NeuO-mediated O-acetylation of uropathogenic Escherichia coli K1 capsule enhances resistance to phage and neutrophil killing
Lachlan L. Walker, Nguyen Thi Khanh Nhu, Zheng Jie Lian, Kate M. Peters, Mercedes Monteleone, James P. R. Connolly, Mark J. Walker, Brian M. Forde, Kate Schroder, Matthew J. Sweet, Minh-Duy Phan, Mark A. Schembri

TL;DR
This study shows that O-acetylation of the K1 capsule in uropathogenic E. coli helps it resist phage and neutrophil attacks, but makes it more vulnerable to human serum.
Contribution
The study reveals the role of phase-variable O-acetylation in enhancing UPEC survival against phage and neutrophils.
Findings
NeuO-mediated O-acetylation increases resistance to lytic K1 phage and human neutrophils.
O-acetylation increases susceptibility to human serum.
The neuO gene is prevalent in the pandemic ST95 UPEC clone.
Abstract
Uropathogenic Escherichia coli (UPEC) strains that express the K1 capsule are associated with severe invasive disease, including pyelonephritis, urosepsis, and neonatal meningitis. The K1 capsule can be modified by NeuO, a phage-encoded phase-variable O-acetyltransferase. The role of O-acetylation in pathogenesis of K1 UPEC remains to be fully elucidated. Here, we assessed the prevalence of the neuO gene in a K1 E. coli data set comprising 8,659 genomes and observed that 43.5% of genomes harbor neuO, with a high prevalence (88.1%) in the pandemic UPEC clone sequence type (ST) 95. We generated an isogenic ∆neuO mutant in the reference ST95 strain MS7163 and complemented this with a phase-ON locked version of the neuO gene, resulting in constitutive O-acetylation of the K1 capsule. The phase-variable rate of neuO in MS7163 was measured by fragment analysis at 93% phase-ON with 21…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
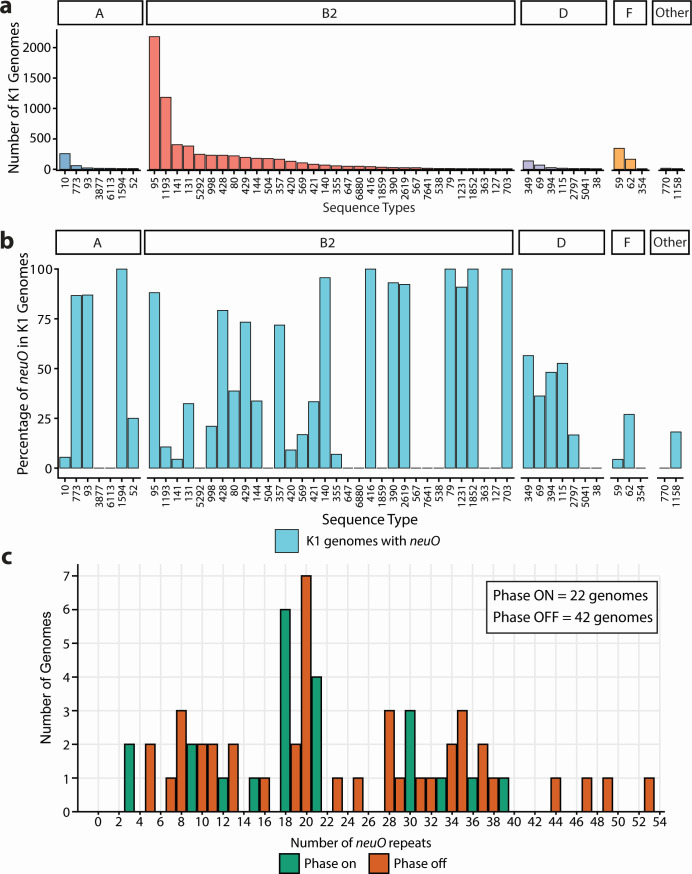 Fig 1
Fig 1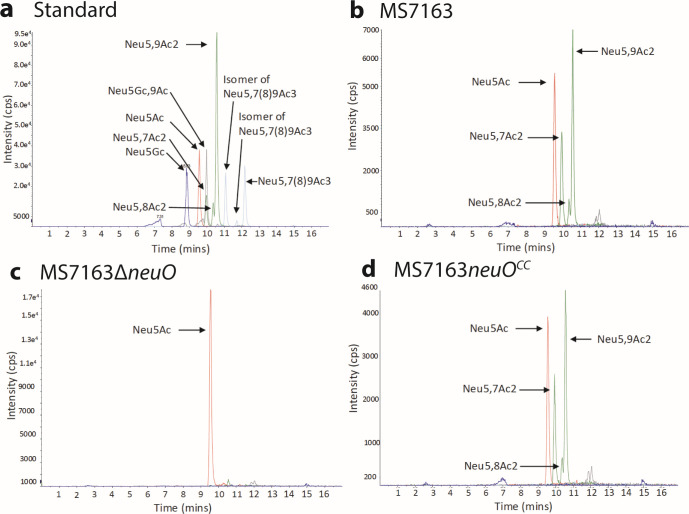 Fig 2
Fig 2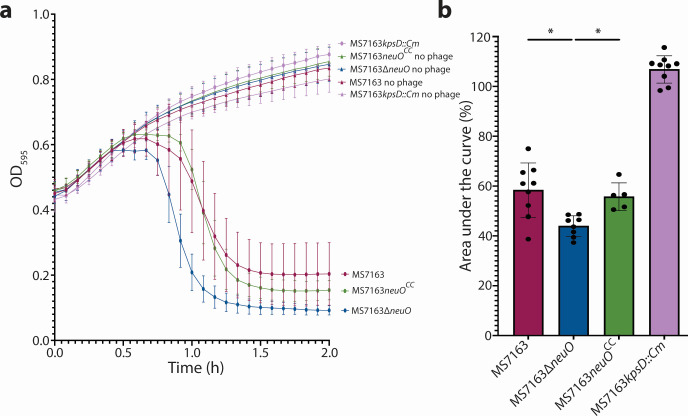 Fig 3
Fig 3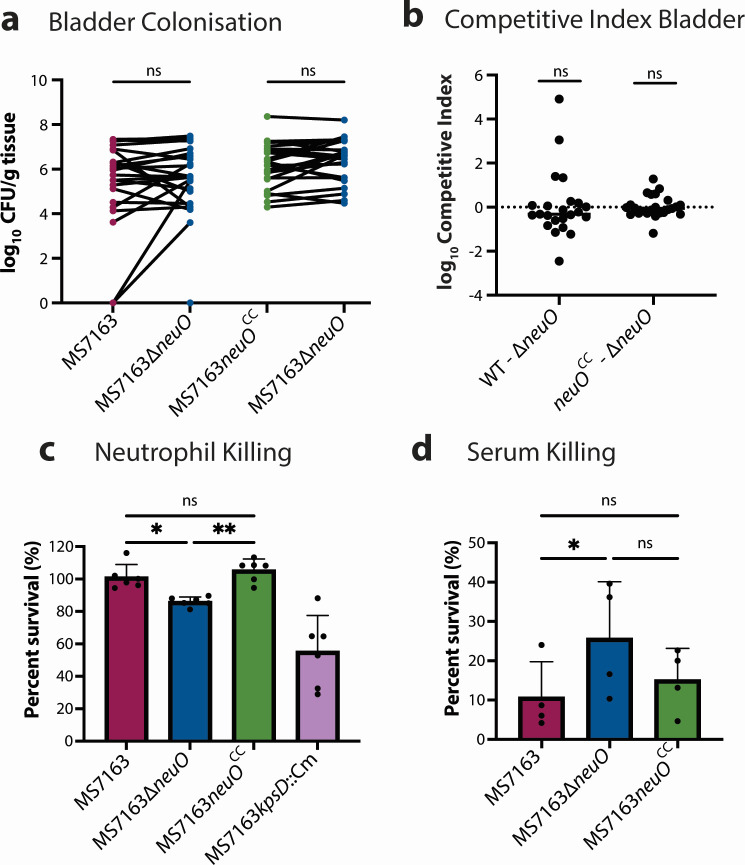 Fig 4
Fig 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Infections and Vaccines · Bacterial Genetics and Biotechnology · Escherichia coli research studies
INTRODUCTION
Urinary tract infections (UTIs) are one of the most common bacterial infections, with an estimated 400 million cases annually across the globe (1). Approximately 75% of all UTIs are caused by uropathogenic Escherichia coli (UPEC) (2). At the severe end of the UTI disease spectrum, acute pyelonephritis can be life-threatening and progress to sepsis. The polysaccharide capsule is an important UPEC virulence factor, and despite its extensive variation, some capsule types are more frequently associated with severe disease. The K1 capsule is particularly significant; longitudinal surveillance studies show that K1-encapsulated E. coli account for ~25% of all bloodstream infections and are associated with invasive diseases, including pyelonephritis and neonatal meningitis (3–6). UPEC strains that express the K1 capsule survive efficiently in human serum and blood (7, 8).
The K1 capsule is an α2,8-linked polysialic acid homopolymer that consists of 5-N-acetylneuraminic acid (Neu5Ac) building blocks (9). Polysialic acid of the K1 capsule can be recognized by host immune receptors, including Siglecs, and O-acetylation of the K1 capsule has been reported to modulate immune recognition (10, 11). O-acetylation is mediated by the phase-variable O-acetyltransferase gene neuO, a cargo gene of the CUS-3 bacteriophage (12–14). Phase variation of neuO is caused by DNA strand slippage of a heptanucleotide tandem repeat (5′-AAGACTC-3′) in the 5′ region of the coding sequence (13). Full-length translation of the protein-coding sequence occurs when the number of repeats is divisible by 3 (13).
The role of neuO in the pathogenesis of K1 E. coli remains to be fully elucidated. Some reports suggest that isolates with an O-acetylated K1 capsule have increased virulence and enhanced immune evasion in both in vitro and in vivo models (11, 15). In contrast, other studies have shown that O-acetylation does not provide a selective advantage in blood during in vivo animal studies and does not significantly affect adherence to human brain endothelial cells (16–18). Despite these conflicting reports, population-based studies show that 91% of human invasive E. coli K1 encapsulated strains and 43% of K1 encapsulated UTI strains harbor the neuO gene (19). Notably, 98% of sequence type complex 95 strains harbor neuO (19). Sequence type (ST) 95 is one of several major UPEC pandemic clones, including ST131, ST73, and ST69 (20). Given the broad conservation of neuO within the ST95 lineage, we aimed to explore if O-acetylation of the K1 capsule contributes to the success of this globally disseminated clone.
In this study, we analyzed a large collection of 8,659 publicly available E. coli genomes that contained the K1 capsule locus, which revealed 43.5% harbor the neuO gene. Molecular analysis demonstrated NeuO-mediated O-acetylation of the K1 capsule provides protection against lytic K1 phage. In terms of UPEC virulence, NeuO-mediated O-acetylation resulted in variable phenotypes; O-acetylation provided protection against killing by human neutrophils, yet increased susceptibility to human serum and did not alter bladder colonization in an experimental mouse model. This suggests that phase variation of neuO provides niche-specific advantages for K1 E. coli.
MATERIALS AND METHODS
Bioinformatic analyses
The data set consisting of 8,659 K1 E. coli genomes was identified from 192,234 complete and draft genomes (downloaded 13 December 2022 from the NCBI, Data S1), using Kaptive 3.0 (21) and the K1 capsule locus from the E. coli group 2 and 3 capsule database (22). Kaptive results that were considered “typeable” were reported (Data S1). Sequence types were determined by MLST (23), and genomes with an undefined ST were removed (24). Blastn was used to query this database for the genes of interest (25). To determine the prevalence of neuO, the neuO gene sequence from the MS7163 genome was used (accession number CP026853.1), with the variable 5′ heptanucleotide tandem repeat region excluded (26). The BLASTn results were filtered with the thresholds of >90% identity and >80% alignment length. Phylogroup labeling for STs was performed with data available on Enterobase (27).
Bacterial strains and growth conditions
Bacteria were grown at 37°C on solid or in liquid lysogeny broth (LB) medium. Media were supplemented with chloramphenicol (30 μg/mL), kanamycin (50 μg/mL), or gentamicin (20 μg/mL) when appropriate. Bacterial strains used in this study are described in Table S1.
Molecular methods
DNA extraction, purification, and Sanger sequencing were performed as previously described (28, 29). Primers used in this study are listed in Table S2. Fragment analysis was performed by the Genetic Research Services at the University of Queensland. Briefly, a primer was labeled with a 5′ 6-carboxyfluorescein (6-FAM) fluorophore. The subsequent PCR reaction labeled the PCR products with a 5′ 6-FAM fluorophore, and these samples were loaded onto the 3730 DNA analyzer (Thermo Fisher Scientific), which allowed for gel capillary electrophoresis to differentiate the sizes of the PCR product.
Generation of isogenic mutant strains
The λ-Red recombinase system was used to generate a neuO mutant and to undertake subsequent chromosome complementation in MS7163 (30). To mutate the neuO gene, we performed a three-way PCR that involved fusing ~500 bp PCR products with overhangs from the upstream and downstream of the neuO gene to the chloramphenicol resistance gene cassette isolated from the pKD3 vector following a previously described method (31). The complement plasmid, pSU2718-pNeuO (pNeuO), carrying a neuO gene with a codon optimized 5′ region replacing the tandem repeats to prevent DNA slippage, resulting in a non-phase-variable (i.e., always ON) construct and a total of 39 heptanucleotide tandem repeats, was synthesized by Epoch Life Science, Inc. (Texas, USA). To perform chromosome complementation, a construct that contained the non-phase-variable version of neuO fused to the chloramphenicol cassette from the pKD3 plasmid was used. The non-phase-variable version of neuO and the chloramphenicol cassette were flanked by ~500 bp of upstream and downstream regions of the neuO gene using PCR as described (31). The resulting complementation construct was electroporated into MS7163∆neuO, followed by removal of the chloramphenicol cassette via the FLP recombinase system (30), resulting in MS7163neuO^CC^.
Mass spectrometry of K1 capsule O-acetylation
K1 capsular sialic acid was extracted from overnight cultures and prepared using Signal DMB Labeling Kit (Agilent) (11). Time of flight mass spectrometry was performed on a Shimadzu Nexera uHPLC (Japan) at The University of Queensland Institute for Molecular Bioscience Mass Spectrometry facility.
K1 phage killing assay
The K1 lytic phage vB_EcoM_SHAK7163 was used (32). Overnight bacterial cultures were standardized to an OD_600_ = 1 (~10^9^ colony-forming units [CFU]/mL) and infected with phage (2 × 10^8^ PFU/mL) at a multiplicity of infection (MOI) of 1:10 (phage:bacteria). Growth of bacteria was measured at 37°C with shaking at 200 rpm using a FLUOstar OPTIMA microplate reader (BMG Labtech).
Neutrophil killing assay
Blood from a single donor was collected across multiple days using Vacuette tubes containing EDTA (Greiner Bio-One). Neutrophils were isolated using the EasySep Direct Human Neutrophil Isolation kit (EasySep). Purified neutrophils (~2.5 × 10^5^) were mixed with bacteria (2.5 × 10^6^ CFU) at an MOI of 1:10 at 37°C with shaking at 50 rpm and enumerated to determine CFU prior to incubation with neutrophils (time 0) and at 1 h post-incubation.
Serum killing assay
Fresh blood from a single donor was collected using BD Vacutainer SST II Advanced tubes (BD). Isolated human serum (90 µL) was inoculated with bacteria (10 µL) at ~10^7^ CFU and incubated at 37°C for 1 h with shaking at 50 rpm. Bacteria were enumerated to determine CFU prior to incubation with human serum (time 0) and at 1 hour post-incubation.
Mouse UTI model
All experiments were performed using the following housing conditions: (light:dark cycle 12:12 h, room temperature 21 ± 1°C, humidity 50 ± 10%). The mouse model of UTI was employed as previously described (33, 34). Briefly, female C57BL/6 mice (8–10 weeks) were infected via transurethral administration at 5 × 10^8^ CFU. Mice were euthanized by cervical dislocation at 24 h post-infection. Bacterial bladder loads were enumerated by plating onto LB agar. Mixed competitive infections were performed using a 1:1 mixture of strains tagged with different antibiotic resistance genes (chloramphenicol or kanamycin). Groups consisted of either wild-type MS7163 and MS7163∆neuO, or MS7163neuO^CC^ and MS7163∆neuO. Statistical analyses were performed using the two-tailed Wilcoxon matched-pairs signed-rank test (Prism 9, GraphPad).
RESULTS
The neuO gene is highly prevalent in the UPEC ST95 clone
We curated a data set of 8,659 K1 genomes using Kaptive 3.0 (21) and screened for the presence of neuO. This analysis identified that the neuO gene is harbored in 43.5% of K1 genomes. To understand the distribution of neuO in K1 E. coli, we determined the STs of all K1 genomes and assessed the carriage of neuO. In total, 255 STs contained at least 1 K1-positive genome, and 51 STs contained 10 or more K1 genomes (Fig. 1a). The majority of STs with more than 10 K1 genomes belonged to phylogroups B2 and D (39/51) that primarily comprise UPEC and other extraintestinal pathogenic E. coli pathotypes (35). The STs with the highest number of K1-positive genomes were ST95 (2,181 genomes) and ST1193 (1,184 genomes); these STs accounted for 25.2% and 13.7% of total K1 genomes in our data set, respectively (Fig. 1a). The carriage of neuO among K1 STs with 10 or more genomes exhibited a variable distribution, ranging from 0% to 100% (Fig. 1b). The neuO gene was present at 85% or more in 12 STs (ST416, ST79, ST1594, ST1852, ST703, ST140, ST390, ST2619, ST1231, ST95, ST93, and ST773) (Fig. 1b). Within this group, ST95 was dominant, with 1,693/1,922 (88.1%) genomes positive for neuO. The remaining 11 STs comprised 10–66 K1 genomes. In contrast, neuO gene carriage varied among other large K1 lineages, including ST1193 (10.6%), ST141 (4.4%), ST131 (32.4%), and ST59 (4.3%). The neuO gene was absent in seven K1-positive STs (Fig. 1b). Overall, this analysis demonstrates that the prevalence of neuO is highly variable among K1 STs and primarily found in E. coli genomes from phylogroups B2 and D.
Prevalence of the neuO gene in 8,659 K1 E. coli genomes. (a) STs that contain at least ten K1 genomes are presented as a column with the y-axis indicating the total number of K1 genomes in each ST. The STs are split by phylogroups, with the phylogroups indicated above the column graph. (b) The percentage of genomes harboring neuO as determined by BLASTn (>90% identity and >80% alignment length thresholds) is shown in cyan. (c) The distribution of neuO repeats in complete genomes that harbored neuO (n = 64). Each column represents the number of repeats identified in each individual genome. The green color represents genomes with the number of repeats divisible by 3, while orange represents genomes not divisible by 3.
Analysis of E. coli K1 complete genomes reveals a phase-variable genotype
To assess the phase status of neuO, we determined the proportion of K1 E. coli strains in the NCBI refseq complete genome database with phase “ON” versus “OFF” neuO genotypes. This data set comprised 64 completely sequenced K1 genomes that harbored neuO. These genomes were probed to identify the number of neuO heptanucleotide tandem repeats, revealing a range from 3 to 53 repeats, with 20 repeats being the most common and found in seven genomes (Fig. 1c). Of the 64 genomes that harbored neuO, 65.6% (42 genomes) were not in a multiple of three based upon the repeat region, indicative of a phase-OFF genotype (Fig. 1c).
O-acetylation of the K1 capsule by NeuO confirmed by mass spectrometry
To dissect the role of neuO in O-acetylation of the K1 capsule, we utilized the ST95 UPEC strain MS7163. MS7163, which was isolated from a patient with acute pyelonephritis, belongs to the highly virulent neonatal meningitis-associated clone, O45:K1:H7 (5, 26). Long-read sequencing of MS7163 showed neuO was primarily in the phase-ON state with 21 heptanucleotide repeats, which was also verified by Sanger sequencing of the repeat region. To determine the phase status of MS7163 cells, we performed fragment analysis on four colonies, which revealed that 93% of the MS7163 cells contained 21 repeats, while the remainder contained 20 (1.92%) and 22 (5.35%) repeats, respectively. Thus, the majority of the MS7163 population is neuO phase-ON during laboratory culture in LB broth.
Next, we mutated the neuO gene in MS7163 using λ-Red recombination to generate the strain MS7163∆neuO. We also generated a chromosomally complemented neuO phase-ON locked strain (referred to as MS7163neuO^CC^). This strain was constructed by codon-optimizing the 5′ repeat region of the neuO gene to abolish the tandem repeat pattern (thereby preventing DNA strand slippage) and increase the size of the repeat region to 39 heptanucleotide tandem repeats (in line with previous literature indicating the length of the neuO repeat region increases the catalytic activity of NeuO [36]). To confirm the O-acetylation status of the K1 capsule in these isogenic strains, we used mass spectrometry with known O-acetylated sialic acid standards (Fig. 2a). The capsule of wild-type MS7163 exhibited peaks for Neu5Ac (sialic acid) as well as the O-acetylated forms of sialic acid: Neu5,7Ac2, Neu5,8Ac2, and Neu5,9Ac2 (Fig. 2b). MS7163∆neuO did not exhibit O-acetylation of the capsule, demonstrated by the single peak for Neu5Ac (Fig. 2c), while the MS7163neuO^CC^ strain possessed an O-acetylation pattern consistent with successful complementation (Fig. 2d).
Mass spectrometry of K1 capsule O-acetylation. (a) Standards for the DMB sialic acid labeling kit (Agilent) with the corresponding peaks labeled. The subsequent panels represent mass spectrometry data for (b) wild-type MS7163, (c) MS7163∆neuO, and (d) MS7163neuOCC.
NeuO is important for protection against lytic K1 phage
To investigate the effect of O-acetylation on K1 capsule recognition, we utilized the K1-specific lytic phage vB_EcoM_SHAK7163 (32). Strains MS7163, MS7163∆neuO, and MS7163neuO^CC^ were grown to mid-exponential phase, at which time phage was added to the culture and the optical density was tracked over 2 h. In these experiments, MS7163*∆neuO* was more susceptible to phage lysis compared to the O-acetylated strains MS7163 and MS7163neuO^CC^ (Fig. 3). As a control, an MS7163 strain mutated for capsule production (MS7163kpsD::Cm) was also incubated with phage in the same manner; this mutant was not susceptible to phage lysis, demonstrating the K1 capsule specificity of the phage (Fig. 3). Together, these results support a previous analysis that suggested O-acetylation of the K1 capsule protects K1 E. coli from K1 phage recognition and lysis (16).
*K1-specific lytic phage killing assay. (a) Growth of bacteria over 2 h in LB media with or without vB_EcoM_SHAK7163 phage (no phage control). Data represent the mean ± SD. Data for wild-type MS7163 and MS7163kpsD represent nine biological replicates, while data for MS7163∆neuO and MS7163neuOCC represent eight and five biological replicates, respectively. (b) Area under the curve analysis is presented as a bar graph, representing mean ± SD. GraphPad Prism was used to visualize data and determine statistical significance by Brown-Forsythe and Welch one-way ANOVA. Significance level is indicated by an asterisk: P ≤ 0.05.
O-acetylation does not enhance K1 UPEC bladder colonization
MS7163 has been shown to effectively colonize the bladder of experimentally infected mice (26, 28). To determine if O-acetylation of the K1 capsule impacts UPEC bladder colonization, C57BL/6 mice were transurethrally infected with a 1:1 mixed ratio of an O-acetylation-positive and O-acetylation-negative strain tagged with different antibiotic resistance markers to enable differential selection and comparative enumeration of bacterial loads. Two sets of mixed infections were performed. Groups consisted of wild-type MS7163 and MS7163∆neuO, or MS7163neuO^CC^ and MS7163*∆neuO*. In these experiments, no significant difference in colonization of the bladder was observed for either group (Fig. 4a). This was also reflected by analysis of the competitive index for both groups, which showed no significant difference at 24 h post-infection (Fig. 4b). Together, these results show that O-acetylation of the K1 capsule does not influence UPEC bladder colonization in mice.
*Infection impacts of K1 capsule O-acetylation. The mixed 50:50 infection consisted of two groups of strains (n = 24 mice per group). The first group contained MS7163 (WT) and MS7163∆neuO (∆neuO), and the second group contained MS7163neuOCC (neuOCC) and MS7163∆neuO. (a) Data from the bladder are represented as CFU count. (b) Competitive index that indicates the ratio of CFU between strains in the mixed infection before and after infection. GraphPad Prism was used to visualize the data. To determine statistical significance, the Wilcoxon matched-pair signed-rank test was used for CFU, and the Wilcoxon signed-rank test with a hypothetical mean of 0 was used for CI. (c) Human neutrophil killing assay, with survival of bacteria after 1-h incubation with human neutrophils being plotted as a bar graph. Data represent mean ± SD from six biological replicates, and statistical significance was determined using Brown-Forsythe and Welch ANOVA. (d) Serum killing assay, showing mean percentage survival of bacteria ± SD after 1-h incubation with human serum. Data represent matched data of n = 4 biological replicates. Statistical significance was determined by Friedman’s test with Dunn’s multiple comparisons to account for day-to-day variation in serum activity. Data were visualized in GraphPad Prism. Significance levels are indicated by asterisks: *P ≤ 0.05, *P ≤ 0.01; ns = P > 0.05.
O-acetylation increases K1 UPEC survival against human neutrophils
A hallmark of the mouse UTI model is the rapid influx of neutrophils into the bladder post-infection (37). Neutrophils employ multiple antibacterial mechanisms required for the resolution of UTI, including phagocytosis, the release of antimicrobial peptides, generation of reactive oxygen species, and formation of neutrophil extracellular traps (38–40). Previous studies have shown that the K1 capsule is essential for UPEC protection against neutrophil-mediated killing and contributes to the capacity of UPEC to form intracellular bacterial communities (IBCs) within the bladder uroepithelium (41). Thus, to assess the effect of O-acetylation on neutrophil killing in a controlled experimental system that avoids the complexity of the mouse UTI model, we tested the survival of the isogenic strains MS7163, MS7163∆neuO, and MS7163neuO^CC^ following incubation with human neutrophils. Here, we observed the O-acetylated strains MS7163 and MS7163neuO^CC^ exhibited significantly increased survival compared to the non-O-acetylated strain MS7163*∆neuO* (Fig. 4c). To further evaluate the contribution of neuO-mediated O-acetylation against innate immune killing, we performed a serum bactericidal assay. Here, we observed deletion of the neuO gene (MS7163∆neuO) led to increased survival in human serum compared to O-acetylated wild-type and complemented strains (Fig. 4d). In these experiments, the bactericidal activity of human serum was confirmed by complete (100%) killing of the K-12 MG1655 strain (Fig. S1). Overall, these results demonstrate that O-acetylation of the K1 capsule confers protection against human neutrophil-mediated killing but contrastingly leads to increased susceptibility to human serum. This is consistent with the role of phase variation in promoting different phenotypes.
DISCUSSION
O-acetylation of the K1 polysialic acid capsule in UPEC is controlled by the CUS-3 phage-encoded phase-variable NeuO O-acetyltransferase. By curating a large genomic data set of K1 E. coli, we assessed the prevalence, phase status, and function of the neuO gene. We show the neuO gene is a common feature in the ST95 clone and exists in phase-ON and phase-OFF states, reflecting active phase switching. We further show NeuO-mediated O-acetylation confers diverse phenotypes, ranging from protection against phage and neutrophil killing to increased susceptibility against human serum, revealing its multifaceted role in UPEC survival.
By curating a large collection of K1 E. coli genomes (n = 8,659), we showed the neuO gene is present in 43.5% of K1 E. coli, a finding consistent with a previous study that analyzed a smaller data set (19). The high prevalence of neuO (and thus the CUS-3 phage) in ST95 aligns with its conserved K1 genotype (3, 28), which is estimated to have emerged ~250 years ago (3). Intriguingly, this is in contrast to ST1193, which transitioned from a K5 to K1 capsule type ~30 years ago (42), and contained only a small proportion of K1 genomes carrying neuO (10.6%). This suggests that the pattern of neuO prevalence is associated with CUS-3 phage infection and clonal expansion of successful K1 lineages such as ST95.
Expression of the K1 capsule leads to increased susceptibility to infection and lysis by K1-specific phage (43). K1 phage recognize the Neu5Ac monomers that are the building blocks of the K1 capsule (44). Here, we showed O-acetylation of the K1 capsule reduces susceptibility to a lytic K1 phage, consistent with a previous study that used a different strain-phage combination (16). It appears that NeuO-mediated O-acetylation of the K1 capsule is a mechanism of phage superinfection exclusion, whereby the CUS-3 phage encodes the neuO gene to O-acetylate the K1 capsule, preventing recognition and infection by similar K1-specific bacteriophage. We speculate that O-acetylation of the K1 capsule may also generate novel receptor motifs that newly evolved phage could preferentially recognize. Thus, NeuO-mediated O-acetylation could impact phage-driven ecology in multiple ways, and it remains to be determined how this phenotype may more broadly impact horizontal gene transfer in K1 E. coli.
The impact of NeuO-mediated O-acetylation on virulence is complicated by its phase-variable expression. We addressed this problem by generating a set of isogenic strains, including a neuO mutant (MS7163∆neuO) and a locked neuO phase-ON strain (MS7163neuO^CC^). While mutation of neuO did not alter colonization of the mouse bladder, we showed that expression of NeuO does enhance survival against neutrophil killing. Although neutrophil influx is characteristic of infection of the bladder in experimental mice, ST95 K1 UPEC can resist this innate immune response (41), and thus a more nuanced experimental design was used to investigate the impact of O-acetylation. This involved a direct killing assay employing ex vivo human neutrophils, where we observed a protective effect mediated by K1 O-acetylation. Given the complexity of UTI, including the establishment of IBCs that shield bacteria from innate immune attack (41), this is not surprising and is in line with previous work that showed NeuO-mediated O-acetylation modulates immune evasion of macrophages (11).
The interpretation of these results has some limitations, for example, the design of the mouse UTI experiments and known differences between mouse and human neutrophils (45). First, our experimental design employed mixed 50:50 infections to assess relative survival; however, we note this may mask the contribution of O-acetylation toward neutrophil-mediated immunity. Indeed, since half of the inoculum expresses an O-acetylated capsule, we cannot rule out the possibility that this may be sufficient to alter the immune environment of the bladder in a way that limits our ability to observe differences. Second, O-acetylation of the K1 capsule has been shown to reduce recognition by Siglecs 5/7/11/14 on human macrophages (11), receptors that are not conserved in mice (46). These species-specific differences in Siglec receptors may also contribute to the observed differences between our human neutrophil killing assays and mouse UTI experiments.
We further explored the role of K1 capsule O-acetylation and immune system interaction through a serum killing assay. Our results show that O-acetylation of the K1 capsule increased susceptibility to human serum, despite its protective effect against neutrophil killing. These findings align with a previous study that demonstrated K1 E. coli recovered from the bloodstream of neonatal rats expressed non-O-acetylated polysialic acid, although that study did not directly assess the impact of O-acetylation (17). In contrast, Yang et al. reported enhanced virulence of a serially passaged phase-ON neuO variant compared to a primarily phase-OFF isogenic neuO parent strain in a neonatal mouse infection model (11). Differences in UPEC strains, neuO phase status, and infection models may account for discrepancies between these studies. An independent challenge UTI study comparing the spatial and temporal patterns of colonization of K1 UPEC neuO phase-ON and phase-OFF strains may provide deeper insight into the contribution of K1 capsule O-acetylation in pathogenesis and dissemination to the bloodstream.
In conclusion, our findings indicate that phase variation of neuO enables UPEC to adapt to different niches. The ability to reversibly switch between O-acetylated and non-O-acetylated K1 capsule states is likely to enhance UPEC survival across diverse host and environmental settings.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yang X, Chen H, Zheng Y, Qu S, Wang H, Yi F. 2022. Disease burden and long-term trends of urinary tract infections: a worldwide report. Front Public Health 10:888205. doi:10.3389/fpubh.2022.888205 PMC 936389535968451 · doi ↗ · pubmed ↗
- 2Flores-Mireles AL, Walker JN, Caparon M, Hultgren SJ. 2015. Urinary tract infections: epidemiology, mechanisms of infection and treatment options. Nat Rev Microbiol 13:269–284. doi:10.1038/nrmicro 343225853778 PMC 4457377 · doi ↗ · pubmed ↗
- 3Arredondo-Alonso S, Blundell-Hunter G, Fu Z, Gladstone RA, Fillol-Salom A, Loraine J, Cloutman-Green E, Johnsen PJ, Samuelsen Ø, Pöntinen AK, Cléon F, Chavez-Bueno S, De la Cruz MA, Ares MA, Vongsouvath M, Chmielarczyk A, Horner C, Klein N, Mc Nally A, Reis JN, Penadés JR, Thomson NR, Corander J, Taylor PW, Mc Carthy AJ. 2023. Evolutionary and functional history of the Escherichia coli K 1 capsule. Nat Commun 14:3294. doi:10.1038/s 41467-023-39052-w 37322051 PMC 10272209 · doi ↗ · pubmed ↗
- 4Bonacorsi S, Bingen E. 2005. Molecular epidemiology of Escherichia coli causing neonatal meningitis. Int J Med Microbiol 295:373–381. doi:10.1016/j.ijmm.2005.07.01116238014 · doi ↗ · pubmed ↗
- 5Bonacorsi S, Clermont O, Houdouin V, Cordevant C, Brahimi N, Marecat A, Tinsley C, Nassif X, Lange M, Bingen E. 2003. Molecular analysis and experimental virulence of French and North American Escherichia coli neonatal meningitis isolates: identification of a new virulent clone. J Infect Dis 187:1895–1906. doi:10.1086/37534712792866 · doi ↗ · pubmed ↗
- 6Mårild S, Jodal U, Orskov I, Orskov F, Svanborg Edén C. 1989. Special virulence of the Escherichia coli O 1:K 1:H 7 clone in acute pyelonephritis. J Pediatr 115:40–45. doi:10.1016/s 0022-3476(89)80326-92661790 · doi ↗ · pubmed ↗
- 7Goh KGK, Phan M-D, Forde BM, Chong TM, Yin W-F, Chan K-G, Ulett GC, Sweet MJ, Beatson SA, Schembri MA. 2017. Genome-wide discovery of genes required for capsule production by uropathogenic Escherichia coli. m Bio 8:e 01558-17. doi:10.1128/m Bio.01558-1729066548 PMC 5654933 · doi ↗ · pubmed ↗
- 8Vermeulen C, Cross A, Byrne WR, Zollinger W. 1988. Quantitative relationship between capsular content and killing of K 1-encapsulated Escherichia coli. Infect Immun 56:2723–2730. doi:10.1128/iai.56.10.2723-2730.19883047064 PMC 259635 · doi ↗ · pubmed ↗