphtheriae strains 1737 (NC_002935) and C7ß (CP003210). Sequencing was done on a NovaSeq X Plus, producing paired-end 150 bp reads. Demultiplexing, quality control, and adapter trimming were performed using bcl-convert. Quality control and adapter trimming were also performed using bcl-convert, read mapping used HISAT2, and read quantification was performed using Subread’s featureCounts functionality. Mapping statistics and raw, quantified counts were provided. Normalization of read counts used edgeR’s Trimmed Mean of M values algorithm, and values were converted to counts per million. Differential expression analysis used edgeR’s glmQLFTest. Differentially expressed genes were defined as those with logFC (base 2) > 1 and P < 0.05; see Supplemental material for all differentially expressed genes.
RNA-seq raw data can be found at the NCBI SRA database: ID- PRJNA1336240.
cDNA synthesis and qPCR
The ProtoScript II First-Strand cDNA synthesis kit (New England Biolabs, Inc.) was used to synthesize cDNA from 100 ng of total RNA. qPCR was performed using the ViiA7 (Thermo Fisher Scientific) with Luna Universal qPCR master mix (New England Biolabs, Inc.). Primers used for qPCR were designed using Primer3 (49) and are listed in Table S5. Data were analyzed using the ΔΔCq method and significance determined using unpaired t tests on ΔCq values. ΔCq values for gyrB were used for normalization. GyrB (dip0005) was not differentially expressed in our analysis.
Electrophoretic mobility shift assays
EMSA experiments using DtxR were performed as described previously (34). Biotinylated DNA probes were generated through PCR amplification of C. diphtheriae strain 1737 genomic DNA with biotinylated primers (Integrated DNA Technologies). For minimal DtxR-binding sites, complementary oligonucleotides were annealed by mixing equimolar amounts and denaturation for 2 m at 98°C in a thermocycler. The temperature was reduced from 98°C to 4°C in 1-m intervals; annealed oligomers were stored at 4°C until use. Primers used to generate DNA probes for the EMSAs are described in Table S6. Biotinylated DNA was detected using the LightShift chemiluminescent EMSA kit (Thermo Scientific).
DtxR was purified using the two-plasmid system for temperature-inducible expression as described previously (34). For testing metal-dependent DNA-binding activity of DtxR, elution fractions containing DtxR were dialyzed sequentially against (i) PBS with 1 mM DTT and 1 mM EDTA for 2 h followed by (ii) PBS with 1 mM DTT and 2 g/L Chelex100 for 2 h, and lastly (iii) PBS with 1 mM DTT, 2 g/L Chelex100, and 15% glycerol for 12 h. For DtxR-binding assays, purified DtxR was incubated at room temperature with target DNA in DtxR-binding buffer (20 mM Na_2_HPO_4_, 50 mM NaCl, 2 mM DTT, 5 mM MgCl_2_, 0.2 µg/µL BSA, 0.05 µg/µL sonicated salmon sperm DNA, and 0.5 mM FeSO_4_ in 10% glycerol at pH 7.0). A reaction in which DtxR was omitted was prepared in parallel. Samples were separated by gel electrophoresis (5% acrylamide with 45 mM Tris-borate [0.5×TB]) and transferred onto nylon in 45 mM Tris-borate, 1 mM EDTA (0.5× TBE).
Beta-galactosidase assays
Overnight cultures of C. diphtheriae with indicated plasmids were grown in mPGT and 0.5 µM FeCl_3_. Cultures were diluted 1:1 with fresh medium and grown for 4–6 h. Cultures were then diluted to a final OD_600_ of 0.03 in fresh medium with indicated metal supplementation. Following overnight growth, cells were pelleted and treated with 10 mg/mL lysozyme in PBS at 37°C for 30 min. Following lysozyme treatment, β-galactosidase (LacZ) activity assays were performed as described by Miller (50). Experiments were performed using cultures initiated from different colonies on different days for a total of three replicates per strain and plasmid.
Generation of antisera against ferritin
The gene encoding ferritin (ftn, dip1866) was cloned into pET24 with an N-terminal Strep II tag to generate the plasmid pET-1866. The plasmid was transformed into E. coli BL21(DE3), and protein was purified using Strep-Tactin XT resin (IBA Lifesciences) following the manufacturer’s recommendations. Elution fractions containing the target protein were dialyzed against PBS and used to immunize guinea pigs for antisera production (Cocalico Biologicals, Inc.).
Cell lysis, electrophoresis, and Western blotting
Cells used for gel electrophoresis and Western blotting were grown in mPGT with FeCl_3_ supplementation as indicated and normalized by OD_600_ at harvest. After centrifugation and harvest of supernatant, cell pellets were lysed using lysozyme and sodium lauroyl sarcosinate as previously described (24). Normalized quantities of cell lysate or supernatant were boiled for 10 min in Laemmli buffer and separated on 4%–15% denaturing precast TGX (Tris-Glycine eXtended) PAGE gels using tris-glycine-SDS running buffer (all reagents from Bio-Rad). Coomassie staining, transfer, and western blot procedures were done as previously described (24). Anti-ferritin primary antibody was used at 1:2,000; IutE primary antibody at 1:5,000; HbpA primary antibody at 1:10,000; and DT (toxin) primary antibody at 1:10,000.
Statistical analyses
GraphPad Prism v10.6.0 was used for data analysis. Specific tests and P values are noted in the figure legends.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Pappenheimer AM Jr. 1977. Diphtheria toxin. Annu Rev Biochem 46:69–94. doi:10.1146/annurev.bi.46.070177.00044120040 · doi ↗ · pubmed ↗
- 2Boyd J, Oza MN, Murphy JR. 1990. Molecular cloning and DNA sequence analysis of a diphtheria tox iron-dependent regulatory element (dtx R) from Corynebacterium diphtheriae. Proc Natl Acad Sci USA 87:5968–5972. doi:10.1073/pnas.87.15.59682116013 PMC 54451 · doi ↗ · pubmed ↗
- 3Schmitt MP, Holmes RK. 1991. Iron-dependent regulation of diphtheria toxin and siderophore expression by the cloned Corynebacterium diphtheriae repressor gene dtx R in C. diphtheriae C 7 strains. Infect Immun 59:1899–1904. doi:10.1128/iai.59.6.1899-1904.19911828057 PMC 257940 · doi ↗ · pubmed ↗
- 4Schmitt MP, Twiddy EM, Holmes RK. 1992. Purification and characterization of the diphtheria toxin repressor. Proc Natl Acad Sci USA 89:7576–7580. doi:10.1073/pnas.89.16.75761502169 PMC 49753 · doi ↗ · pubmed ↗
- 5White A, Ding X, vander Spek JC, Murphy JR, Ringe D. 1998. Structure of the metal-ion-activated diphtheria toxin repressor/tox operator complex. Nature 394:502–506. doi:10.1038/288939697776 · doi ↗ · pubmed ↗
- 6Schmitt MP. 2014. Edited by A. Burkovski. Iron acquisition and iron-dependent gene expression in Corynebacterium diphtheriae, p 95–121. Springer, Dordrecht, Netherlands.
- 7Thelander L, Reichard P. 1979. Reduction of ribonucleotides. Annu Rev Biochem 48:133–158. doi:10.1146/annurev.bi.48.070179.001025382982 · doi ↗ · pubmed ↗
- 8Gregory EM, Yost FJ Jr, Fridovich I. 1973. Superoxide dismutases of Escherichia coli: intracellular localization and functions. J Bacteriol 115:987–991. doi:10.1128/jb.115.3.987-991.19734580575 PMC 246346 · doi ↗ · pubmed ↗