Decoding plant defense signaling using the defenseless mutant
Bikash Baral, Mikael Brosché

TL;DR
Scientists created a plant mutant lacking six defense pathways and found that plants can survive without defenses in good conditions, revealing hidden backup systems.
Contribution
The study introduces a novel Arabidopsis mutant lacking six defense pathways to explore redundancy in plant defense networks.
Findings
The defenseless mutant showed no stress phenotypes under optimal conditions, indicating plant defenses are dispensable in favorable environments.
Ozone-triggered ROS signaling remained functional, revealing alternative defense mechanisms.
Transcriptome analysis identified downregulated immune genes, but pathogen susceptibility was not increased beyond known mutants.
Abstract
Can plants live without defenses? Mutant analysis in Arabidopsis thaliana has identified numerous regulators of biotic, abiotic, and hormone‐based defenses, but the redundancy among separate defense pathways remains unexplored.We constructed an Arabidopsis mutant, defenseless, lacking six canonical defense pathways using abi1‐1 (abscisic acid), coi1 (jasmonic acid), sid2 (salicylic acid), ein2 (ethylene), eds1 (biotic defense signaling), and rbohD (apoplastic reactive oxygen species production), enabling dissection of defense network resilience.In optimal growth conditions, defenseless exhibited no stress phenotypes, demonstrating that plant defenses are dispensable under favorable environments. Stress assays revealed paradoxical responses: some defenses remained functional in defenseless, while others were severely compromised. Notably, ozone‐triggered apoplastic ROS signaling was…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
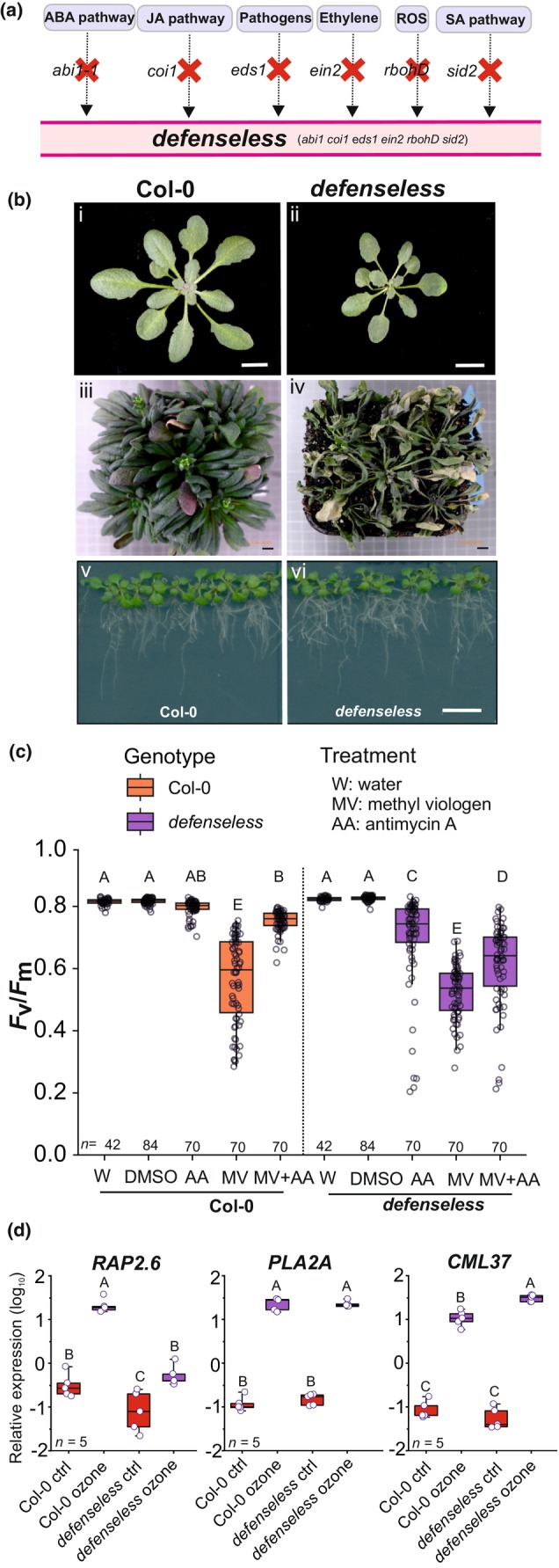 Fig. 1
Fig. 1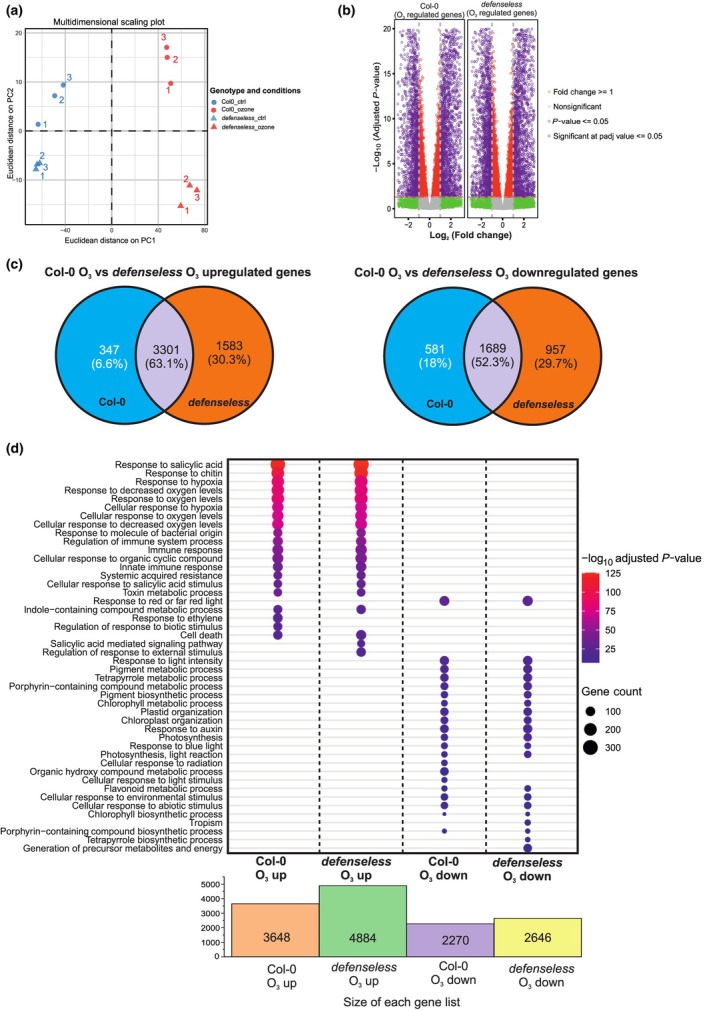 Fig. 2
Fig. 2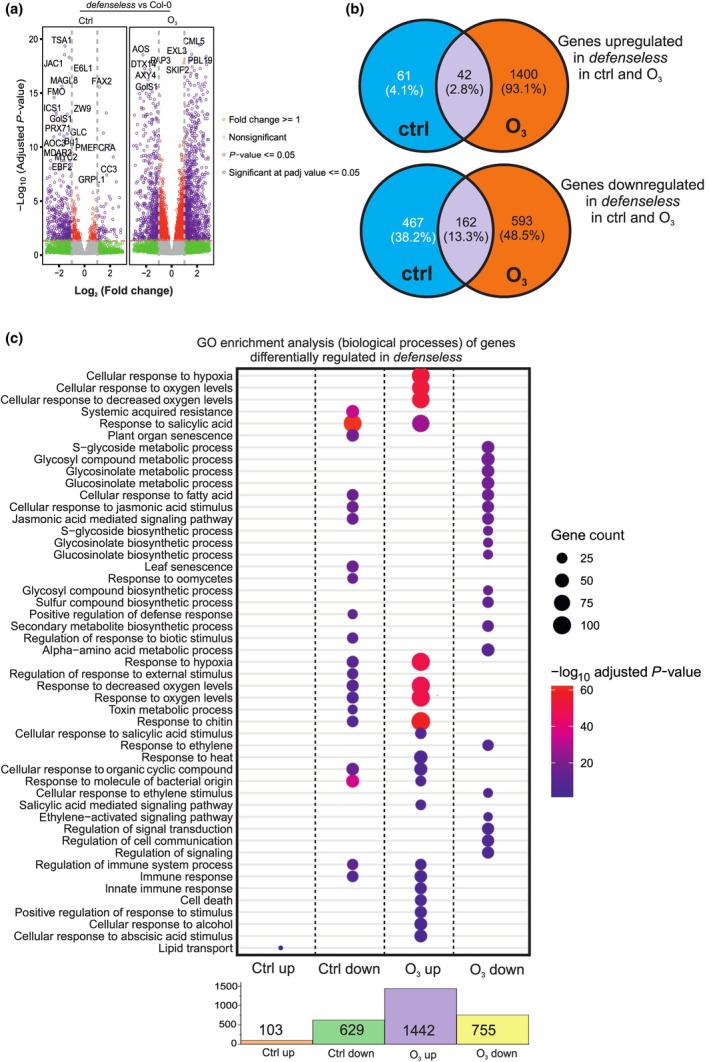 Fig. 3
Fig. 3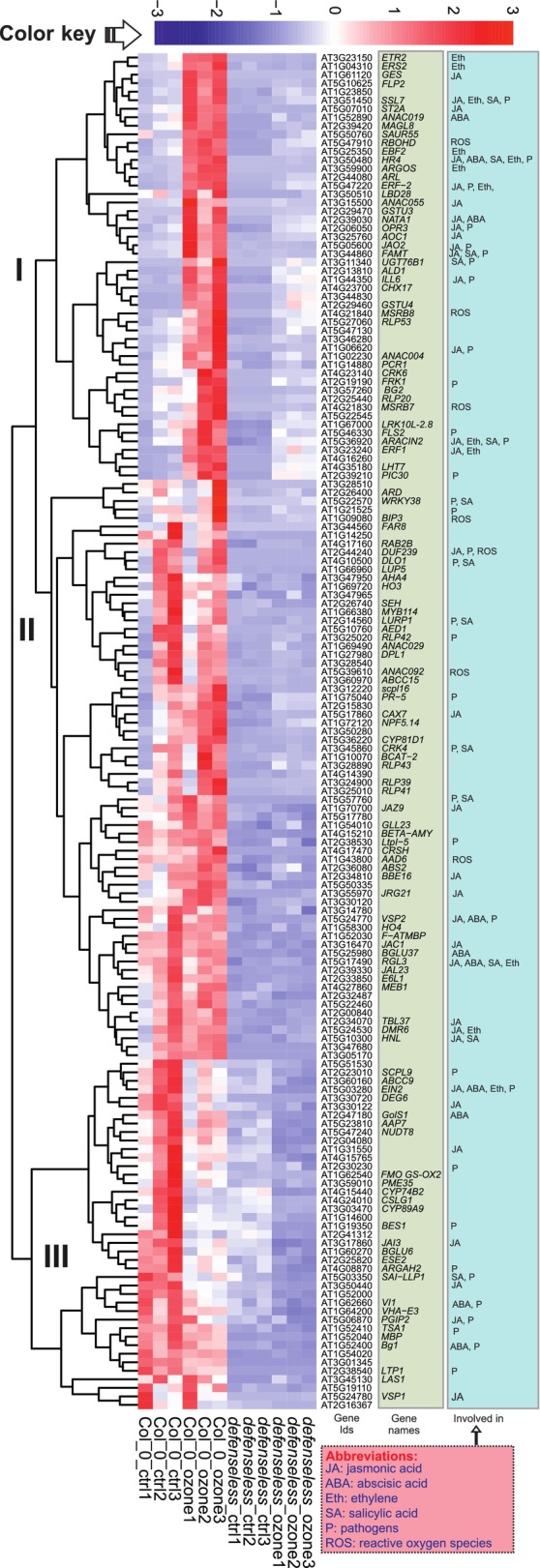 Fig. 4
Fig. 4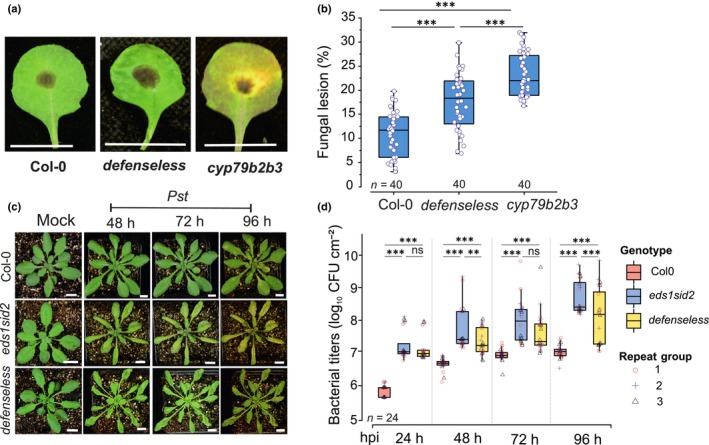 Fig. 5
Fig. 5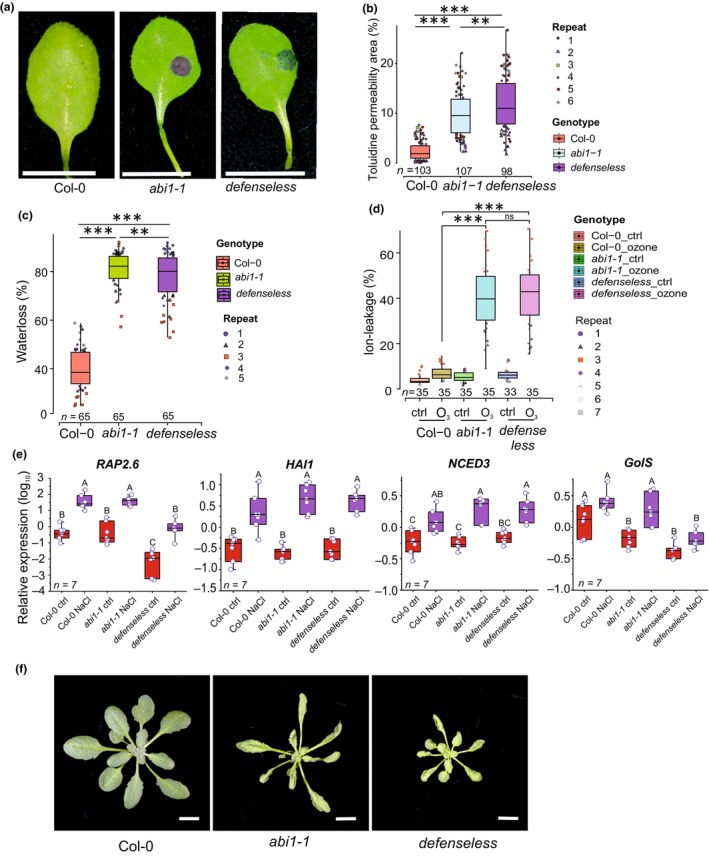 Fig. 6
Fig. 6- —Research Council of Finland10.13039/501100002341
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect-Plant Interactions and Control · Plant-Microbe Interactions and Immunity · Plant Gene Expression Analysis
Introduction
Immunodeficiency renders organisms highly vulnerable to a wide range of pathogens, as demonstrated by the emergence of acquired immunodeficiency syndrome, first identified in humans in the early 1980s (Bosma et al., 1983). Development of mice that lack the adaptive immune system is a key resource to understand disease in animals and humans (Walsh et al., 2017). In plants, protection against invaders relies on complex signaling networks for pathogen recognition and defense activation (Dodds et al., 2024). Plants are constantly exposed to pathogenic threats, both above‐ and belowground (De Coninck et al., 2015). Moreover, plants are also exposed to abiotic stress from changes in the environment, including drought, temperature extremes, air pollutants, and excess light (Zhang et al., 2022).
Plants are effective in combating invading pathogens, and only a fraction of these invaders are successful in causing disease (Wei et al., 2015; Saur & Hückelhoven, 2021). Plant defenses are activated by both biotic and abiotic stressors and rely on several signaling pathways that allow the plant to adapt to different stressors (Zhang et al., 2022; Dodds et al., 2024). Several hormones, jasmonic acid (JA), salicylic acid (SA), abscisic acid (ABA), and ethylene (ET) – play central roles in orchestrating these responses (Xu et al., 2015; Overmyer et al., 2018; Sun & Zhang, 2022; Zhang et al., 2022; Dodds et al., 2024). JA and SA regulate defense against pathogens (Ma & Ma, 2016), while ABA primarily regulates responses to environmental stress (Zhang et al., 2022). Plants detect invaders through pattern‐recognition receptors, which recognize microbe‐associated molecular patterns (MAMPs) and damage‐associated molecular patterns, initiating pattern‐triggered immunity (Dodds et al., 2024). Pathogens combat plant defenses by using effector proteins, where in turn, plants can recognize the effector to activate effector‐triggered immunity (ETI; Remick et al., 2023), which provides a stronger response, sometimes including localized cell death known as the hypersensitive response (Dalio et al., 2021). In the signaling pathway following pathogen perception by surface receptors or intracellular nucleotide‐binding leucine‐rich repeat, the lipase‐like protein ENHANCED DISEASE SUSCEPTIBILITY1 (EDS1) has a key function to link pathogen perception to downstream responses (Dongus & Parker, 2021; Dodds et al., 2024). EDS1 together with interacting partners PHYTOALEXIN DEFICIENT 4 (PAD4) or SENESCENCE‐ASSOCIATED GENE 101 activate defense signaling, programmed cell death and calcium (Ca^2+^) channels. In defense toward abiotic stress, plants rely on various sensing mechanisms, for example changes in turgor pressure that activate Ca^2+^‐channels (Zhang et al., 2022). The sensing mechanisms take place throughout the cell and include cell wall, plasma membrane, chloroplast, and mitochondria (Zhang et al., 2022). After perception of abiotic or biotic stress, downstream signaling components include reactive oxygen species (ROS), Ca^2+^ and kinases (Waszczak et al., 2018; Luan & Wang, 2021; Sun & Zhang, 2022; Dodds et al., 2024). Outputs from signaling pathways include activation of transcription factors and transcriptional reprogramming tailored to combat the specific stress.
ROS include hydrogen peroxide (H_2_O_2_) and superoxide (O_2_ ^·−^), and ROS have several roles during plant defense; they can strengthen cell walls and be toxic to invading pathogens, and they can act as signaling molecules during defense and plant development (Waszczak et al., 2018; Bleau & Spoel, 2021; Mittler et al., 2022). There are several challenges in understanding how ROS act as signals: (1) ROS are generated at multiple cellular locations, including the apoplast, mitochondria, chloroplast, peroxisome, and cytosol. (2) ROS are produced through diverse mechanisms, such as metabolic processes and catalytic formation by RESPIRATORY BURST OXIDASE HOMOLOGUEs (RBOHs). (3) ROS are perceived through specific molecular mechanisms, for example, via cysteine modifications in proteins (Waszczak et al., 2018). One way to start addressing the role of ROS in signaling is to use treatments in which the ROS are formed at a specific cellular location, rather than a general treatment that raises ROS at several places. Another approach is to use mutants defective in specific subcellular ROS production or scavenging. For example, extensive work with the peroxisomal cat2 (catalase 2) has established how peroxisomal H_2_O_2_ contributes to plant defenses and development (Yang et al., 2019; Baker et al., 2023). In defense signaling, and especially plant–pathogen interactions, ROS produced in the apoplast are considered a key signal, exemplified by extensive regulation of RBOH enzymes that produce superoxide in the apoplast (Castro et al., 2021). Both RBOHD and RBOHF contribute to ROS used for signaling in abiotic and biotic stress (Torres et al., 2002) but RBOHD transcription levels are higher than RBOHF transcription levels (Morales et al., 2016), and RBOHD appears to be more important for producing ROS used in cell‐to‐cell communication (Miller et al., 2009; Castro et al., 2021). A direct approach to study the signaling function of apoplastic ROS is to apply the air pollutant ozone (O_3_). Ozone enters the plant via stomata and, in the cell wall, breaks down into hydrogen peroxide and superoxide; thus, ozone provides a tool to directly activate apoplastic ROS signaling without the need for any other manipulation of the plant (Xu et al., 2015; Waszczak et al., 2018).
As the global threat of phytopathogens and abiotic stress intensifies due to climate change, food security is at risk (Zandalinas & Mittler, 2022). A challenge to understand plant defenses is the redundancy and multiple different defense signaling pathways. For example, using higher‐order Arabidopsis mutants where several hormone and defense signals are removed uncovered that if one hormone signal is removed, compensatory mechanisms are present that still provide defenses (Hillmer et al., 2017). Inspired by animal models, where the entire immune system can be compromised (Walsh et al., 2017), we tested whether the same could be done in Arabidopsis by combining several mutations in key defense and hormone‐related genes. A plant without defenses would then act as a platform to study how plant defenses toward abiotic and biotic stress are organized.
Here, we describe the defenseless mutant (abi1‐1 coi1 eds1 ein2 rbohD sid2; Fig. 1a), lacking signaling or biosynthesis of hormones (ABA, ET, JA, and SA), pathogen signaling (EDS1), and apoplastic ROS production (RBOHD). With this set of mutants, we expect to remove defense signaling, but not necessarily all plant defenses, as defenses also rely on various other mechanisms, including antioxidants, secondary metabolites, and barriers, including the cell wall and cuticle (Göhre & Robatzek, 2008; Waszczak et al., 2018; Zaynab et al., 2018). We use defenseless in multiple assays related to biotic and abiotic stress, intra‐organellar signaling, and transcriptomics to define the contribution of these key regulators toward plant defenses. While defenseless was compromised in plant defenses, it also retained a largely intact transcriptome response, indicating that the plant defense signaling network is redundant, and it is difficult to eliminate plant defense signaling.
Overview of pathway disruptions in defenseless and its growth and stress phenotypes. (a) Schematic overview of the signaling pathways disrupted in the defenseless mutant. (b) Phenotypic comparison between Columbia‐0 (Col‐0) and defenseless: (i, ii) images showing 3 wk old Col‐0 and defenseless under standard growth conditions; (iii, iv) when grown in growth chambers for seed production, the defenseless mutant occasionally shows complete collapse and dying, (v, vi) root growth of Col‐0 and defenseless on ½ MS‐medium. (c) Leaf disk assay of Col‐0 and defenseless under different treatments with methyl viologen (MV), which creates reactive oxygen species (ROS) in chloroplast, and antimycin A (AA) that inhibits mitochondrial complex III. Data from seven biological repeats are presented as box‐and‐whisker plots (n = the number of leaf disks). The boxes represent the 25th to the 75th percentiles, with the horizontal line plotted at the median values. The individual data points are shown as circles, and the whiskers represent the minimum and maximum values. Statistical significance was determined using two‐way ANOVA, followed by Tukey's test (P < 0.05). Samples with different letters are significantly different. (d) Reverse transcription quantitative polymerase chain reaction (RT‐qPCR) was used to assess the relative expression of marker genes under control and ozone (see Supporting Information Fig. S1b for additional genes). Data represent five biological replicates, and statistical significance was determined using two‐way ANOVA, followed by Tukey's test (P < 0.05). Samples with different letters are significantly different.
Materials and Methods
Plant lines and growth conditions
Seeds of Arabidopsis thaliana Columbia‐0 (Col‐0) and mutants were obtained from Nottingham Arabidopsis Stock Centre or were gifts from Prof. Jane Parker (eds1‐2 in Col‐0), Dr Miguel Torres (rbohD), and Prof. Julian Schroeder (abi1‐1 in Col‐0). We initially made the mutants coi1 ein2 sid2 (Xu et al., 2015) and coi1 eds1 ein2 sid2 (Overmyer et al., 2018). As coi1‐16 is partially male sterile, higher‐order mutants containing coi1‐16 were used as pollen acceptors in crosses to abi1‐1 and rbohD to generate the sextuple abi1‐1 coi1‐16 eds1‐2 ein2‐1 rbohD sid2‐1, which we will refer to as the defenseless mutant. All the single mutants have been extensively studied for their function in plant defense and hormone signaling (Supporting Information Table S1). Identification of higher‐order mutants was based on initial screenings for hormone phenotypes: ein2 was identified via the triple response, coi1‐16 was identified via MeJA root growth assay and abi1‐1 was identified via ABA root growth assay. Subsequently, the presence of all mutations were identified with PCR‐based markers (Table S1).
Seeds were sown on 1 : 1 peat/vermiculite, stratified for 3 d, and then grown at 22°C/19°C for a week. Seedlings were then transplanted into a new 1 : 1 peat/vermiculite mixture. All plants were grown in a controlled chamber (Weiss Bio1300; Weiss Gallenkamp) or growth rooms, at 22°C/19°C, in relative humidity of 60/90%, under a 12 h : 12 h, light : dark cycle, at 220–250 μmol m^−2^ s^−1^ light intensity. Experiment details for stress experiments and transcriptome analysis are provided in the Methods S1.
Data analysis
All data analyses were performed using the R software (v.4.4.0; R Core Team, 2024; https://www.R‐project.org) or Graphpad Prism 10.4.2. To account for inter‐replicate variability, linear mixed‐effects models were implemented via the ‘lmer’ function from the ‘lme4’ package (Bates et al., 2015), incorporating biological replicates as random effects. Post hoc pairwise comparisons were conducted using Tukey's honest significant difference (HSD) method as implemented in the ‘emmeans’ package (Lenth & Piaskowski, 2017), to adjust for multiple tests and accurately identify significant group‐wise differences. A P‐value of < 0.05, < 0.01, and < 0.001 was considered to denote significant, moderately significant, and highly significant differences, respectively.
Results
Construction of the defenseless mutant
Plant defenses to abiotic and biotic stresses are dependent on plant hormones ABA, ET, JA, SA, and on small signaling molecules like ROS. In plant–pathogen signaling, after recognition of the pathogen, EDS1 is an essential signaling intermediate required for activation of defenses (Locci et al., 2023). We aimed to generate a plant with removed signaling (or biosynthesis) from hormones, ROS, and EDS1. The selected mutants should have strong impairment, but at the same time, the final plant with all mutations should also be viable and produce seeds. We selected the following mutants (Fig. 1a): abi1‐1 that has strongly impaired ABA signaling (Merilo et al., 2013); coi1‐16, a conditional allele for the COI1 JA receptor (Ellis & Turner, 2002) – as complete knockouts of COI1 are infertile, the use of coi1‐16 allows seeds to be produced; eds1‐2 that lacks EDS1 (Bartsch et al., 2006); ein2‐1 with impaired ET signaling (Alonso et al., 1999); rbohD that lacks the apoplastic ROS producer RBOHD (Torres et al., 2002; Miller et al., 2009); and sid2‐1 impaired in the main Arabidopsis SA biosynthesis pathway (Wildermuth et al., 2001). We used the JA receptor coi1 rather than a JA biosynthesis mutant (e.g. aos, allene oxide synthase), as biosynthesis mutants also lack intermediates that can act as signals (Wasternack & Feussner, 2018). The focus of this work was to characterize defenseless (abi1‐1 coi1 eds1 ein2 rbohD sid2), but where appropriate single mutants (especially abi1‐1), and other mutants defective in production of defense molecules were used for comparison.
When grown in clean growth rooms or chambers, the defenseless mutant has no visible signs of stress (Fig. 1b). The leaves of defenseless are narrower and smaller than Col‐0 and are similar to the abi1‐1 single mutant. We quantified fresh weight, which confirmed the smaller size of defenseless (Fig. S1a). The smaller size of defenseless and abi1‐1 probably reflects the role of hormones as regulators of growth, particularly ABA (Cutler et al., 2010). Although defenseless can complete the entire life cycle to produce seeds, it is a poor seed producer, like the single coi1‐16. While defenseless showed normal growth in clean growth conditions, we observed occasional collapse and dying of the plants (Fig. 1b) and moving of plants from the growth chamber to the glasshouse often led to infection from (yet unidentified) opportunistic pathogens (Fig. S1e). Growing plants in sterile in vitro growth medium, defenseless, showed similar growth as Col‐0 (Fig. 1b).
Organelle ROS metabolism and signaling
One of the most sensitive assays to monitor the health status of plants is to measure the quantum yield of photosystem II, F v/F m (Murchie & Lawson, 2013). In nonstressed, control plants, the defenseless F v/F m values were similar to Col‐0 (Fig. 1c), indicating that as long as plants are maintained in clean growth conditions, they do not need active defenses. Plants use active production of apoplastic ROS for intra‐ and inter‐cellular signaling (Waszczak et al., 2018). We confirmed that defenseless, similarly to rbohd, lack activation of the apoplastic ROS burst, seen after treatment with the MAMP flg22 (Fig. S2).
Plant cells contain three genomes (nuclear, chloroplast, and mitochondrial), where signals need to be exchanged between organelles via anterograde and retrograde signaling for proper transcriptional regulation and plant function. Through applications of chemicals, the operation of these signals can be monitored as changes in F v/F m (Shapiguzov & Kangasjärvi, 2022). Methyl viologen (MV) extracts electrons from photosystem I to create ROS in the chloroplasts, and antimycin A (AA) inhibits mitochondrial complex III activity leading to ROS production in the intermembrane space. With single treatments, this allows monitoring of the capacity of the chloroplast or mitochondria to withstand stress. In addition, the double MV + AA treatment also allows testing signaling and metabolic interactions between organelles, as the AA treatment will activate mitochondrial retrograde signaling to the nucleus, resulting in upregulation of mitochondrial ALTERNATIVE OXIDASE1a (AOX1a), which in turn will help to protect the chloroplasts from the ROS damage caused by the MV treatment (Shapiguzov et al., 2019). We used a previously established leaf disk assay, where the disks are incubated overnight in darkness with the single MV, AA, and double MV + AA treatments, followed by incubation in repeated cycles of blue light to activate ROS production from MV in the chloroplast (Shapiguzov & Kangasjärvi, 2022). Importantly, the overnight AA incubation allows increased AOX1a transcription to take place before the MV‐mediated ROS production is activated.
In Col‐0, MV alone induced strong damage, whereas AA treatment alone had minor effects (Fig. 1c). In line with previous results, the combined MV + AA treatment led to protection from the damage caused by MV (Fig. 1c; Shapiguzov et al., 2019). In defenseless, MV also caused damage, but this was not statistically significantly different from Col‐0, suggesting that the response to chloroplast ROS production is not influenced by the impaired defenses in defenseless. By contrast, the single AA treatment gave minor damage to defenseless (but not Col‐0). The combined MV + AA treatment also led to protection from ROS damage in defenseless, although to a lower level than in Col‐0. Overall, the single and double MV + AA assays suggest that defenseless has a wild‐type (WT) chloroplast ROS response and minor defects to mitochondrial stress responses.
To probe the contribution of other ROS sources, especially the peroxisome, we used the leaf disk assay with the catalase inhibitor 3‐AT (Gechev et al., 2002). Increasing concentrations of 3‐AT led to decreased F v/F m, indicating that improper ROS scavenging in the peroxisome feeds back into chloroplast function (Fig. S3a). In this assay, defenseless performed better than Col‐0. SA is not only activating defense signaling via its primary receptor NONEXPRESSER OF PR GENES 1 (Zavaliev & Dong, 2024), but SA also inhibits catalase to regulate the ROS balance during defense (Yuan et al., 2017). We tested a concentration series of SA in the leaf disk assay (Fig. S3b). At low concentration (0.1 mM), no difference between genotypes was observed, and at high concentration (0.5 mM), F v/F m was severely decreased in both genotypes. The intermediate concentration (0.25 mM) was informative, where defenseless was more tolerant than Col‐0 (Fig. S3b).
Application of the air pollutant ozone that enters the plant through stomata allows the monitoring of ROS signaling via apoplast to nucleus (Xu et al., 2015). In our previous work, we used real‐time quantitative PCR (qPCR), microarrays, and RNA‐seq to investigate how apoplastic ROS signaling regulates transcriptional responses (Brosché et al., 2014). With qPCR, we tested seven different marker genes after 2‐h 350 nl l^−1^ ozone treatment (Figs 1d, S1b). RAP2.6 (RELATED to AP2 6) expression was highly induced by ozone in Col‐0. By contrast, the defenseless mutant exhibited a significant reduction in RAP2.6 expression compared with Col‐0 (Fig. 1d). However, within the defenseless background itself, ozone treatment still led to a significant upregulation of RAP2.6 relative to its control, suggesting a partial response retained despite the impaired defenses. For PHOSPHOLIPASE A 2A and CALMODULIN‐LIKE 37 (CML37), transcript levels were significantly elevated in response to ozone treatment in both genotypes. However, CML37 was markedly upregulated in the defenseless mutant relative to Col‐0 under ozone treatment (Fig. 1d). Thus, we observed three different transcriptional ozone responses: lower, the same, and higher in defenseless. This suggests that the impaired signaling in defenseless can function as both positive and negative regulators of ROS transcriptional responses.
The defenseless transcriptome
To gain full insights into which signaling is impaired in defenseless we performed RNA‐seq analysis after 2‐h ozone treatment. Multidimensional scaling (MDS) plots revealed distinct clustering of the samples (Fig. 2a). The ozone treatment was the main separator, followed by genotype. The separation by genotype was more pronounced during the ozone treatment than in control samples. We selected genes with significantly altered expression based on an adjusted P‐value < 0.05 and log_2_ fold change (< −1 for downregulated and > 1 for upregulated; Table S2). This filtering provided us with 3648 genes upregulated in Col‐0, 4884 genes upregulated in defenseless, 2270 genes downregulated in Col‐0, and 2646 genes downregulated in defenseless, which we displayed as volcano plots (Fig. 2b) and to get the differences between the genotypes they were compared in Venn diagrams (Fig. 2c). More than half of the differentially expressed genes were shared between the genotypes, suggesting that apoplastic ROS signaling is largely independent from hormone, EDS1, and RBOHD pathways.
Transcriptional responses induced by ozone in Columbia‐0 (Col‐0) and defenseless mutant. Twenty‐two‐day‐old plants were exposed to 350 nl l−1 of O3 for 2 h, and transcriptome changes were accessed with RNA‐seq (n = 3). (a) Multidimensional scaling plot illustrating sample relationships based on transcriptome profiles. (b) Volcano plot illustrating the distribution of ozone‐regulated genes in Col‐0 and defenseless. Dots representing genes are color‐coded based on statistical significance and fold‐change thresholds: red dots indicate genes with P‐values < 0.05, green dots represent genes with log2FC ≥ 1, and purple dots highlight differentially expressed genes meeting both criteria. Gray dots correspond to nonsignificant genes. Upregulated genes are positioned on the right, while downregulated genes are on the left. (c) Venn diagrams of genes with significantly increased and decreased expression by ozone in Col‐0 and defenseless. (d) Dot plot of enriched Gene Ontology (GO) biological processes among ozone‐responsive genes. Bar diagram represents the gene list sizes used for the enrichment analysis depicted in the dot plot.
To get the biological context of the differentially expressed genes, we performed Gene Ontology (GO) analysis on the GO category biological process. Again, defenseless displayed a remarkably similar profile as Col‐0, with only subtle differences; for example, ‘response to ethylene’ was upregulated by ozone in Col‐0 but not in defenseless.
The analysis above was based on the identification of genes significantly differentially expressed in Col‐0 and defenseless through analysis of control vs ozone samples. The complementary approach is to instead analyze Col‐0 vs defenseless control samples, and Col‐0 vs defenseless ozone samples (Fig. 3; Table S3). We used the same selection criteria as above (adjusted P‐value < 0.05 and log_2_ fold change < −1 for downregulated and > 1 for upregulated) and plotted the genes as a volcano plot (Fig. 3a) and in Venn diagrams (Fig. 3b). We identified 103 genes with higher expression in defenseless in control, 629 genes with lower expression in defenseless in control, 1442 genes with higher expression in defenseless in ozone and 755 genes with lower expression in defenseless in ozone. As already suggested by the MDS plot (Fig. 2a), the defenseless mutant had more differentially expressed genes in the ozone treatment than in control, and with the most genes belonging to the category upregulated in defenseless in the ozone treatment (Fig. 3b). This matches the profile of the CML37 marker gene used in qPCR (Fig. 1d). To put the transcriptional differences between Col‐0 and defenseless into a biological context we performed GO analysis for biological process (Fig. 3c). As anticipated based on the impaired signaling pathways in defenseless, many GO categories related to hormones and defenses showed enrichment among the genes differentially expressed in defenseless.
Difference between the Columbia‐0 (Col‐0) and defenseless transcriptome. (a) Volcano plots illustrating differentially expressed genes between Col‐0 and defenseless under control and ozone conditions. Upregulated genes appear on the right, downregulated on the left. Dots are color‐coded by statistical and fold‐change thresholds: red (P < 0.05), green (log2FC ≥ 1), purple (differentially expressed genes meeting both criteria with gene labels), and gray (nonsignificant). (b) Venn diagrams displaying the genotype difference between the Col‐0 and defenseless differentially expressed genes under control and ozone conditions. These gene sets were used for subsequent enrichment analyses. (c) Dot plot of enriched Gene Ontology (GO) biological processes among differentially expressed genes in defenseless under control and ozone treatments. The accompanying bar chart indicates gene set sizes used for enrichment analysis.
In control conditions, the biological process lipid transport was upregulated in defenseless, whereas genes associated with SA signaling, systemic acquired resistance, and bacterial molecular recognition were downregulated in defenseless. Under ozone treatment, there was a significant upregulation of genes involved in cellular response to hypoxia, oxygen homeostasis, chitin response, and SA‐mediated signaling pathways, indicating a broad activation of stress‐responsive pathways. Conversely, genes implicated in S‐glycoside and glucosinolate biosynthesis, ET signaling, signal transduction regulation, and cell communication were downregulated in defenseless. Plants manage ROS levels via antioxidants and enzymes that scavenge ROS, for example catalase that removes H_2_O_2_. In Arabidopsis, at least 150 genes are involved in determining the ROS levels (Mittler et al., 2004) and we checked their expression levels (Table S4). In control treatment, few genes differed between Col‐0 and defenseless. In ozone, more genes had differential expression between defenseless and Col‐0, but for many of these, the expression was higher in defenseless, suggesting that the mutant would have increased capacity to handle ROS. We continued to measure reduced and oxidized ascorbic acid, where defenseless had increased amount of reduced ascorbic acid (Fig. S4).
To identify a core set of genes with decreased expression in defenseless and their biological context, we selected 162 genes that were significantly downregulated in defenseless in both control and ozone treatment (Fig. 3b), and performed hierarchical clustering using Euclidean distance and complete linkage (Fig. 4). Cluster I contained genes strongly upregulated by ozone in Col‐0 but not in defenseless, Cluster II had genes that have high expression in Col‐0 (both control and ozone) and low expression in defenseless, Cluster III had genes with high expression in Col‐0 control and low expression in defenseless. In this core set of genes, there are numerous regulators of hormone and pathogen signaling that have been extensively studied for their function in plant–pathogen interaction, for example the FLAGELLIN‐SENSITIVE 2 (FLS2) receptor that detects flagellin (Chinchilla et al., 2006), FLG22‐INDUCED RECEPTOR‐LIKE KINASE 1, which has been used as a marker gene for pathogen signaling (Asai et al., 2002), and AGD2‐LIKE DEFENSE RESPONSE PROTEIN 1 involved in the pipecolic acid biosynthetic pathway that is a key signal for systemic acquired resistance (Bernsdorff et al., 2016). Strikingly, there are also multiple genes from JA biosynthesis or catabolism (ALLENE OXIDE CYCLASE 1 (AOC1), OXOPHYTODIENOATE‐REDUCTASE 3 (OPR3), and JASMONIC ACID OXIDASE 2 (JAO2)) and the ET signaling pathway (ETHYLENE RESPONSE 2 (ETR2), ETHYLENE RESPONSE SENSOR 2 (ERS2), and EIN3‐BINDING F BOX PROTEIN 2 (EBF2)). Overall, this shows that defenseless is impaired in the signaling pathways that would be expected from the mutants used to create defenseless.
Heatmap of the 162 genes downregulated in the defenseless mutant under both control and ozone conditions. The first column lists gene accession numbers, the second lists gene symbols (retrieved from the TAIR database), and the third indicates associated functional pathways (see also Supporting Information Table S5). ABA, abscisic acid; Eth, ethylene; JA, jasmonic acid; P, pathogens; ROS, reactive oxygen species; SA, salicylic acid.
The response to pathogens in defenseless
To test defenseless in plant–pathogen interaction, we performed assays with both fungal and bacterial pathogens (Fig. 5). After 1 wk, fungal lesions were quantified in Col‐0 and defenseless, and in cyp79b2b3 (defective in indolic glucosinolates biosynthesis) used as positive control (Fig. 5a). Fungal lesion areas were significantly larger in defenseless mutants compared with Col‐0 WT (P < 0.01; Fig. 5b), but the lesion area of infection of the defenseless mutant was less than that of the cyp79b2b3 mutant. Here, it should be noted that the abi1‐1 mutation provides a permeable cuticle (Fig. 6), which was previously shown to give strong resistance toward fungal pathogens (Bessire et al., 2007). As defenseless was susceptible to Alternaria infection, this suggests that potential resistance from a permeable cuticle is not enough to overcome the other impaired defenses in defenseless.
*Pathogen infection assays of Columbia‐0 (Col‐0) and different mutants (defenseless, cyp79b2b3, and eds1 sid2) by fungal and bacterial pathogens. (a) Alternaria alternata infection in Col‐0, defenseless, and cyp79b2b3 (positive control). Representative images show necrotic lesions indicative of fungal colonization. (b) The lesion area was quantified as a percentage of the total leaf surface. Each genotype included 40 biological leaves (n = 40). (c) Pseudomonas syringae pv tomato DC3000 (Pst) infection in Col‐0, defenseless, and eds1sid2. (d) A time‐course analysis measured bacterial titers at 24‐, 48‐, 72‐, and 96‐h postinoculation (hpi). Individual measurements from 40 biological replicates (b) and 24 measurements across three repeat groups (d) are displayed as box‐and‐whisker plots. Boxes denote the interquartile range (25th–75th percentiles) with the median indicated by a horizontal line. Circles (and distinct symbols for repeat groups) represent individual data points, and whiskers indicate the minimum and maximum values. Statistical significance was assessed using one‐way ANOVA followed by Tukey's honest significant difference (HSD) test. Significance levels: ***, P < 0.0001; *, P < 0.05; ns, not significant (P > 0.05). Bacterial assays were conducted with 24 biological replicates across three independent experiments (n = 8 per replicate).
Developmental and abiotic stress assays in Columbia‐0 (Col‐0) and mutants (abi1‐1 and defenseless). (a) Toluidine blue (TB) staining of leaves following 2‐h incubation. Col‐0 showed limited staining, indicative of an intact cuticle, while abi1‐1 and defenseless displayed strong blue staining, consistent with cuticular defects. (b) Quantification of TB permeability revealed significant differences between abi1‐1 and defenseless (P < 0.05), and a highly significant difference between Col‐0 and both mutants (P < 0.01). Data were analyzed using one‐way ANOVA. (c) Water loss (%) across genotypes (n = 65, 5 biological repeat groups). One‐way ANOVA with Tukey's honest significant difference (HSD) test indicated highly significant water loss in abi1‐1 and defenseless compared with Col‐0 (P < 0.01), and a significant difference between the two mutants (P < 0.05). (d) Ion leakage as a measure of ozone‐induced cell death following 6‐h O3 exposure. Box plots represent data from seven biological repeats (n = 33–35 plants). Two‐way ANOVA with Tukey's HSD test showed highly significant differences (P < 0.01) between Col‐0 and both mutants under ozone treatment, while no significant difference (ns, P > 0.05) was observed between abi1‐1 and defenseless. (e) Reverse transcription quantitative polymerase chain reaction (RT‐qPCR) was used to assess the relative expression of marker genes (RAP2.6, HAI1, NCED3, and GolS) under control and NaCl (see Supporting Information Fig. S5d for additional genes). Data represent seven biological replicates, and statistical significance was determined using two‐way ANOVA, followed by Tukey's test (P < 0.05). Samples with different letters are significantly different. (f) Representative plant images following 2‐h ozone exposure. Bars, 1 cm.
For bacterial pathogen assays, we used spray inoculation with Pseudomonas syringae pv tomato DC3000. In addition to defenseless, we also used the eds1 sid2 double mutant, as SA is considered the main defense against bacterial pathogens. Both eds1 sid2 and defenseless exhibited significantly higher susceptibility to bacterial infection relative to Col‐0. Visible disease symptoms (chlorosis, necrosis, and tissue collapse) became apparent after 48 h postinoculation (hpi; Fig. 5c). Quantitative bacterial growth assays revealed markedly elevated bacterial titers as early as 24 hpi, with statistically significant differences persisting through 96 hpi (Fig. 5d). While both eds1 sid2 and defenseless were highly susceptible, the eds1 sid2 double mutant was the most susceptible mutant. This suggests that other mutations in defenseless can provide some level of resistance, for example, the abi1‐1 mutation that not only provides a permeable cuticle but also leads to more open stomata (Merilo et al., 2013), which alters the aquatic environment inside of the leaf and hence bacterial proliferation (Lajeunesse et al., 2023).
The response to abiotic stress in defenseless
ABA is considered the main hormone in various signaling pathways related to abiotic stress. In addition, ABA is also a regulator of developmental responses, for example, seed germination and cuticle formation (Cui et al., 2016). To check for cuticle permeability, we used toluidine blue stain in abi1‐1 and defenseless (Fig. 6a). Quantitative analysis of the toluidine‐stained surface area revealed increased cuticle permeability in both mutants, with a subtle increase in defenseless vs abi1‐1. This suggests that ABA signaling is the main hormone in regulating cuticle formation, with a limited contribution from other hormone signaling pathways impaired in defenseless.
ABA is also the main regulator of guard cell function in the opening and closing of stomata. This impacts many stresses, including the response to bacterial pathogens, entry of ozone into the plant, and water balance. A classical assay for guard cell function is to measure water loss from detached leaves (Klein et al., 1996; Outlaw Jr, 2003), but here it should be considered that this assay will not distinguish between water that is lost through permeable cuticle vs open stomata, as both will contribute to the water that evaporates from leaves. Both abi1‐1 and defenseless mutants demonstrated significantly higher water loss compared with Col‐0. While there was a statistically significant change between abi1‐1 and defenseless, the difference in water loss was so minor that it is probably not biologically relevant. Thus, for both cuticle permeability and water loss, it is the abi1‐1 mutation in defenseless that acts as the main contributor to the defenseless phenotype.
We use treatments with the air pollutant ozone not only to study the role of apoplastic ROS in activation of defense signaling but also to study how ROS regulates cell death (Brosché et al., 2014). We quantified cell death after a 6‐h exposure to ozone, in abi1‐1 and defenseless (Fig. 6d). As mutants with open stomata and/or permeable cuticle are highly ozone sensitive due to increased amount of ozone entering the plant, we predicted that the abi1‐1 mutant would be the main source of ozone sensitivity in the defenseless mutant. Both mutants were highly ozone sensitive (Fig. 6d,f), but with no significant difference between the mutants, suggesting that abi1‐1 provides the high ozone sensitivity to defenseless.
Another abiotic stress that uses ABA as a signaling intermediate is NaCl stress. We first tested defenseless in a NaCl root growth assay, where root elongation was observed at increasing concentrations of NaCl. No difference in root growth was observed between Col‐0 and defenseless (Fig. S5d). To study transcriptional responses to NaCl in Arabidopsis research, it is common to use relatively young seedlings. As we wanted to stay consistent in our use of adult plants for all transcript profiling assays, we performed the assay by putting a 10 μl drop of 150 mM NaCl on the surface of fully developed leaves. Samples were harvested after 6 h and from corresponding control samples treated with water. We selected seven marker genes for qPCR (Figs 6e, S5d), based on a recent NaCl transcriptome study (Lamers et al., 2025), as well as marker gene and transcriptome data from the ozone experiments (Figs 1, 2, 3, 4). As in the ozone treatment (Fig. 1d), increased transcript levels were seen after NaCl treatment for RAP2.6 (Fig. 6e). Notably, RAP2.6 expression was decreased in defenseless (but not in abi1‐1), again consistent with the data from the ozone treatment (Fig. 1e). Also, ERF109 showed the same profile (Fig. S5d). HIGHLY ABA‐INDUCED PP2C GENE 1 (HAI1) and RESPONSIVE TO DESICCATION 20 were induced in response to NaCl stress in all three genotypes (Figs 6e, S5d). Similarly, NCED3 (9‐cis‐Epoxycarotenoid Dioxygenase 3), a key rate‐limiting enzyme in the ABA biosynthesis pathway, was also significantly upregulated under salt stress in all genotypes. One of the few genes associated with ABA signaling in the core 162 genes downregulated in defenseless was Gol‐S (Galactinol synthase; Fig. 4). GolS is involved in the biosynthesis of galactinol, a precursor of raffinose family oligosaccharides important for osmotic and oxidative stress tolerance. Consistent with our transcriptome data, transcript levels for Gol‐S were low in defenseless. However, it was significantly upregulated by NaCl in the abi1‐1 mutant. Overall, the defenseless mutant was more impaired in NaCl transcriptional responses than the abi1‐1 mutant, suggesting contributions from other hormones than ABA to signaling in the response to NaCl.
Discussion
In this work, we investigated whether it is possible to generate a plant that lacks all the major plant defense signaling pathways, and how this plant would respond to perturbations of organellar function, abiotic and biotic stress. We selected mutants impaired in hormone signaling or biosynthesis (ABA – abi1‐1, JA – coi1‐16, ET – ein2, and SA – sid2). In pathogen signaling, EDS1 is a key node in basal and ETI (Cui et al., 2017), and ROS generated from RBOHD act as signaling molecules in both abiotic and biotic stress (Miller et al., 2009). In defenseless (abi1‐1 coi1‐16 eds1 ein2 rbohd sid2), the majority of the well‐characterized plant defense signals should be removed. The rationale for choosing these mutants was also based on previous work with double, triple, and quadruple mutants used to study defense signaling. Characterization of eds1 sid2 showed that EDS1 and SA act redundantly in pathogen defenses (Venugopal et al., 2009; Cui et al., 2017). Extensive work with dde2 ein2 pad4 sid2, deficient in JA biosynthesis (dde2, a mutation in aos), ET signaling, SA biosynthesis, and for PAD4 (an interacting partner protein to EDS1) showed that using single mutants is not enough to understand defense signaling: when one signal pathway is missing other signals can compensate to still provide defense (Hillmer et al., 2017). We have previously also analyzed the transcriptional responses of many of the single (abi1‐1, coi1‐16, ein2, and sid2), double (coi1 sid2, coi1 ein2, and ein2 sid2), and triple mutants (coi1 ein2 sid2) that are part of defenseless (Xu et al., 2015; Vuorinen et al., 2021), and these mutants have a mostly intact transcriptional response. Hence, defenseless should be a valuable complement to single mutant studies to get a broader view of the redundancy in signaling. With the defenseless mutant, we were able to evaluate the contribution of hormone and pathogen signaling pathways to defense and defense signaling. As a large part of transcriptional regulation appears to be intact in defenseless, this suggests that additional signaling pathways contribute to plant defense and defense signaling, via for example Ca^2+^ or kinases.
Transcriptional regulation in defenseless
We used treatment with the air pollutant ozone to initiate ROS signaling from the apoplast (Xu et al., 2015; Waszczak et al., 2018). In line with our previous results (Xu et al., 2015), several thousand genes were up‐ and downregulated by the ozone treatment in both Col‐0 and in defenseless (Fig. 2). The large number of ozone‐regulated genes in defenseless suggests that there are alternative signaling pathways that operate in parallel with hormone signaling. These independent signaling pathways could, for example, be connected to Ca^2+^, an important signaling molecule in many stress signaling pathways (Luan & Wang, 2021), or to kinase‐mediated signaling, for example, via mitogen‐activated protein (MAP) kinases that are activated in response to many stresses, including ozone (Ahlfors et al., 2004; Sun & Zhang, 2022). In this scenario, ozone initiates ROS signaling in the apoplast, leading to activation of Ca^2+^ channels and/or activation of kinase signaling that relays the signal to activation of transcription factors in the nucleus and transcriptional reprogramming.
However, even if there is a large overlap in ozone‐regulated genes between WT and defenseless, there are numerous genes that show altered expression in the mutant. The largest category of defenseless differentially expressed genes included genes with increased expression in the ozone‐treated mutant. This can be seen for the marker gene CML37 in qPCR (Fig. 1d), and in transcriptome data with 1442 genes (Fig. 3). GO analysis of these genes suggests functions related to oxygen sensing, SA, and immunity signaling. As these genes have higher expression in defenseless, it suggests that hormone signaling acts as a negative regulator to keep their expression levels low. This is in line with studies of autoimmune mutants, where too strong activation of plant defenses leads to impaired growth (Chakraborty et al., 2018). Thus, defense signaling in plants should balance appropriate activation of defense with overactivation that could lead to detrimental effects. The other category of genes with differential expression are genes downregulated in defenseless, in control, and/or ozone treatment. GO enrichment of these genes indicated their role in SA, JA, ET, and pathogen/defense signaling, and that these genes are downregulated in defenseless is consistent with this mutant being impaired in hormone and defense signaling (Figs 3, 4).
ROS functions as a crucial secondary messenger in diverse plant stress responses, including plant–pathogen interactions. Previous work by Vuorinen et al. (2021) demonstrated a substantial overlap between genes transcriptionally regulated by ozone exposure and those responsive to Botrytis cinerea infection, suggesting a shared transcriptional signature indicative of ROS‐mediated signaling. To further investigate the relevance of ozone‐induced transcriptional changes as a readout for ROS‐associated plant responses (Vaahtera et al., 2014), we used data from the Genevestigator database to evaluate transcriptional regulation of four marker genes we used for qPCR. This revealed that strongly ozone‐responsive genes (CML37, CRK37, MDHAR, and RAP2.6) were consistently and similarly regulated by multiple biotic stress conditions, including infections by bacterial and fungal pathogens (Fig. S6). This supports the notion that ozone initiates apoplastic ROS signaling (Waszczak et al., 2018), similarly to the ROS signaling activated during pathogen attack. Consequently, our ozone RNA‐seq dataset provides a useful resource for dissecting ROS‐dependent regulatory networks involved in plant immune responses.
To get a further biological context of the genes downregulated in defenseless, we focused on the core 162 genes downregulated in defenseless in both control and ozone treatment (Fig. 4). Among these core genes are many genes that belong to JA biosynthesis/signaling (AOC1, OPR3, JAO2, and JAZ9), ET signaling (ETR2, ERS2, and EBF2), and transcription factors characterized as executors of ET/JA signaling (ERF1; Lorenzo et al., 2003) and SA signaling (WRKY38; Kim et al., 2008). Overall, the defenseless transcriptome analysis indicates that the mutant is severely impaired in specific aspects of JA/SA/ET/defense signaling, but at the same time the mutant also has other signaling pathways that are functional to mediate the ROS signal into transcriptional changes.
The transcription factor RAP2.6 used as a marker gene in qPCR illustrates another aspect of signaling to take into consideration (Figs 1d, 6e). Expression of RAP2.6 is regulated by several biotic and abiotic stresses, and by JA and ABA signaling (Zhu et al., 2010; Vuorinen et al., 2021). In defenseless, its expression levels were very low, but both ozone and salt stress led to increased transcript levels in defenseless. This indicates that many different signaling pathways can target this gene, both the hormone‐dependent and independent signaling pathways.
Stress responses in defenseless
We established the defenseless mutant as a platform to study defense and hormone signaling. However, the transcriptional response is only one aspect of defense, and to get a broader view of the defenseless phenotype, we evaluated its stress and developmental phenotypes. When grown in vitro or in a clean growth room or growth chambers, the defenseless mutant does not appear to be visibly stressed, and its F v/F m value is the same as WT, suggesting that as long as it is maintained in nonstressful growth conditions, it performs well. However, defenseless is smaller than Col‐0 (Fig. S1a). This is likely a reflection of hormone regulation of growth. For example, it has long been observed that ABA biosynthesis and signaling mutants are smaller than WT (Cutler et al., 2010).
ROS are used as signal molecules in many stress signaling pathways, but their location differs according to stress; for example, ozone and pathogens initiate ROS signaling from the apoplast (Waszczak et al., 2018). To investigate organellar (chloroplast, mitochondria, and peroxisome) ROS production, we used chemicals to perturb the function in chloroplast (MV), mitochondria (AA), and peroxisome (3‐AT). In the single MV treatment, this showed similar damage in WT and defenseless, indicating that impaired signaling in defenseless is not contributing to chloroplast function (Fig. 1c). By contrast, the defenseless mutant was sensitive to the AA treatment, which initiates ROS production in mitochondria through inhibition of the electron transport chain. Proper exchange of signals between chloroplast, mitochondria, and nucleus is needed to protect against stress that affects the organelles. This can be evaluated in a combined MV + AA assay, where signaling initiated from AA stressed mitochondria goes to the nucleus, resulting in transcriptional reprogramming that ultimately helps the chloroplast to better tolerate MV (Shapiguzov et al., 2019). The combined MV + AA treatment gave protection toward MV damage both in WT and defenseless, but to a slightly lower level in defenseless. Application of the catalase inhibitor 3‐AT showed increased tolerance in defenseless (Fig. S3a). Overall, the 3‐AT, MV, and AA assays suggest a slightly altered response from the chemical that impairs mitochondria function, but, as defenseless can still execute protection for the chloroplast, this suggests a mostly intact organellar signaling network in defenseless.
Most of the previous work to impair multiple signaling pathways has focused on ET, JA, SA, and pathogen signaling (Xu et al., 2015; Hillmer et al., 2017). In defenseless, we also impaired ABA signaling with the abi1‐1 mutant. To further evaluate defenseless, we performed a series of assays where ABA is considered the main regulatory hormone, notably cuticle development and regulation of stomatal aperture. The plant cuticle constitutes a critical barrier limiting pathogen entry. Both abi1‐1 and defenseless mutants exhibited significantly increased toluidine blue uptake relative to Col‐0, indicating compromised cuticular integrity (Fig. 6). Water loss from detached leaves acts as an indicator for both permeable cuticle and open stomata, as these are the main places where water can evaporate from leaves. Again, both abi1‐1 and defenseless showed high water loss compared with Col‐0 (Fig. 6).
To probe defenseless responses in defense against pathogens, we used both fungal and bacterial pathogens. The lesions observed following Alternaria (a necrotrophic pathogen) infection directly implicate compromised antifungal defenses, likely due to attenuated JA‐ and ET‐mediated signaling pathways (Qi et al., 2012). As a positive control, we employed the Arabidopsis glucosinolate biosynthesis mutant cyp79b2b3, which had larger lesions than the defenseless mutants (Fig. 5a,b). Interpretation of defenseless fungal defenses is complicated by the permeable cuticle observed in defenseless (Fig. 6). A permeable cuticle, including that seen in strongly impaired ABA mutants, provides strong resistance to fungal pathogens (Serrano et al., 2014). However, as defenseless was susceptible to Alternaria infection, this suggests that the resistance provided by a permeable cuticle was insufficient to overcome the other impaired defenses in defenseless.
Both defenseless and the eds1 sid2 double mutant showed high susceptibility to the bacterial pathogen Pst. However, defenseless was not more susceptible than eds1 sid2. As SA is considered the main defense hormone, this implicates SA‐mediated transcriptional activation as the main reason for susceptibility in defenseless. Also, the interpretation of bacterial susceptibility is challenging. As defenseless has a permeable cuticle and more open stomata from the abi1‐1 mutation, this allows additional entry points for the bacteria. In addition, mutants with more open stomata are more resistant to Pst infection due to loss of water, preventing the formation of an aqueous phase required for Pst growth (Kemppinen et al., 2025). As defenseless was susceptible to Pst to almost the same level as eds1 sid2, this suggests that even if these other mechanisms may have an impact, they are not enough to override the loss of SA and EDS1 in defense against Pst.
Defense against damage from ozone relies on several components, where the first barrier consists of preventing ozone entry into the plants. Mutants with more open stomata, permeable cuticle, or other epidermal defects are ozone sensitive due to higher levels of ozone entering the plant (Waszczak et al., 2024). Once inside of the plant, the balance of hormone signaling influences to which extent ozone will give tissue damage through activation of programmed cell death (Xu et al., 2015). In both abi1‐1 and defenseless, there was extensive tissue damage in leaves, which was further supported by quantification of the damage with ion leakage (Fig. 6). As the damage was at similar levels in both abi1‐1 and defenseless, this suggests that very high entry of ozone into both mutants via the permeable cuticle and open stomata caused the damage and other signaling pathways did not have a major contribution.
NaCl acts as a stress in plants via ABA‐dependent and ABA‐independent signaling pathways (Yoshida et al., 2014). We employed two different assays to evaluate defenseless responses to NaCl, root growth inhibition, and transcriptional responses. Root growth was similar in Col‐0 and defenseless in increasing concentrations of NaCl (Fig. S5b,c). In qPCR, we used several marker genes, including ABA biosynthesis (NCED3), ABA signaling (HAI1), transcription factors (RAP2.6, ERF109), and osmoprotectant biosynthesis (GolS). Both HAI1 and NCED3, considered as markers for ABA signaling (Huang et al., 2018), were similarly induced in all three genotypes, suggesting that their regulation by NaCl is through an ABA‐independent signaling pathway. By contrast, increased expression by NaCl for RAP2.6 and ERF109 was impaired only in defenseless (and not in abi1‐1). This suggests that it is not impaired ABA signaling in defenseless that leads to their lower expression, and instead it's another signaling pathway. This is likely to be JA, as both genes are also suggested to be JA‐regulated (Cai et al., 2014; Vuorinen et al., 2021). We selected Gol‐S as a marker gene as it belonged to the core 162 genes downregulated in defenseless (Fig. 4). Consistent with the transcriptome data, Gol‐S had decreased transcript levels in defenseless. However, it was not downregulated in abi1‐1, suggesting that similarly to RAP2.6 and ERF109, an ABA‐independent signaling pathway is activated by the NaCl treatment.
Which defense signals and plant defenses remain in defenseless?
One intriguing observation was the occasional collapse of defenseless in clean good growth conditions (Fig. 1b). This suggests that even if most of the plants show no signs of stress as indicated by F v/F m (Fig. 1c), the lack of defenses makes defenseless very susceptible to stress once it is initiated. However, there are many nuances to consider; defenseless retained some level of resistance to both A. alternata and Pst DC3000. While susceptibility was elevated relative to Col‐0, it remained lower than that observed in positive control mutants deficient in specific, individual defense components. Transcriptomic profiling via RNA‐seq further revealed that relatively few genes were differentially expressed in both Col‐0 and defenseless mutants, indicating that the overall transcriptional response to ozone remains largely intact. These findings suggest that complete removal of defense signaling requires simultaneous inactivation of multiple, potentially redundant defense mechanisms. The challenge of testing other additional signaling components, for example, Ca^2+^ signaling or MAP kinase signaling, comes from large genetic redundancy in both Ca^2+^ channels and the MAP kinase signaling cascade (Luan & Wang, 2021; Sun & Zhang, 2022). Applications of specific chemical inhibitors, for example Ca^2+^ channel inhibitors or chelators, in the defenseless mutant could reveal which other signaling pathways are still active.
Plant defenses rely not only on signaling once the stress is detected but also on many different types of defenses via antioxidants and scavengers to manage ROS levels, secondary metabolites, and barriers (cell wall, cuticle, callose, suberin deposition, trichomes, etc.). Here, defenseless showed both impairments, for example a permeable cuticle (Fig. 6a), but at the same time an intact or possibly enhanced defense, for example an increase in reduced ascorbate and better tolerance to catalase inhibitors. To make a true defenseless mutant, we will likely need to combine mutations that not only impair signaling but also those that disrupt various preformed or metabolite‐based defenses, such as pad3 impaired in CYP71B15 (Cytochrome P450 71B15) that catalyze the last step in camalexin biosynthesis (Schuhegger et al., 2006) or pad2 impaired in glutamate‐cysteine ligase that catalyze the rate‐limiting step of glutathione biosynthesis (Parisy et al., 2007).
Collectively, our data underscore the complexity and robustness of the plant defense signaling network and provide a foundation for future studies aimed at disentangling the interplay and relative contributions of discrete signaling pathways within a combinatorial framework.
Competing interests
None declared.
Author contributions
BB and MB performed the experiments. BB analyzed the RNA‐seq data. MB generated the defenseless mutant and designed the project. BB and MB wrote the manuscript.
Disclaimer
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
Supporting information
Fig. S1 Growth and stress responses in defenseless. Fig. S2 Flg22 induced ROS burst in Col‐0, defenseless and rbohD from three biological repeats, each consisting of 24 leaf disks. Fig. S3 Leaf disk assay of Col‐0 and defenseless under treatments with 3‐AT (a) or SA (b). Fig. S4 Ascorbate redox status in Col‐0 and defenseless under control conditions. Fig. S5 Cuticle and NaCl responses in defenseless. Fig. S6 Transcriptomic profiles of selected genes induced under various treatment conditions, extracted from the Genevestigator Plants database. Methods S1 Experimental details and materials and methods for transcriptome analysis, abiotic and biotic stress assays.
Table S1 Primer sequences for genotyping and RT‐qPCR, along with references.
Table S2 Differentially expressed genes after ozone treatment in Col‐0 and defenseless.
Table S3 Differentially expressed genes between Col‐0 and defenseless.
Table S4 Genes involved in ROS metabolism or scavenging.
Table S5 List of 162 consistently downregulated genes in defenseless under control and ozone conditions; includes expression counts (used for Fig. 4 heatmap), gene descriptions, and GO terms.Please note: Wiley is not responsible for the content or functionality of any Supporting Information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahlfors R , Macioszek V , Rudd J , Brosché M , Schlichting R , Scheel D , Kangasjärvi J . 2004. Stress hormone‐independent activation and nuclear translocation of mitogen‐activated protein kinases in Arabidopsis thaliana during ozone exposure. The Plant Journal 40: 512–522.15500467 10.1111/j.1365-313X.2004.02229.x · doi ↗ · pubmed ↗
- 2Alonso JM , Hirayama T , Roman G , Nourizadeh S , Ecker JR . 1999. EIN 2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 284: 2148–2152.10381874 10.1126/science.284.5423.2148 · doi ↗ · pubmed ↗
- 3Asai T , Tena G , Plotnikova J , Willmann MR , Chiu W‐L , Gomez‐Gomez L , Boller T , Ausubel FM , Sheen J . 2002. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 415: 977–983.11875555 10.1038/415977 a · doi ↗ · pubmed ↗
- 4Baker A , Lin C‐C , Lett C , Karpinska B , Wright MH , Foyer CH . 2023. Catalase: a critical node in the regulation of cell fate. Free Radical Biology & Medicine 199: 56–66.36775107 10.1016/j.freeradbiomed.2023.02.009 · doi ↗ · pubmed ↗
- 5Bartsch M , Gobbato E , Bednarek P , Debey S , Schultze JL , Bautor J , Parker JE . 2006. Salicylic acid–independent ENHANCED DISEASE SUSCEPTIBILITY 1 signaling in Arabidopsis immunity and cell death is regulated by the monooxygenase FMO 1 and the nudix hydrolase NUDT 7 . Plant Cell 18: 1038–1051.16531493 10.1105/tpc.105.039982 PMC 1425861 · doi ↗ · pubmed ↗
- 6Bates D , Mächler M , Bolker B , Walker S . 2015. Fitting linear mixed‐effects models using lme 4. Journal of Statistical Software 67: 1–48.
- 7Bernsdorff F , Döring A‐C , Gruner K , Schuck S , Bräutigam A , Zeier J . 2016. Pipecolic acid orchestrates plant systemic acquired resistance and defense priming via salicylic acid‐dependent and ‐independent pathways. Plant Cell 28: 102–129.26672068 10.1105/tpc.15.00496 PMC 4746677 · doi ↗ · pubmed ↗
- 8Bessire M , Chassot C , Jacquat AC , Humphry M , Borel S , Petétot JMDC , Métraux JP , Nawrath C . 2007. A permeable cuticle in Arabidopsis leads to a strong resistance to Botrytis cinerea . EMBO Journal 26: 2158–2168.17396154 10.1038/sj.emboj.7601658 PMC 1852784 · doi ↗ · pubmed ↗