From recognition to response: integrated signaling pathways determining pollen acceptance and rejection in Brassicaceae
Tong Zhang, Shuyan Li, Shengwei Dou, Qiaohong Duan

TL;DR
This review explains how plants in the Brassicaceae family control pollen acceptance and rejection through complex signaling pathways in their stigmas.
Contribution
The paper provides a unified framework linking cellular events to physiological outcomes in pollen-pistil interactions in Brassicaceae.
Findings
Initial recognition mechanisms distinguish self from nonself pollen in Brassicaceae.
Stigma responses like calcium signaling and reactive oxygen species regulate compatible and incompatible pollen interactions.
Understanding these mechanisms can improve breeding strategies for economically important Brassicaceae crops.
Abstract
Generation of competent offspring is vital for the prosperity of flowering plants. The pistil not only functions as a conduit for pollen tubes to grow to the ovary but also provides a selective venue for facilitating the growth of compatible pollen tubes and discouraging invaders and incompatible pollen. This review integrates recent advances in pollen–pistil interactions on dry stigmas of the Brassicaceae in the domains of self‐incompatibility (SI) and cross‐compatibility. We first outline the initial recognition mechanisms that distinguish self from nonself pollen and then highlight how key stigma responses are differentially regulated during compatible and incompatible responses, including calcium signaling, exocytosis, cytoskeleton dynamics, reactive oxygen species, aquaporin activity, and cell wall permeability. By linking these discrete cellular events to their physiological…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
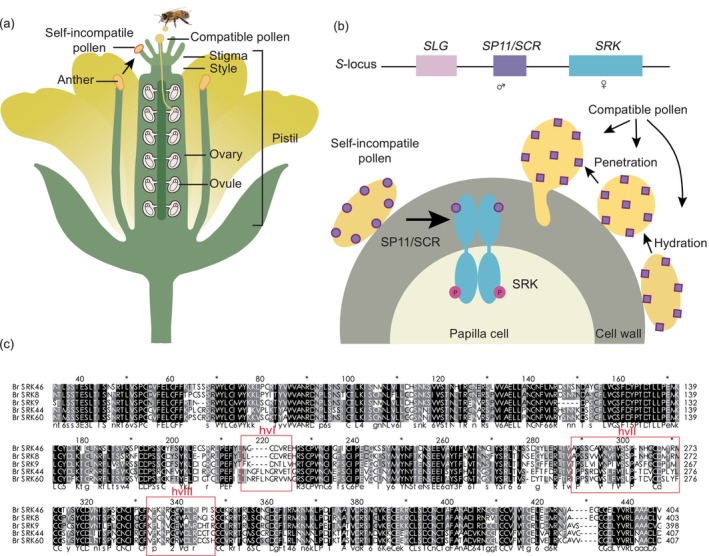 Fig. 1
Fig. 1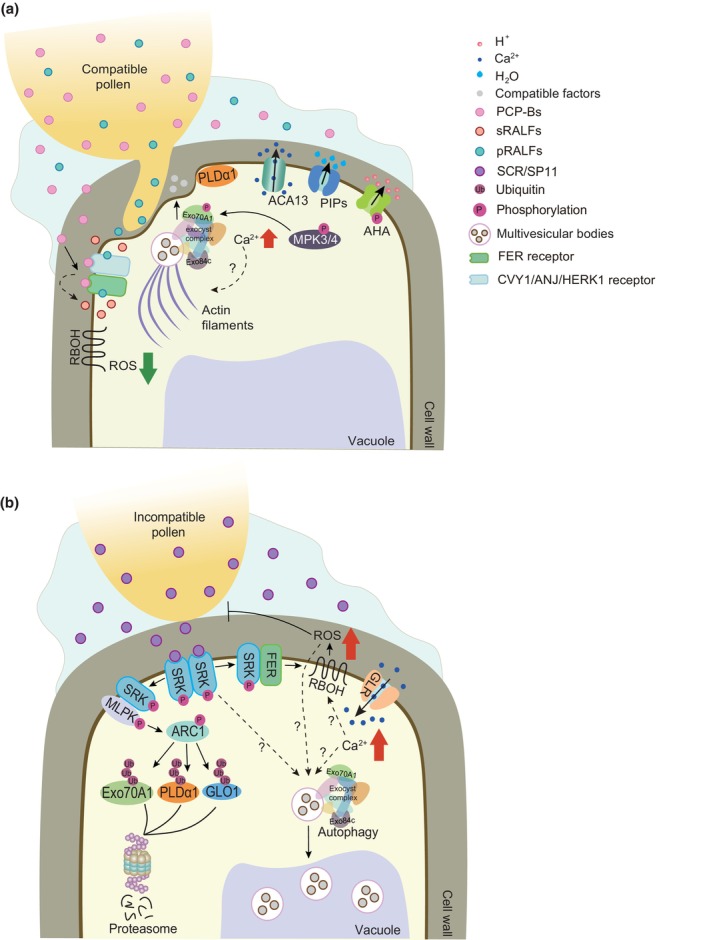 Fig. 2
Fig. 2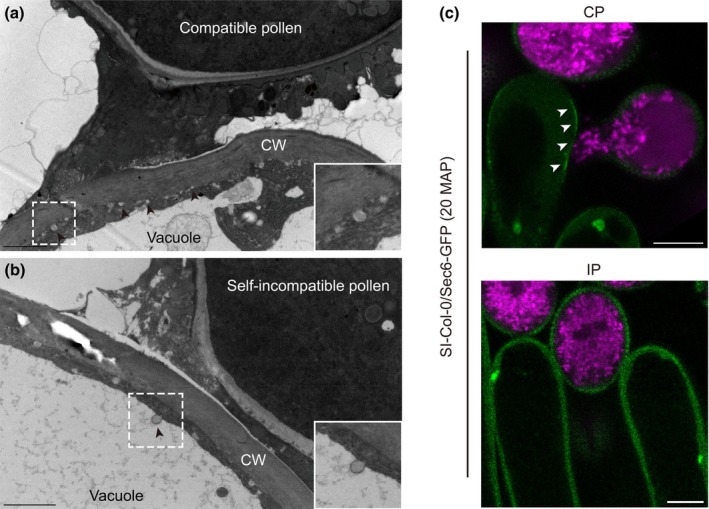 Fig. 3
Fig. 3- —China National Key Technology Research and Development Program
- —National Natural Science Foundation of China10.13039/501100001809
- —Taishan Scholar Foundation of Shandong Province10.13039/501100010029
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Reproductive Biology · Plant Molecular Biology Research · Plant and animal studies
Contents Summary 788I. Introduction 788II. The recognition of self‐incompatible pollen 790III. Basal compatible responses and their ‘suppression’ in self‐incompatible responses 790IV. Stigma barriers actively inhibiting pollen hydration and penetration 793V. Conclusions 795 Acknowledgements 795 References 796
Introduction
I.
In flowering plants, sexual reproduction is actively regulated by cell–cell interactions between the pollen (male reproductive cells released from the anther) and the pistil (the female reproductive organ; Fig. 1a). Successful fertilization begins at the receptive stigma surface of the pistil, in which pollen hydrates and germinates. The pollen tube then grows extracellularly through the stigma, style, and ovary to deliver sperm cells for double fertilization (Broz & Bedinger, 2021; Zhong et al., 2024). Throughout this journey, the pistil functions not merely as a passive conduit for pollen tube growth but also as an active gatekeeper (Bedinger et al., 2017; Broz & Bedinger, 2021; Zhong et al., 2025). Following pollination, two primary signaling pathways for pollen–pistil recognition are induced in the stigma: the compatibility pathway which ensures acceptance of suitable pollen, and the self‐incompatibility (SI) pathway which prevents inbreeding in outcrossing species (Takayama & Isogai, 2005; Doucet et al., 2016; Abhinandan et al., 2022). Among the several molecularly characterized SI systems in flowering plants – including gametophytic systems in the Papaveraceae and Solanaceae – the sporophytic system of the Brassicaceae provides a key model for dissecting pollen–stigma interactions at the stigma surface (Goring et al., 2023; Bosch & Franklin‐Tong, 2024; Xue, 2025). Notably, even during self‐incompatible pollinations, pollen is still recognized as ‘compatible’ and initiates basal compatible responses in the stigma. However, the self‐incompatible response always dominates, leading to pollen rejection (Jany et al., 2019).
Schematic demonstration of cross‐ and self‐pollination in Brassicaceae. (a) Schematic of a self‐incompatible Brassicaceae flower. Cross‐pollen hydrates, germinates, and penetrates the stigma to complete fertilization, while self‐pollen is rejected at the stigma surface. (b) The S‐locus. This comprises the female determinant SRK, the male determinant SP11/SCR, and the modulator S‐locus glycoprotein (SLG) (which enhances SRK expression). Haplotype‐specific binding of SP11/SCR to SRK activates the receptor, triggering incompatibility responses to reject self‐pollen. SLG, while not essential for recognition, enhances the self‐incompatibility reaction. (c) Sequence alignment of the extracellular domains of SRKs from different S‐haplotypes. The three hypervariable regions (hvI, hvII, and hvIII) critical for specific ligand binding are highlighted in red boxes.
Brassicaceae species have dry stigmas lacking abundant exudates; consequently, the hydration of pollen grains can only occur upon release of water and nutrients from the stigma (Dickinson, 1995). In this system, self‐pollen recognition is sporophytically controlled by the multi‐allelic S‐locus, which encodes tightly linked male‐ and female determinants (Fig. 1b). These allelic variants, known as S‐haplotypes, determine specificity: pollen carrying an S‐haplotype identical to that of the stigma, that is pollen from the same or a genetically closely related individual, is recognized as self‐pollen and triggers the SI response (Nasrallah & Nasrallah, 1993; Goring et al., 2023). Previous research has established key mechanisms that promote compatible factors during basal compatible responses and their suppression during SI response (Doucet et al., 2016; Nasrallah, 2023). Recent findings show that the stigma also constitutively maintains active barriers that inhibit pollen hydration and penetration (Liu et al., 2021; Zhang et al., 2021; Huang et al., 2023; Lan et al., 2023). Following pollen–stigma interaction, this barrier is differentially modulated: the recognition of compatible pollen weakens the inhibition to facilitate pollen tube growth, whereas the recognition of self‐incompatible pollen reinforces it (Liu et al., 2021; Zhang et al., 2021; Lan et al., 2023).
The Brassicaceae family includes economically important crops, such as Brassica rapa, Brassica oleracea, and Brassica napus, and various Arabidopsis species. While closely related to the outcrossing species Arabidopsis lyrata, the model plant Arabidopsis thaliana is fully self‐compatible (Nasrallah et al., 2004; Vekemans et al., 2014; Tsuchimatsu et al., 2017). The loss of SI in A. thaliana has been attributed to the mutation and deletion of male and/or female determinants (Bechsgaard et al., 2006; Shimizu et al., 2008; Tsuchimatsu et al., 2010, 2017; Vekemans et al., 2014). The introduction of functional S‐genes from A. lyrata or related species into A. thaliana has successfully restored its SI system (Nasrallah et al., 2002; Boggs et al., 2009b; Rea et al., 2010; Zhang et al., 2019). The transgenic restoration has helped overcome limitations in studying SI in nonmodel Brassica species, facilitating research into the molecular and cellular events in stigmatic papillae following compatible and incompatible pollination (Goring et al., 2023; Nasrallah, 2023).
In this review, we synthesize the complex molecular and cellular processes of pollen–stigma interactions in the Brassicaceae. As most findings derive from studies in Brassica species and Arabidopsis species, we refer to genes without specifying species names for conciseness. We focus on three integrated aspects: (1) the initial recognition of self‐incompatible pollen; (2) the regulation of stigma responses that promote compatible factors during basal compatible responses and suppress them during incompatible responses; and (3) the mechanisms by which the stigma barrier actively inhibits pollen hydration and penetration and how this inhibition is dynamically regulated.
The recognition of self‐incompatible pollen
II.
The receptive surface of the stigma is composed of numerous elongated papilla cells, each capable of accepting a pollen grain. Upon landing, proteins and lipids from the pollen coat mix with the stigmatic surface at the contact site, forming a specialized interface known as the ‘pollen foot’. This structure establishes the essential physical and hydraulic connection for pollen adhesion, creating the platform for the subsequent molecular dialogue of recognition (Roberts et al., 1980; Dickinson et al., 2000).
The recognition of self‐pollen is genetically controlled by a highly polymorphic S‐locus encoding tightly linked S‐genes expressed in both reproductive tissues (Suzuki et al., 1997). SP11/SCR is a small, cysteine‐rich protein localized in the pollen coat (Schopfer et al., 1999; Takayama et al., 2000). In plants heterozygous for two S‐haplotypes with a dominant‐recessive relationship, a pollen grain expresses only the dominant allele due to trans‐acting small RNA‐mediated silencing (e.g. Smi and Smi2 in Brassica; miR‐based systems in Arabidopsis) of recessive SP11/SCR alleles in the diploid tapetum (Tarutani et al., 2010; Durand et al., 2014; Yasuda et al., 2016; Bachmann et al., 2021; Dou et al., 2021, 2024; Yew et al., 2023). SRK (S‐locus receptor kinase) is a transmembrane receptor kinase in the papilla plasma membrane that acts as the receptor for its cognate SCR ligand (Stein et al., 1991; Goring & Rothstein, 1992; Takasaki et al., 2000). Another gene in the S‐locus is S‐locus glycoprotein (SLG), which is expressed in the stigmatic papilla cells. Although its precise role remains under investigation, SLG can enhance the SI response in B. rapa (Nasrallah et al., 1987; Takayama et al., 1987; Kandasamy et al., 1989).
At the contact site, this diffusible SCR signal binds with high specificity to its cognate SRK (Kachroo et al., 2001; Takayama et al., 2001; Shimosato et al., 2007). This haplotype‐specific interaction at the pollen–stigma interface is the foundation of the SI response (Fig. 1c). Hypervariable regions (hv) in both SP11/SCR and the extracellular domain of SRK are key for this interaction (Mishima et al., 2003; Kemp & Doughty, 2007; Boggs et al., 2009a; Ma et al., 2016). Structural analysis revealed that SCR interacts simultaneously with hvI and hvII of its cognate SRK, forming a 2 : 2, eSRK : SCR heterotetramer (Ma et al., 2016). This complex triggers SRK autophosphorylation and activation, initiating the downstream signaling cascades that lead to pollen rejection (Kachroo et al., 2001; Takayama et al., 2001). If the pollen is not recognized as self – that is, if the SCR signal and SRK receptor are not from the same S‐locus, or if one or both of the S‐determinants are mutated or lost – basal compatible responses ultimately promote the hydration and penetration of compatible pollen (Jany et al., 2019).
Basal compatible responses and their ‘suppression’ in self‐incompatible responses
III.
1. Calcium signaling
Calcium (Ca^2+^) is a fundamental secondary messenger known to regulate pollen tube growth (Lin et al., 2025). During pollen–stigma interactions, distinct dynamic changes in stigmatic Ca^2+^ levels are observed (Dearnaley et al., 1997; Iwano et al., 2004, 2014, 2015), which are thought to be associated with either pollen acceptance or rejection.
During compatible pollination, given that the Ca^2+^ gradient at the tip region of the pollen tube is essential for its growth, it is speculated that Ca^2+^ is pumped out from the papilla cell (Iwano et al., 2014; Lin et al., 2025). Experiments coating the B. rapa and A. thaliana stigmas with the Ca^2+^ indicator Calcium Green revealed elevated Ca^2+^ levels on the papilla surface following cross‐pollination, but not following self‐incompatible pollination. This Ca^2+^ increase is dependent on the Ca^2+^ pump ACA13 (autoinhibited Ca^2+^‐ATPase 13), as mutants lacking ACA13 failed to exhibit this response, leading to impaired pollen germination and reduced seed set. Together, these results indicate that ACA13‐mediated Ca^2+^ export from papillae is essential for compatible pollen growth, although precisely how this surface Ca^2+^ increase facilitates pollen acceptance remains unknown (Iwano et al., 2014; Fig. 2a).
Model of signaling pathways during compatible and self‐incompatible responses in Brassicaceae. (a) Compatible response. A slight increase in papilla cytosolic calcium (Ca2+) is observed at the pollen contact site, and ACA13 is supposed to be responsible for Ca2+ export to support pollen germination. The compatible factor Exo70A1 is phosphorylated by mitogen‐associated protein kinase 3/4 (MPK3/4) and trafficked to the papilla plasma membrane. PLDα1 produces phosphatidic acid (PA) to facilitate vesicle tethering. Actin filaments are reorganized, a process suggested to support vesicle transport. Pollen‐derived pollen coat‐derived cysteine‐rich peptides (PCP‐Bs) and pRALFs compete for binding to CrRLK1L receptor complexes, leading to a decrease in stigmatic reactive oxygen species (ROS) and promoting pollen tube penetration. Concurrently, plasma membrane H+‐ATPases (AHAs) are activated, pumping H+ out of the cell. This elevates cytosolic pH, promoting water export through plasma membrane intrinsic proteins (PIPs). (b) Self‐incompatible response. GLR channels mediate Ca2+ influx, although the upstream activating signal remains unknown. SCR binding induces SRK autophosphorylation and activation. Activated SRK interacts with M‐locus protein kinase (MLPK) at the plasma membrane. Together, they activate the E3 ubiquitin ligase ARM‐repeat containing (ARC1), which targets and degrades downstream compatible components (Exo70A1, PLDα1, and GLO1). Exocyst complex subunits are also degraded via Exo84c‐mediated autophagy. Simultaneously, SRK interacts with FERONIA (FER), which further activates RBOH‐mediated ROS production, blocking pollen hydration and germination. PLD, phospholipase D; GLO1, Glyoxalase 1. Solid arrows represent experimentally confirmed pathways, while dashed arrows represent predicted potential pathways that have not been experimentally verified.
During self‐incompatible pollination, A. thaliana expressing the Ca^2+^ probe YC3.6 showed a rapid and large Ca^2+^ influx into stigmatic papilla cells, but compatible pollination only triggered a small Ca^2+^ increase (Iwano et al., 2015). This influx is mediated primarily by glutamate receptor‐like (GLR) channels (Iwano et al., 2015). A glr3.7 knockout mutant shows a diminished stigmatic Ca^2+^ elevation, while application of GLR agonists (e.g. D‐Ser, D‐Glu) elevated cytosolic [Ca^2+^] levels in wild‐type (WT) Arabidopsis (Iwano et al., 2015; Fig. 2b). However, how the GLR channels are activated by the initial SCR‐SRK recognition remains unclear.
Thus, Ca^2+^ signaling bifurcates at the stigma: compatible pollination triggers efflux (via ACA13) to nourish the pollen, while incompatible pollination induces influx (via GLRs) to initiate rejection. This Ca^2+^ signal likely acts upstream and integrates with other rejection mechanisms (Bosch & Franklin‐Tong, 2024; Li et al., 2024; Zhou et al., 2024). Establishing the specific signaling networks in the Brassicaceae during compatibility responses and SI responses remains a key area for future research (Fig. 2b).
2. Compatible factors
Successful hydration and germination of compatible pollen depends on the export of water and compatible factors from papilla cells (Dickinson, 1995). Although the nature of compatible factors remains unknown, they are likely components that promote the compatible responses. It is hypothesized that these compatible factors are delivered to the stigmatic papillae surface by secretory vesicles. Cytoplasmic streaming and vesicle‐like structures have been observed in B. oleracea stigmatic papillae upon compatible response (Dickinson, 1995; Elleman & Dickinson, 1996; Fig. 3a,b). However, these secretory events are specifically abrogated upon pollination with self‐incompatible pollen, effectively halting the delivery of resources to the pollen grain (Elleman et al., 1992; Safavian & Goring, 2013; Fig. 2a,b). Thus, the regulation of vesicle‐mediated secretion represents a key cellular checkpoint determining pollen fate.
Vesicle trafficking during pollen–stigma interactions in Brassicaceae. (a, b) Transmission electron microscopy (TEM) images of Brassica napus stigmatic papillae following compatible (a) and incompatible pollination (b). After compatible pollination, vesicle‐like structures (indicated by arrowheads) are observed fusing to the plasma membrane (PM). Following incompatible pollination, multivesicular bodies or autophagosomes (arrowheads) are observed fusing with the vacuolar membrane, suggesting the degradation of compatible components in the vacuole. The dashed squares indicate the secretory vesicles (a) or multivesicular bodies/autophagosomes (b) are shown in the lower right corner at an enlarged scale. Bars, 2 μm. CW, cell wall. (c) Exocytosis is monitored by Sec6‐GFP upon compatible (CP) and incompatible pollination (IP). The image, reprinted from Zhang et al. (2024), shows that exocytosis is activated during compatible pollination, with Sec6‐GFP signal accumulated at the PM adjacent to pollen (white arrowheads). By contrast, the Sec6‐GFP signal is distributed evenly in the cytoplasm after incompatible pollination. Bars, 10 μm. CP, compatible pollination. IP, incompatible pollination. MAP, minutes after pollination. GFP, green fluorescent protein.
Regulation of compatible factors in compatible responses
The secretory vesicles delivering compatible factors rely on the exocyst complex – a complex containing eight subunits, Sec3, Sec5, Sec6, Sec8, Sec10, Sec15, Exo70, and Exo84 – for tethering to the plasma membrane of stigmatic papilla (Chong et al., 2010). Live‐cell imaging shows that Sec6‐GFP‐ and GFP‐Exo70A1‐labelled exocytic vesicles accumulate at the papilla plasma membrane adjacent to pollen grains during compatible, but not incompatible, pollinations (Zhang et al., 2024; Fig. 3c). The trafficking of Exo70A1 is activated following its phosphorylation by mitogen‐associated protein kinase 3 (MPK3)/4 (Jamshed et al., 2020; Fig. 2a). Silencing Exo70A1 or other subunits of the exocyst complex impairs compatible pollen hydration in A. thaliana (Samuel et al., 2009; Safavian et al., 2015), confirming the function of exocyst complex in vesicle transport during compatible pollen hydration.
Vesicle fusion is further regulated by phospholipase D (PLD), which produces phosphatidic acid (PA), a key messenger for vesicle fusion to target membrane (Ammar et al., 2013). PLDα1 activity is crucial for hydration and pollen tube germination in compatible responses (Scandola & Samuel, 2019). Overexpressing PLDα1 or exogenous application of PA to B. napus stigmas breaks down the SI and promotes the growth of incompatible pollen tubes into the papillae (Scandola & Samuel, 2019; Fig. 2a). Optimal PLDα1 activity requires cytosolic Ca^2+^, suggesting that a compatible pollination‐induced Ca^2+^ increase may activate PLDα1 to promote vesicle fusion (Hong et al., 2016). However, the specific cargos transported by these vesicles and the upstream signals that initiate their delivery remain unidentified.
In addition to vesicle trafficking, papilla cells must maintain a suitable intracellular environment. Glyoxalase 1 (GLO1), which detoxifies the byproduct of glycolysis (methylglyoxal, MG), is identified as another key compatible factor (Sankaranarayanan et al., 2015). Overexpression of GLO1 partially breaks down SI of B. napus, while its silencing reduces seed set (Sankaranarayanan et al., 2015). It remains unclear whether GLO1 activity is upregulated after compatible pollination and how it is regulated.
Degradation of compatible factors in self‐incompatible responses
In contrast to the compatible response, the secretory system responsible for delivering compatible factors is inhibited in stigmatic papillae after incompatible pollination (Elleman et al., 1992; Safavian & Goring, 2013; Zhang et al., 2024). This inhibition is executed through two parallel degradation pathways targeting compatible factors.
One is the ubiquitin‐proteasome pathway. Following the recognition of self‐incompatible pollen, the activated SP11/SCR‐SRK complex triggers signaling events in the stigmatic papilla that cause pollen rejection (Fig. 2b). The downstream signaling proteins have proven to be rather elusive and so far, only two SRK‐interacting proteins were shown to function in this response: the M‐locus protein kinase (MLPK) and the ARM‐repeat containing 1 (ARC1) E3 ligase (Murase et al., 2004; Abhinandan et al., 2022). MLPK is phosphorylated by SRK after incompatible pollination (Murase et al., 2004; Kakita et al., 2007; Chen et al., 2019). Then, SRK and MLPK together phosphorylate ARC1 (Samuel et al., 2009; Scandola & Samuel, 2019). After phosphorylation, activated ARC1 relocates to the proteasome, mediating the ubiquitin‐dependent degradation of specific downstream compatible components, including Exo70A1, PLDα1, and GLO1 (Stone et al., 2003; Samuel et al., 2008; Abhinandan et al., 2022; Fig. 2b). The degradation of GLO1 leads to cytotoxic MG accumulation (Sankaranarayanan et al., 2015). Knocking down or knocking out ARC1 breaks down the SI response in both B. napus and Arabidopsis (Stone et al., 1999; Abhinandan et al., 2023), confirming the role of the ubiquitin‐proteasome pathway in pollen rejection during SI.
The other is the autophagy‐mediated vacuolar degradation. Autophagy is a major inducible degradation pathway that participates in plant development and stress response (Zhuang et al., 2024). This pathway is also recruited during self‐incompatible pollination, which induces the accumulation of autophagosomes in the vacuole (Safavian & Goring, 2013; Goring, 2017). Loss‐of‐function mutants of the key autophagy genes ATG5 and ATG7 partially break down the self‐incompatible response in A. thaliana (Macgregor et al., 2022). Recent work identified the exocyst complex subunit Exo84c as a critical mediator of this pathway. Exo84c promotes the autophagic degradation of other exocyst complex subunits (e.g. Sec6, Exo70A1) upon incompatible pollination. An exo84c knockout mutant delays the degradation of Sec6 and Exo70A1 (Zhang et al., 2024). This demonstrates that the exocyst complex subunits are the direct targets of autophagy during SI responses (Fig. 2b).
While autophagy is clearly engaged, its upstream induction signal in self‐incompatible responses remains unknown. Given that intracellular Ca^2+^ is a key regulator of autophagy in mammals (Høyer‐Hansen et al., 2007; Decuypere et al., 2011; Li et al., 2017; Saikia & Joseph, 2021; Zheng et al., 2022; Liu et al., 2023), it will be interesting to investigate whether cytosolic Ca^2+^ promotes autophagy in Brassicaceae self‐incompatible responses.
Cytoskeleton
The plant cytoskeleton, comprising actin filaments and microtubules, plays central roles in cell shape maintenance, vesicle trafficking, cell division, primary cell wall assembly, and immunity (Chebli et al., 2021; Sinha et al., 2024; Schoofs et al., 2025).
During compatible pollinations, papilla cells rapidly reorganize their actin cytoskeleton. In B. rapa, rhodamine‐phalloidin staining reveals that actin bundles polymerize and accumulate in the cytoplasm of papilla cells adjacent to pollen grains (Iwano et al., 2007). Similar actin dynamic changes have also been observed in Arabidopsis expressing LifeAct‐Venus (Rozier et al., 2020; Fig. 2a). This reorganization is absent in papillae cells during incompatible pollination (Iwano et al., 2007). Disrupting actin dynamics with cytochalasin D inhibits compatible pollen hydration and germination, confirming that actin polymerization is essential for promoting a compatible response (Iwano et al., 2007).
The functional significance of this actin remodeling is likely twofold. First, it may facilitate vesicle trafficking required for exocytosis of compatible factors. In B. rapa papillae, compatible pollination induces actin polymerization associated with an apical vacuole network, which is suggested to support pollen hydration (Iwano et al., 2007). Second, it could represent a mechanosensitive response to the pressure at the pollen tube penetration site, similar to actin accumulation at pathogen contact sites (Takemoto & Hardham, 2004; Hardham et al., 2008).
Microtubules have also been proposed to participate in pollen–stigma interactions. In Arabidopsis, the orientation of cortical microtubules regulates the direction of pollen tube growth on stigmatic papillae by influencing the deposition of cellulose microfibrils in the cell wall (Riglet et al., 2020). Mutants for KATANIN 1 (ktn1), a microtubule‐severing enzyme, exhibit disordered cortical microtubules and cellulose microfibrils. This leads to a loss of mechanical anisotropy in the papilla cell wall, which causes compatible pollen tubes to coil around the papillae instead of penetrating them. A similar coiled phenotype was observed in aged WT papillae with disordered cellulose microfibril pattern, confirming the importance of microtubule‐guided wall mechanics for proper pollen tube guidance (Riglet et al., 2020).
This cytoskeletal remodeling is likely coordinated with Ca^2+^ signaling. A well‐established reciprocal crosstalk exists, where the cytoskeleton influences Ca^2+^ homeostasis, and Ca^2+^ signals, in turn, regulate cytoskeletal dynamics (Naveed et al., 2024). This interaction could integrate the mechanical and chemical signals during compatibility. The availability of live‐cell markers for both actin/microtubule and secretory vesicles now enables direct testing of this model to determine how cytoskeletal reorganization affects vesicle delivery.
Stigma barriers actively inhibiting pollen hydration and penetration
IV.
Conceptually, beyond the suppression of basal compatible responses during self‐incompatible responses, the stigma also imposes selective barriers actively inhibiting pollen hydration and pollen tube entry. These barriers are maintained by stigmatic signaling modules that are weakened by specific pollen‐derived compatible signals and reinforced by specific pollen‐derived incompatible signals.
1. Basal barrier maintenance via endogenous sRALF–CrRLK1L modules
A key regulatory node involves the Catharanthus roseus receptor‐like kinase 1‐like (CrRLK1L) members, FERONIA (FER)/SIRÈNE and ANJEA (ANJ), which, together with LORELEI‐like GPI‐anchored protein 1 (LLG1), form receptor complexes on the papilla cell surface (Huck et al., 2003; Rotman et al., 2003; Escobar‐Restrepo et al., 2007; Duan et al., 2010, 2014, 2020; Li et al., 2015; Cheung, 2024). These complexes bind the stigma autocrine RAPID ALKALINIZATION FACTOR 23/33 (sRALF23/33) peptides and maintain a basal inhibitory stigma barrier before pollination (Liu et al., 2021; Zhang et al., 2021; Lan et al., 2023). The sRALF–CrRLK1L interaction maintains this barrier by initiating three pathways involving reactive oxygen species (ROS) accumulation, aquaporin gating (water channel), and cell wall permeability.
Reactive oxygen species accumulation
ROS plays important roles in diverse physiological processes, but high levels of ROS burst are a common defense mechanism against pathogens (Qi et al., 2017; Otulak‐Kozieł et al., 2020). In the stigma, the sRALF–CrRLK1L interaction activates downstream NADPH oxidase RESPIRATORY‐BURST OXIDASE HOMOLOG D (RBOHD), leading to constitutive accumulation of ROS. This basal level of ROS in unpollinated stigmas is inhibitory for pollen hydration (Liu et al., 2021; Zhang et al., 2021). Accordingly, stigmas of fer‐4 mutant or rbohD mutant, which have reduced levels of stigmatic ROS, exhibit faster hydration of compatible pollen (Liu et al., 2021; Zhang et al., 2021).
Aquaporin closure
On the dry stigma of Brassicaceae, pollen hydration depends on effective water transport from the stigmatic papilla cells, a process facilitated by plasma membrane aquaporins, such as plasma membrane intrinsic proteins (PIPs; Windari et al., 2021). Notably, many PIPs are gated by cytosolic pH, undergoing conformational closure upon protonation (Törnroth‐Horsefield et al., 2006; Anderberg et al., 2011; Groszmann et al., 2017). The sRALF–CrRLK1L module inhibits plasma membrane H^+^‐ATPases (AHAs), preventing proton efflux. The resulting lower cytosolic pH promotes the protonation and closure of aquaporins (e.g. PIP1;2), effectively blocking water release (Liu et al., 2025). Consistent with this, ralf33 mutant stigmas, which have a higher stigmatic pH before pollination, facilitate faster pollen hydration (Liu et al., 2025).
Cell wall remodeling
After pollen germination, the pollen tube must penetrate the papilla cell wall. FER and other CrRLK1L family members are known to be important for maintaining cell wall integrity (Cheung & Wu, 2011; Duan et al., 2020; Liu et al., 2024). The cell wall barrier is maintained by a broader CrRLK1L module involving FER, ANJ, HERCULES1 (HERK1), CURVY1 (CVY1), together with four stigmatic autocrine RALF peptides (sRALFs: RALF1, RALF22, RALF23, and RALF33; Lan et al., 2023). Stigmas of mutants deficient in key components – such as fer, cvy1, anj, herk1, and a ralf quad mutant (which carries mutations in all four sRALFs) – are unable to maintain the cell wall barrier, allowing the penetration of interspecific pollen tubes that would be rejected by WT stigmas (Lan et al., 2023).
A second layer of control involves the transcription factor STIGMATIC PRIVACY 2 (SPRI2) and its homolog SPRI2‐like, which promote the expression of the cell wall biosynthesis enzymes TBL40 and TBL45. The finding that mutants for either SPRI2 or TBL genes compromise the barrier – allowing interspecific pollen tube penetration – directly implicates this transcriptional module in fortifying the stigma cell wall during pollination (Fujii et al., 2023).
2. Weakening the barrier by compatible pollen‐derived signals
To allow pollen hydration and penetration, the stigma barriers – ROS accumulation, aquaporin closure and cell wall permeability – must be weakened. This process is triggered by signals from compatible pollen.
Reducing ROS
Following compatible pollination, a rapid reduction in stigmatic ROS is essential for successful pollen hydration and pollen tube growth (Liu et al., 2021; Zhang et al., 2021). Compatible pollen employs at least three strategies to achieve this.
Pollen coat‐derived cysteine‐rich peptides (PCP‐Bs) are crucial regulators of pollen hydration (Wang et al., 2017). PCP‐Bs compete with stigma RALF23/33 for binding to CrRLK1L receptors, preventing the activation of RBOHD‐mediated ROS production and thus leading to pollen hydration (Wang et al., 2017; Liu et al., 2021; Fig. 2a). PCP‐B peptides also trigger high levels of nitric oxide (NO) in the stigma, which is intimately engaged with ROS (Domingos et al., 2015). Stigma NO triggered by compatible pollen inactivates FER and RBOHD via nitrosation, shutting down the core ROS‐production machinery (Huang et al., 2023). Scavenging stigmatic NO causes ROS accumulation and inhibits compatible pollen tube growth (Huang et al., 2023). Furthermore, ROS homeostasis is regulated by several scavenging enzymes. For example, knocking out Ascorbate peroxidase 1 (APX1), a key enzyme that detoxifies hydrogen peroxide (H_2_O_2_), in B. napus stigmas leads to delayed compatible pollen hydration (Liang et al., 2024). However, how the activity of scavenging enzymes is regulated during compatibility responses remains unclear. The structural mechanisms by which PCP‐Bs outcompete sRALFs for binding with the receptor complex await elucidation.
Opening aquaporin water channels
Water channels are also opened by signals from compatible pollen. The replacement of sRALFs by PCP‐Bs activates plasma membrane AHA activity, which pumps protons out of the papilla cell. This elevates the cytosolic pH near the plasma membrane, causing deprotonation and the opening of aquaporins (PIP1/2s). Targeted water efflux is thereby to facilitate pollen hydration (Wang et al., 2017; Liu et al., 2021; Liu et al., 2025; Fig. 2a).
Loosening the cell wall barrier
A distinct set of pollen‐derived RALF peptides (pRALFs) targets the cell wall permeability maintained by the sRALFs–CrRLK1L module. These pRALFs compete with sRALFs (RALF1, RALF22, RALF23, RALF33) for binding to a set of CrRLK1L receptors (FER/ANJ/HERK1/CVY1). This ligand switch is proposed to modify the papilla cell wall, possibly through interactions with leucine‐rich repeat extensin (LRX) proteins, making it locally permeable to pollen tube penetration (Lan et al., 2023; Zhong et al., 2025; Fig. 2a). In practice, compatible pollen or application of pRALFs from compatible pollen helps interspecific pollen overcome the hybridization barrier on the stigma. Future research to elucidate the complete structures of these ligand–receptor complexes will reveal how the RALF–CrRLK1L–LLG module is assembled and how ligand–receptor specificity is achieved.
3. Reinforcing the barrier by signals from incompatible pollen
In contrast to compatibility, self‐incompatible or interspecific pollination actively reinforces the barrier. The binding of incompatible pollen signals – such as the self‐pollen ligand SCR or the interspecific pollen signal SRK‐interacting Interspecific Pollen Signal – to the stigma SRK receptor enhances the interaction between SRK and FER. This rapidly activates the ROS‐production pathway, triggering a strong oxidative burst that irreversibly arrests incompatible pollen (Zhang et al., 2021; Huang et al., 2023; Cao et al., 2025; Fig. 2b). In practice, spraying stigmas with ROS scavengers effectively breaks down the self‐incompatible response, permitting the growth of incompatible pollen and enabling seed production from otherwise sterile self‐incompatible inbred lines (Zhang et al., 2021).
The inhibitory effect of the elevated stigmatic ROS on pollen was demonstrated by a ‘stigma transfer experiment’ (Zhang et al., 2021). In this assay, stigmas were pollinated with self‐pollen to induce ROS elevation and then transferred at different time points (0–60 min) to a culture medium containing ROS scavengers. The results showed a sharp, time‐dependent decline in rescue: immediate transfer (0 min) allowed c. 80 self‐pollen tubes to grow, but delays of 15 and 30 min reduced this number to only 42 and 10 tubes, respectively. This stands in contrast to the complete rejection of self‐pollen observed in nontransferred, self‐incompatible stigma (Zhang et al., 2021). These results suggest that prolonged exposure to high ROS levels irreversibly arrests self‐pollen development on the stigma.
Mechanistically, the FER‐mediated activation of ROS production may be linked to a broader signaling cascade. In other contexts, stress‐induced increases in cytosolic Ca^2+^ are known to activate NADPH oxidases (Arfaoui et al., 2018; Ravi et al., 2023). It is therefore plausible that during the SI response in Brassicaceae, Ca^2+^ influx into papilla cells serves as the upstream signal that induces the ROS burst.
An important open question is whether signals from self‐incompatible pollen and interspecific pollen also enhance the water channel and the stigma cell wall barrier. Ca^2+^ influx is suggested to be the upstream signal for cytosolic acidification, which might be responsible for aquaporin inactivation in rejecting undesired pollen (Bosch & Franklin‐Tong, 2024; Wang et al., 2024, 2025). Future studies should determine whether these pathways are similarly amplified during incompatibility responses.
Conclusions
V.
In the Brassicaceae, pollen–stigma interactions leading to either acceptance or rejection follow a conserved sequence: Initial signal recognition is followed by the execution of a specific cellular response.
The compatible response orchestrates pollen acceptance by coordinating the stigmatic supply of water and essential factors with the active dismantling of inhibitory barriers, including reducing ROS and weakening the cell wall. Conversely, the incompatible response utilizes a parallel yet opposing pathway, blocking this supportive supply while simultaneously amplifying inhibitory signals to enforce pollen rejection.
Strikingly, the signaling architecture is not unique to the Brassicaceae stigmas but shares deep similarities with the SI system in Papaveraceae and with the plant innate immune system (Kessler et al., 2010; Goring et al., 2023). Despite utilizing distinct ligand–receptor pairs for initial recognition, these pathways converge on conserved downstream events, including Ca^2+^ influx, ROS bursts, cytoskeletal remodeling, and targeted trafficking (Qi et al., 2017; Wang et al., 2022; Goring et al., 2023; Ravi et al., 2023; Bosch & Franklin‐Tong, 2024; Naveed et al., 2024). This mechanistic overlap suggests plants that employ a modular and economical signaling toolkit to respond to diverse external stimuli, whether from pollen or pathogens. Elucidating the evolutionary origin of these SI systems, whether potentially derived from ancient defense‐related recognition pathways, will be a compelling challenge for the future.
Despite significant progress, key questions remain. The molecular basis for how SI signaling decisively overrides compatible pathways is still unknown, and the cross‐regulation between the various downstream cellular events is poorly understood. A more comprehensive understanding of these processes is not only fundamental to plant biology but also of direct practical importance. SI is a widely used tool in breeding programs to produce F_1_ hybrid seeds. Elucidating its mechanisms will therefore provide new strategies for manipulating fertility and improving crop breeding in economically important Brassicaceae species.
Competing interests
None declared.
Disclaimer
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abhinandan K , Hickerson NMN , Lan X , Samuel MA . 2023. Disabling of ARC 1 through CRISPR‐Cas 9 leads to a complete breakdown of self‐incompatibility responses in Brassica napus . Plant Communications 4: 100504.36518081 10.1016/j.xplc.2022.100504 PMC 10030360 · doi ↗ · pubmed ↗
- 2Abhinandan K , Sankaranarayanan S , Macgregor S , Goring DR , Samuel MA . 2022. Cell‐cell signaling during the Brassicaceae self‐incompatibility response. Trends in Plant Science 27: 472–487.34848142 10.1016/j.tplants.2021.10.011 · doi ↗ · pubmed ↗
- 3Ammar MR , Kassas N , Chasserot‐Golaz S , Bader MF , Vitale N . 2013. Lipids in regulated exocytosis: what are they doing? Frontiers in Endocrinology 4: 125.24062727 10.3389/fendo.2013.00125 PMC 3775428 · doi ↗ · pubmed ↗
- 4Anderberg HI , Danielson J , Johanson U . 2011. Algal MI Ps, high diversity and conserved motifs. BMC Evolutionary Biology 11: 110.21510875 10.1186/1471-2148-11-110PMC 3111385 · doi ↗ · pubmed ↗
- 5Arfaoui A , El Hadrami A , Daayf F . 2018. Pre‐treatment of soybean plants with calcium stimulates ROS responses and mitigates infection by Sclerotinia sclerotiorum . Plant Physiology and Biochemistry 122: 121–128.29223021 10.1016/j.plaphy.2017.11.014 · doi ↗ · pubmed ↗
- 6Bachmann JA , Tedder A , Fracassetti M , Steige KA , Lafon‐Placette C , Köhler C , Slotte T . 2021. On the origin of the widespread self‐compatible allotetraploid Capsella bursa‐pastoris (Brassicaceae). Heredity 127: 124–134.33875831 10.1038/s 41437-021-00434-9PMC 8249383 · doi ↗ · pubmed ↗
- 7Bechsgaard JS , Castric V , Charlesworth D , Vekemans X , Schierup MH . 2006. The transition to self‐compatibility in Arabidopsis thaliana and evolution within S‐haplotypes over 10 Myr. Molecular Biology and Evolution 23: 1741–1750.16782760 10.1093/molbev/msl 042 · doi ↗ · pubmed ↗
- 8Bedinger PA , Broz AK , Tovar‐Mendez A , Mc Clure B . 2017. Pollen‐pistil interactions and their role in mate selection. Plant Physiology 173: 79–90.27899537 10.1104/pp.16.01286 PMC 5210727 · doi ↗ · pubmed ↗