Metabolomics-guided identification of bioactive phytometabolites from South African plants targeting neuroblastoma
Mmei Cheryl Motshudi, Clarissa Marcelle Naidoo, Chikwelu Lawrence Obi, Benson Chucks Iweriebor, Earl Prinsloo, Muhammad Sulaiman Zubair, Nqobile Monate Mkolo

TL;DR
This study identifies bioactive compounds from two South African plants that show potential in fighting neuroblastoma, a deadly childhood cancer.
Contribution
The study introduces an integrated metabolomics and mitochondrial assay approach to discover phytometabolites with neuroblastoma-targeting potential.
Findings
Acorus calamus and Lippia javanica extracts inhibited SH-SY5Y cell viability with IC50 values of 0.2886 and 0.3066 μg/μL, respectively.
Metabolomic profiling revealed distinct phytochemical signatures, including flavonoids and alkaloids, with significant species-level differentiation.
Key metabolites showed favorable ADME properties and blood-brain barrier permeability, suggesting therapeutic potential.
Abstract
Neuroblastoma constitutes a solid tumor in pediatric populations, characterized by a dismal prognosis and a scarcity of effective therapeutic interventions. Medicinal flora from South Africa represents valuable sources of bioactive phytometabolites with potential relevance to neuroblastoma. This study employed an integrated workflow merging untargeted UPLC-MS/MS metabolomics, mitochondrial functional assays, and in silico absorption, distribution, metabolism, and excretion (ADME) prediction to systematically identify bioactive metabolites from Acorus calamus and Lippia javanica with activity against SH-SY5Y neuroblastoma cells. Cytotoxic effects were quantified utilizing the CCK-8 assay, while mitochondrial membrane potential (ΔΨm) was conducted through JC-1 flow cytometry. Untargeted UPLC-MS/MS profiling yielded metabolomic fingerprints, through PCA, PLS-DA, and OPLS-DA. ADME and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
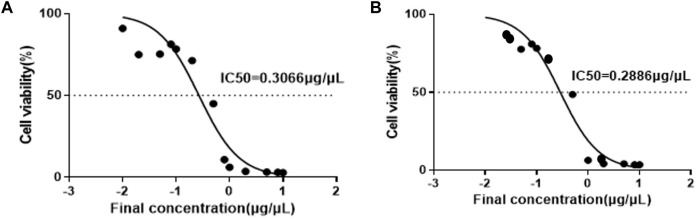 FIGURE 1
FIGURE 1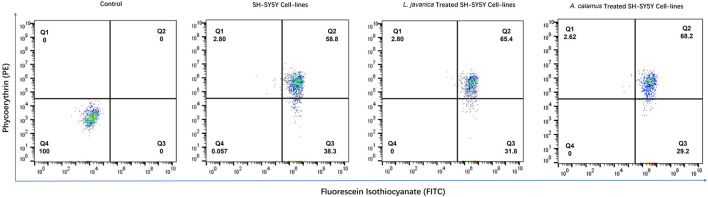 FIGURE 2
FIGURE 2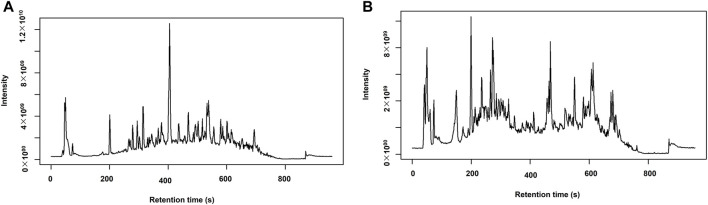 FIGURE 3
FIGURE 3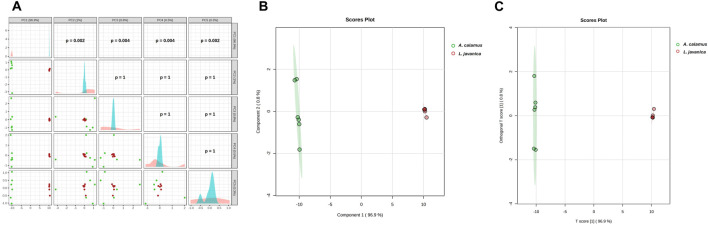 FIGURE 4
FIGURE 4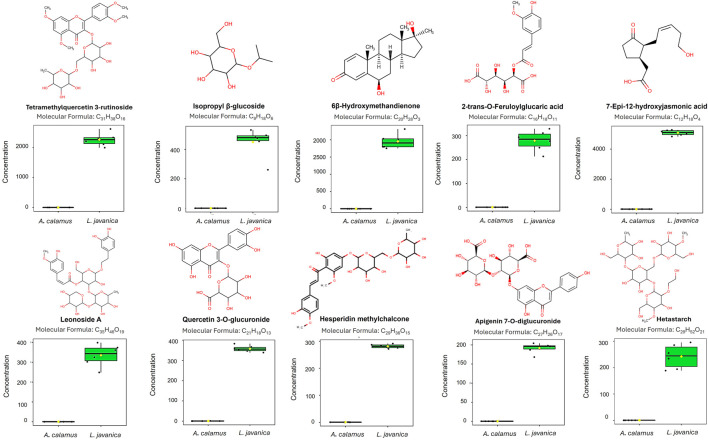 FIGURE 5
FIGURE 5| Compound name | RT (min) | Molecular Weight |
| HMDB_ID | Formula | Log2(FC) | T-Test | FDR Log10 (p-adj) | VIP |
|---|---|---|---|---|---|---|---|---|---|
| ALKALOIDS | |||||||||
| 5-Isothiocyanatoindane | 3,015 | 115,17 | 157,1334 | CSID642931 | C6H13NO | -6,7198 | 4,54E-05 | 4,342703 | 1,830521 |
| 3-Indolehydracrylic acid | 3,336 | 205,0736 | 206,0809 | HMDB0059765 | C11H11NO3 | 5,338881 | 2,95E-05 | 4,52994 | 1,634694 |
| Hydrocotarnine | 4,956 | 221,1048 | 222,1121 | HMDB0033701 | C12H15NO3 | 5,347002 | 4,08E-08 | 7,38938 | 1,6638 |
| Mescaline | 2,991 | 211,26 | 229,1543 | HMDB0254474 | C11H17NO3 | 5,105346 | 4,88E-05 | 4,311856 | 1,602307 |
| Perlolyrine | 4,972 | 264,0895 | 265,0967 | HMDB0030327 | C16H12N2O2 | -5,99531 | 0,001842 | 2,73468 | 1,680894 |
| Prodolic acid | 5,075 | 273,33 | 296,1276 | HMDB0256794 | C16H19NO3 | -6,27265 | 0,001789 | 2,747286 | 1,732869 |
| CARBOHYDRATES | |||||||||
| Isopropyl β-glucoside | 3,5 | 222,24 | 245,1016 | HMDB0032705 | C9H18O6 | 10,83254 | 3,52E-05 | 4,453856 | 2,331036 |
| Muramic acid | 3,206 | 251,1014 | 252,1087 | HMDB0003254 | C9H17NO7 | 8,105161 | 9,48E-05 | 4,023081 | 2,006866 |
| 1-O-Caffeoyl-beta-glucose | 5,172 | 342,0944 | 343,1016 | HMDB0302440 | C15H18O9 | 7,814332 | 1,86E-06 | 5,730844 | 1,977503 |
| Cellulose, microcrystalline | 2,156 | 370,35 | 393,136 | HMDB0032197 | C14H26O11 | 6,787933 | 8,9E-06 | 5,050728 | 1,842234 |
| O-Desmethyltramadol glucuronide | 3,259 | 425,50 | 448,1957 | HMDB0060856 | C21H31NO8 | -6,19442 | 2,12E-05 | 4,673131 | 1,758105 |
| Persicogenin 3'-glucoside | 4,747 | 456,1638 | 479,1532 | HMDB0041398 | C23H26O11 | 8,080171 | 1,16E-06 | 5,934704 | 2,012623 |
| Hetastarch | 4,045 | 736,70 | 759,2936 | HMDB0253113 | C29H52O21 | 8,836521 | 2,09E-05 | 4,679721 | 2,134599 |
| LIPIDS | |||||||||
| 4-Methyl-2-pentenoic acid | 4,046 | 114,14 | 156,1018 | HMDB0031561 | C6H10O2 | 6,668955 | 6,85E-05 | 4,164162 | 1,833963 |
| Undecylenic acid | 4,113 | 184,27 | 226,1798 | HMDB0033724 | C11H20O2 | 7,923542 | 8,84E-08 | 7,053706 | 2,029285 |
| 12-Hydroxydodecanoic acid | 7,277 | 216,32 | 239,1638 | HMDB0002059 | C12H24O3 | -5,60027 | 1,14E-05 | 4,941239 | 1,679658 |
| 11-Dodecenoic acid | 4,664 | 198,30 | 262,1797 | HMDB0032248 | C12H22O2 | 6,21621 | 1,46E-06 | 5,834954 | 1,771416 |
| 9-Pentadecenoic acid | 10,834 | 240,38 | 263,2002 | HMDB0029765 | C15H28O2 | -5,45158 | 0,000253 | 3,59735 | 1,674411 |
| Stearidonic acid | 9,676 | 276,40 | 299,1999 | HMDB0006547 | C18H28O2 | 4,57766 | 1,96E-07 | 6,707939 | 1,521083 |
| MG(18:3/0:0/0:0) | 8,725 | 352,2605 | 353,2678 | HMDB0011570 | C21H36O4 | 5,971525 | 2,8E-07 | 6,553359 | 1,761732 |
| Prostaglandin A2 | 7,707 | 334,40 | 357,2028 | HMDB0002752 | C20H30O4 | 7,579842 | 0,000121 | 3,916245 | 1,940373 |
| Eicosanedioic acid | 10,522 | 342,50 | 365,2678 | HMDB0242141 | C20H38O4 | 6,161779 | 5,82E-06 | 5,23503 | 1,764398 |
| Thromboxane B3 | 4,446 | 368,50 | 386,2529 | HMDB0005099 | C20H32O6 | -6,81951 | 0,000823 | 3,084822 | 1,814574 |
| MG(5-iso PGF2VI/0:0/0:0) | 6,209 | 400,50 | 423,2344 | HMDB0260485 | C21H36O7 | 6,891431 | 1,09E-05 | 4,962081 | 1,86861 |
| LysoPA(18:2/0:0) | 4,868 | 434,50 | 457,2398 | HMDB0007856 | C21H39O7P | 6,062893 | 1,42E-06 | 5,847397 | 1,754746 |
| DG(20:4-2OH/0:0/2:0) | 8,79 | 452,60 | 475,2682 | HMDB0297002 | C25H40O7 | -8,77228 | 1,31E-05 | 4,882486 | 2,094746 |
| DG(2:0/PGD2/0:0) | 7,165 | 468,60 | 491,2629 | HMDB0296907 | C25H40O8 | -5,07408 | 0,000151 | 3,820683 | 1,583894 |
| DG(6 keto-PGF1alpha/2:0/0:0) | 4,095 | 486,60 | 528,3155 | HMDB0296912 | C25H42O9 | 5,338498 | 0,000864 | 3,063593 | 1,605199 |
| LysoPE(22:2/0:0) | 10,093 | 533,3475 | 534,3547 | HMDB0011522 | C27H52NO7P | -6,18444 | 0,00202 | 2,694655 | 1,568301 |
| NITROGEN COMPOUND | |||||||||
| 1,3-Hexadien-3-amine | 1,032 | 97,0895 | 98,09678 | CSID67029750 | C6H11N | 6,805566 | 9,38E-06 | 5,027828 | 1,856891 |
| 2-Aminocyclohexanecarboxylic acid | 1,229 | 143,0945 | 144,1017 | CSID133327 | C7H13NO2 | 5,478209 | 4,02E-06 | 5,395528 | 1,656544 |
| 7-Aminomethyl-7-carbaguanine | 1,725 | 179,0804 | 180,0877 | HMDB0011690 | C7H9N5O | 7,746392 | 4,59E-06 | 5,33773 | 2,019233 |
| 5-Fluoromethylornithine | 4,624 | 164,18 | 187,0863 | HMDB0245493 | C6H13FN2O2 | 5,535211 | 8,43E-08 | 7,073976 | 1,674376 |
| Succinyl proline | 3,047 | 215,20 | 238,0682 | CSID168469 | C9H13NO5 | 7,81002 | 9,47E-08 | 7,02377 | 1,978274 |
| erythro-4-Hydroxyarginine | 3,109 | 190,20 | 229,0704 | HMDB0034326 | C6H14N4O3 | 6,393116 | 3,43E-07 | 6,464989 | 1,81208 |
| Pantothenic acid | 9,416 | 219,23 | 237,1402 | HMDB0000210 | C9H17NO5 | -6,60822 | 0,000512 | 3,290701 | 1,785286 |
| Phenylalanyl-Glycine | 4,839 | 222,24 | 245,0917 | HMDB0304788 | C11H14N2O3 | 4,586087 | 6,19E-06 | 5,208442 | 1,522839 |
| Glutaminylleucine | 3,04 | 259,30 | 277,1906 | HMDB0028801 | C11H21N3O4 | 7,790423 | 1,82E-05 | 4,74028 | 1,971685 |
| Glycyl-Tryptophan | 3,196 | 261,279 | 279,1448 | HMDB0028852 | C13H15N3O3 | -6,59597 | 0,001815 | 2,741203 | 1,589188 |
| N-Acetyllactosamine | 2,923 | 383,35 | 406,1337 | HMDB0001542 | C14H25NO11 | 6,16663 | 0,000286 | 3,544322 | 1,745963 |
| NUCLEOSIDE / NUCLEOTIDE | |||||||||
| 2-Deoxy-ribono-1,4-lactone | 5,407 | 132,0422 | 133,0495 | HMDB0033958 | C5H8O4 | 4,608802 | 2,27E-05 | 4,644565 | 1,515139 |
| 2',3'-Didehydro-2',3'-dideoxycytidine | 4,434 | 209,0811 | 210,0884 | HMDB0245545 | C9H11N3O3 | -8,76604 | 1,28E-05 | 4,892618 | 2,092533 |
| Keto-3-deoxy-manno-octulosonic acid | 3,079 | 238,19 | 261,0601 | HMDB0244292 | C8H14O8 | 6,750766 | 4,57E-06 | 5,340459 | 1,856524 |
| 5-(Hydroxymethyl)cytidine | 2,821 | 273,0955 | 274,1027 | CSID32720391 | C10H15N3O6 | 4,788076 | 2,22E-06 | 5,653688 | 1,616127 |
| 2',3'-Dideoxyadenosine | 6,718 | 235,24 | 277,1405 | HMDB0245544 | C10H13N5O2 | -6,84114 | 4,18E-05 | 4,379233 | 1,84559 |
| 5-Methyldeoxycytidine | 2,036 | 241,24 | 283,1396 | HMDB0002224 | C10H15N3O4 | -8,43195 | 0,00119 | 2,924626 | 1,881698 |
| 3'-C-Ethynylcytidine | 3,174 | 267,24 | 309,1174 | HMDB0252093 | C11H13N3O5 | 8,752045 | 3,19E-08 | 7,496043 | 2,107001 |
| 2'-Deoxy-5-formylcytidine | 1,99 | 255,23 | 319,0993 | CSID10291642 | C10H13N3O5 | 5,632298 | 2,15E-06 | 5,666959 | 1,744307 |
| Dimp | 4,25 | 332,0524 | 333,0597 | HMDB0006555 | C10H13N4O7P | 8,036495 | 2,38E-07 | 6,623169 | 2,006318 |
| 2-Methylguanosine | 4,679 | 297,27 | 339,1407 | HMDB0005862 | C11H15N5O5 | -5,73448 | 0,000162 | 3,789346 | 1,681938 |
| OTHER | |||||||||
| 2,4-Pentadienal | 10,135 | 82,04236 | 83,04964 | HMDB0031597 | C5H6O | 7,510876 | 1,41E-06 | 5,850963 | 1,943115 |
| 5-Methylcytosine | 6,097 | 125,0603 | 126,0676 | HMDB0002894 | C5H7N3O | -8,4848 | 4,81E-06 | 5,317795 | 2,059602 |
| xi-4-Hydroxy-4-methyl-2-cyclohexen-1-one | 3,795 | 126,0681 | 127,0754 | HMDB0033629 | C7H10O2 | 8,258472 | 6,02E-06 | 5,22045 | 2,034825 |
| gamma-Butyrolactone | 3,78 | 86,09 | 128,0706 | HMDB0000549 | C4H6O2 | 7,403507 | 6,82E-06 | 5,166232 | 1,925084 |
| Indol-2-one | 3,251 | 131,0371 | 132,0444 | HMDB0253466 | C8H5NO | 5,862663 | 3,16E-07 | 6,500593 | 1,718906 |
| Methyl 2-thiofuroate | 0,897 | 142,0115 | 143,0188 | HMDB0037762 | C6H6O2S | 4,48824 | 1,48E-05 | 4,829513 | 1,511626 |
| Norcamphoric acid | 3,195 | 158,15 | 200,0915 | CSID207959 | C7H10O4 | 5,611922 | 7,34E-06 | 5,134455 | 1,720562 |
| 4-Guanidino-1-butanol | 0,787 | 131,18 | 173,1396 | CSID4476579 | C5H13N3O | -5,88882 | 0,001006 | 2,997208 | 1,515761 |
| 1-Oxo-1H-2-benzopyran-3-carboxaldehyde | 4,626 | 174,0315 | 175,0388 | HMDB0030577 | C10H6O3 | 6,426311 | 1,56E-05 | 4,807221 | 1,825317 |
| 3-Hydroxysuberic acid | 2,866 | 190,19 | 232,1191 | HMDB0000325 | C8H14O5 | 4,813224 | 4,4E-06 | 5,356159 | 1,568506 |
| 6-(2-Hydroxyethoxy)-6-oxohexanoic acid | 3,717 | 190,19 | 213,0755 | HMDB0061681 | C8H14O5 | 4,69247 | 4,65E-06 | 5,332964 | 1,552665 |
| N-Lactoylphenylalanine | 4,391 | 215,1177 | 238,1069 | HMDB0062175 | C12H15NO4 | 5,606803 | 5,75E-07 | 6,240409 | 1,704328 |
| 2-(2-Furylmethyl)-1-indanol | 3,905 | 214,26 | 237,0907 | CSID40514807 | C14H14O2 | -5,2121 | 0,000218 | 3,660803 | 1,591246 |
| Piperdial | 6,936 | 250,1565 | 251,1638 | HMDB0035798 | C15H22O3 | -5,12368 | 4,52E-06 | 5,345311 | 1,600008 |
| 1-(Ribofuranosyl)indoline | 2,641 | 251,1153 | 252,1226 | CSID67029342 | C13H17NO4 | 4,946661 | 1,05E-05 | 4,977423 | 1,570688 |
| 2-Hydroxyacorenone | 9,271 | 236,35 | 259,1664 | HMDB0030916 | C15H24O2 | -7,68915 | 1,73E-06 | 5,761784 | 1,961428 |
| Risbitin | 5,609 | 222,32 | 264,1954 | HMDB0302980 | C14H22O2 | -5,92921 | 0,001049 | 2,979129 | 1,703258 |
| Linamarin | 1,312 | 247,24 | 270,0944 | HMDB0033699 | C10H17NO6 | -6,29487 | 0,000877 | 3,056864 | 1,759135 |
| 1-Hydroxyacorenone | 7,673 | 250,33 | 273,1456 | HMDB0030917 | C15H22O3 | -7,10849 | 1,82E-06 | 5,739609 | 1,887368 |
| 4-Hydroxycyclohexylcarboxylic acid | 4,619 | 144,17 | 289,1637 | HMDB0001988 | C7H12O3 | 6,014314 | 5,86E-06 | 5,232144 | 1,73907 |
| Pollenin A | 4,254 | 302,042 | 303,0493 | HMDB0303704 | C15H10O7 | 8,76912 | 2,16E-07 | 6,664657 | 2,126655 |
| 9,10-DiHODE | 4,767 | 312,40 | 330,2632 | HMDB0010221 | C18H32O4 | -4,97006 | 0,000693 | 3,159427 | 1,553509 |
| 6β--Hydroxymethandienone | 10,138 | 316,40 | 339,1922 | HMDB0005832 | C20H28O3 | 10,73503 | 2,66E-06 | 5,575855 | 2,317918 |
| Bisoprolol | 5,313 | 325,40 | 348,2162 | HMDB0014750 | C18H31NO4 | 5,110144 | 2,27E-05 | 4,643337 | 1,607036 |
| 6'-Hydroxyenterolactone | 4,893 | 314,30 | 356,1482 | HMDB0041697 | C18H18O5 | 6,599311 | 0,00027 | 3,568419 | 1,804397 |
| Caryoptosidic acid | 2,971 | 392,35 | 415,1204 | HMDB0034249 | C16H24O11 | 6,037738 | 0,000224 | 3,649573 | 1,731817 |
| Tyromycic acid | 12,521 | 452,3278 | 453,3351 | HMDB0035888 | C30H44O3 | -5,71222 | 4,22E-05 | 4,374993 | 1,690313 |
| Fluocinolone | 2,966 | 412,4 | 454,2062 | HMDB0252347 | C21H26F2O6 | -5,03287 | 0,000493 | 3,307314 | 1,561579 |
| Persiconin | 4,285 | 478,1466 | 479,1539 | HMDB0037482 | C23H26O11 | 8,487737 | 1,03E-06 | 5,987367 | 2,064423 |
| Phaseolus epsilon | 3,657 | 526,50 | 544,2378 | HMDB0035039 | C25H34O12 | 7,87868 | 7,04E-05 | 4,152464 | 1,979816 |
| 19-Nor-5-androstenediol | 9,685 | 276,40 | 553,4239 | HMDB0004590 | C18H28O2 | 7,24471 | 8,58E-06 | 5,066603 | 1,926738 |
| 7-Dehydrologanin tetraacetate | 3,488 | 556,1755 | 557,1828 | CSID391585 | C25H32O14 | -6,16628 | 0,003738 | 2,427367 | 1,709022 |
| Procyanidin | 3,823 | 578,1412 | 579,1484 | HMDB0013690 | C30H26O12 | -7,188 | 0,000208 | 3,681349 | 1,883821 |
| Desglucocheirotoxol | 13,088 | 552,70 | 591,2582 | HMDB0033828 | C29H44O10 | 5,752651 | 2,39E-05 | 4,621974 | 1,696989 |
| Neolicuroside | 3,869 | 550,50 | 592,201 | HMDB0040728 | C26H30O13 | 8,370981 | 5,7E-06 | 5,243925 | 2,048135 |
| Marmesin rutinoside | 2,724 | 554,50 | 596,2325 | HMDB0041413 | C26H34O13 | -6,94662 | 0,0022 | 2,657484 | 1,81168 |
| Hesperidin methylchalcone | 4,557 | 624,2032 | 625,2106 | HMDB0253112 | C29H36O15 | 9,090381 | 1,04E-08 | 7,982463 | 2,139523 |
| Glucoliquiritin apioside | 3,01 | 712,60 | 730,2684 | HMDB0041149 | C32H40O18 | -6,08309 | 0,00112 | 2,950734 | 1,673043 |
| Leonoside A | 4,557 | 770,70 | 788,2936 | HMDB0040342 | C35H46O19 | 9,611505 | 1,25E-05 | 4,904459 | 2,19735 |
| PHENOLIC | |||||||||
| Coumarin | 2,503 | 146,0365 | 147,0438 | HMDB0001218 | C9H6O2 | 5,452435 | 0,000132 | 3,879057 | 1,64696 |
| 3 Hydroxycoumarin | 4,771 | 162,0314 | 163,0387 | HMDB0002149 | C9H6O3 | 4,910609 | 2,63E-07 | 6,580547 | 1,573768 |
| 4,8-Dimethyl-7-hydroxycoumarin | 3,343 | 190,19 | 208,0965 | CSID4512230 | C11H10O3 | 5,339346 | 1,67E-05 | 4,77794 | 1,635275 |
| Vanillactic acid | 1,727 | 212,0682 | 213,0755 | HMDB0000913 | C10H12O5 | 7,713922 | 6,68E-07 | 6,175003 | 1,979313 |
| Apigenin | 5,177 | 270,0523 | 271,0596 | HMDB0002124 | C15H10O5 | 5,845909 | 7,92E-05 | 4,101263 | 1,718134 |
| Kaempferol | 4,724 | 286,047 | 287,0542 | HMDB0005801 | C15H10O6 | 7,038508 | 1,48E-06 | 5,830917 | 1,898751 |
| 3-Feruloyl-1,5-quinolactone | 4,631 | 350,30 | 373,091 | HMDB0029289 | C17H18O8 | 7,663669 | 2,15E-05 | 4,668116 | 1,956575 |
| 2-trans-O-Feruloylglucaric acid | 3,606 | 386,0841 | 387,0913 | HMDB0302546 | C16H18O11 | 9,917521 | 9,38E-06 | 5,027846 | 2,226979 |
| Glucosyloxyanthraquinone | 3,844 | 386,40 | 409,0911 | CSID389045 | C20H18O8 | -5,23257 | 0,000103 | 3,987626 | 1,611837 |
| Eriodictin | 4,559 | 434,1204 | 435,1277 | HMDB0037480 | C21H22O10 | 8,417998 | 1,65E-10 | 9,781596 | 2,058779 |
| Catechin 7-glucoside | 3,177 | 452,131 | 453,1383 | HMDB0037949 | C21H24O11 | -5,3561 | 0,003285 | 2,483526 | 1,591291 |
| Quercetin 3-O-glucuronide | 4,241 | 478,0739 | 479,0812 | HMDB0029212 | C21H18O13 | 9,420172 | 6,67E-08 | 7,175578 | 2,186479 |
| Sesaminol glucoside | 7,259 | 532,50 | 555,1482 | HMDB0041209 | C26H28O12 | 4,976722 | 7,32E-05 | 4,135256 | 1,570614 |
| Apigenin 7-O-diglucuronide | 4,027 | 622,1149 | 623,1222 | HMDB0301685 | C27H26O17 | 9,063099 | 3,86E-07 | 6,413527 | 2,131068 |
| Tetramethylquercetin 3-rutinoside | 4,052 | 666,60 | 708,2469 | HMDB0039337 | C31H38O16 | 11,67918 | 1,57E-06 | 5,804899 | 2,418616 |
| POLYKETIDES | |||||||||
| 1(2H)-Pentalenone | 2,502 | 118,042 | 119,0493 | CSID66739022 | C8H6O | 4,673748 | 0,000182 | 3,739442 | 1,516804 |
| Phthalide | 2,503 | 134,13 | 157,0282 | HMDB0032469 | C8H6O2 | 8,291181 | 0,000232 | 3,634062 | 2,029866 |
| Aflatoxin G2 | 6,708 | 330,0731 | 331,0804 | HMDB0030475 | C17H14O7 | 7,294113 | 0,000298 | 3,526371 | 1,900981 |
| TERPENOIDS | |||||||||
| Shinanolone | 4,667 | 192,075 | 193,0823 | HMDB0030580 | C11H12O3 | -5,65953 | 0,005135 | 2,289432 | 1,589457 |
| 4,7-Megastigmadien-9-ol | 8,59 | 194,1668 | 195,1741 | HMDB0038731 | C13H22O | -5,78981 | 3,46E-05 | 4,460445 | 1,700406 |
| Vomifoliol | 3,517 | 224,1409 | 225,1482 | HMDB0303570 | C13H20O3 | 6,685312 | 3,63E-06 | 5,439498 | 1,850491 |
| 7-Epi-12-hydroxyjasmonic acid | 3,942 | 226,12 | 227,1273 | HMDB0303749 | C12H18O4 | 9,757035 | 2,83E-08 | 7,548945 | 2,218914 |
| 8-Hydroxygeraniol 8-O-glucoside | 4,457 | 332.39 | 374,2165 | HMDB0035025 | C16H28O7 | 5,392225 | 0,000565 | 3,248129 | 1,621572 |
| 3beta-Hydroxy-5-cholestenal | 9,848 | 400,60 | 423,3249 | HMDB0060131 | C27H44O2 | -5,19836 | 1,33E-05 | 4,874901 | 1,62304 |
| 7a,12a-Dihydroxy-cholestene-3-one | 10,46 | 416,60 | 439,3199 | HMDB0002197 | C27H44O3 | -7,53264 | 7,79E-06 | 5,108677 | 1,956674 |
| Limonoate a-ring-lactone | 6,644 | 488,2015 | 489,2088 | HMDB0302537 | C26H32O9 | -5,00973 | 0,001182 | 2,927482 | 1,561349 |
| Cucumerin A | 2,79 | 552,50 | 575,1571 | HMDB0301967 | C29H28O11 | 6,765172 | 0,000685 | 3,164562 | 1,819729 |
| Cholic acid glucuronide | 12,017 | 584,70 | 623,2843 | HMDB0002577 | C30H48O11 | 4,8136 | 9,02E-06 | 5,04476 | 1,549668 |
| Linalool (8-hydroxydihydro-) | 4,627 | 650,70 | 692,2884 | HMDB0304700 | C32H42O14 | 6,441235 | 4,01E-05 | 4,396548 | 1,794936 |
| Parameters | Isopropyl β-glucoside | Hetastarch | 6β--hydroxymethandienone | Hesperidin methylchalcone | Leonoside A | 2-trans-O-Feruloylglucaric acid | Quercetin 3-O-glucuronide | Apigenin 7-O-diglucuronide | Tetramethylquercetin 3-rutinoside | 7-Epi-12-hydroxyjasmonic acid |
|---|---|---|---|---|---|---|---|---|---|---|
| Physicochemical Properties | ||||||||||
| Formula | C9H18O6 | C29H52O21 | C20H28O3 | C29H36O15 | C35H46O19 | C16H18O11 | C21H18O13 | C27H26O17 | C31H38O16 | C12H18O4 |
| Molecular weight | 222.24 g/mol | 736.71 g/mol | 316.43 g/mol | 624.59 g/mol | 770.73 g/mol | 386.31 g/mol | 478.36 g/mol | 622.49 g/mol | 666.62 g/mol | 226.27 g/mol |
| No. heavy atoms | 15 | 50 | 23 | 44 | 54 | 27 | 34 | 44 | 47 | 16 |
| No. arom. heavy atoms | 0 | 0 | 0 | 12 | 12 | 10 | 16 | 16 | 12 | 0 |
| Fraction Csp3 | 1.00 | 1.00 | 0.60 | 0.48 | 0.57 | 0.44 | 0.24 | 0.37 | 0.48 | 0.50 |
| No. of rotatable bonds | 0 | 15 | 6 | 11 | 13 | 4 | 4 | 7 | 13 | 9 |
| No. H-bond acceptors | 6 | 21 | 3 | 15 | 19 | 11 | 13 | 17 | 16 | 4 |
| No. H-bond donors | 0 | 11 | 0 | 9 | 11 | 6 | 8 | 9 | 8 | 0 |
| Molar Refractivity | 49.89 | 156.69 | 93.83 | 148.42 | 179.64 | 87.17 | 110.77 | 139.71 | 158.16 | 61.42 |
| TPSA | 55.38 Å2 | 314.83 Å2 | 43.37 Å2 | 245.29 Å2 | 304.21 Å2 | 179.28 Å2 | 227.58 Å2 | 283.34 Å2 | 251.36 Å2 | 52.60 Å2 |
| Lipophilicity | ||||||||||
| Log Po/w (iLOGP) | 3.13 | 2.21 | 4.00 | 2.15 | 2.28 | 0.90 | 1.13 | 0.28 | 3.47 | 3.17 |
| Log Po/w (XLOGP3) | 1.84 | −7.70 | 5.49 | −0.50 | −1.60 | −0.99 | 0.61 | −0.24 | 0.07 | 2.22 |
| Log Po/w (WLOGP) | 2.05 | −7.35 | 4.39 | −1.12 | −2.27 | −1.61 | −0.45 | −1.94 | −0.55 | 2.01 |
| Log Po/w (MLOGP) | 1.50 | −7.42 | 3.48 | −2.37 | −3.75 | −2.23 | −2.60 | −3.68 | −1.99 | 1.94 |
| Log Po/w (SILICOS-IT) | 1.09 | −6.09 | 4.87 | −1.14 | −2.61 | −1.13 | −1.04 | −2.66 | −0.61 | 2.53 |
| Consensus Log Po/w | 1.92 | −5.27 | 4.45 | −0.60 | −1.59 | −1.01 | −0.47 | −1.65 | 0.08 | 2.37 |
| Water solubility | ||||||||||
| Log S (ESOL) | −2.38 | 1.43 | −4.86 | −2.87 | −2.92 | −1.62 | −3.27 | −3.36 | −3.35 | −2.05 |
| Solubility | 9.33e-01 mg/mL; 4.20e-03 mol/L | 2.00e+04 mg/mL; 2.71e+01 mol/L | 4.32e-03 mg/mL; 1.37e-05 mol/L | 8.36e-01 mg/mL; 1.34e-03 mol/L | 9.33e-01 mg/mL; 1.21e-03 mol/L | 9.24e+00 mg/mL; 2.39e-02 mol/L | 2.54e-01 mg/mL; 5.32e-04 mol/L | 2.75e-01 mg/mL; 4.41e-04 mol/L | 2.99e-01 mg/mL; 4.49e-04 mol/L | 2.03e+00 mg/mL; 8.96e-03 mol/L |
| Class | Soluble | Highly soluble | Moderately soluble | Soluble | Soluble | Very soluble | Soluble | Soluble | Soluble | Soluble |
| Log S (Ali) | −2.62 | 1.83 | −6.16 | −4.18 | −4.28 | −2.29 | −4.96 | −5.25 | −4.90 | −2.96 |
| Solubility | 5.29e-01 mg/mL; 2.38e-03 mol/L | 4.95e+04 mg/mL; 6.72e+01 mol/L | 2.19e-04 mg/mL; 6.94e-07 mol/L | 4.09e-02 mg/mL; 6.55e-05 mol/L | 4.05e-02 mg/mL; 5.26e-05 mol/L | 1.99e+00 mg/mL; 5.14e-03 mol/L | 5.20e-03 mg/mL; 1.09e-05 mol/L | 3.48e-03 mg/mL; 5.59e-06 mol/L | 8.35e-03 mg/mL; 1.25e-05 mol/L | 2.48e-01 mg/mL; 1.10e-03 mol/L |
| Class | Soluble | Highly soluble | Poorly soluble | Moderately soluble | Moderately soluble | Soluble | Moderately soluble | Moderately soluble | Moderately soluble | Soluble |
| Log S (SILICOS-IT) | −1.45 | 5.00 | −3.83 | −0.22 | 1.04 | −0.34 | −1.04 | 0.05 | −0.81 | −2.55 |
| Solubility | 7.89e+00 mg/mL; 3.55e-02 mol/L | 7.45e+07 mg/mL; 1.01e+05 mol/L | 4.65e-02 mg/mL; 1.47e-04 mol/L | 3.77e+02 mg/mL; 6.04e-01 mol/L | 8.55e+03 mg/mL; 1.11e+01 mol/L | 1.75e+02 mg/mL; 4.52e-01 mol/L | 4.36e+01 mg/mL; 9.12e-02 mol/L | 6.95e+02 mg/mL; 1.12e+00 mol/L | 1.03e+02 mg/mL; 1.55e-01 mol/L | 6.35e-01 mg/mL; 2.81e-03 mol/L |
| Class | Soluble | Soluble | Soluble | Soluble | Soluble | Soluble | Soluble | Soluble | Soluble | Soluble |
| Pharmacokinetics | ||||||||||
| GI absorption | High | Low | High | Low | Low | Low | Low | Low | Low | High |
| BBB permeant | Yes | No | Yes | No | No | No | No | No | No | Yes |
| P-gp substrate | No | Yes | No | Yes | No | No | Yes | Yes | Yes | No |
| CYP1A2 inhibitor | No | No | No | No | No | No | No | No | No | No |
| CYP2C19 inhibitor | No | No | Yes | No | No | No | No | No | No | No |
| CYP2C9 inhibitor | Yes | No | Yes | No | No | No | No | No | No | No |
| CYP2D6 inhibitor | No | No | Yes | No | No | No | No | No | No | No |
| CYP3A4 inhibitor | No | No | Yes | No | No | No | No | No | No | No |
| Log Kp (skin permeation) | −6.35 cm/s | −16.26 cm/s | −4.33 cm/s | −10.46 cm/s | −12.14 cm/s | −9.36 cm/s | −8.78 cm/s | −10.27 cm/s | −10.32 cm/s | −6.10 cm/s |
| Drug likeness | ||||||||||
| Lipinski | Yes; 0 violation | No; 3 violations: MW >500, NorO >10, NHorOH >5 | Yes; 0 violation | No; 3 violations: MW >500, NorO >10, NHorOH >5 | No; 3 violations: MW >500, NorO >10, NHorOH >5 | No; 2 violations: NorO >10, NHorOH >5 | No; 2 violations: NorO >10, NHorOH >5 | No; 3 violations: MW >500, NorO >10, NHorOH >5 | No; 3 violations: MW >500, NorO >10, NHorOH >5 | Yes; 0 violation |
| Ghose | Yes | No; 4 violations: MW >480, WLOGP <−0.4, MR >130, #atoms >70 | Yes | No; 4 violations: MW >480, WLOGP <−0.4, MR >130, #atoms >70 | No; 4 violations: MW >480, WLOGP <−0.4, MR >130, #atoms >70 | No; 1 violation: WLOGP <−0.4 | No; 1 violation: WLOGP<−0.4 | No; 3 violations: MW >480, WLOGP<−0.4, MR >130 | No; 4 violations: MW >480, WLOGP <−0.4, MR >130, #atoms >70 | Yes |
| Veber | Yes | No; 2 violations: Rotors >10, TPSA >140 | Yes | No; 2 violations: Rotors >10, TPSA >140 | No; 2 violations: Rotors >10, TPSA >140 | No; 1 violation: TPSA >140 | No; 1 violation: TPSA >140 | No; 1 violation: TPSA >140 | No; 2 violations: Rotors >10, TPSA >140 | Yes |
| Egan | Yes | No; 1 violation: TPSA >131.6 | Yes | No; 1 violation: TPSA >131.6 | No; 1 violation: TPSA >131.6 | No; 1 violation: TPSA >131.6 | No; 1 violation: TPSA >131.6 | No; 1 violation: TPSA >131.6 | No; 1 violation: TPSA >131.6 | Yes |
| Muegge | Yes | No; 5 violations: MW >600, XLOGP3 <−2, TPSA >150, H-acc >10, H-don >5 | No; 1 violation: XLOGP3 >5 | No; 4 violations: MW >600, TPSA >150, H-acc >10, H-don >5 | No; 4 violations: MW >600, TPSA >150, H-acc >10, H-don >5 | No; 3 violations: TPSA >150, H-acc >10, H-don >5 | No; 3 violations: TPSA >150, H-acc >10, H-don >5 | No; 4 violations: MW >600, TPSA >150, H-acc >10, H-don >5 | No; 4 violations: MW >600, TPSA >150, H-acc >10, H-don >5 | Yes |
| Bioavailability score | 0.55 | 0.17 | 0.55 | 0.17 | 0.17 | 0.17 | 0.11 | 0.11 | 0.17 | 0.55 |
| Medicinal Chemistry | ||||||||||
| PAINS | 0 alert | 0 alert | 0 alert | 1 alert: catechol_A | 1 alert: catechol_A | 0 alert | 1 alert: catechol_A | 0 alert | 1 alert: catechol_A | 0 alert |
| Brenk | 1 alert: peroxide | 0 alert | 1 alert: isolated_alkene | 2 alerts: catechol, michael_acceptor_1 | 2 alerts: catechol, michael_acceptor_1 | 1 alert: cumarine | 1 alert: catechol | 0 alert | 3 alerts: catechol, michael_acceptor_1, more_than_2_esters | 2 alerts: mchael_acceptor_1, more_than_2_esters |
| Leadlikeness | No; 1 violation: MW 250 | No; 2 violations: MW >350, Rotors >7 | No; 1 violation: XLOGP3 >3.5 | No; 2 violations: MW >350, Rotors >7 | No; 2 violations: MW >350, Rotors >7 | No; 1 violation: MW >350 | No; 1 violation: MW >350 | No; 1 violation: MW >350 | No; 2 violations: MW >350, Rotors >7 | No; 2 violations: MW < 250, Rotors >7 |
| Synthetic accessibility | 4.34 | 8.06 | 4.72 | 6.37 | 7.45 | 5.05 | 5.26 | 6.26 | 6.57 | 2.31 |
- —National Research Foundation10.13039/501100001321
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMedicinal Plants and Neuroprotection · Bioactive natural compounds · Phytochemicals and Medicinal Plants
Impact statement
This manuscript serves to investigate the medicinal flora for natural bioactive compounds as probable sources of therapeutic anticancer agents targeting SH-SY5Y neuroblastoma cells. Utilizing a multifaceted methodology that incorporates cytotoxicity assays, untargeted metabolomics, and in silico Absorption, Distribution, Metabolism, and Excretion modeling, we have identified phytometabolites exhibiting promising anticancer efficacy and have also predicted their permeability across the blood–brain barrier. The findings will offer different perspectives and insights into the metabolomic profiling and therapeutic potential of the medicinal plants, thereby enhancing the pharmacological comprehension of phytochemical agents in pediatric oncology.
This manuscript also highlights the current strategic plans for neuroblastoma treatment and approaches that could be applied in the future to improve the quality of life of patients with neuroblastoma, as well as to provide early diagnostic measures to reduce mortality rates of this fatal cancer. This information could be of great use for both clinical and scientific research.
Introduction
Neuroblastoma (NB) is a primary cancer detected in infants; it is a predominant solid tumor commonly found in the extracranial area in children [1]. Neuroblastoma accounts for approximately 10–15% of childhood malignancies, and the primary tumour generally stems from the sympathetic chain, usually in the abdomen or in the adrenal gland [2]. Patients with neuroblastoma tend to exhibit a wide range of biological, clinical, accompanied by prognostic heterogeneity, and a fundamental prognostic trait pertaining to neuroblastoma patients is the location of the primary tumour, which can originate in the adrenal gland, abdominal/retroperitoneal area, neck, thorax, pelvis, or, less frequently, in other sites [3, 4]. Due to the heterogeneous biological nature of neuroblastoma, its prognosis, along with clinical course, differs between spontaneous regression to high-risk cases, with several neuroblastoma tumours exhibit poorly responsive to intensive multimodal therapy [5, 6]. Presently treatment strategies involve aggressive amalgamation of radiotherapy, induction chemotherapy, elevated-dose chemotherapy with autologous stem-cell rescue, surgical removal of tumors, and applications of post-consolidation methods such as immunotherapy, as well as differentiation therapy [7]. The 5-year overall survival rate for high-risk neuroblastoma individuals continues to hover around 50–60% despite these integrated multimodal approaches, and late effects are usually imminent and frequently experienced by long-term survivors [8]. Regardless of advancement in multimodal therapy, outcomes for high-risk neuroblastoma persist to be suboptimal, highlighting the necessity for alternative and complementary therapeutic strategies.
The SH-SY5Y human neuroblastoma cell line comprises an N-type catecholaminergic subclone of neuroblastoma commonly used in research areas pertaining to neuroblastoma and neurobiology [9]. SH-SY5Y cell lines convey crucial elements of human catecholaminergic systems, which encompass components such as dopamine-β-hydroxylase and tyrosine hydroxylase [10]. Induced SHSY5Y cells can be differentiated into phenotypes that resemble neurons, therefore rendering them an adaptable in vitro model [11]. However, limitations such as inadequate maturation levels of neurons and restricted functionality of network systems, irrespective of the differentiation, emphasize the necessity of cautious interpretation of pharmacological responses [12]. Changes in mitochondrial membrane potential (ΔΨm) and cellular redox balance can play context-dependent roles in cancer biology, acting as regulators of cell survival and cell death conditional on cellular state and therapeutic pressure [13]. Although moderate stabilization of mitochondrial function and redox homeostasis might permit tumour cell adaptation, excessive or dysregulated perturbation of these pathways can encourage cytotoxic or cytostatic outcomes via mitochondrial dysfunction [14, 15]. Accordingly, modulation of ΔΨm and redox-related processes has been explored as a functional vulnerability in cancer cells, also neuroblastoma, instead as a unidirectional pro-survival mechanism [16, 17].
Approximately 65% of the world’s population, along with an estimated 80% of the population in Africa as well as Asia, depends on traditional herbal remedies to cure and furthermore prevent infectious ailments and chronic diseases [18, 19]. Ethnobotanical knowledge from southern Africa has showcased many species of aromatic origin known for their anticancer, anti-inflammatory, and anti-infective properties [20]. South African based medicinal plants are rich sources of structurally diverse phytometabolites with documented biological activities, and systematic investigation of these resources might permit the discovery of candidate anti-cancer chemotypes or complementary approaches to present therapies [21–25]. Lippia javanica (Burm.f.) is a multi-branched woody shrub, that is commonly known as fever tea [9, 26]. L. javanica (Verbenaceae family) is an aromatic medicinal plant indigenous to eastern and southern Africa that has been broadly explored for its phytochemical composition and biological activities [26–30]. Previous studies have recognized phenolic, terpenoids, and flavonoids constituents, with apigenin- and luteolin-associated derivatives, which in vitro exhibit antioxidant and cytotoxic activities, supporting its importance for further assessment within a metabolomics-directed anticancer framework [31–36]. Acorus calamus Linn., (Aceraceae family), is classified as a perennial, semi-aquatic herb, that is commonly known as sweet flag {25]. A. calamus is an aromatic medicinal plant that has been extensively studied for its phytochemical diversity and biological activities, and it is frequently utilized in traditional Chinese and Indian medicine [37–41]. Notably, this plant comprises phenylpropanoid constituents for instance α- and β-asarone, for which extracts and isolated compounds have established anticancer activity in preclinical cancer models, also permitting its inclusion in metabolomics-directed anticancer investigations [42, 43]. Untargeted LC-MS metabolomics is commonly utilized to map systematic methods for enhancing quality control, distinguishing chemotypes, associating chemical properties with bioactivity of samples, and prioritizing key metabolites for further pharmacology analysis, alongside in silico ADME assessment through tools like SwissADME and the BOILED-Egg model [44–47].
Building on the above insight, while the phytochemistry and bioactivity of L. javanica and A. calamus have been previously reported in various biological contexts, their assessment within an integrated framework relating metabolomic composition to mitochondrial functional outcomes and pharmacokinetic possibility remains limited. In this study, we integrated untargeted UPLC-MS/MS metabolomics, mitochondrial membrane potential evaluation in SH-SY5Y neuroblastoma cells, and in silico ADME analysis to select metabolites with possible neuroblastoma importance.
Materials and methods
Plant collection and identification
Specimens of L. javanica and A. calamus were collected from Hartbeespoort in the North-West Province of South Africa (25.7236° S, 27.9653° E) in February 2023. Organ-matched sampling was applied to follow metabolomics reporting recommendations that highlight control of tissue-specific variability in comparative chemical analyses [48]. Species identification of both plants was authenticated by taxonomists at the National Herbarium, where voucher specimens were deposited under accession numbers NR 904 (L. javanica) and NR 905 (A. calamus). Plant material collection was completed in accordance with national regulatory requirements, under permit CF6-0234 released by the Department of Agriculture and Rural Development-Nature Conservation, South Africa (Permit Holder: Prof. Nqobile Monate Mkolo).
Anti-cancer assay analysis
Sample preparation
The L. javanica and A. calamus samples were pulverized into a powder using a mortar and pestle under liquid nitrogen. A precise weight of 0.5 g from each powdered sample was combined into a single 50 mL centrifuge tube. A total of 10 mL of buffer was added to the tube followed by extraction at a 4 °C. The extractant was filtered through a 0.45 μm filter, and the clearer supernatants were transferred into a new centrifuge tube. Crude extracts and intermediate preparations were stored at −20 °C in amber, light-protected containers to reduce degradation. Extracts were permitted to equilibrate to room temperature and were used within 1 week of preparation to allow stability and duplicability.
SH-SY5Y cell preparation
Human neuroblastoma SH-SY5Y cells (Cellonex ™, Johannesburg South Africa) were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% (v/v) heat-inactivated fetal bovine serum, 1% penicillin-streptomycin obtained from Gibco (New York, NY, United States), and maintained at 37 °C in a humidified incubator with 5% CO_2_ [49]. Cells were washed by utilizing a 1 mL pipette to aspirate and discard the old medium. Gradually, 2 mL of Phosphate Buffered Saline (PBS) from Sigma-Aldrich (St. Louis, MO, United States), was poured along the edge of the T25 flask, and this flask was gently swirled to wash away any remaining medium. Cell digestion was conducted by gradually adding 1 mL of 0.25% trypsin digest solution (Sigma-Aldrich, St. Louis, MO, United States) into the T25 flask, ensuring it covered all the cells. The flask was then placed in a 37 °C CO_2_ incubator for 2 min. Cells were examined, using an inverted microscope, for separation and formation of a spherical shape. To stop digestion, 4 mL of cell growth medium was added to the cells. Passage was done at a ratio of 1:4, where the cells were carefully pipetted for dispensation, and a single-cell suspension was confirmed under the microscope.
CCK-8 assay
The effect of chosen plant tissue (final concentration of 0.01–10 μg/mL) on SH-SY5Y cell viability was assessed using the CCK-8 assay according to the protocol [50]. SH-SY5Y cells were collected from an approximately 80% confluent T-25 flask using the standard trypsinization method. Afterward, 2 × 10^4^ cells were seeded in a 96-well plate and incubated for 24 h at 37 °C. Then, the media was removed, rinsed with PBS, and the cells were treated with 100 μL of clear supernatants from the sample, while the cell medium served as the negative control. Following 24 h, 10 μL of CCK-8 was added to every well and allowed to incubate for 2 h. A microplate reader (Infinite M200, Tecan, Switzerland) scanned the 96-well plate at a wavelength of 450 nm. All CCK-8 experiments were performed utilizing three independent biological replicates, with each condition examined in technical triplicate. The subsequent formula was applied to determine the percentage of cell viability:
Wherein Asample, Ab, and Ac are the absorbance/Optical density of cells exposed to test reagents, absorbance/Optical density of blank, and negative control, respectively.
Mitochondrial membrane potential detection
The mitochondrial membrane potential (ΔΨm) of SHSY-5Y cells against L. javanica and A. calamus was determined by adopting a procedure of Sakamuru and co-workers [51]. SH-SY5Y cells were seeded in a 96-well plate (2 × 10^4^ cells per well) according to experimental groups, which included the blank control group (non-treated SH-SYSY cell line), negative control of non-treated SH-SYSY cell line for gating, and the treated SH-SYSY cell line group (L. javanica and A. calamus at 1 µg/uL). After 24 h of treatment, the cells were collected using trypsin for adherent cells to avoid over-digestion and to prevent membrane potential damage. A JC-1 working solution was prepared using a diluted JC-1 stock solution with pre-warmed (37 °C) medium, which was added to the final concentration. The JC-1 working solution was then added to the cells and incubated at 37 °C in the dark for 15 min. JC-1 solution was removed, and cells were washed 3 times with pre-cooled PBS to remove unbound dye. The cells were collected using trypsin, then centrifuged and resuspended into PBS. The set flow cytometry channels were green fluorescence, which was the FL1 channel (Ex 488 nm, Em 530/30 nm), and red fluorescence, which was the FL2 channel (Ex 488 nm, Em 585/42 nm). The red fluorescence (normal mitochondria) and green fluorescence (decreased membrane potential) were then merged for analysis. Mitochondrial membrane potential (ΔΨm) assessments were conducted utilizing three independent biological replicates, with technical triplicates for each experimental condition.
Metabolomic characterization
Chemicals and instrumentation
Multiple chemicals and reagents, such as acetonitrile (Merck, Rahway, NJ, United States), methanol (Merck, Rahway, NJ, United States), DL-o-Chlorophenylalanine (Merck, Rahway, NJ, United States), and formic acid (Merck, Rahway, NJ, United States) were utilized. Additionally, instrumentation, namely ACQUITY UPLC HSS T3 (100 × 2.1 mm × 1.8 μm), Ultimate 3000LC combined with Q Exactive MS (Thermo, Waltham, MA, United States), and Temp functional Centrifugation (Eppendorf, Enfield, CT, United States) were utilized.
Sample preparation
The initial stages of the sample preparation comprised a freeze-drying process (lyophilization) of A. calamus and L. javanica leaf samples. The respective samples were subsequently dried and ground into powder; the resultant powder for each sample was transferred into a 5 mL homogenizing tube. A MM 400 mixer with four 5 mm diameter metal balls was utilized to mix the samples at 30 Hz. The subsequent product for each sample was added with 80% methanol at a volume of 800 µL. The resultant solution was vortexed for 30 s and sonicated thereafter for 30 min at 4 °C. Samples were stored at a temperature of −20 °C for an hour. Each sample was centrifuged thereafter at 12,000 rpm, 4 °C for 5 min. A total of 200 µL of each supernatant from the respective samples, along with 5 µL of DL-o-Chlorophenylalanine (140 μg/mL), were added to vials. The vials were then used for liquid chromatography–mass spectroscopy (LC-MS) analysis.
Untargeted plant metabolomics analysis using UPLC-MS/MS
Separation of compounds was carried out using the Ultimate 3000LC (Thermo, Waltham, MA, United States) combined with QExactive MS (Thermo) and additionally, screened with ESI-MS [27]. The first solvent of the mobile phase comprised 0.05% formic acid in water and the second solvent was acetonitrile with a subsequent gradient elution (0–1 min, 95% A; 1–12 min, 95%–5% A; 12–13.5 min, 5% A; 13.5–13.6 min, 5–95% A; 13.6–16 min, 95% A). The mobile phase had a flow rate of 0.3 mL·min^−1^. The column temperature was kept at 40 °C, while the sample manager temperature was adjusted to 4 °C. The mass spectrometry settings for electrospray ionization ESI+ mode are detailed as follows: heater temperature 300 °C; sheath gas flow rate, 45 arb; auxiliary gas flow rate, 15 arb; sweep gas flow rate, 1 arb; spray voltage, 3.0 kV; capillary temperature, 350 °C; S-Lens RF level, 30%.
Quality control samples
Extracts of the respective plants were mixed as quality control samples (QC) to evaluate the methodology and stability of the LC-MS system. Raw data for the UPLC–MS/MS were obtained from Compound Discover (3.0, Thermo) based on the m/z value and the retention time of the ion signals. To ensure consistency and efficacy of the system, multiple QC samples were formulated through a combination of identical quantities of each isolated sample. The instrument’s performance and repeatability were evaluated through pooled QC samples that were injected before the sample analysis until equilibrium was reached. The QC samples were subsequently run in positive mode. The ion characteristics of the QC samples were utilized to determine the Relative Standard Deviation (RSD). Supplementary Figure S1 of the Supplementary Material displays the percentage of RSD distribution, with a significant portion of the RSD being below 30%. This indicates that the analytical method is reliable and appropriate for application to future sample analyses.
Identification of metabolites
Metabolite identification was performed at a putative level (MSI level 2) utilizing accurate mass measurements and MS/MS fragmentation data. Candidate metabolites were annotated by matching experimental spectra against the Human Metabolome Database1, ChemSpider2 and MassBank3, all accessed 19 August 2024. Then manual evaluation of retention time consistency, isotope patterns, and MS/MS fragmentation spectra was done. Cross-validation across databases was achieved to enhance annotation confidence. Features detected in the ESI^+^ mode were cross-checked for chromatographic and spectral consistency and combined where appropriate. Redundant ions and adducts were resolved utilizing mass accuracy criteria (<5 ppm), retention-time coherence, signal intensity, and % RSD across technical replicates to reduce duplication or erroneous assignments.
Pharmacokinetic and drug-likeness activity of differential metabolites
The properties of the ADME of significant metabolites were extrapolated through in silico methods, and their potential, along with their suitability as drug candidates, was evaluated. The SWISSADME platform4 (accessed on 12 July 2025) was utilized for primary screening, which facilitated the high ranking of compounds that exhibited drug-like profiles and advantageous pharmacokinetic properties. Subsequently, further assessments were carried out with the assistance of recognized cheminformatics inclined tools for a more thorough evaluation of the pharmaceutical relevance of the metabolites. The parameters examined comprised physicochemical descriptors, which included molecular weight, number of heavy aromatic atoms, topological polar surface area (TPSA), molecular refractivity, number of hydrogen bonds, lipophilicity factors (logP), water solubility (logS), along other parameters were examined. Pharmacokinetic traits such as blood–brain barrier (BBB) permeation, gastrointestinal (GI) absorption, substrate specificity for P-glycoprotein (P-gp), inhibition potential essential key cytochrome P450 (CYP) isoenzymes (CYP1A2, CYP2C19, CYP2C9, CYP2D6, and CYP3A4), as well as skin permeation (logKp) were evaluated, and these aspects conjointly affect the metabolism of the potential and the likelihood of drug-to-drug interactions. Ultimately, the final analysis for drug-likeness was conducted using Lipinski, Ghose, Veber, Egan, Muegge, and bioavailability scores. These assessments provide a comprehensive view of the compounds’ appropriateness for further progression.
Statistical analysis
In terms of anti-cancer analysis, GraphPad Prism version 8.2.0 (GraphPad Software, Inc., San Diego, CA, United States) was utilized to plot the dose-response curve and determine the IC_50_ of plant treatments.
While for metabolomics analysis, the raw data was collected and aligned utilizing Compound Discover (3.0, Thermo) according to the m/z values and the retention time of the ion signals. Ions from both ESI+ were combined and subsequently transferred into the SIMCA-P software (version 14.1) and Metaboanalyst version 6.05 (accessed on 12 August 2025) for multivariate analysis. Unsupervised principal component analysis (PCA) was achieved to evaluate intrinsic clusters and the structure of variance. Supervised partial least squares-discriminant analysis (PLS-DA) and orthogonal PLS-DA (OPLS-DA) were also achieved for maximizing class segregation and assessing statistically significant metabolites. Cross-validation of these models was completed, and the variance percentages clarified through orthogonal and predictive components were verified.
Statistically significant metabolites were verified and filtered by integrating the outcomes of the Variable Importance in Projection (VIP) score greater than 1.5, a p-value less than 0.05, FDR-adjusted p < 0.05 and a fold change (FC) greater than 2.0. The R ^ 2 ^ and Q ^ 2 ^ values can explain the quality of the fitted model. R ^ 2 ^ shows the variance accounted for in the model and reflects the goodness of the fit. Q ^ 2 ^ shows the variability in the data, reflecting the model’s predictability.
Results
Anti-cancer activities
The anti-cancer activities of A. calamus and L. javanica extracts on SH-SY5Y neuroblastoma cells were assessed utilizing the CCK-8 assay. These plant extracts displayed a clear dose-dependent decrease in cell viability, as demonstrated by the sigmoidal dose-response curves (Figure 1). The estimated half maximal inhibitory concentration (IC_50_) values were 0.2886 μg/μL for A. calamus and 0.3066 μg/μL for L. javanica, with 0.2976 μg/μL as an overall IC_50_ mean. However, A. calamus displayed slightly higher effectiveness in comparison to L. javanica.
Dose-response curves for (A) A. calamus and (B) L. javanica extracts on SH-SY5Y neuroblastoma cells utilizing the CCK-8 assay.
Mitochondrial membrane potential analysis
Mitochondrial function in SH-SY5Y neuroblastoma cells was evaluated using flow cytometry and the JC-1 fluorescent probe to quantify variations in membrane potential (ΔΨm). As illustrated in Figure 2, in terms of untreated SH-SY5Y control cells, 58.8% of the population was within the Q2 gate, signifying cells with high ΔΨm (red fluorescence), although 38.3% of the population was situated in the Q3 gate, conforming to mitochondrial depolarization (green fluorescence). Following plant extract treatment, the proportion of the cells with high ΔΨm increased to 68.2% for A. calamus and 65.4% for L. javanica, implying increased mitochondrial polarization comparative to untreated controls.
Mitochondrial membrane potential (ΔΨm) analysis of SH-SY5Y neuroblastoma cells using the JC-1 fluorescent probe and flow cytometry.
UPLC-MS/MS base peak intensity chromatograms
The UPLC-MS/MS analysis of A. calamus and L. javanica completed in ESI+ mode produced (BPI) chromatograms, a wide profile of metabolites eluting over a specified retention time frame, as shown in Figure 3.
Base peak intensity (BPI) chromatograms from (A) A. calamus and (B) L. javanica in ESI+ mode.
The chromatogram of A. calamus with the dominating of a particularly intense peak at ∼400 s, with a highest intensity of ∼1.2 × 10^10^. Subsequent moderate peaks were detected at ∼700, ∼550, ∼200, and ∼50 s. Conversely, L. javanica offered an evenly dispersed chromatographic profile with multiple peaks of medium to high intensity detected between 100s and 600s. The most intense peak occurred at ∼220 s (∼4.0 × 10^9^). Additional prominent peaks were detected at ∼720, ∼600, ∼400, and ∼100 s.
Multivariate analysis of A. calamus and L. javanica data
Principal Component Analysis (PCA) score model of the associated features displays complete species segregation across PC1, with a variance of 96.9% (PC2 = 1.0%). The PCA pair-plot validates that cluster variances are concentrated on the first component and also noticeable, but smaller on higher components. The dispersals of scores vary significantly for PC2: p = 0.002, PC3: p = 0.004, PC4: p = 0.004, and PC5: p = 0.002 (Figure 4A). These results signify the possession of distinct global metabolomic fingerprints of the two species, dominated by variance expressed on PC1. The observed clustering patterns were statistically significantly validated using the PERMANOVA test, consisting of an F-value of 936.71, a *p-*value = 0.005, and R ^ 2 ^ = 0.98944.
Multivariate analysis: (A) Principal Component Analysis (PCA) pairwise scatterplots, (B) Partial Least Squares Discriminant Analysis (PLS-DA) scores plot, and (C) Orthogonal Partial Least Squares Discriminant Analysis (OPLS-DA) scores plot of A. calamus (green) and L. javanica (red).
Moreover, supervised Partial Least Squares-Discriminant Analysis (PLS-DA) generated the same qualitative pattern of PCA score model with distinct separated clusters driven by component 1 (96.9%), though component 2 represented insignificant variance of 0.8% (Figure 4B). Cross-validation metrics also validate with R ^ 2 ^ and Q ^ 2 ^ values that were close to 1.0 for all 5 tested components, which displayed consistently high performance. Moreover, the permutation test additionally validated the PLS-DA model, with a noticeable statistic positioned (p < 0.01, 0/100) that was distant outside the permutation distribution (Supplementary Figure S2).
Orthogonal PLS-DA (OPLS-DA) model additionally displays variation into a single predictive component (T score [1] = 96.9%) that segregates the plant species and an orthogonal component (Orthogonal T score [1] = 0.8%) representing within the class structure (Figure 4C). The model was further validated by cross-validation, where the model generated high explanatory and predictive performance, with R ^ 2 ^Y = 1.0, R ^ 2 ^X = 0.969, and Q ^ 2 ^ = 0.999. The related p-values for the predictive component (p1) were R ^ 2 ^X: 0.969; R ^ 2 ^Y: 1.0; Q ^ 2 ^: 0.999, whereas for the orthogonal component (o1) were R ^ 2 ^X: 0.00813, R ^ 2 ^Y: 0.00031, and Q ^ 2 ^: 7.04 × 10^-5^, underscoring the model’s explanatory and predictive capability. Further validation of the model was done using a permutation test, with Q ^ 2 ^ and R ^ 2 ^Y values of 0.969 and 0.979, respectively (Supplementary Figure S2).
Univariate analysis of A. calamus and L. javanica data
Univariate analysis recognized the top ten metabolites with VIP >2.0, FC >2.0, p < 0.05 and FDR-adjusted p < 0.05 that significantly differentiated the clusters of A. calamus and L. javanica (Table 1). Figure 5 depicts major significant metabolites comprising Tetramethylquercetin 3-rutinoside, Isopropyl β-glucoside, and 6β-Hydroxymethandienone, altogether presenting an extreme fold of enrichment. Further significant metabolites comprised of 2-trans-O-Feruloylglucaric acid, 7-Epi-12-hydroxyjasmonic acid, and Leonoside A. Flavonoid derivatives, for instance Quercetin 3-O-glucuronide, Hesperidin methylchalcone, and Apigenin 7-O-diglucuronide were also significantly differentiated. Additionally, Hetastarch also provided a contribution to the metabolic variance. Together, these compounds underscore jasmonates, flavonoid glycosides, and phenolic derivatives as crucial, statistically significant metabolites, supporting the distinct chemical divergence amongst the two plant species. These compounds show higher abundances constantly in L. javanica (Figure 5). A detailed chemical profiling of each compound is represented in the Supplementary Table S1.
Differential metabolites distinguished between A. calamus and L. javanica. Molecular formulas, chemical structures, and boxplots of the top ten compounds (VIP >1.5, FC >2.0, p < 0.05, FDR-adjusted p < 0.05) are shown, with abundances in A. calamus and L. javanica.
Pharmacokinetic and drug-likeness activity of differential metabolites
The assessment of the ten most significant metabolites from A. calamus and L. javanica was conducted through the SWISSADME platform to acquire the ADME profiles of each significant metabolite. This yields essential insights into their potential efficacy as neuroactive inhibitors targeting SH-SY5Y neuroblastoma cells (Table 2).
Physicochemical properties
There was a considerable amount of variability in molecular weight, polarity, and solubility of the ten-highest ranked metabolites, which was determined using the SWISSADME. Smaller molecules such as 7-epi-12-hydroxyjasmonic acid (MW 226.27 g/mol) and isopropyl β-glucoside (MW 222.24 g/mol) had a low topological polar surface area (TPSA <60 Å^2^) and the absence of hydrogen bond donors, indicating a pronounced potential for passive permeability across biological membranes, and these molecules also met the physicochemical criteria for drug-like entities. In contrast, more complex glycosylated flavonoids, such as apigenin 7-O-diglucuronide (MW 622.49 g/mol) and Leonoside A (MW 770.73 g/mol), exhibited slightly elevated TPSA (>250 Å^2^) and standout characteristics such as multiple hydrogen bond donors/acceptors, which are conventionally linked to restricted oral bioavailability.
The indices (consensus Log Po/w) for lipophilicity varied from −5.27 for hetastarch to 4.45 for 6β-hydroxymethandienone. This specifically emphasizes the solubility-permeability trade-offs across the metabolite library. Remarkably, 6β-hydroxymethandienone displayed favorable lipophilicity alongside an acceptable molecular weight, implying a significant potential for permeability; conversely, polar glucuronides (e.g., quercetin 3-O-glucuronide, consensus Log Po/w −0.47) are likely impeded in their capacity for cellular penetration.
Water solubility
Predictions regarding water solubility indicated that hetastarch is highly soluble (>2.0 × 10^4^ mg/mL), while 6β-hydroxymethandienone displayed suboptimal solubility (4.32 × 10^-3^ mg/mL). Intermediate solubility profiles were noted for flavonoid derivatives, such as hesperidin methylchalcone and tetramethylquercetin 3-rutinoside. These findings suggest that formulation strategies may be necessary for highly lipophilic compounds exhibiting poor intrinsic solubility.
Pharmacokinetics
Predictions regarding gastrointestinal (GI) absorption depicted that 6β-hydroxymethandienone, isopropyl β-glucoside, and 7-epi-12-hydroxyjasmonic acid demonstrate high absorption potential, whereas the majority of glycosylated flavonoids exhibited low absorption due to their extensive polar surface areas. Blood-brain barrier (BBB) permeability was anticipated for the smaller hydrophobic compounds, particularly isopropyl β-glucoside, 6β-hydroxymethandienone, and 7-epi-12-hydroxyjasmonic acid, implying a potential for central nervous system (CNS) activity against SH-SY5Y neuroblastoma cells. Significantly, numerous compounds, including hesperidin methylchalcone and apigenin diglucuronide, were identified as substrates for P-glycoprotein (P-gp), suggesting a vulnerability to efflux and diminished intracellular accumulation.
Skin permeation (Log Kp) values were more favorable for lipophilic scaffolds (e.g., −4.33 cm/s for 6β-hydroxymethandienone) in comparison to highly polar compounds (−16.26 cm/s for hetastarch). The analysis of Cytochrome P450 (CYP) inhibition identified 6β-hydroxymethandienone as a multi-isoform inhibitor (CYP2C19, CYP2C9, CYP2D6, CYP3A4), indicating potential drug-drug interaction liabilities, whereas the majority of glycosides exhibited no alerts for CYP inhibition.
Drug-likeness
Filters for Drug-likeness indicated that only isopropyl β-glucoside, 6β-hydroxymethandienone, and 7-epi-12-hydroxyjasmonic acid fully conformed to Lipinski’s rule of five, as well as Veber, Egan, and Muegge criteria, while the majority of glycosides surpassed thresholds for hydrogen bonding, molecular weight, and TPSA. Bioavailability scores corroborated these findings, revealing favorable values of 0.55 for the smaller drug-like molecules in contrast to ≤0.17 for the larger glycosides. These outcomes underscore isopropyl β-glucoside and 6β-hydroxymethandienone as promising candidates for oral bioavailability.
Medicinal chemistry filters
The identification of Brenk alerts, as well as PAINS (Pan-Assay Interference Compounds), has highlighted various flavonoid derivatives (such as hesperidin methylchalcone, Leonoside A, and quercetin 3-O-glucuronide) that exhibit Michael acceptor motifs or catechol, thereby indicating the likelihood of assay interferences. The synthetic accessibility scores exhibited a range from 2.31 (7-epi-12-hydroxyjasmonic acid, which is readily synthesizable) to 8.06 (hetastarch, which is characterized by high complexity), reflecting considerations of feasibility for prospective lead optimization endeavors. Lead-likeness was attained solely for 6β-hydroxymethandienone, while the remaining metabolites were hindered by excessive molecular dimensions or flexibility.
Therefore, the ADME profiling has prioritized three metabolites, namely isopropyl β-glucoside, 6β-hydroxymethandienone, and 7-epi-12-hydroxyjasmonic acid, which emerge as the most promising lead-like candidates for targeting of SH-SY5Y neuroblastoma cells. Larger polar flavonoid glycosides, despite their bioactivity, may necessitate structural simplification or the implementation of advanced delivery systems to mitigate challenges related to poor absorption and permeability. Their favourable oral bioavailability, blood-brain barrier permeability, and adherence to drug-likeness criteria substantiate their potential for subsequent preclinical validation.
Discussion
This study validates that extracts of L. javanica and A. calamus alter mitochondrial functional status and exert dose-dependent cytotoxic effects in SH-SY5Y neuroblastoma cells. The observed shifting toward increased mitochondrial membrane polarization, as evaluated by JC-1 staining, implies preservation or enrichment of mitochondrial reliability at levels of the tested exposure, rather than clear mitochondrial depolarization. Such effects are consistent with complex phytochemical concoctions in which mitochondrial responses might vary depending on exposure duration, concentration, and cellular context [14, 52].
Untargeted metabolomics analysis facilitated the chemical delineation of the phenotypic expressions of substances. With multivariate statistics verifying the distinct chemical differences between the two species, the metabolomic profiling identified a wide range of bioactive substances, including alkaloids, phenolics, flavonoid glycosides, and jasmonates. Separated and distinct groupings were exhibited by L. javanica and A. calamus in PCA, PLS-DA and OPLS-DA, with cross-validation by OPLS-DA and permutation analyses corroborating the non-random nature of the segregation of classes [53]. Given the propensity for supervised models to exhibit overfitting traits, the interpretation of data of the nearly perfect R ^ 2 ^/Q ^ 2 ^ values with circumspection, anchoring conclusions within the framework of unsupervised PCA delineation and independent univariate filters (VIP, fold-change, p values), which is indicative of optimal methodological procedures [54]. The principal discriminative compounds identified were isopropyl β-glucoside, 6β-hydroxymethandienone, 7-epi-12-hydroxyjasmonic acid, 2-trans-O-feruloylglucaric acid, and flavonoid glycosides (e.g., conjugates of quercetin and apigenin) [55]. The identification of a jasmonate (7-epi-12-hydroxyjasmonic acid) is of significant mechanistic relevance; most jasmonates and their derivatives possess the capability to induce cancer cell apoptosis through the disassociation of hexokinase II from the mitochondria, inadvertently, disrupting metabolic processes [56, 57], and multiple research investigations have documented ROS-mediated apoptotic responses across several tumor models [57, 58]. If jasmonate-like chemistry was enriched in L. javanica, the plant may contribute to cytotoxic effects at elevated concentrations, notwithstanding the net mixture’s antioxidative behavior under the experimental conditions employed [35, 36].
The chemistry of L. javanica varies by chemotype (piperitenone-, carvone-, linalool, or myrcenone-rich), which can modulate biological effects [59]. The metabolomic analysis confirms the divergence of composition from A. calamus and focuses on phenylpropanoid, flavonoid, or jasmonate classes for fractionation [33]. The profile for A. calamus inherently invokes concerns pertaining to translation, although β-asarone and some crude extracts have the potential to exhibit antiproliferative activity both in vitro and in vivo [60–62]. β-asarone has been identified as a carcinogenic and genotoxic component in rodents as well as hepatotoxic in preclinical models; this necessitates the establishment of regulatory limits in phytomedicine as well as food products [62]. Consequently, any trajectory for the development of products must prioritize asarone-depleted fractions or alternatives devoid of asarone if the exploration of efficacy is to be conducted safely [42, 43, 62]. It is crucial to note that putative annotations of mescaline-and steroid-like compounds probable indicate structurally associated plant-derived analogues sharing common triterpenoid or phenethylamine biosynthetic scaffolds, instead of definitive identification of canonical reference molecules [48, 63].
Accordingly, SwissADME analysis was employed to provide functional prioritization of annotated metabolites. The SwissADME software was utilized alongside complementary cheminformatics principles (Lipinski, Veber, Ghose, Egan, Muegge), and in conjunction with gastrointestinal absorption/BBB predictions and alert panels (PAINS, Brenk, lead-likeness), this was done to examine the most discriminative metabolites [64–66]. Compounds such as isopropyl β-glucoside, 6β-hydroxymethandienone, and 7-epi-12-hydroxyjasmonic were identified through the internal ADME heatmap as advantageous lead-like candidates that are grounded in minimal rule infractions, bioavailability assessments, and feasible synthetic accessibility [44]. Although SwissADME’s BOILED-Egg framework serves as an ideal tool for preliminary evaluation for BBB and gastrointestinal predictions, further orthogonal verification is usually required through experimental assays such as PAMPA, MDCK/P-gp transport, and microsomal stability as described in this study [47, 67]. Within this limitation-aware framework, in silico ADME predictions still serve as a critical early-stage filter for selecting candidate phytometabolites with translational possibility in neuroblastoma research [68]. Predicted BBB permeability is mostly applicable for neuroblastoma, given the connection of paraspinal, intracranial, or central nervous system-adjacent disease locations, where incomplete tissue penetration can restrict therapeutic efficacy [44]. Moreover, predicted P-glycoprotein (P-gp) substrate status alerts the probability of active efflux, which might decrease intracellular accumulation and influence multidrug resistance [69]. Lastly, cytochrome P450 (CYP) inhibition profiles present understanding into possible metabolic liabilities and drug-drug interaction risks, which are important considerations in pediatric oncology settings where integration therapies are common [70]. In general, these parameters reinforce rational prioritization of phytometabolites for downstream optimization, while emphasizing compounds that might need structural refinement or different delivery approaches.
Natural products remain significantly important as complementary or primary scaffolds for neuroblastoma-based therapies, particularly polyphenols, which are a class of secondary metabolites known to influence apoptosis activity and autophagy, and may also specifically target neuroblastoma stem-like cell populations [71, 72]. A compilation of recent scholarly reviews has systematically outlined a variety of phytochemicals exhibiting preclinical activity against neuroblastoma and highlighted proposed combinations of probable strategies aimed at augmenting responses to standard therapeutic regimens [73]. Within this body of literature, our findings motivate a chemotype-specific follow-up: (i) to deprioritize asarone-rich fractions of A. calamus or to specifically eliminate β-asarone before experimental testing due to safety concerns [62], (ii) to assess and evaluate flavonoid conjugates for their potential to modulate redox status and kinase signaling pathways in neuroblastoma [74], and (iii) to extract jasmonate-like metabolites from L. javanica for the purpose of investigating the direct targeting of mitochondrial pathways [57].
Together, these results offer a rationale for structured future investigations that incorporate chemotype-informed isolation approaches, bioactivity-guided fractionation, and target-focused mechanistic assessment.
Future directions
Building on potential chemotypic signatures identified by incorporating univariate and multivariate metabolomic analyses, future work should prioritize targeted isolation of potential metabolites from dominant chemical classes. Bioactivity-guided fractionation methods can be critical to connect specific phytochemical fractions to functional effects studies in neuroblastoma cells and to isolate active constituents from complex extract mixtures. In silico structure-based molecular docking and molecular dynamics can be used to screen relevant molecular targets against neuroblastoma. To support rational prioritization of candidate chemotypes and guide hypothesis-driven mechanistic investigations.
Conclusion
This study employed an integrated workflow of untargeted metabolomics, mitochondrial functional assessment, and in silico ADME to explore phytometabolites from L. javanica and A. calamus in human SH-SY5Y neuroblastoma cells. Univariate and multivariate investigations uncovered distinct chemotypic signatures, with L. javanica characterized by flavonoid- and polyphenol-enriched profiles and A. calamus was dominated by jasmonate-, phenylpropanoid- and lipophilic terpenoid-associated metabolites. Extract exposure related to dose-dependent reductions in cell viability and modulation of mitochondrial membrane potential. In silico pharmacokinetic predictions supported early-stage prioritization of selected chemotypes such as isopropyl β-glucoside, 6β-hydroxymethandienone, and 7-epi-12-hydroxyjasmonic acid. These results are preliminary and hypothesis-generating, and further mechanistic confirmation and in vivo studies are recommended to establish biological mechanisms and translational importance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1National Cancer Institute. Childhood neuroblastoma treatment (PDQ®)–health professional version (2025). Available online at: https://www.cancer.gov/types/neuroblastoma/hp (Accessed September 12, 2025).26389190 · pubmed ↗
- 2D’Ambrosio N Lyo JK Young RJ Haque SS Karimi S . Imaging of metastatic CNS neuroblastoma. AJR Am J Roentgenol (2010) 194(5):1223–9. 10.2214/AJR.09.3203 20410407 · doi ↗ · pubmed ↗
- 3Vo KT Matthay KK Neuhaus J London WB Hero B Ambros PF Clinical, biologic, and prognostic differences on the basis of primary tumor site in neuroblastoma: a report from the international neuroblastoma risk group project. J Clin Oncol (2014) 32(28):3169–76. 10.1200/JCO.2013.54.2315 25154816 PMC 4171360 · doi ↗ · pubmed ↗
- 4Motshudi MC Naidoo CM Mkolo NM . The race against time for the enhancement of African national strategic plans in the neuroblastoma research heterogeneity. Publications (2024) 12(4):45. 10.3390/publications 12040045 · doi ↗
- 5Davidoff AM . Neuroblastoma. Semin Pediatr Surg (2012) 21(1):2–14. 10.1053/j.sempedsurg.2011.10.009 22248965 PMC 3261589 · doi ↗ · pubmed ↗
- 6Speleman F Park JR . Henderson TO. Neuroblastoma: a tough nut to crack. Am Soc Clin Oncol Educ Book (2016) 35:548–57. 10.1200/EDBK_159169 27249766 · doi ↗ · pubmed ↗
- 7Cohn SL Pearson ADJ London WB Monclair T Ambros PF Brodeur GM The international neuroblastoma risk group (INRG) classification system: an INRG task force report. J Clin Oncol (2009) 27(2):289–97. 10.1200/JCO.2008.16.6785 19047291 PMC 2650388 · doi ↗ · pubmed ↗
- 8Matthay KK Maris JM Schleiermacher G Nakagawara A Mackall CL Diller L Neuroblastoma. Nat Rev Dis Primers (2016) 2:16078. 10.1038/nrdp.2016.78 27830764 · doi ↗ · pubmed ↗