Brucella abortus histidine auxotrophs are copper sensitive
Charline Focant, Agnès Roba, Elisabeth Wanlin, Katy Poncin, Xavier De Bolle

TL;DR
This study finds that histidine auxotrophy, not copper sensitivity, is the main reason for reduced growth of Brucella abortus in macrophages.
Contribution
The study reveals that histidine biosynthesis is crucial for Brucella's survival in macrophages, not copper resistance.
Findings
Histidine auxotrophs of Brucella abortus are sensitive to copper stress.
Suppressor strains show that histidine auxotrophy, not copper sensitivity, causes attenuation in macrophages.
Overproduction of a homolog of OppA restores copper resistance in histidine auxotrophs.
Abstract
Despite decades of investigation into bacterial pathogens, the conditions met by intracellular bacteria are still unclear. These conditions can include access to nutrients, such as amino acids, and exposure to toxic compounds, like copper. To investigate the ability of Brucella abortus, a facultative intracellular pathogen responsible for a major zoonosis, to cope with copper, we performed a Tn-seq analysis to identify copper-sensitive mutants. Unexpectedly, we realized that classical copper resistance systems (involving CopA and CueO homologs) do not appear to be robustly needed, while histidine and purine biosynthesis pathways are crucial to cope with copper. We show that hisA, hisB, hisC, and hisD mutants are auxotrophic for histidine and sensitive to copper. This suggests that the reported attenuation of his mutants in macrophages could be based on auxotrophy and/or copper…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
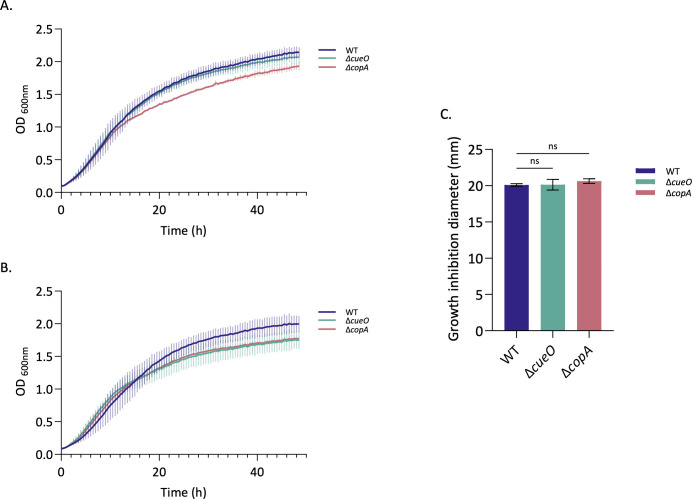 Fig 1
Fig 1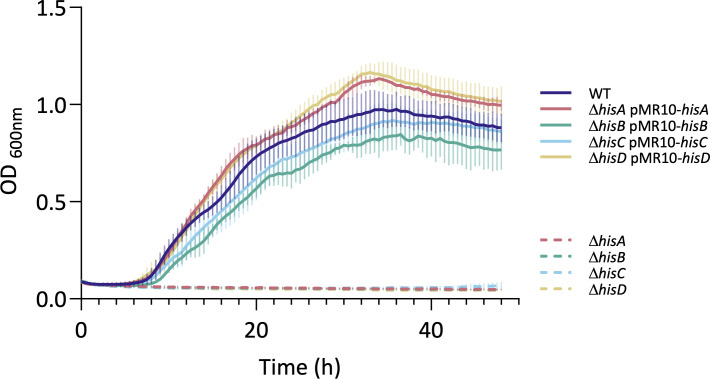 Fig 2
Fig 2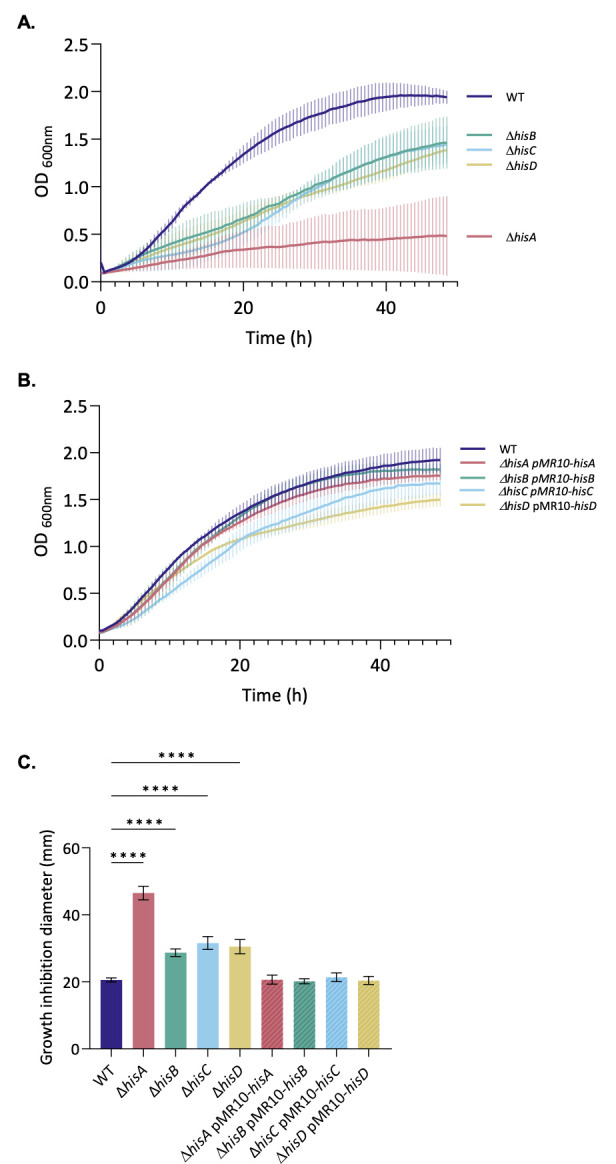 Fig 3
Fig 3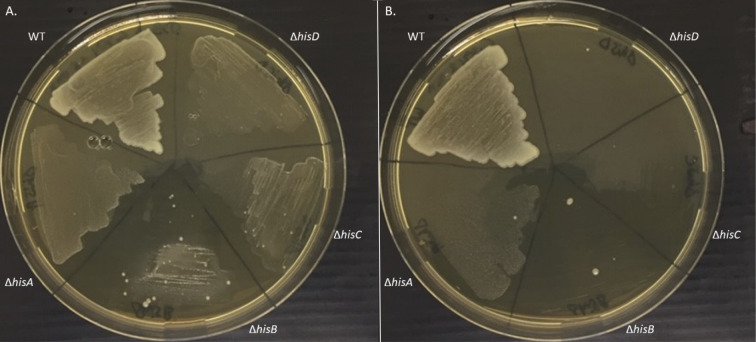 Fig 4
Fig 4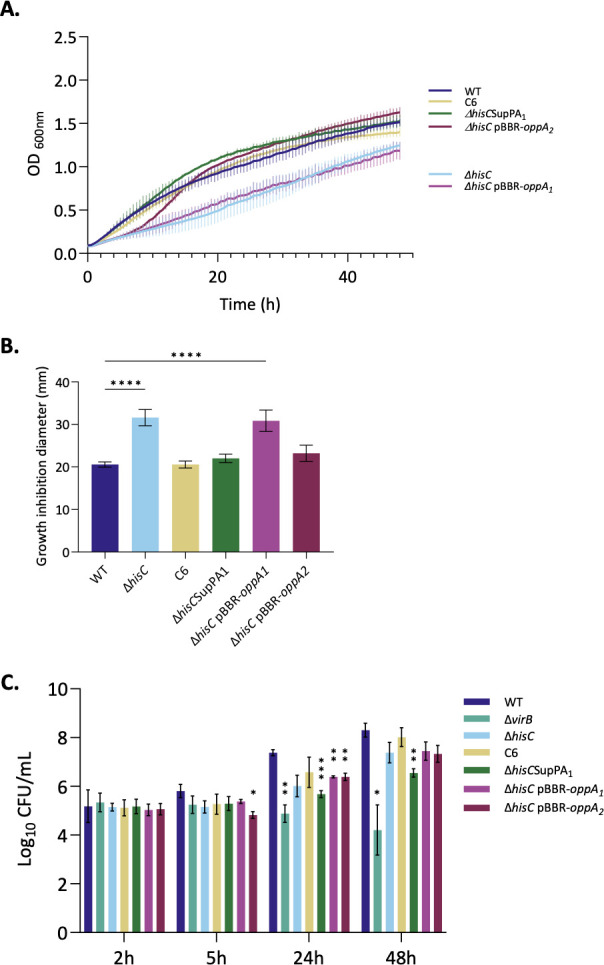 Fig 5
Fig 5| Name | Locus tag | Product | TnIF (CT) | TnIF (Cu) | ΔTnIF | |
|---|---|---|---|---|---|---|
| 544 | 2308 | |||||
| Cue system | ||||||
| | BAB_v1_a0229 | BAB1_0222 | DNA-binding transcriptional dual regulator CueR | 2.61 | 0.95 | −1.66 |
| | BAB_v1_a0228 | BAB1_0221 | Cu(+) exporting P-type ATPase | 2.76 | 0.55 | −2.22 |
| | BAB_v1_b0532 | BAB2_0534 | Blue copper oxidase CueO | 2.75 | 0.39 | −2.36 |
| Histidine biosynthesis | ||||||
| | BAB_v1_a0293 | BAB1_0285 | Histidinol dehydrogenase | 3.04 | 0.55 | −2.49 |
| | BAB_v1_b0179 | BAB2_0182 | ATP phosphoribosyltransferase regulatory subunit | 2.57 | 0.41 | −2.16 |
| | BAB_v1_b0180 | BAB2_0183 | ATP phosphoribosyltransferase | 2.63 | 0.61 | −2.02 |
| | BAB_v1_a2099 | BAB1_2086 | Imidazole glycerol phosphate synthase subunit HisF | 2.31 | 0.58 | −1.73 |
| | BAB_v1_a1109 | BAB1_1098 | Phosphoribosyl-AMP cyclohydrolase | 2.92 | 1.22 | −1.70 |
| | BAB_v1_a2097 | BAB1_2084 | Imidazole glycerol phosphate synthase subunit HisH | 2.62 | 0.98 | −1.65 |
| | BAB_v1_a2095 | BAB1_2082 | Imidazoleglycerol-phosphate dehydratase | 2.45 | 0.87 | −1.59 |
| | BAB_v1_a2100 | BAB1_2087 | Phosphoribosyl-ATP pyrophosphatase | 3.10 | 1.53 | −1.57 |
| | BAB_v1_a2005 | BAB1_1988 | Histidinol-phosphate aminotransferase | 1.71 | 0.67 | −1.04 |
| | BAB_v1_a2098 | BAB1_2085 | 1-(5-phosphoribosyl)−5-[(5-phosphoribosylamino) | 2.49 | 0.38 | −2.11 |
| Purine biosynthesis | ||||||
| | BAB_v1_a1839 | BAB1_1824 | Fragment of bifunctional AICAR transformylase/IMP | 1.74 | 0.73 | −1.01 |
| | BAB_v1_a1838 | BAB1_1824 | Fragment of bifunctional AICAR transformylase/IMP | 1.58 | 0.70 | −0.87 |
| | BAB_v1_a0751 | BAB1_0730 | Phosphoribosylglycinamide formyltransferase 1 | 1.93 | 1.32 | −0.60 |
| | BAB_v1_a1774 | BAB1_1758 | N5-carboxyaminoimidazole ribonucleotide synthase | 1.91 | 1.32 | −0.59 |
| | BAB_v1_a0449 | BAB1_0442 | Phosphoribosylamine--glycine ligase | 2.04 | 1.49 | −0.55 |
| | BAB_v1_a1773 | BAB1_1757 | N (5)-carboxyaminoimidazole ribonucleotide mutase | 2.14 | 1.59 | −0.54 |
| | BAB_v1_a0480 | BAB1_0472 | Amidophosphoribosyltransferase | 1.91 | 1.45 | −0.46 |
| | BAB_v1_a0881 | BAB1_0862 | Phosphoribosylaminoimidazole-succinocarboxamide synthase | 1.74 | 1.29 | −0.46 |
| | BAB_v1_a0879 | BAB1_0860 | Phosphoribosylformylglycinamidine synthase subunit PurQ | 2.40 | 1.95 | −0.45 |
| | BAB_v1_a0752 | BAB1_0731 | Phosphoribosylformylglycinamide cyclo-ligase | 2.00 | 1.62 | −0.39 |
| Pyridoxin biosynthesis | ||||||
| | BAB_v1_a1420 | BAB1_1404 | Pyridoxine 5′-phosphate synthase | 2.54 | 0.80 | −1.74 |
| | BAB_v1_a0724 | BAB1_0705 | 4-hydroxythreonine-4-phosphate dehydrogenase | 2.26 | 0.63 | −1.63 |
| | BAB_v1_a0451 | BAB1_0444 | Pyridoxine/pyridoxamine 5′-phosphate oxidase | 1.95 | 0.89 | −1.06 |
| Others | ||||||
| | BAB_v1_a0442 | BAB1_0434 | D-2-hydroxyglutarate dehydrogenase | 2.57 | 0.85 | −1.72 |
| | BAB_v1_a0329 | BAB1_0318 | Phosphoglycolate phosphatase | 2.47 | 1.07 | −1.40 |
| | BAB_v1_a0196 | BAB1_0188 | Cobalamin-dependent methionine synthase | 2.35 | 1.57 | −0.77 |
| | BAB_v1_a0920 | BAB1_0903 | Sec-independent protein translocase protein TatC | 1.36 | 0.76 | −0.61 |
| | BAB_v1_a0918 | BAB1_0901 | Sec-independent protein translocase protein TatA | 1.65 | 1.21 | −0.44 |
| | BAB_v1_a0661 | BAB1_0639 | LPSA protein | 1.83 | 0.60 | −1.23 |
| | BAB_v1_a0061 | BAB1_0055 | Phosphoglucomutase | 1.31 | 0.77 | −0.54 |
| | BAB_v1_b0989 | BAB2_1005 | FtsZ-localized protein A | 1.70 | 1.17 | −0.53 |
| | BAB_v1_a2001 | BAB1_1984 | Diaminopimelate decarboxylase | 2.04 | 1.60 | −0.44 |
| | BAB_v1_a1939 | BAB1_1916 | Tyrosine recombinase XerC | 2.02 | 1.58 | −0.44 |
| | BAB_v1_a2110 | BAB1_2099 | Adenosylhomocysteinase | 1.85 | 1.42 | −0.43 |
| | BAB_v1_b0358 | BAB2_0361 | Ribosome-dependent GTPase%2C ribosome assembly factor | 1.95 | 1.58 | −0.37 |
| Mutant | Suppressor | Position | Type | Ref. | Alt. | Locus_tag | Name | |
|---|---|---|---|---|---|---|---|---|
|
| B1 | chr2. 695058 | Del | * | C | BAB_v1_b0690 | BAB2_0699 |
|
| B9 | chr2. 695451 | SNP | G | C | BAB_v1_b0691 | BAB2_0700 |
| |
| chr2. 695457 | Complex | TCG | ACC | |||||
| chr2. 695468 | Complex | CATC | TCTG | |||||
| chr2. 695483 | SNP | G | A | |||||
| chr2. 695489 | SNP | A | T | |||||
| B10 | chr2. 694185 | SNP | C | T | BAB_v1_b0690 | BAB2_0699 |
| |
| chr2. 694191 | Complex | GAAA | AAAG | |||||
| chr2. 694200 | Complex | ACAG | GGAT | |||||
| chr2. 694219 | Complex | CCG | TCT | |||||
|
| C6 | chr2. 693379 | SNP | G | T | Potential promoter region operon | ||
| C8 | chr2. 693942 | SNP | C | T | BAB_v1_b0690 | BAB2_0699 |
| |
| chr2. 693957 | SNP | C | G | |||||
| chr2. 693966 | SNP | A | C | |||||
| chr2. 695612 | Complex | AAG | CTC | BAB_v1_b0691 | BAB2_0700 |
| ||
- —Fédération Wallonie-Bruxelleshttp://dx.doi.org/10.13039/501100002910
- —Fédération Wallonie-Bruxelleshttp://dx.doi.org/10.13039/501100002910
- —FRFS-Welbio
- —FRS-FNRS
- —FRS-FNRS
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBrucella: diagnosis, epidemiology, treatment · Trace Elements in Health · Metal Extraction and Bioleaching
INTRODUCTION
Brucella spp. are gram-negative bacteria that belong to the Hyphomicrobiales order, which is part of the alpha-proteobacteria class (1, 2). Bacteria from the genus Brucella are intracellular, facultatively extracellular pathogens responsible for brucellosis (3). This worldwide zoonosis disease affects a wide range of hosts, including domestic animals and humans, illustrating the complexity of Brucella spp. (4). The tropism of the bacteria for the reproductive systems correlates with abortion and sterility as main symptoms in animals, while humans present undulant fever symptoms (3, 5, 6). Brucella employs sophisticated strategies to enter, survive, and proliferate within host cells—encompassing both professional phagocytes and non-phagocytic cells—thereby effectively ensuring its pathogenicity (3, 6). Once within the cell, Brucella is found enclosed in a membrane-bound compartment, the Brucella-containing vacuole (BCV), which traffics along the endocytic pathway and undergoes an acidification of the compartment, becoming the endocytic BCV (eBCV) (7, 8). The vacuole acidification is crucial to produce the Brucella VirB Type IV secretion system (7, 9), which secretes effector proteins inside the cell (10, 11). The establishment of the replicative niche (rBCV) inside the endoplasmic reticulum (ER) is dependent on the VirB function (7, 8). As a last step, the rBCVs are converted into autophagic BCVs (aBCVs) by getting associated to atypical autophagic membranes (12). Inside the aBCVs, Brucella can egress from the cell, being therefore able to reinfect the surrounding cells (12). During its trafficking, Brucella probably encounters many stresses, most of which are unknown or indirectly deduced from attenuated mutants.
In the arms race of host-pathogen co-evolution, host cells use several strategies, one of which is the sequestration of key metal ions for the pathogenic bacteria (13). Iron deprivation by host cells is well established. Most iron ions are predominantly sequestered, for example, bound by transferrin and hemoglobin, rendering them unavailable to bacteria (14, 15). Neutrophils recruited to the infection site release calprotectin, which sequesters manganese and zinc (16, 17). Manganese and iron can also be depleted from the phagosomes through the natural resistance-associated macrophage protein 1 (NRAMP1) in macrophages and neutrophils (13, 18). Despite the well-established role of metal starvation as a defense mechanism against pathogens, it is becoming increasingly evident that metal intoxication also occurs during infection to fight invading pathogens (16, 19). Host cells exploit copper toxicity to poison intrusive pathogens (20). In response to interferon-γ (IFN-γ) stimulation, activated macrophages accumulate copper within phagolysosomes (21). IFN-γ induces the expression of the copper transporter CTR1, allowing copper import into the cytosol (21, 22), where it is delivered by the ATOX1 chaperone to the ATP7A transporter associated with the phagolysosomal membrane (21, 23–25) and able to pump copper into the lumen of the phagosome. ATP7A expression is also upregulated by IFN-γ (24). Copper resistance has been linked to the virulence of intravacuolar pathogens, such as Mycobacterium tuberculosis (25, 26) and Salmonella enterica (27).
Several defensive mechanisms are used by bacteria to combat toxic concentrations of metals. A simple mechanism of averting metal toxicity is to sequester metal ions within the periplasm or the cytoplasm, resulting in fewer than one copper atom being available per cell (15, 28, 29). When metal ions accumulate to toxic levels, bacteria induce the expression of specific adapted genes. This defense response includes the induction of efflux systems as well as proteins involved in metal sequestration and storage (16). The bacterial response to copper could involve a core cue regulon, which is relatively well conserved among proteobacteria (28). The cue regulon includes CueR, a copper-responsive metalloregulatory protein that upregulates cueO and copA expression upon increased intracellular copper concentration (30). CueO, the periplasmic multicopper oxidase, oxidizes cuprous ion to cupric ion, which is less harmful (31). CopA is a copper efflux P-type ATPase (32). A cuprochaperone, CopZ, is also part of the Cue efflux system (33). Most S. enterica serotypes possess a duplication of the cue regulon, known as the gol regulon (34, 35). The expanded anti-copper arsenal of S. enterica is presumably an adaptation for survival within macrophages (28, 34). A second copper efflux system, the Cus system, is also present in Escherichia coli and S. enterica. This second regulon is controlled by the two-component system CusRS (28), which activates the expression of the cusCFBA operon in response to elevated copper levels (31). The cusCBA genes encode the CusCBA complex, a resistance, nodulation, and division (RND) proton-cation antiporter (28, 31), while cusF encodes a periplasmic cuprochaperone (28).
The specific environmental conditions encountered by intracellular bacteria remain poorly characterized. These conditions may involve limited access to nutrients, such as amino acids, and exposure to compounds like copper. To explore how B. abortus responds to copper stress, we conducted a transposon sequencing (Tn-seq) analysis to identify mutants with increased copper sensitivity. Although the classical CueO and CopA homologs do not appear to be required for B. abortus resistance to copper, the histidine biosynthesis pathway was found to be necessary for growth in the presence of copper. Since it is known (and confirmed here) that his mutants are attenuated in macrophages, this phenotype could be due to auxotrophy for histidine and/or to sensitivity to copper. By generating suppressors with a restored copper resistance, we show that the attenuation of the his mutants in macrophages is probably not related to copper sensitivity.
RESULTS
Identification of genes involved in copper resistance in B. abortus
The mechanism by which Brucella copes with copper toxicity remains unknown. To identify genes required to face copper stress, a Tn-seq was performed on B. abortus 544 using a mini-Tn5 (36). To select the appropriate copper concentration for the Tn-seq, B. abortus and E. coli mini-Tn5 carrier strains were first mated. After incubation and optical density (OD) normalization, serial 10-fold dilutions were spotted on TSB agar plates supplemented with an increasing CuSO₄ concentration, and CFUs were then counted to identify the optimal copper concentration. A final concentration range of 1.6–2 mM CuSO₄ was chosen for the selection corresponding to the concentration where wild type (WT) was not impacted. Once the copper concentration to use for the Tn-seq was determined, the mating was repeated, and the Brucella library was grown on rich medium for the control condition and 2 mM CuSO_4_-supplemented media for the test condition. From the library, 4.8 × 10^6^ random mutants were recovered and sequenced using a deep sequencing method. We identified 1,684,839 and 1,743,143 unique insertion sites for control and copper conditions, respectively, illustrating genome saturation with a unique insertion every 1.95 bp for control and every 1.89 bp for the stress condition, on average. The raw data were analyzed by an automatic process called TnBox, following a previously described method (37). The transposon insertion frequency (TnIF) for each gene was computed according to the following definition: TnIF is defined as the per-gene average of log₁₀(r + 1)/l, where r is the number of miniTn5 insertions at a given nucleotide and l is the coding sequence length. The calculation was performed considering only the central 80% of the coding sequence (37). A TnIF value below the genome-wide average indicates that disruption of the gene decreases bacterial fitness. Then, the ΔTnIF was determined as the difference between the TnIF under copper stress conditions and the TnIF under control conditions (ΔTnIF = TnIFcopper − TnIFcontrol). A gene will be annotated as required to grow in the presence of copper if the ΔTnIF is lower than two times the standard deviation of all ∆TnIF values (ΔTnIF > −0.357). Through the Tn-seq analysis, 38 genes were identified as essential for growth under copper-supplemented conditions (Table 1).
The genes coding for the Cue system, copA, cueO, and the regulator cueR, involved in copper resistance, appeared to be essential in the Tn-seq copper condition (Table 1), as expected compared to the data available from the work in E. coli (31). CopA is an efflux P-type ATPase of the cue regulon, while CueO is a periplasmic multi-copper oxidase (32). The expression of both copA and cueO is regulated by the copper-responsive transcriptional regulator CueR (30). The cue regulon also includes copZ (BAB1_0960 in B. abortus 2308 and BAB_v1_a0976 in B. abortus 544 [33]), which is not scored as required in the Tn-seq copper condition.
Genes coding for the whole histidine biosynthesis pathway (hisA, hisB, hisC, hisD, hisE, hisF, hisG, hisH, and hisI) are highlighted in our Tn-seq to be required to cope with copper stress. Each step of the pathway is catalyzed by a specific enzyme, where the five first steps lead to the production of imidazole glycerol phosphate (IGP) and 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR), an intermediate in purine biosynthesis (see Fig. S1). Histidine is produced from IGP after four other enzymatic reactions (see Fig. S1). While in E. coli and S. enterica Typhimurium, the his pathway is genetically organized in a single operon, it is distributed across both chromosomes in Brucella spp. (38).
Linked to the histidine biosynthesis pathway, the purine de novo synthesis pathway appeared to be also crucial to face copper stress. Indeed, most of the genes*—purH* (part I and II), purN, purK, purD, purE, purF, purC, purQ, and *purM—*required for inosine monophosphate (IMP) production were found to be required in the presence of copper.
Next, three genes from the vitamin B6 biosynthesis (pdxJ, pdxA, and pdxH) also emerged from our analysis as being required. These genes code for enzymes allowing the biosynthesis of vitamin B6, in its active form, pyridoxal 5′-phosphate (39). Vitamin B6 is utilized as a cofactor by the HisC enzyme (40), which may explain why it is required to face copper stress.
Finally, there were 12 other genes that also presented a ΔTnIF lower than two times the standard deviation. Among them, glcD, tatC, pgm, and fzlA were already essential in the control condition, while metH, tatA, wadA, lysA, xerC, ahcY, and bipA had low fitness in the control condition. Only gph did not present any impact on growth in the absence of copper.
CopA and CueO are not required for copper resistance in Brucella abortus
To date, the mechanisms underlying copper resistance in B. abortus remain largely uncharacterized. Only a few homologs of genes involved in copper homeostasis are predicted from the Brucella genomes, including copA, cueO, and cueR (41). These three genes were identified as required under copper stress conditions in our Tn-seq screen (Table 1). Given their predicted function in copper transport and detoxification, we first focused on testing the role of copA and cueO in B. abortus copper resistance by constructing markerless single-gene deletion mutants called ΔcopA and ΔcueO. The growth of ΔcopA and ΔcueO was monitored during 48 h in liquid culture in rich medium in the presence of 2 mM CuSO_4_ (Fig. 1B). Unexpectedly, both mutants did not present any sensitivity to copper stress, displaying growth curves similar to the WT strain. ΔcopA presented a slight growth defect in the control condition (Fig. 1A) compared to the WT, which was also found in the copper condition (Fig. 1B). The minor effect of copper on ΔcueO was not confirmed when the copper concentration was increased.
The copA and cueO genes are not required for copper resistance. WT, ΔcopA, and ΔcueO strains were tested for their sensitivity to copper toxicity for growth. (A) WT, ΔcopA, and ΔcueO strains were grown in liquid TSB rich medium. (B) WT, ΔcopA, and ΔcueO strains were grown in TSB rich medium containing 2 mM CuSO4. (A and B) The OD at 600 nm was measured every 30 min for 48 h. The data represent three independent replicates. (C) Five hundred microliters of an overnight culture of each B. abortus strain, normalized to an OD of 0.1, was added to 4.5 mL of TSB soft agar (0.7% agar) and then plated on a TSB plate. In the middle, a well was dug and filled with 100 µL of 200 mM CuSO4. Inhibition zones around the agar well were measured after 3 days of incubation at 37°C, and the average diameter (± standard deviation, SD) was reported in a histogram. The data represent the mean ± SD and were compiled from three independent replicates. Statistical analysis was carried out by a one-way ANOVA followed by Dunnett’s multiple comparisons test (n.s., non-significant, P > 0.05).
We also conducted a soft agar well diffusion assay where the well was filled with 100 µL of 200 mM CuSO_4_. The inhibition zone was measured around the well and reported in millimeters, with the results presented as a bar graph. As with the growth curves, ΔcopA and ΔcueO displayed the same inhibition zone as the WT strain around the copper source, confirming the absence of sensitivity of the two mutants in these conditions (Fig. 1C).
Taken together, these results demonstrate that the deletion of copA or cueO does not impact copper resistance in B. abortus, contrary to what has been shown in E. coli, and also in sharp contrast to our Tn-seq data (Table 1).
The his mutants are auxotrophic for histidine and sensitive to copper
In addition to our Tn-seq results, histidine biosynthesis genes have already been unveiled as required in B. abortus during RAW 264.7 macrophages infection (36), as well as previous studies with B. suis (42, 43) and Tn-seq with B. melitensis in mice (37). A series of deletion mutants in the histidine biosynthesis pathway has been constructed: hisA, hisB (previously reported [44]), hisC, and hisD. The auxotroph phenotypes of these four mutants have been confirmed by their inability to proliferate in minimal medium in the absence of amino acids. Their growth was rescued either when the gene was introduced in trans on a single-copy plasmid (Fig. 2) or if histidine was provided in the culture medium.
B. abortus ΔhisA, ΔhisB, ΔhisC, and ΔhisD are auxotrophic for histidine. WT and his mutant strains were tested for their histidine auxotrophy. Strains were grown in liquid PE minimal medium. OD600 nm was measured every 30 min for 48 h. The pMR10 is a low-copy plasmid. The data represent three independent replicates.
Since the classical copper homeostasis actors (CopA, CueO) were not required for the growth of B. abortus in the presence of a copper stress, we decided to investigate the histidine biosynthesis pathway in the context of copper stress. The growth of the his mutants in the presence of copper was studied through diverse approaches. First, all strains were cultivated in liquid rich medium with a high copper concentration (1.6 mM), and the OD was measured over a period of 48 h (Fig. 3A), as done with ΔcopA and ΔcueO strains. As suggested by the Tn-seq results, all auxotrophic mutants struggled to grow in the presence of the metal compared to the WT strain, which did not demonstrate a growth defect. The hisA mutant was particularly affected. However, it already struggled in the control condition compared to the other strains (see Fig. S2). The same sensitivity phenotype for all the his mutants was observed with 2 mM CuCl_2_, indicating that the growth defect was due to copper rather than sulfate. The copper sensitivity of the his auxotroph mutants was rescued upon genetic complementation (Fig. 3B). Similar observations were made with the same strains cultivated with a concentration of 2 mM of CuSO_4_, but with an incomplete complementation (see Fig. S3). These preliminary results suggested that impairment of the histidine biosynthesis pathway leads to copper sensitivity. The sensitivity of his mutants to copper was confirmed by a soft agar well diffusion assay in which the inhibition zone around the copper-containing well was measured. As with growth curves, the copper sensitivity was rescued when the strains were complemented with the missing genes (Fig. 3C). The growth of the four his auxotroph mutants was tested on solid TSB medium supplemented with 1.6 or 2 mM CuSO_4_. Results showed a growth defect for each his mutant, in contrast to the WT strain, which grew normally (Fig. 4). On these plates, a few colonies among the mutants were observed. The presence of mutations in these colonies could potentially enable them to withstand the presence of copper. In this case, these colonies could be suppressor clones, that is, clones in which a second mutation cancels out the phenotypic effect of the first mutation (ΔhisA, ΔhisB, ΔhisC, or ΔhisD). Identifying these secondary mutations could therefore provide valuable insight to understand how these strains can cope with copper.
*Histidine auxotrophic mutants are copper sensitive. WT, ΔhisA, ΔhisB, ΔhisC, ΔhisD, ΔhisA pMR10-hisA, ΔhisB pMR10-hisB, ΔhisC pMR10-hisC, and ΔhisD pMR10-hisD strains were tested for their sensitivity to copper toxicity for growth. (A) WT, Δhis, and (B) complemented strains were grown in liquid TSB rich medium containing 1.6 mM of CuSO4. OD600 nm was measured every 30 min for 48 h. The data represent three independent replicates. (C) Experiment performed as in Fig. 1C. Data represent the mean ± SD and were compiled from three independent replicates. Statistical analysis was carried out by a one-way ANOVA followed by Dunnett’s multiple comparisons test (***P <0.0001).
B. abortus WT and mutant strains are copper sensitive on plates. WT and ΔhisA, ΔhisB, ΔhisC, and ΔhisD mutant strains (in this counterclockwise order) were tested for their copper sensitivity on TSB medium containing 1.6 mM (A) or 2 mM (B) of CuSO4. Both plates were divided into five parts where 20 μL of each culture was deposited and spread after their OD600 nm was normalized to 0.1. Pictures were taken after a 4-day incubation at 37°C. The data represent three independent replicates. It is interesting to note the presence of individual colonies for ΔhisA, ΔhisB, ΔhisC, and ΔhisD, suggesting that suppressor mutants could be obtained for this phenotype.
Most suppressor mutations affect a pair of periplasmic proteins
To verify the suppressor phenotype of the colonies isolated from the copper-supplemented TSB plates, it was necessary to streak each of them onto a new plate with and without copper. Of the 29 potential suppressors isolated, all mutant backgrounds combined, eight were unable to retain the ability to grow on copper-supplemented media. The remaining potential suppressors were able to grow at 2 mM CuSO_4_. The genomic DNA of the remaining 21 copper-resistant clones was extracted and analyzed by whole-genome sequencing to identify the mutations that could confer copper resistance in these suppressors. As a control, the genomes of the WT and the parental ΔhisA, ΔhisB, ΔhisC, and ΔhisD strains were also sequenced. To investigate mutations that were strictly present in the potential suppressor, only the mutations present in the suppressor and not in the parental strain were retained. Strikingly, of the 22 unique mutations identified, 15 appeared within the same operon, the opp operon (Table 2). Opp is homologous to an oligopeptide transport system, and it is encoded by five genes. The first two genes, oppA1 and oppA2, encode periplasmic proteins. The next two genes, oppB and oppC, encode permeases, while the final gene, oppD/F, encodes an ATP-binding protein. Interestingly, only the first two genes were touched by a mutation in the suppressors (Table 2).
The suppressive mutation helps to face copper stress in vitro
In this study, we opted to exclude suppressors derived from the hisB mutant. Indeed, hisB is located downstream of the intersection between the histidine synthesis pathway and purine metabolism, via the production of AICAR (Fig. 1). Moreover, in the laboratory, hisB mutant had been studied and demonstrated to exhibit a morphological abnormality by the formation of chains, which is associated with a deficiency in cell division (44). It is also noteworthy that suppressors exhibiting a unique mutation within hisA or hisD background did not affect the opp operon (see Table S1). Out of all mutations in the hisC mutant background, one stood out as particularly interesting. It is located in the suppressor named C6 (the sixth suppressor isolated within the hisC mutant), 70 pb upstream of the oppA1 ATG, in a potential distal promoter region (Table 2). The mutation was investigated as it could influence the expression of the first two coding sequences of the operon.
For the purpose of confirming that the suppressor phenotype is attributable to the identified mutation, it was introduced back into the hisC mutant. The potential distal promoter region where the SNP occurred was amplified from C6 genomic DNA and subsequently inserted into ΔhisC strain by allelic exchange. The new strain, including the C6 mutation, was named ΔhisCSupPA1 (standing for Suppressor Promoter OppA1). To investigate the hypothesis that the suppressive mutation in the potential promoter enhances the abundance of OppA1 and OppA2 substrate proteins, an overexpression vector for each of the genes was constructed in the pBBR-MCS2 medium-copy plasmid and introduced into the ΔhisC mutant, generating ΔhisC pBBR-oppA1 and ΔhisC pBBR-oppA2 strains, respectively. The histidine auxotroph phenotype was first evaluated by measuring the OD in a minimal medium, as previously performed with the his mutants. All strains retained the auxotrophic phenotype of the mutant parental strain. Indeed, the growth was rescued when the minimal medium was supplemented with 1 mM of histidine (See Fig. S4).
The suppression phenotype was then studied to confirm its role in the rescue process in the presence of copper stress. For this purpose, the same experiments as for histidine auxotrophic mutants were carried out. The growth of the strains WT, ΔhisC, C6, ΔhisCSupPA1, ΔhisC pBBR-oppA1, and ΔhisC pBBR-oppA2 was observed in the presence of 2 mM of copper by measuring the OD during 48 h (Fig. 5A). Confirming the previous results, ΔhisC had a growth defect. The impaired growth due to copper sensitivity was rescued in C6, ΔhisCSupPA1, and ΔhisC pBBR-oppA2 strains. These results confirmed the suppressor phenotype in C6 and the role of the mutation identified in this strain to cope with copper stress. Indeed, when reinserted in the ΔhisC parent strain, ΔhisCSupPA1, the mutation resolved the growth defect observed due to copper sensitivity. Having oppA2 on a medium-copy vector allowed bacteria to grow as the WT strain in the presence of copper (Fig. 5A).
*Suppressive mutation saves the sensitivity phenotype to copper stress. WT, ΔhisC ΔhisC, C6, ΔhisCSupPA1, pBBR-oppA1, and ΔhisC pBBR-oppA2 strains were tested for their sensitivity to copper toxicity for growth and their intracellular replication capacity. (A) WT, ΔhisC strains, and derivatives were grown in liquid TSB rich medium containing 2 mM of CuSO4. OD600 nm was measured every 30 min for 48 h. Data represent three independent replicates. (B) Experiment performed as in Fig. 1C. The data represent the mean ± SD and were compiled from three independent replicates. Statistical analysis was carried out by a one-way ANOVA followed by Dunnett’s multiple comparisons test (****P <0.0001). (C) Intracellular replication of WT, ΔhisC mutant, and derivatives was assessed by CFU at 2, 5, 24, and 48 h post-infection (PI) of J774.A1 macrophages. Data represent the mean ± SD and were compiled from three independent replicates. Statistical analysis was carried out by a two-way ANOVA followed by Dunnett’s multiple comparisons test (*P<0.05; **P <0.01; **P <0.001).
The soft agar assay was repeated with these strains, and the inhibition zone around the copper well was measured (Fig. 5B). As was evidenced in the case of the liquid growth culture, the C6, ΔhisCSupPA1, and ΔhisC pBBR-oppA2 strains exhibited an inhibition zone, akin to that observed in the WT. In contrast, ΔhisC and ΔhisC pBBR-oppA1 both displayed a copper sensitivity.
As an intracellular pathogen, B. abortus is subject to various stresses within the cell, including possibly copper intoxication induced by the host cell. We examined the ability of the ΔhisC strain and its different derivatives to replicate intracellularly. This was assessed by counting the number of CFUs after J774A.1 macrophage infection. The intracellular trafficking of the different strains was monitored at two early time points (2 and 5 h PI) and two late time points (24 and 48 h PI) following replication (Fig. 5C). The WT strain replicated after 24 h PI (positive control). In contrast, the mutant strain deleted for the entire virB operon was used as a negative control, since it is known to remain blocked in eBCVs (7, 8) and therefore was unable to replicate in macrophages. At 24 h PI, the hisC mutant appeared to have a weaker replication capacity than the WT. However, by 48 h PI, the CFUs increased and approached the WT level. The C6 suppressor had a replication profile that was slightly higher than that of the hisC mutant. Meanwhile, ΔhisC pBBR-oppA1 and ΔhisC pBBR-oppA2 replicated similarly to ΔhisC. The strain in which the suppressor mutation was reintroduced, ΔhisCSupPA1, appeared to be more affected than the hisC mutant itself, particularly at 48 h. Compared to the virB mutant, the hisC mutant and its derivatives could replicate within the cell but to a lesser extent than the WT (Fig. 5C).
DISCUSSION
Copper is an essential transition metal for microorganisms, yet it becomes rapidly toxic at slightly elevated concentrations (45, 46). In E. coli, the total intracellular copper pool (free, labile, and sequestered) is around 10^4^ atoms (~10 µM), where the free cuprous ion amount is at zeptomolar level, thus less than one copper atom per cell (15, 28). This implies strong buffering systems, likely mediated by cysteine-rich proteins (15, 20, 45). Metal ions are critical in numerous mechanisms, and their controlled availability is therefore important in the host-pathogen interplay as well. During intracellular pathogenic infection, host cells both deprive pathogens of essential metals such as iron, zinc, or manganese and also exploit copper toxicity as antimicrobial defense, a strategy known as nutritional immunity (13, 14, 16, 17). Notably, IFN-γ-activated macrophages upregulate the copper transporter ATP7A and relocalize it into the phagolysosome membrane to poison invading microbes by pumping copper into the pathogen-containing vacuole (13, 24). Resistance to copper stress is a determinant for virulence in intravacuolar bacteria such as M. tuberculosis (25, 26) and S. enterica (27). Understanding how Brucella holds copper homeostasis may be a key to a better understanding of its mechanisms of pathogenicity. In the absence of knowledge regarding the way Brucella copes with copper stress, the objective of the present study was to identify potential key actors involved in copper homeostasis through a transposon insertion analysis.
Genes involved in histidine biosynthesis emerged as critical for copper resistance. In this work, the role of this synthesis pathway was therefore investigated through the construction of hisA, hisB, hisC, and hisD deletion mutants. Brucella became auxotrophic for histidine and markedly sensitive to copper, as demonstrated in growth curves and soft agar assays with the loss of one of these genes (Fig. 2). Given the ability of histidine to coordinate metal ions through its imidazole ring, this observation suggests that histidine, free or ligated to a tRNA, might play a role of buffer during copper stress. Our findings, therefore, revealed an unexpected link between histidine biosynthesis and copper detoxification in B. abortus. The copper sensitivity phenotype does not result from a defect in histidine uptake, as supplementation of the minimal medium with histidine restores prototrophy, indicating that the bacterium can import histidine from the extracellular environment (Fig. S4). It has been demonstrated that, among a variety of organisms, histidine performs a role in metal resistance or as a buffer. In Caenorhabditis elegans, disruption of the histidine ammonia lyase (HALY-1) or dietary histidine supplementation results in an increase in zinc and nickel resistance, possibly through histidine’s capacity to chelate zinc and nickel (47). In Aspergillus fumigatus, hisB deletion decreases resistance to several metals, which can be rescued by histidine supplementation. During iron starvation, an increase in histidine content was observed in this pathogenic fungus (48). Likewise, in Acinetobacter baumannii, HutH, the HALY, increases zinc availability by degrading histidine, which under normal conditions stores zinc in a labile histidine-zinc complex. This finding provides direct evidence for histidine-based buffering mechanisms (49). To support this connection between histidine biosynthesis and metal resistance, a histidine-enrichment analysis of the Brucella predicted proteome was performed, and the results indicated that the majority of proteins exhibiting a histidine percentage exceeding 5% are proteins implicated in metal homeostasis or using metals as cofactors (see Table S2). From this enrichment list, only CueO, the multicopper oxidase, was identified in the Tn-seq analysis performed in the presence of copper, suggesting that histidine-rich proteins cannot buffer copper in a non-redundant manner.
Whole-genome sequencing of the suppressor mutants revealed that, among the suppressors, 15 unique mutations were localized to the opp operon that encodes an ATP-binding cassette transporter, and more specifically, the first two genes encoding the periplasmic substrate-binding proteins OppA1 and OppA2. It is striking to observe that no mutations appeared in the permeases oppB and oppC or the ATPase components, oppD/F. At least eight opp operons are encoded in B. abortus genomes, including seven on the second chromosome. Among all of them, only one was affected by suppressive mutations within an impaired histidine biosynthesis background, highlighting its specificity in rescuing from copper resistance when histidine biosynthesis is lacking. In this study, we demonstrated that when histidine biosynthesis is impaired, and the bacteria have to face a copper stress, mutating the two substrate-binding protein genes, oppA1 and oppA2, generates an efficient compensatory mechanism. The mutation in the C6 suppressor is of particular interest due to its location within the promoter region of oppA1. The C6 suppressor and ΔhisCSupPA1 strain, in which the mutation was reintroduced into the ΔhisC background, were confirmed to display a suppressor phenotype by rescuing growth in the presence of copper in comparison to the parent mutant (Fig. 5A and B). One non-exclusive hypothesis to account for this rescue is that OppA1 and/or OppA2 could facilitate increased histidine uptake, either as free amino acid or within short peptides, thereby enhancing the capacity of the bacterial cytosol to buffer copper ions. In this scenario, the C6 suppressive mutation may lead to oppA overexpression. This is consistent with the results obtained with the oppA1 or oppA2 overexpression strains, where increasing the copy number of the second oppA2 gene rescued the copper sensitivity (Fig. 5A and B). Increasing the copy number of oppA1 did not generate the same suppression, maybe because the overexpression of oppA1 is not achieved in this strain. Nevertheless, the suppressive mutations did not resolve histidine auxotrophy, which makes sense in the absence of histidine or histidine-containing peptides in the medium, if the uptake hypothesis proposed above is true (see Fig. S4). Since the attenuation of hisC mutants in infection can be either due to copper sensitivity or histidine auxotrophy, the suppressor strains offer a way to discriminate between these two hypotheses, since the suppressors are copper resistant but still histidine auxotrophic. As indicated in Fig. 5C, the suppressor mutation in the C6 strain does solve the intracellular replication problem of the hisC mutant, while the suppressive mutation inserted in the hisC mutant background does not. These results thus demonstrated that within the cell, the replication of his mutants is impacted due to a histidine biosynthesis dysfunction rather than a copper stress.
Amino acid starvation is a common feature of host-pathogen interactions. It can be imposed by the host as a nutritional immunity strategy to restrict pathogen growth, or alternatively, it may result from the infection by the pathogen. The source of amino acids for B. abortus in host cells is unknown. It may involve extracellular proteases or the hijacking of host proteases to gain small peptides that could fit into the pore of the major porin of B. abortus. To the best of our knowledge, such proteases have not been identified so far, and the availability of free amino acids in the rBCV of macrophages is unknown. Such lines of research would deserve an in-depth investigation in the future. Therefore, why is histidine the only amino acid whose biosynthesis would be required in macrophage infection? It is notorious that histidine biosynthesis is a costly amino acid to synthesize in terms of the ATP equivalents needed (50). It is therefore possible that the available quantities of histidine in the host cells are a limiting factor for growth. It has been demonstrated that M. tuberculosis depends on de novo histidine biosynthesis to overcome host-imposed histidine deprivation. IFN-γ-mediated upregulation of histidine catabolizing enzymes (HAL, HDC) reduces free histidine in infected tissues, creating a nutritional stress that M. tuberculosis ΔhisD mutant cannot withstand. These results highlight the histidine biosynthetic pathway as a critical bacterial strategy in Mycobacterium to evade host nutritional immunity and sustain intracellular survival (51). In macrophages, S. Typhimurium also relies on histidine biosynthesis genes upregulation to counter free histidine limitation in host cells (52). These studies demonstrate that histidine production is an important adaptation for survival under host-imposed nutrient restriction.
As expected, the results highlighted three genes within the cue operon: copA, cueO, and cueR as important for copper resistance in vitro. In addition, the copA transposon mutant was attenuated in macrophages in a previous Tn-seq from our lab (36). Unexpectedly, however, targeted deletions of copA or cueO did not confer a sensitivity phenotype to copper in our conditions in B. abortus (Fig. 1). These results remained consistent when copper concentration was increased. These findings are in striking contrast with observations in B. melitensis, where the bmcO (Brucella multicopper oxidase) mutant displays a copper sensitivity on minimal medium (53), which could be explained by differences related to the bacterial strain context and culture conditions. Also, in E. coli and S. Typhimurium, the cue system is the primary defense line to cope with copper toxicity (28). Indeed, the deletion of copA or cueO increased copper sensitivity, even stronger under anaerobic conditions for ΔcopA (31). In S. Typhimurium, a similar phenotype has been reported where the loss of copA (34) or cueO (27) also gives a copper susceptibility. Likewise, the cueO deletion sensitivity phenotype is exacerbated in the absence of oxygen (27). A homolog of cueO called mmcO also plays a role in copper resistance in M. tuberculosis (54). E. coli and S. Typhimurium have other defense systems, including cus and gol systems, respectively, expressed under anaerobic conditions or higher copper concentrations (28, 31). Additional copper homeostasis systems are absent in the Brucella genome, according to our current knowledge. The role of CopA and CueO was also investigated in S. Typhimurium replication capacity within the host. While CopA is described as having a role in macrophage replication (34), CueO was more important in the mouse model (27). This demonstrated the importance of copper homeostasis in S. Typhimurium for its virulence. Despite the lack of sensitivity to copper in vitro, it would be interesting to test the two mutants constructed in B. abortus during macrophage or mice infection to define their role in a more complex virulence model. Even more so, as copA was identified as required for survival in macrophages (36). It would also be interesting to repeat our experiments in different conditions, such as minimal media. Nevertheless, despite the indications of Tn-seq experiments, the cue system does not appear to be the primary strategy used by Brucella, at least under the conditions tested in this work. This could also demonstrate the limitations of Tn-seq analysis in predicting deletion mutant phenotypes.
Our study reveals an intriguing new aspect of copper homeostasis in B. abortus. While key copper-associated proteins such as CopA and CueO are critical in other bacteria, they appear surprisingly dispensable in B. abortus. Instead, histidine biosynthesis emerges as a possible frontline defense against copper stress. Indeed, we found that disruption of histidine synthesis leads to copper sensitivity in B. abortus, highlighting the importance of this pathway. Interestingly, the attenuation of hisC mutant in macrophages stems from histidine auxotrophy rather than copper sensitivity. The mechanism underlying nutrient uptake within the rBCV remains poorly understood, representing a promising avenue for future research into how B. abortus acquires amino acids in its intracellular replicative niche.
MATERIALS AND METHODS
Bacterial strains and media
Escherichia coli DH10B (Invitrogen) and S17-1 (55) strains were grown in Luria-Bertani (LB Lennox) medium at 37°C. E. coli MFDpir pXMCS2-Tn5 (36) was grown in LB medium supplemented with 300 µM *meso-*2,6-diaminopimelic acid. Brucella abortus 544 Nal^R^ (referred to as the WT in this work; J-M. Verger, INRA, Tours) and its derivative strains were grown in 3% Bacto tryptic soy broth rich medium (TSB; Difco ref. 211825) at 37°C. For auxotrophy experiments, the defined medium Plommet erythritol (PE) (Plommet 1991) was used, composed of 7 or 9.2 g/L K_2_HPO_4_, 3 g/L KH_2_PO_4_, 0.1 g/L Na_2_S_2_O_3_, 5 g/L NaCl, 0.2 mg/L nicotinic acid, 0.2 mg/L thiamine, 0.04 mg/L pantothenic acid, 0.01 g/L MgSO_4_, 0.01 mg/L MnSO_4_, 0.1 mg/L FeSO_4_, 0.1 μg/L biotin, and 2 g/L erythritol. All the strains used in this study are listed in Table S5.
When it was necessary, the culture medium was supplemented with the appropriate antibiotics at the following concentrations: kanamycin (Kan, 10 or 50 µg/mL for chromosomal locus or plasmid selection, respectively) and nalidixic acid (Nal, 25 µg/mL).
Strain construction
For deletion mutants, the whole gene was removed by homologous recombination. Joined PCR was applied. Two regions (the upstream and downstream regions of the target gene) of more than 500 base pairs were amplified from purified B. abortus 544 gDNA using Q5 High-Fidelity DNA Polymerase (New England Biolabs) and primer pairs (F1/R1 for the upstream region and F2/R2 for the downstream region). The two amplified fragments were fused together through a complementary region designed in the primers used, and the resulting fragment was amplified by PCR using F1 and R2 primers. To ΔhisCSupPA1 construction, the complementary region primer pair contained the desired SNP. The amplicon was purified and inserted into an EcoRV (New England Biolabs)-linearized pNPTS138 plasmid through an overnight ligation (T4 DNA ligase, Promega) at 20°C. The ligation product was transformed in DH10B E. coli, and clones positive in blue-white screening were screened by PCR using GoTaQ DNA polymerase (Promega). The selected plasmid was purified and checked by sequencing. The plasmid was inserted into S17-1 E. coli to allow conjugation to B. abortus 544 Nal^R^ by mating. Allelic exchange on the chromosome occurs via the non-replicative plasmid, pNTPS138 (56). The gene deletion was then verified by PCR using GoTaQ DNA polymerase (Promega). ΔhisA, ΔhisB, ΔhisC, and ΔhisD deletion strains were complemented with pMR10 carrying the deleted gene. Complementation plasmids were obtained by amplification of the genes with a region of 400 base pairs upstream of the coding sequence by PCR using Q5 High-Fidelity DNA Polymerase (New England Biolabs). The PCR product was digested as well as the pMR10 plasmid with BamHI and XbaI enzymes (New England Biolabs). The insert was inserted in the linearized plasmid through ligation in the same direction as the E. coli lac promoter of the vector.
To construct the overexpression strains, the genes were amplified by regular PCR using Q5 High-Fidelity DNA Polymerase (New England Biolabs). The designed primers included restriction sites for KpnI and SacI restriction enzymes (New England Biolabs). PCR product was purified and restricted by KpnI and SacI as well as the multicopy-plasmid pBBR-MCS2 (56). The gene expression is under the control of the E. coli lac promoter.
All the primers and plasmids used in this study are listed in Table S3 and S4, respectively.
Transposon sequencing assay
One milliliter of an overnight culture of B. abortus 544 Nal^R^ and 50 µL of an overnight culture of E. coli MFDpir pXMCS2-Tn5 Kan^R^ were mixed. After recovery in 1 mL of TSB supplemented with meso-2,6-diaminopimelic acid (300 µM), OD of the cultures was measured and normalized to an OD of 1. Using a 96-well plate, serial 10-fold dilutions were carried out. Fifteen microliters of these dilutions were plated on TSB plates supplemented with CuSO4 concentrations ranging from 0.5 mM to 3 mM. CFUs were counted to determine the optimal concentration of copper. A CuSO_4_ concentration of 2 mM was selected. The mating was repeated as described above, and plates were incubated overnight at RT. To ensure complete coverage of the genome, 8 B. abortus and E. coli matings were done in parallel. The matings were recovered in 1 mL of TSB from which 50 µL were diluted in 450 µL of TSB. One hundred microliters of the dilution were spread on TSB agar plates supplemented with kanamycin or kanamycin and CuSO_4_ to allow the selection of the resulting B. abortus mini-Tn5 libraries. After 4 days of incubation, the B. abortus mutants were recovered in 2 mL of TSB. The bacterial suspension was centrifuged for 7 min at 7,000 RPM. The pellet was resuspended in 300 µL of 2% SDS. Bacteria were inactivated for at least 1 h at 80°C. All the samples were pooled together following the two conditions. gDNA was extracted using the Nucleobond AXG 500 kit (Macherey-Nagel). This gDNA was sequenced via Illumina sequencing. The raw data were analyzed by an automatic process, TnBox (https://github.com/fxstubbe/TnBox), following a previously described method (37). TnBox uses the BWA method (57) to map the raw reads from Illumina sequencing on the genome of B. abortus 544 and the SAMtools suite (58) to calculate read counts. The libraries underwent normalization based on read depth. For a detailed protocol, see reference (37). From the library, 4.8 × 10^6^ random mutants were recovered and sequenced using a deep sequencing method. We identified 1,684,839 and 1,743,143 unique insertion sites for control and copper conditions, respectively, illustrating genome saturation with a unique insertion every 1.95 bp for control and every 1.89 bp for the stress condition, on average. To evaluate the contribution of each gene to Brucella fitness under copper stress, a TnIF was calculated. The TnIF was defined as the average log_10_(r + 1)/l, where r is the number of miniTn5 insertions at a given nucleotide and l is the coding sequence length in base pairs. To minimize insertional bias, the calculation was performed considering only the central 80% of the coding sequence. For each gene, the differential TnIF (ΔTnIF) between the interest and control conditions was calculated as ΔTnIF = TnIFcopper – TnIFcontrol. A negative ΔTnIF indicates that the gene is required in the copper condition, as the number of reads associated is lower than in the control condition.
Growth curves
Growth was assessed by using BioTek Epoch2 Microplate Reader by measuring OD_600 nm_ every 30 min for 48 h at 37°C with agitation. Culture of B. abortus 544 and derivative strains was grown in TSB overnight to reach the exponential phase (OD_600 nm_ 0.3–0.8) and normalized at an OD_600 nm_ of 0.1 final the following day. For the growth measurement in the presence of copper, TSB was supplemented with 1.6 or 2 mM of CuSO_4_. For the growth measurements in PE, overnight bacterial cultures were washed twice in PBS and diluted to an OD_600 nm_ of 0.1 in PE. When necessary, PE was supplemented with 1 mM of histidine (Sigma).
Suppressor assays
Culture of B. abortus 544 and derivative strains were grown in TSB overnight to reach the exponential phase (OD_600nm_ 0.3–0.8) and normalized at an OD_600 nm_ of 0.1. Twenty microliters of each strain was spread on a TSB agar plate containing 1.6 or 2 mM CuSO_4_. Plates were incubated at 37°C for 4–5 days.
For all four Δhis mutants, 29 isolated colonies formed on CuSO4 plates were streaked on new TSB agar 1.6 and 2 mM CuSO4 plates to confirm the phenotype of the suppressors. Among the 29 clones, 21 were still resistant to copper on the new plates and were grown overnight and stored. To extract demonic DNA, bacterial cultures of around 7.5 mL were centrifuged at 8,200 × g for 5 min. Supernatants were discarded, and bacterial pellets were resuspended in 300 μL of PBS. Then, bacteria were inactivated at 80°C for at least 1 h. Afterward, 100 μL of SDS 10% was added. The gDNA extraction was achieved following the manufacturer’s instructions of the Macherey-Nagel NucleoSpin Tissue Kit. Bacterial SNP sequencing was performed by BIO, part of the Pathology and Genetics Institute. The analysis of sequencing reads was conducted utilizing the Snippy tool, which is available on the Galaxy.org website (https://usegalaxy.org/). Each parental mutant and suppressor genome was aligned to the reference B. abortus 544 genome.
Soft agar well diffusion assays
Culture of B. abortus 544 and derivative strains were grown in TSB overnight to reach the exponential phase (OD_600 nm_ 0.3–0.8) and normalized at an OD_600 nm_ of 0.1. Five hundred microliters was added to 4.5 mL of TSB soft agar (0.7% agar) and spread on a 20 mL TSB agar plate. After the culture-soft agar mix had dried, a well was dug in the upper part of a pipette tip and removed. The well was filled with 100 µL of 200 mM CuSO_4_. The plates were incubated for 3–4 days at 37°C upward-facing lid on the first day before being flipped upside down. After incubation, the inhibition zone around the wall was measured in three different directions, and the mean was reported in a graph in millimeters.
J774A.1 macrophage culture and infection
J774A.1 macrophages (ATCC) were cultivated at 37°C in a 5% CO_2_ atmosphere in GlutaMAX-supplemented Dulbecco’s Modified Eagle Medium (DMEM, Gibco) in which 10% of heat-inactivated fetal bovine serum (Gibco) was added. The day before infection, J774A.1 macrophages were seeded in a 24-well plate at a concentration of 1.10^5^ cells/mL. On the day of the infection, overnight cultures of Brucella strains at exponential phase (0.3–0.8) were washed twice in PBS and diluted in GlutaMAX-supplemented DMEM medium at a multiplicity of infection of 50. Bacterial suspensions were added to the cells, and the 24-well plates were centrifuged at 169 × g for 10 min at RT. Infected cells were incubated at 37°C in a 5% CO_2_ atmosphere for 1 h. Cell medium was refreshed and supplemented with 50 µg/mL gentamycin to kill extracellular bacteria. Cells were incubated one extra hour before the media was again refreshed and supplemented with 10 µg/mL gentamycin, to kill bacteria egressed and avoid a re-infection event, for the 46 h left. For CFU counts, at the interested time points (2, 5, 24, or 48 h after infection), cells were washed twice with PBS and incubated with 0.1% Triton X-100 PBS for 10 min at RT before being scratched to be detached. Macrophage lysates were 10-fold diluted in PBS, and specific dilutions (20 µL) were spread onto TSB agar plates. Plates were incubated at 37°C for 5 days, and CFUs were counted. The CFU number (log10) was calculated per mL of lysate and reported in a graph.
Statistical analysis
Statistical analyses via one- or two-way ANOVA with Dunnett’s test were carried out provided by the GraphPad Prism software. Values of P < 0.05 were considered to represent a significant difference.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hördt A, López MG, Meier-Kolthoff JP, Schleuning M, Weinhold L-M, Tindall BJ, Gronow S, Kyrpides NC, Woyke T, Göker M. 2020. Analysis of 1,000+ type-strain genomes substantially improves taxonomic classification of Alphaproteobacteria Front Microbiol 11:468. doi:10.3389/fmicb.2020.0046832373076 PMC 7179689 · doi ↗ · pubmed ↗
- 2González-Espinoza G, Arce-Gorvel V, Mémet S, Gorvel JP. 2021. Brucella: reservoirs and niches in animals and humans. Pathogens 10:186. doi:10.3390/pathogens 1002018633572264 PMC 7915599 · doi ↗ · pubmed ↗
- 3Corbel MJ. 1997. Brucellosis: an overview. Emerg Infect Dis 3:213–221. doi:10.3201/eid 0302.9702199204307 PMC 2627605 · doi ↗ · pubmed ↗
- 4Suárez-Esquivel M, Chaves-Olarte E, Moreno E, Guzmán-Verri C. 2020. Brucella genomics: macro and micro evolution. IJMS 21:7749. doi:10.3390/ijms 2120774933092044 PMC 7589603 · doi ↗ · pubmed ↗
- 5Moreno E, Moriyón I. 2006. The genus Brucella, p 315–456. In Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E (ed), The prokaryotes. Springer, New York.
- 6von Bargen K, Gorvel J-P, Salcedo SP. 2012. Internal affairs: investigating the Brucella intracellular lifestyle. FEMS Microbiol Rev 36:533–562. doi:10.1111/j.1574-6976.2012.00334.x 22373010 · doi ↗ · pubmed ↗
- 7Starr T, Ng TW, Wehrly TD, Knodler LA, Celli J. 2008. Brucella intracellular replication requires trafficking through the late endosomal/lysosomal compartment. Traffic 9:678–694. doi:10.1111/j.1600-0854.2008.00718.x 18266913 · doi ↗ · pubmed ↗
- 8Celli J, de Chastellier C, Franchini D-M, Pizarro-Cerda J, Moreno E, Gorvel J-P. 2003. Brucella evades macrophage killing via Vir B-dependent sustained interactions with the endoplasmic reticulum. J Exp Med 198:545–556. doi:10.1084/jem.2003008812925673 PMC 2194179 · doi ↗ · pubmed ↗