Multifaceted roles of BBX transcription factors: impacts on key agronomical traits and environmental resilience
Bruno Silvestre Lira, Juliene Moreira, Luciano Freschi, Magdalena Rossi

TL;DR
This review explores how BBX proteins influence plant growth, stress responses, and crop traits, offering potential for improving agricultural productivity.
Contribution
The paper highlights the multifaceted roles of BBX proteins in integrating light and hormone signaling for agronomic and environmental resilience.
Findings
BBX proteins modulate flowering time, fruit and seed production in crops.
They integrate light and hormone signaling to enhance plant resilience to stresses.
BBX factors influence specialized metabolism and signal transduction pathways.
Abstract
B‐box (BBX) proteins were initially characterized as transcription factors connecting light signaling to the regulation of flowering time and seedling photomorphogenesis. However, over the last decade, increasing evidence has shown that they integrate light and hormone signaling, modulating multiple physiological processes during plant life. In this review, we provide an overview of the structure of this protein family and the function of its members in Arabidopsis thaliana model species. Then, we specifically discuss the role of BBX factors in controlling agronomically important traits in crop species, including flowering time, fruit and seed production, responses to biotic and abiotic stresses, and specialized metabolism. Finally, we address several aspects that require further investigation to dissect the mechanisms behind the multifaceted roles of BBX proteins and their implications…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
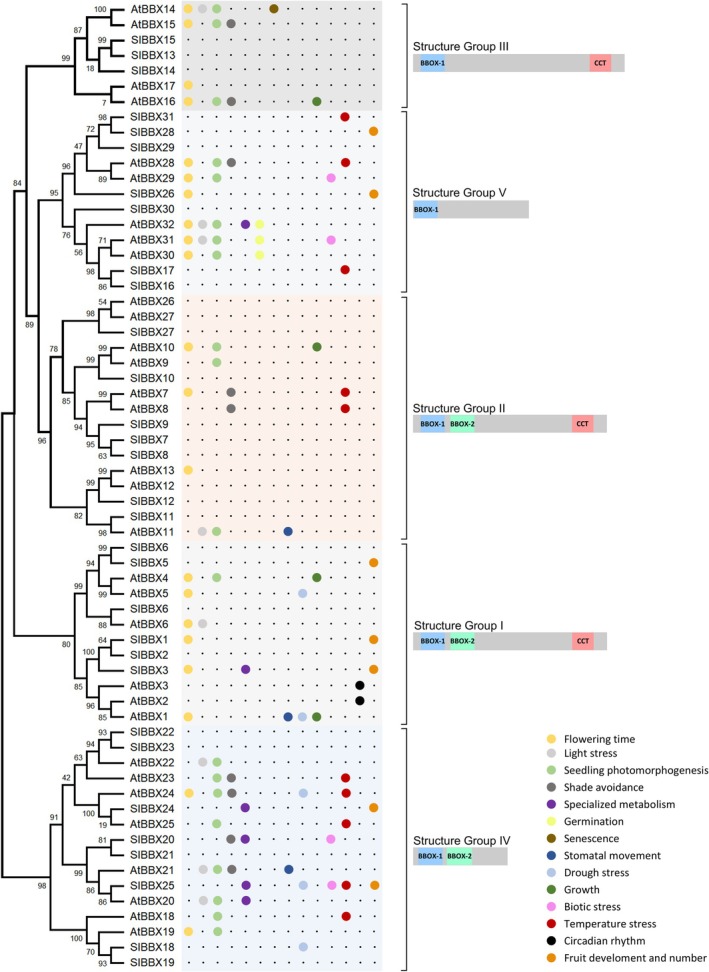 Fig. 1
Fig. 1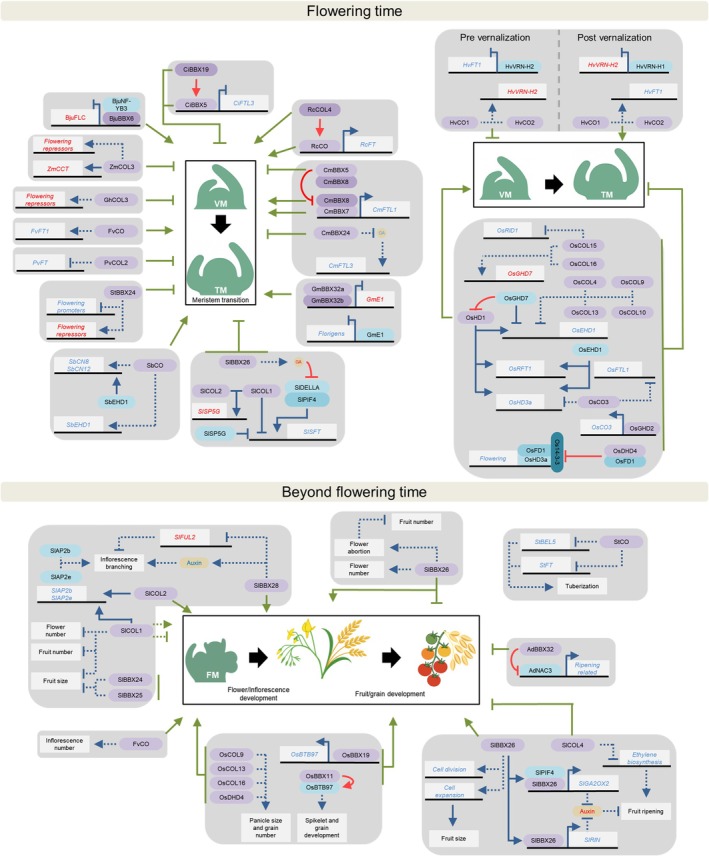 Fig. 2
Fig. 2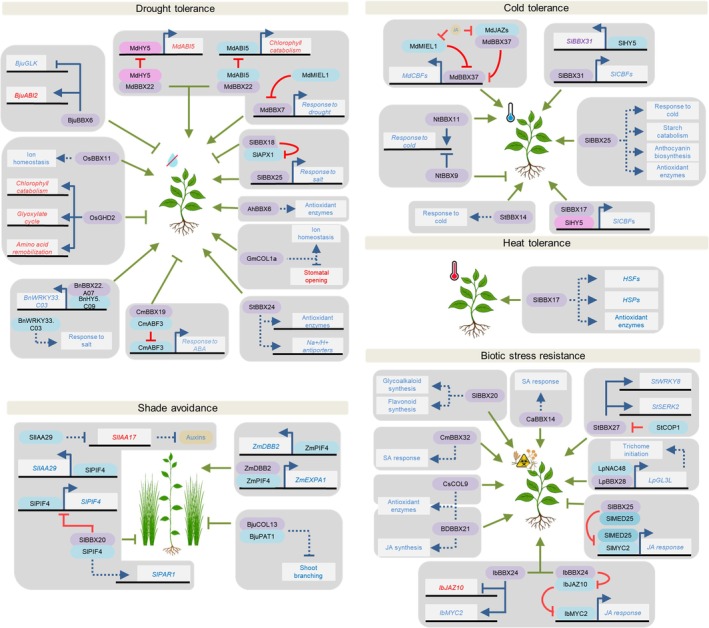 Fig. 3
Fig. 3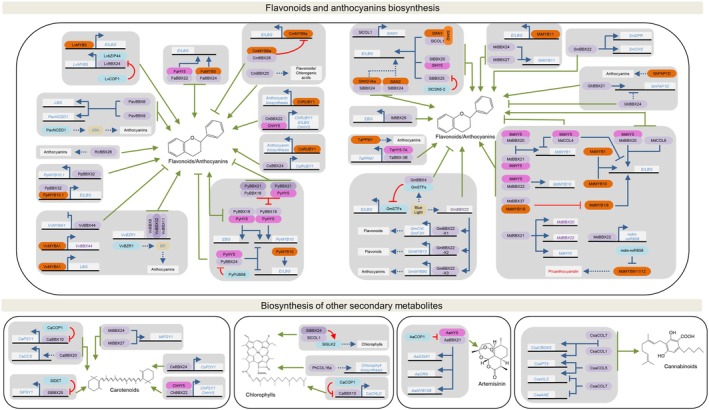 Fig. 4
Fig. 4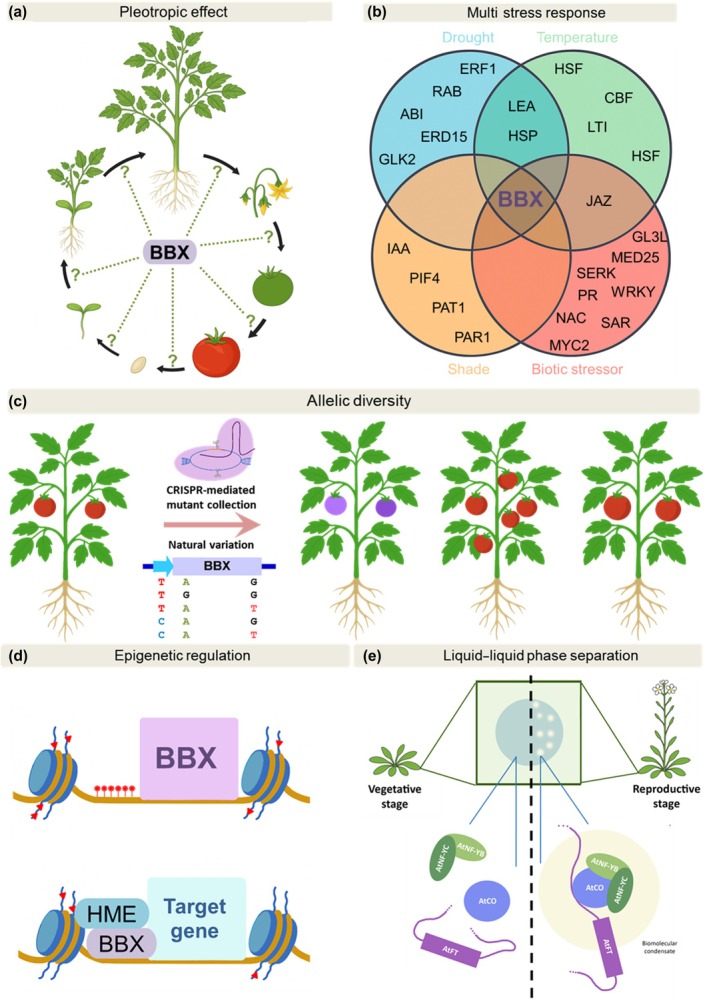 Fig. 5
Fig. 5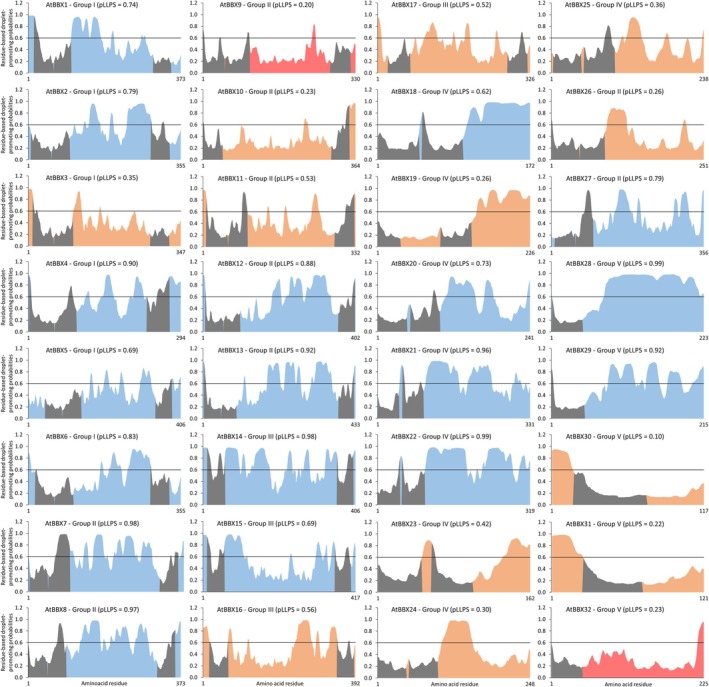 Fig. 6
Fig. 6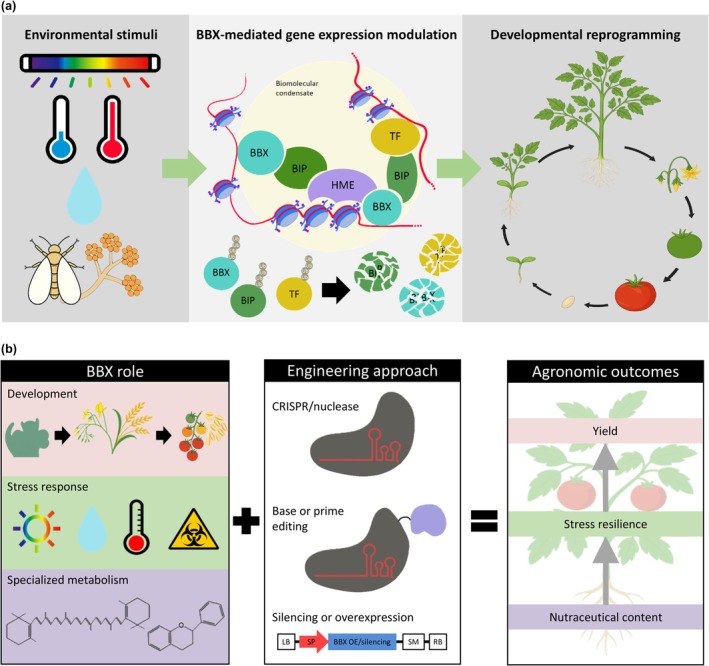 Fig. 7
Fig. 7| Species | Common name | BBX | Function | References |
|---|---|---|---|---|
|
| ||||
|
| ||||
|
| Chinese mustard | BjuBBX6 | Positive | Feng |
|
| Canola | BnCOa1 | Positive | Robert |
|
| Chrysanthemum | CiBBX5 | Negative | C. Yang |
| CiBBX19 | Negative | C. Yang | ||
|
| Chrysanthemum | CmBBX5 | Negative | Wang |
| CmBBX7 | Positive | Zhai | ||
| CmBBX8 | Positive | Wang | ||
| Zhai | ||||
| CmBBX24 | Negative | Y. Yang | ||
|
| Strawberry | FvCO | Positive | Kurokura |
|
| Soybean | GmCOL5 | Positive | Fan |
| GmBBX32a | Negative | C. Gao | ||
| GmBBX32b | Negative | C. Gao | ||
|
| Cotton | GhCOL1A | Positive | Cai |
| GhCOL1D | Positive | Cai | ||
| GhCOL3 | Negative | Z. Song | ||
|
| Barley | HvCO1 | Positive | Mulki & von Korff ( |
| HvCO2 | Positive | Mulki & von Korff ( | ||
|
| Ryegrass | LpCO | Positive | Martin |
|
| Rice | OsCO3 | Negative | Kim |
| OsCOL4 | Negative | Lee | ||
| OsCOL9 | Negative | H. Liu | ||
| OsCOL10 | Negative | Tan | ||
| OsCOL13 | Negative | Sheng | ||
| OsCOL15 | Negative | Wu | ||
| OsCOL16 | Negative | Wu | ||
| OsDHD4 | Negative | Cai | ||
| OsGhd2 | Negative | Fan | ||
| OsHd1 | Positive | Z. Zhang | ||
|
| common bean | PvCOL2 | Negative | González |
|
| Peach | PpCO | Positive | Zhang |
|
| Rose | RcCO | Positive | Lu |
| RcCOL4 | Positive | Lu | ||
|
| Tomato | SlBBX1(SlCOL2) | Negative | J. Song |
| SlBBX3(SlCOL1) | Negative | Cui | ||
| J. Song | ||||
| SlBBX26 | Negative | Moreira | ||
|
| Potato | StBBX24 | Negative | Kiełbowicz‐Matuk |
|
| Sorghum | SbCO | Positive | S. Yang |
|
| Maize | ZmCOL3 | Negative | Jin |
|
| ||||
|
| Kiwi | AdBBX32 | Negative (fruit ripening) | Y. Yang |
|
| Strawberry | FvCO | Positive (inflorescence number) | Kurokura |
|
| Rice | OsBBX11 | Positive (harvestable organs development) | Shalmani |
| OsBBX19 | Positive (harvestable organs development) | Shalmani | ||
| OsCOL9 | Positive (harvestable organs number) | H. Liu | ||
| OsCOL13 | Positive (harvestable organs number) | Sheng | ||
| OsCOL16 | Positive (harvestable organs number) | Wu | ||
| OsDHD4 | Positive (harvestable organs number) | Cai | ||
|
| Tomato | SlBBX1(SlCOL2) | Positive (harvestable organs number) | J. Song |
| SlBBX3(SlCOL1) | Negative (harvestable organs number and size) | Cui | ||
| Positive (inflorescence branching) | J. Song | |||
| SlBBX5(SlCOL4) | Negative (fruit ripening) | Wu | ||
| SlBBX24 | Negative (harvestable organs number and size) | Cui | ||
| SlBBX25 | Negative (harvestable organs size) | Luo et al. ( | ||
| SlBBX26 | Positive (fruit ripening) | Moreira | ||
| Negative (harvestable organs size) | Moreira | |||
| SlBBX28 | Positive (harvestable organs number) | Lira | ||
|
| Potato | StCO | Negative (harvestable organs number) | González‐Schain |
|
| ||||
|
| ||||
| Cold tolerance | ||||
|
| Apple | MdBBX37 | Positive | An |
|
| Tabacco | NtBBX9 | Negative | S. Liu |
| NtBBX11 | Positive | S. Liu | ||
|
| Tomato | SlBBX17 | Positive | Song |
| SlBBX25 | Positive | Ma | ||
| SlBBX31 | Positive | Zhu | ||
|
| Potato | StBBX14 | Positive | Zhang |
| Drought tolerance | ||||
|
| Peanut | AhBBX6 | Positive | Tang |
|
| Chinese mustard | BjuBBX6 | Negative | Feng |
|
| Canola | BnBBX22.A07 | Positive | Zhang |
|
| Chrysanthemum | CmBBX19 | Negative | Xu |
|
| Soybean | GmCOL1a | Positive | C. Xu |
|
| Apple | MdBBX7(MdCOL9) | Positive | Chen |
| MdBBX22 | Positive | An | ||
|
| Rice | OsGhd2 | Negative | J. Liu |
| OsBBX11 | Positive | Lei | ||
|
| Tomato | SlBBX18 | Positive | Li |
| SlBBX25 | Positive | Ma | ||
|
| Potato | StBBX24 | Positive | Kiełbowicz‐Matuk |
| Heat tolerance | ||||
|
| Tomato | SlBBX17 | Positive | Xu |
| Shade avoidance response | ||||
|
| Chinese mustard | BjuCOL13 | Positive | Muntha |
|
| Tomato | SlBBX20 | Negative | Shiose |
|
| Maize | ZmDBB2 | Positive | X. Wang |
|
| ||||
|
| Bamboo | BDBBX21 | Positive (fungi resistance) | Y. Liu |
|
| Pepper | CaBBX14 | Positive (fungi resistance) | Zhou |
|
| Chrysanthemum | CmBBX32 | Positive (fungi resistance) | B. Wang |
|
| Cucumber | CsCOL9 | Positive (white fly resistance) | Xie |
|
| Sweet potato | IbBBX24 | Positive (fungi resistance) | Zhang |
|
| Lily | LpBBX28 | Positive (trichome initiation) | Xin |
|
| Tomato | SlBBX20 | Positive (fungi resistance) | Shiose |
| SlBBX25 | Negative (fungi resistance) | Luo | ||
|
| Potato | StBBX27 | Positive (oomycete resistance) | Sun |
|
| ||||
|
| Sweet wormwood | AaBBX21 | Positive (artemisinin in vegetative organs) | He |
|
| Cannabis | CsaCOL1 | Positive (cannabinoids in hairy roots) | M. Gao |
| CsaCOL5 | Positive (cannabinoids in hairy roots) | M. Gao | ||
| CsaCOL7 | Positive (cannabinoids in hairy roots) | M. Gao | ||
|
| Pepper | CaBBX10 | Positive (chlorophyll and carotenoids in fruits) | J. Wang |
| CaBBX20 | Positive (carotenoids in fruits) | Ma | ||
|
| Chrysanthemum | CmBBX28 | Negative (anthocyanins in flowers ) | L.J. Zhou |
| CmBBX20 | Positive (flavonoids and chlorogenic acid in flowers) | Lu | ||
|
| Orange, pomelo | ChBBX22 | Positive (carotenoids and anthocyanins in fruits) | Fu |
|
| Blood orange | CsBBX24 | Positive (carotenoids and anthocyanins in fruits) | Fu |
|
| Strawberry | FaBBX22 | Positive (anthocyanins in fruits) | Liu |
| FaBBX24 | Positive (anthocyanins in fruits) | Zhang | ||
|
| Soybean | GmBBX4 | Negative (anthocyanins in seeds ) | Song |
| GmBBX22 | Positive (flavonoids/anthocyanins in vegetative organs) | Zhan | ||
|
| Cotton | GhBBX21 | Positive (anthocyanins in vegetative organs) | Li |
| GhBBX24(GhDR) | Negative (anthocyanins in vegetative organs) | X. Wang | ||
| Li | ||||
|
| Sweeto potato | IbBBX29 | Positive (flavonoids in leaves and roots) | Gao |
|
| Lily | LvBBX24 | Positive (anthocyanins in flowers ) | Gao |
|
| Apple | MdCOL4 | Negative (anthocyanins in fruits ) | Fang |
| MdCOL6 | Positive (anthocyanins in fruits ) | Wang | ||
| MdBBX20 | Positive (anthocyanins in fruits ) | Fang | ||
| MdBBX21 | Positive (anthocyanins in fruits ) | Zhang | ||
| MdBBX22 | Positive (anthocyanins in fruits ) | An | ||
| Zhang | ||||
| MdBBX37 | Negative (anthocyanins in leaves) | An | ||
|
| Mango | MiBBX24 | Positive (anthocyanins and carotenoids in fruits) | Pan |
| MiBBX27 | Positive (anthocyanins and carotenoids in fruits) | Pan | ||
|
| Petunia | PhCOL16a | Positive (chlorophyll in flowers) | Ohmiya |
|
| Sweet cherry | PavBBX6 | Positive (anthocyanins in fruits ) | Wang |
| PavBBX9 | Positive (anthocyanins in fruits ) | Wang | ||
|
| Peach | PpBBX32 | Positive (anthocyanins in fruits ) | Huang |
|
| Pear | PyBBX16 | Positive (anthocyanins in fruits ) | Bai |
| PyBBX18 | Positive (anthocyanins in fruits ) | Bai | ||
| PyBBX21 | Negative (anthocyanins in fruits ) | Bai | ||
| PyBBX24 | Negative (anthocyanins in fruits and vegetative organs) | Li | ||
| Yang | ||||
|
| Palmleaf raspberry | RcBBX26 | Positive (anthocyanins in leaves) | Xu |
|
| Tomato | SlBBX3(SlCOL1) | Positive (anthocyanins in vegetative organs) | Liu |
| Positive (chlorophylls in fruits) | Cui | |||
| SlBBX20 | Positive (flavonoids in fruits) | Shiose | ||
| SlBBX24 | Positive (anthocyanins in fruits) | He | ||
| Positive (chlorophylls in fruits) | Cui | |||
| SlBBX25 | Positive (carotenoids and flavonoids in fruits) | Xiong | ||
| Positive (anthocyanins in vegetative organs) | Luo | |||
|
| Eggplant | SmBBX22 | Positive (anthocyanins in fruits) | J. Li |
|
| Wheat | TaBBX3B | Positive (anthocyanins in seeds) | Jiang |
|
| Grape | VvBBX8 | Positive (anthocyanins in fruits) | Qiu |
| VvBBX11 | Positive (anthocyanins in fruits) | Qiu | ||
| VvBBX32 | Positive (anthocyanins in fruits) | Qiu | ||
| VvBBX44 | Negative (anthocyanins in fruits) | W. Liu | ||
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico10.13039/501100003593
- —Fundação de Amparo à Pesquisa do Estado de São Paulo10.13039/501100001807
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLight effects on plants · Plant Gene Expression Analysis · Plant nutrient uptake and metabolism
Contents Summary 762I Introduction 762II The BBX protein family: insights from Arabidopsis thaliana 763III The BBX gene family in crop species 765IV Reproductive development 765V Stress response 771VI Specialized metabolism 774VII Conclusions and perspectives 778 Acknowledgements 781 References 781
Introduction
I.
As sessile organisms, plants rely on dynamic and highly interconnected signaling networks to adapt to continuously changing environmental conditions. Central to these mechanisms, transcription factor (TF)–controlled gene regulatory networks rapidly reprogram gene expression, thereby enabling metabolic and developmental adjustments throughout the plant life cycle. Among the TF families, the B‐box (BBX) zinc finger proteins contain at least one BBX domain, which encompasses c. 40 amino acids and is stabilized by cysteine, histidine and aspartic acid residues along with two zinc ions. Moreover, BBX proteins may contain a CONSTANS, CONSTANS‐like and TIMING OF CAB1 (CCT) domain characterized by its highly conserved 42–43 amino acids. The TF activity of the BBX family members relies on both the BBX and CCT domains. Lastly, a nuclear localization signal occurs in the C‐terminus region of the protein (Song et al., 2024a).
BBX proteins regulate the expression of their target genes by recognizing specific cis‐regulatory motifs within promoter regions, including the CCAAT box (Ben‐Naim et al., 2006), CORE2 (TGTGN_2‐3_ATG, Tiwari et al., 2010), CCACA (Gnesutta et al., 2017), the G‐box (CACGTG; Song et al., 2020) and a modified G‐box motif (TACGTG; Xiong et al., 2019). Additionally, BBX proteins frequently form heterodimers with other TFs, thereby either enhancing or repressing their regulatory activity (Cao et al., 2023). Meanwhile, BBX proteins are often degraded by E3 ubiquitin ligase complex‐mediated post‐translational regulation (Lin et al., 2018). With these modes of action, BBX TFs regulate developmental processes ranging from germination and seedling establishment to flowering and senescence (Song et al., 2024a).
Therefore, with a focus on crop species, this review explores how BBX TFs enable plant adaptation bridging environmental stimuli and developmental processes through multifaceted regulatory mechanisms.
The BBX protein family: Insights from Arabidopsis thaliana
II.
In A. thaliana, the 32 AtBBX proteins cluster into five structural groups based on the number and evolutionary origin of BBX domains and on the presence or absence of the CCT domain. Group I and II proteins display two BBX domains with independent origins, and one CCT domain; Group III structure consists of a unique BBX and a CCT domain; lastly, Groups IV and V lack CCT and contain two and one BBX domains, respectively (Khanna et al., 2009). AtBBX26 and AtBBX27, initially structural Group V members, were recently reassigned to Group II following a robust phylogenetic analysis and the identification of a second BBX domain and a vestigial CCT domain (Lira et al., 2020; Fig. 1).
B‐box (BBX) proteins cluster into five structural groups. Phylogenetic analysis of the 32 Arabidopsis thaliana and the 31 Solanum lycopersicum BBX proteins. Sequences were aligned with the default parameters of the Expresso T‐Coffee algorithm (Armougom et al., 2006). The tree was reconstructed from the obtained alignment using the phyml 3.0 package (Guindon et al., 2010) with the JTT (Jones‐Taylor‐Thornton) substitution model, and the proportion of invariable sites and gamma shape parameter were estimated from the data sample. Then, the tree was optimized by tree topology and branch length, and improved by subtree pruning and regrafting. Finally, the branch support was calculated by the approximate likelihood‐ratio test Shimodaira–Hasegawa‐like (aLTR SH‐like). The protein topology for each structural group was predicted from the group consensus sequence using the InterPro database (Blum et al., 2025) and is shown on the right side. The functions are indicated by colored circles and are listed and referred to in Supporting Information Table S1 for A. thaliana and Table 1 for S. lycopersicum. The loci were numbered according to Lira et al. (2020).
Out of the 32 AtBBX genes, only AtBBX12, AtBBX26 and AtBBX27 remain functionally uncharacterized (Supporting Information Table S1). Interestingly, 66 and 79% of the 29 characterized AtBBXs are associated with flowering time and light responses (i.e. seedling photomorphogenesis, shade avoidance and light stress), respectively. Additionally, most AtBBXs have been associated with other functions, highlighting the pleiotropic effects of BBX family members on plant physiology. Given the current state of knowledge, no structural groups can yet be reliably correlated with specific physiological processes (Fig. 1).
The first AtBBX characterized was AtBBX1 (CONSTANS, AtCO), which directly binds to the florigen FLOWERING LOCUS T (AtFT) promoter, inducing vegetative‐to‐reproductive meristem transition (Putterill et al., 1995; Tiwari et al., 2010). Similarly, AtBBX6/24 also induce AtFT expression (Hassidim et al., 2009; Li et al., 2014), while AtBBX4/5/32 inhibit the florigen transcription (Cheng & Wang, 2005; Tripathi et al., 2017; Steinbach, 2019). The other flowering‐associated AtBBXs regulate AtBBX1, either repressing its expression (i.e. AtBBX7; Cheng & Wang, 2005) or modulating its transcriptional regulatory activity over AtFT through physical interaction. The latter is the case of the negative AtBBX10/13/14/15/16/17/19/30/31 (Wang et al., 2014; Graeff et al., 2016; Ordoñez‐Herrera et al., 2018; Xu et al., 2022b; Susila et al., 2023; Rahul et al., 2024) and the positive AtBBX28/29 factors (Wang et al., 2021).
Photomorphogenesis is the modulation of plant development in response to variations in light conditions. One extensively studied aspect is seedling photomorphogenesis, the transition from a heterotrophic to a photoautotrophic organism. If light is limiting, the seedling will exhibit elongated hypocotyls with underdeveloped cotyledons; in contrast, upon light exposure, the seedlings will show thick and short hypocotyls with open cotyledons (Nemhauser & Chory, 2002). Twenty AtBBXs have been characterized as seedling photomorphogenesis regulators, participating in an intricate transcriptional and post‐translational regulatory network together with ELONGATED HYPOCOTYL 5 (AtHY5) and/or PHYTOCHROME INTERACTING FACTORS (AtPIFs), which are the light‐signaling master positive and negative TFs, respectively. Among the positive regulators of seedling photomorphogenesis, AtBBX11/14/15/16 induce AtHY5 expression (Zhao et al., 2020; Nasim et al., 2024), AtBBX22/23 regulate AtHY5 protein activity through dimerization (X. Zhang et al., 2017; Podolec et al., 2022), while AtBBX20/21 act at both transcriptional and post‐translational levels (Wei et al., 2016; Xu et al., 2016; Podolec et al., 2022). By contrast, AtBBX4 interacts with AtPIF3, alleviating the AtPIF3‐mediated repression of photomorphogenic genes (Heng et al., 2019). Moreover, AtBBX9/14/15/16/20/21 induce photomorphogenesis by downregulating auxins, brassinosteroids (BRs), and gibberellins (GAs) metabolism and signaling (Fan et al., 2012; Xu et al., 2017; Nasim et al., 2024; Song et al., 2024b). On the contrary, the negative regulators AtBBX24/25/28/29 physically interact with AtHY5, hindering its ability to bind to its target genes' promoters, thereby inhibiting photomorphogenesis (Gangappa et al., 2013; Song et al., 2020). Interestingly, AtHY5 represses the expression of AtBBX30/31, whose protein products induce AtBBX28/29, resulting in a feedback loop that fine‐tunes seedling development (Song et al., 2020). The negative effect of AtBBXs on photomorphogenesis is also mediated by hormones. For instance, AtBBX18, which induces hypocotyl elongation by upregulating GA biosynthesis (Wang et al., 2011), and AtBBX28/29/32, which regulate BRs signaling (Cao et al., 2022; Ravindran et al., 2021). Finally, AtBBX19 promotes hypocotyl growth by facilitating AtCOP1‐mediated degradation of EARLY FLOWERING 3 (AtELF3; Wang et al., 2014).
AtBBXs also regulate cotyledon development during seedling photomorphogenesis. The GENOMES UNCOUPLED 1 (AtGUN1)/GOLDEN2 LIKE 1 (AtGLK1) retrograde signaling module regulates the expression of AtBBX14/16, which are positive regulators of cotyledon development (Veciana et al., 2022; Atanasov et al., 2024). Moreover, as Chl biosynthetic genes are upregulated by AtBBX11, its expression is tightly regulated by AtPIF3 and AtHY5 to avoid photobleaching (Job & Datta, 2021). AtPIF1‐AtBBX10 dimer inhibits photo‐oxidative damage in de‐etiolating seedlings by repressing Chl biosynthesis in an AtGUN5‐mediated manner in darkness and by inducing antioxidant enzymes under light conditions (L. Zhou et al., 2025). Similarly, AtBBX32 acts with AtPIF3, maintaining the repression on AtGUN4/5 and photosynthesis‐associated genes during soil emergence, fine‐tuning cotyledon greening (K. Wang et al., 2025).
Shade avoidance syndrome (SAS) is a photomorphogenic response triggered by the reduction in the red : far‐red (R : FR) ratio of incident light. SAS is characterized by hypocotyl, internodes and petiole elongation, hyponasty, reduced branching and early flowering (Ballaré & Pierik, 2017). The overexpression of AtBBX16 results in longer hypocotyls and reduced branching under shade conditions (Wang et al., 2013). AtBBX24 also positively regulates SAS, which, through the interaction with the GA repressors AtDELLAs, releases AtPIF4 to induce the expression of auxin signaling genes (Crocco et al., 2015). AtBBX28 triggers SAS by inducing auxin‐related genes in an AtCOP1‐dependent manner (Saura‐Sánchez et al., 2024). Finally, AtBBX7/8 positively regulate the low R : FR‐induced AtPIF4 expression and promote hypocotyl growth (Bian et al., 2025). Conversely, AtBBX21 reduces BRs biosynthesis, repressing hypocotyl elongation under shade conditions (Gómez‐Ocampo et al., 2024).
The role of several AtBBX proteins in plant stress responses has been increasingly attracting attention. AtBBX1 antagonizes ABSCISIC ACID‐RESPONSIVE ELEMENT BINDING FACTORS (AtABFs) through dimerization, downregulating salinity‐responsive genes and, consequently, stress tolerance (Du et al., 2023). AtBBX21/24 negatively regulate ABSCISIC ACID INSENSITIVE 5 (AtABI5) expression, reducing abscisic acid (ABA) sensitivity during germination (Xu et al., 2014; Chiriotto et al., 2023). Similarly, Atbbx5 exhibits increased sensitivity to ABA and salt stress during seed germination (Min et al., 2015).
AtBBX11/31 induce photoprotection and DNA damage repair genes under UV‐B light stress (Yadav et al., 2019; Job et al., 2022). Similarly, AtBBX20/21/22 are needed for hypocotyl growth inhibition and photoprotective pigment accumulation in response to UV‐B (Podolec et al., 2022). AtBBX14 is required for acclimation to light‐stress conditions, as Atbbx14 plants display compromised photosynthetic activity when exposed to high light (Atanasov et al., 2024). By contrast, AtBBX32 impairs light‐stress acclimation by downregulating photosynthesis‐associated gene expression under high‐light conditions (Alvarez‐Fernandez et al., 2021).
Finally, the AtPIF4‐mediated hypocotyl elongation triggered by high temperature is induced by the partially redundant AtBBX18/23/24/25 (Ding et al., 2018; Malakar et al., 2025b) and repressed by AtBBX21 (Malakar et al., 2025a). A recent report advanced the limits of current understanding by highlighting the importance of both sense and antisense AtBBX28 transcript expression in enhancing plant freezing tolerance (Meena et al., 2024). Finally, under cold stress, blue light‐induced phosphorylation stabilizes CRYPTOCHROME 2 (AtCRY2), which interacts with AtCOP1, preventing AtHY5 degradation. As direct targets of AtHY5, AtBBX7/8 positively regulate freezing tolerance by modulating cold‐responsive genes (Li et al., 2021).
The BBX gene family in crop species
III.
A gene family is a group of genes that arose from duplication of a single ancestral gene and have retained sequence similarities. In plants, gene families expand through whole‐genome (WGD), single‐gene and segmental duplication events, which enhance genetic diversity and provide raw material for emergence of new functions (Fang et al., 2022). Within plant genomes, tandem gene duplication is a major single‐gene duplication mechanism, characterized by the occurrence of two or more adjacent homologous genes. Yet, segmental duplications arise from the duplication of larger genomic fragments containing variable numbers of genes that are identified as syntenic blocks. By surveying 18 representative plant species, Yu et al. (2022) proposed that the expansion of the BBX gene family is primarily driven by WGD, with segmental duplication acting as a secondary contributing mechanism.
Whole‐genome surveys over the last decade have identified BBX gene families in 79 plant species, most of which contain between 16 and 40 members. In all cases, the loci cluster into the five structural groups originally described (Table S2). In some species, the copy number is notably higher. Such is the case of cotton (Gossypium hirsutum), which has 127 BBX proteins, mainly originated from lineage‐specific WGD events (Paterson et al., 2012; Sun et al., 2023). Moreover, the number of BBX‐coding genes in the genomes also reflects the ploidy of the species. In strawberry, while the diploid species (Fragaria vesca) contains 22 BBX genes, 51 are found in the polyploid genotype (F. ananassa; D. Xu et al., 2023; Ye et al., 2021). In this line, the tetraploid genome of Vaccinium corymbosum blueberry harbors 83 BBX genes, while the diploid V. darrowii has only 24 loci (Xue et al., 2025). The number of BBX genes in the genomes of the polyploid species Brassica napus, B. carinata, B. juncea and wheat (Triticum aestivum) is approximately three times that of the number of loci in A. thaliana (Chen et al., 2021; Singh et al., 2021; Zheng et al., 2021).
Most genome‐wide BBX identification studies also characterize their expression patterns showing that these genes are differentially expressed across organs and in response to hormonal and stress treatments. Based on this information, subsequent studies have investigated the mechanisms through which BBX proteins modulate agronomically important traits in crop species, including reproductive development, abiotic and biotic stress responses, and specialized metabolism. Below, we review the current knowledge on the involvement of BBX proteins in these processes.
Reproductive development
IV.
1. Flowering time
The first function identified for BBX proteins was the regulation of flowering time. As a key determinant of crop yield in many species, this discovery prompted investigations into whether crop BBXs regulate flowering through analogous mechanisms (Fig. 2; Table 1). In this sense, by recovering Atco late flowering phenotype, BBXs were identified as positive regulators of flowering, including canola (Brassica napus) BnCOa1 (Robert et al., 1998), cotton (G. hirsutum) GhCOL1A and GhCOL1D (Cai et al., 2017), peach (Prunus persica) PpCO (Zhang et al., 2015), ryegrass (Lolium perenne) LpCO (Martin et al., 2004), and soybean (Glycine max) GmCOL5 (Fan et al., 2014).
B‐box (BBX) genes that regulate reproductive development in crop species. The species are indicated by the genus and species initials: Actinidia deliciosa (Ad), Brassica juncea (Bju), Chrysanthemum indicum (Ci), Chrysanthemum morifolium (Cm), Fragaria vesca (Fv), Glycine max (Gm), Gossypium hirsutum (Gh), Hordeum vulgaris (Hv), Oryza sativa (Os), Phaseolus vulgaris (Pv), Rosa chinensis (Rc), Solanum lycopersicum (Sl), Solanum tuberosum (St), Sorghum bicolor (Sb) and Zea mays (Zm). Black lines and white boxes represent DNA and genes, respectively. BBX proteins are represented in purple, other proteins in blue and hormones in yellow. Blue lines indicate transcriptional regulation, red lines indicate post‐translational regulation, and green lines indicate the effect of the BBX protein on the phenotype. Arrows represent positive regulation, and blunt‐ended arrows represent inhibition. Solid and dotted lines indicate direct or indirect effects, respectively. Both gene and hormone names are written in red or blue, depending on whether they have a negative or a positive effect on the phenotype, respectively. Gene, protein and hormone abbreviations are detailed in the text. VM, TM and FM: vegetative, transition and floral meristem, respectively.
In potato (Solanum tuberosum), StBBX24 was identified as a repressor of floral transition, as knockdown plants exhibited early flowering and up‐ and downregulation of key flowering positive and negative regulators, respectively (Kiełbowicz‐Matuk et al., 2022). Similarly, silencing GhCOL3 in cotton plants resulted in an early‐flowering phenotype likely acting upstream of florigen repressors (Z. Song et al., 2025).
For other BBXs, the molecular mechanisms underlying the regulation of flowering time have been more extensively explored. In the Brassica juncea (Chinese mustard), BjuBBX6 interacts with the NUCLEAR FACTOR YB3 (BjuNF‐YB3) and, alone or as a dimer, downregulates the expression of the flowering suppressor FLOWERING LOCUS C (BjuFLC; Feng et al., 2025). The strawberry FvCO, in a long‐day‐flowering accession, controls the expression of FvFT1, regulating the differentiation of inflorescence‐producing branch crowns, rather than into vegetative runners (Kurokura et al., 2017). In Sorghum bicolor, SbCO not only induces the florigens CENTRORADIALIS 8 (SbCN8) and SbCN12, but also activates the expression of the florigens activator EARLY HEADING DATE 1 (SbEHD1; S. Yang et al., 2014).
The three tomato (Solanum lycopersicum) BBXs characterized as flowering regulators act as repressors. SlCOL1 and SlCOL2 inhibit the expression the florigen SINGLE FLOWER TRUSS (SlSFT), either directly (Cui et al., 2022) or inducing SELF PRUNNING 5G (SlSP5G) flowering repressor (J. Song et al., 2025). SlBBX26, on the contrary, indirectly regulates flowering by promoting the accumulation of GAs, which disrupt the SlSFT inducer SlPIF4‐SlDELLA dimer (Moreira et al., 2025).
In Chrysanthemum morifolium, the long‐day‐associated florigen FLOWERING LOCUS T LIKE 1 (CmFTL1) is directly activated by the synergistic action of CmBBX7 and CmBBX8 (Wang et al., 2020; Zhai et al., 2023). Conversely, CmBBX5 dimerizes with CmBBX8, reducing the transcriptional promoting activity of the latter (Wang et al., 2024). The short‐day florigen CmFTL3 is indirectly repressed by CmBBX24, as this protein inhibits GA synthesis, which is a florigen‐inductor hormone in this species (Y. Yang et al., 2014). In another chrysanthemum species, C. indicum, CiBBX5 directly represses CiFTL3 expression, which is further strengthened through heterodimerization with CiBBX19 (C. Yang et al., 2025).
In the common bean (Phaseolus vulgaris), PvCOL2 represses PvFT expression under noninductive long‐day conditions. Mutations in PvCOL2 reduce photoperiod sensitivity, and when combined with loss‐of‐function alleles of PHYTOCHROME A3 (PvPHYA3), result in photoperiod‐insensitive flowering in domesticated cultivars (González et al., 2021). Similarly, continuous flowering in rose (Rosa chinensis) relies on the interplay between RcCO and RcCOL4. Under short days, RcCO is upregulated and activates RcFT. Under long days, RcCOL4 is induced and, by dimerizing with RcCO, increases its affinity for the RcFT promoter (Lu et al., 2020). In soybean, under long day, the GmBBX32a‐GmBBX32b dimer inhibits the transcription of the flowering negative regulator GmE1 (C. Gao et al., 2025).
In maize (Zea mays), ZmCOL3 directly promotes the expression of flowering repressors, such as ZmCCT. Interestingly, polymorphisms in the ZmCOL3 promoter and in the 5′ untranslated region may have contributed to the adaptation from tropical to temperate regions (Jin et al., 2018). Similarly, in the Japanese morning glory (Ipomoea nil), it has been suggested that a single base deletion in InCO was responsible for an early‐flowering QTL, which might have facilitated adaptation to temperate Asian regions (Katsuyama et al., 2025).
To date, rice (Oryza sativa) is the crop species with the largest number of BBX proteins identified as flowering regulators. The vegetative‐to‐reproductive growth transition is triggered by EARLY HEADING DATE 1 (OsEHD1), which activates the expression of the florigens HEADING DATE 3A (OsHD3a) and RICE FLOWERING LOCUS T 1 (OsRFT1). Conversely, OsEHD1 transcript accumulation is downregulated by GRAIN NUMBER, PLANT HEIGHT AND HEADING DATE 7 (OsGHD7; J. Song et al., 2024). The only known rice BBX that positively regulates flowering is OsHD1, which upregulates OsEHD1, OsHD3a and OsRFT1. The transcriptional activity of this BBX is, however, suppressed through dimerization with OsGHD7 (Z. Zhang et al., 2017). Nine OsBBXs are flowering repressors, although it remains unclear whether some act directly or indirectly. Notably, OsGHD7 is upregulated by OsCOL15 (Wu et al., 2018) and OsCOL16 (Wu et al., 2017). Additionally, OsEHD1 is downregulated by OsCOL4 (Lee et al., 2010), OsCOL9 (H. Liu et al., 2016), OsCOL10 (Tan et al., 2016) and OsCOL13 (Sheng et al., 2016). Additionally, the BBX OsGHD2 directly upregulates OsCO3 (Fan et al., 2023), which in turn represses the florigen‐encoding genes OsHD3a and OsFTL1 (Kim et al., 2008). Moreover, the BBX protein DELAYED HEADING DATE 4 (OsDHD4) interacts with the TF FLOWERING LOCUS D1 (OsFD1), disrupting the florigen activation complex composed of OsHD3a, Os14‐3‐3 and OsFD1 (Cai et al., 2021). Furthermore, OsCOL15 inhibits the expression of the flowering activator RICE INDETERMINATE 1 (OsRID1; Wu et al., 2018).
Finally, an interesting regulatory mechanism has been identified in barley (Hordeum vulgaris), where HvCO1 and HvCO2 may either repress or promote the flowering transition (Mulki & von Korff, 2016). Before vernalization, both BBXs induce VERNALIZATION2 (HvVRN‐H2) expression, which downregulates the florigen HvFT1. During vernalization, the accumulation of HvVRN‐H1 represses HvVRN‐H2, altering the activity of HvCO1 and HvCO2 that upregulate HvFT1 expression, promoting flowering.
2. Beyond flowering time
While the regulation of BBXs over flowering induction has been extensively studied, less is known about their roles in reproductive processes beyond flowering time (Fig. 2; Table 1). Some of the flowering time regulators mentioned above have been reported to affect other reproductive traits. For instance, FvCO positively controls the number of inflorescences per plant (Kurokura et al., 2017).
In rice, delaying flowering time through the overexpression of OsCOL13 (Sheng et al., 2016), OsCOL16 (Wu et al., 2017) and OsDHD4 (Cai et al., 2021) increased panicle size, grain number and yield. Conversely, these parameters were reduced in Oscol9 mutant, even though no differences were detected in overexpressing genotypes (H. Liu et al., 2016). For most of these genes, the increased yield was interpreted as a consequence of the prolonged vegetative phase, without further investigation into whether these BBXs also affect panicle development. Furthermore, Shalmani et al. (2023) identified OsBBX19 and OsBBX11 as positive regulators of spikelet size and seed production through the transcriptional and post‐translational positive regulation of BROAD‐COMPLEX, TRAM TRACK and BRIC‐A‐BRAC (OsBTB97), respectively.
To date, tomato is the crop species with the highest number of BBXs identified as regulators of the reproductive development beyond flowering transition. SlBBX28 regulates inflorescence branching and fruit number per plant by upregulating auxin biosynthesis and by repressing the expression of the inflorescence branching inhibitor FRUITFULL 2 (SlFUL2; Lira et al., 2022). Conflicting results have been reported regarding the effect of SlCOL1 in flowers and fruit production. While Cui et al. (2022) describe this gene as a negative regulator of fruit size and flowers and fruits number, J. Song et al. (2025) showed that SlCOL1 and SlCOL2 promote inflorescence branching. Interestingly, the overexpression of SlBBX24, a SlCOL1‐interacting protein, reduces fruit size (Cui et al., 2022). Furthermore, SlBBX25 (as originally Solyc01g110180 locus was named by Chu et al., 2016) also negatively regulates fruit size and number (Luo et al., 2023). SlBBX26 is another regulator of processes beyond flowering time. Although SlBBX26 deficiency reduced the abortion rate, increasing fruit production, fruits were smaller due to the downregulation of cell division‐ and expansion‐related genes. SlBBX26 also promotes fruit ripening by lowering auxin levels through two coordinated mechanisms. During preripening stages, the SlBBX26–SlPIF4 complex directly upregulates GA2 OXIDASE 2 (SlGA2OX2), promoting GA degradation that reduces auxin accumulation. In addition, SlBBX26 directly activates the master ripening TF RIPENING INHIBITOR (SlRIN), which diminishes auxin concentrations (Moreira et al., 2025). On the contrary, SlCOL4 inhibits fruit ripening by downregulating ethylene biosynthesis‐related genes (Wu et al., 2025).
In kiwi (Actinidia deliciosa), AdBBX32 interacts with the ripening‐promoting TF NAM, ATAF1/2 and CUC2 3 (AdNAC3), impairing its activity and, consequently, inhibiting ripening‐related genes (Y. Yang et al., 2025). Lastly, potato StCO reduces tuberization in repressive and weakly inductive photoperiods by downregulating the tuberization‐inducers BEL1‐LIKE PROTEIN 5 (StBEL5) and StFT expression (González‐Schain et al., 2012).
Stress response
V.
1. Abiotic stress response
Throughout the life cycle, plants are exposed to diverse environmental stressors, including drought, salinity, extreme temperatures and fluctuations in light conditions. These abiotic challenges can strongly impair development and yield, especially as climate change increases their frequency and severity (Zhang et al., 2025b). To cope with these conditions, plants have evolved complex adaptive strategies, in which BBX proteins have emerged as key regulators (Bandara et al., 2022; Fig. 3; Table 1).
B‐box (BBX) proteins involved in stress‐response regulation in crop species. The species are indicated by the genus and species initials. Arachis hypogaea (Ah), Bambusa pervariabilis × Dendrocalamopsis grandis (BD), Brassica juncea (Bju), Brassica napus (Bn), Capsicum annuum (Ca), Chrysanthemum morifolium (Cm), Cucumis sativus (Cs), Glycine max (Gm), Ipomoea batatas (Ib), Lilium pumilum (Lp), Malus domestica (Md), Nicotiana tabacum (Nt), Oryza sativa (Os), Solanum lycopersicum (Sl), Solanum tuberosum (St) and Zea mays (Zm). Black lines and white boxes represent DNA and genes, respectively. BBX proteins are represented in purple, HY5 in pink, other proteins in blue and hormones in yellow. Blue lines indicate transcriptional regulation, red lines indicate post‐translational regulation and green lines indicate the effect of the BBX protein on the phenotype. Arrows represent positive regulation and blunt‐ended arrows represent inhibition. Solid and dotted lines indicate direct or indirect effects, respectively. Both gene and hormone names are written in red or blue depending on whether they have a negative or a positive effect on the phenotype, respectively. Gene, protein and hormone abbreviations are detailed in the text.
In rice, the BBX OsGHD2 negatively regulates drought tolerance by promoting leaf senescence. It directly controls the expression of the Chl catabolism‐related genes STAY GREEN 1 (OsSGR1), NON‐YELLOW COLORING 1 (OsNYC3) and PHEIDE A OXYGENASE (OsPAO); the glyoxylate cycle enzyme encoding genes ISOCITRATE LYASE (OsICL) and MALATE SYNTHASE (OsMS); and PYRUVATE ORTHOPHOSPHATE DIKINASE B (OsPPDKB) and ASPARAGINE SYNTHASE 1 (OsAS1), whose encoded proteins are involved in amino acid remobilization (J. Liu et al., 2016). Conversely, deficiency in OsBBX11 resulted in salt hypersensitivity due to excessive accumulation of Na^+^ and K^+^ in shoots, revealing this protein as a positive regulator of ion homeostasis (Lei et al., 2023). In a similar way, the soybean GmCOL1a enhances salt and drought tolerance by modulating ion transport between roots and shoots. The underlying mechanisms involve the direct induction of the stress‐responsive genes LATE EMBRYOGENESIS ABUNDANT PROTEIN (GmLEA) and PYRROLINE‐5‐CARBOXYLATE SYNTHASE (GmP5CS), as well as the ABA‐mediated stomatal closure (C. Xu et al., 2023). Some other BBX proteins that modulate drought response also act by regulating ABA signaling. In apple (Malus domestica), MdBBX22 enhances drought tolerance by delaying ABA‐induced leaf senescence through the reduction in MdHY5‐mediated activation of ABA INSENSITIVE 5 (MdABI5) transcription and the physical interaction with MdABI5, interfering with its ability to activate the Chl catabolic genes MdSGR1 and MdNYC1 (An et al., 2021b). Furthermore, under water‐limiting conditions, MdBBX7 directly activates drought‐responsive genes, including ETHYLENE RESPONSE FACTOR 1 (MdERF1), EARLY RESPONSIVE TO DEHYDRATION 15 (MdERD15) and MdGLK1; however, MdBBX7 is degraded by MYB30‐INTERACTING E3 LIGASE 1 (MdMIEL1) under water availability. Overexpression of MdBBX7 increases the activity of the antioxidant enzymes peroxidase and catalase, elevates ABA levels and reduces H_2_O_2_ accumulation, conferring higher stress resilience (Chen et al., 2022).
In tomato, SlBBX25 enhances salt tolerance by increasing antioxidant enzyme activities, improving water content and photosynthesis, thereby reducing malondialdehyde (MDA) and reactive oxygen species (ROS) levels. It directly induces stress‐protective genes, such as PYRROLINE‐5‐CARBOXYLATE REDUCTASE (SlP5CR) and SMALL HEAT SHOCK PROTEINS (SlHSP21.6A and SlHSP26.2; Ma et al., 2024). Allelic variation in BBX18 among tomato species underscores its role in drought adaptation. The SlBBX18 allele, from the drought‐sensitive cultivated tomato, encodes a truncated protein compared with drought‐tolerant wild species, Solanum pennellii. Although the full‐length SpBBX18 transcriptionally represses ASCORBATE PEROXIDASE 1 (SlAPX1), the reduction in SlAPX1 activity caused by its dimerization with the truncated SlBBX18 is even stronger than the transcriptional repression, ultimately weakening the drought tolerance mechanisms of cultivated tomato (Li et al., 2024).
The conserved role of BBX proteins in drought response is also evident in other crops. The silencing of peanut (Arachis hypogaea) AhBBX6 leads to reduced antioxidant enzyme activity and elevated MDA levels (Tang et al., 2024). Similarly, loss of StBBX24 function in potato diminished salinity tolerance, as reflected by reduced Na^+^/H^+^ antiporter transcript levels and lowered antioxidant enzyme activity following salt treatment (Kiełbowicz‐Matuk et al., 2022). In canola, BnBBX22.A07 enhances salinity tolerance by heterodimerizing with BnHY5.C09 to boost its activation of the BnWRKY33.C03 promoter (Zhang et al., 2024). Conversely, in Chinese mustard, BjuBBX6 negatively regulates drought tolerance by lowering Chl levels and antioxidant activity through direct repression of BjuGLK and activation of BjuABI2 (Feng et al., 2025). Finally, under normal conditions, CmBBX19 from C. morifolium dimerizes with CmABF3 suppressing its transcriptional activation of ABA‐responsive genes, such as RESPONSIVE TO ABA 18 (CmRAB18). Under drought stress, ABA accumulation downregulates CmBBX19, relieving CmABF3 repression and inducing ABA‐responsive genes (Xu et al., 2020).
Several BBX genes have been characterized as regulators of extreme temperature tolerance, acting via distinct mechanisms. For example, the expression of C‐REPEAT BINDING FACTORs (SlCBFs), whose proteins bind cold‐ and dehydration‐responsive elements in target promoters to activate cold‐responsive genes, is upregulated by SlBBX17 and SlBBX31 (Song et al., 2023; Zhu et al., 2023). Interestingly, SlHY5 is stabilized by SlBBX17 and, together, they induce SlCBFs, while SlHY5 also upregulates SlBBX31. This induction is strengthened in domesticated tomato genotypes as consequence of a 27 bp deletion in the SlBBX31 promoter. Under low‐temperature conditions, the upregulation of SlBBX25 induces the expression of genes associated with cold response, antioxidant activity, anthocyanin biosynthesis and starch catabolism. These molecular changes mitigate ROS and MDA accumulation, enhancing photosynthetic efficiency under stress (Ma et al., 2025). The overexpression of the potato StBBX14 reduces leaf damage and electrolyte leakage under low temperature, although the underlying molecular mechanism remains uncharacterized (Zhang et al., 2025a).
The contrasting effects of tobacco (Nicotiana tabacum) NtBBX9 and NtBBX11 in response to low temperatures highlight the complex regulatory networks in which BBX factors participate to modulate stressful conditions. Both genes are upregulated by cold treatment; however, while NtBBX11 induces the expression of the cold‐responsive genes NtCBFs, NtLEA14 and LOW‐TEMPERATURE‐INDUCED 65 (NtLTI65), NtBBX9 represses them (S. Liu et al., 2023). Finally, under normal conditions, the activity of apple MdBBX37 is suppressed by MdMIEL1‐mediated ubiquitination and by the interaction with the jasmonic acid (JA) response repressors JASMONATE‐ZIM‐DOMAIN PROTEIN 1 (MdJAZ1) and MdJAZ2. Cold‐induced JA accumulation triggers the degradation of MdMIEL1 and JAZ proteins, releasing MdBBX37 to activate MdCBF1 and MdCBF4 expression, promoting cold tolerance (An et al., 2021a).
Although heat stress is increasingly relevant to crop improvement, studies on BBX regulation of high‐temperature responses have lagged behind. In tomato, the overexpression of SlBBX17 enhances heat tolerance by stabilizing membranes, reducing ROS and upregulating the expression of HEAT SHOCK FACTORS (SlHSFs), SlHSPs and ROS detoxification genes (Xu et al., 2022a).
Beyond drought and extreme temperatures, BBX proteins are also involved in responses to light‐stress conditions, particularly SAS, often in a PIFs and/or HY5‐mediated manner. In B. juncea, BjuCOL13 is a negative regulator of shade avoidance that interacts with PHYTOCHROME A SIGNAL TRANSDUCTION 1 (BjuPAT1) factor under low R : FR light to suppress shoot branching (Muntha et al., 2019). In maize, DOUBLE B‐BOX ZINC FINGER PROTEIN 2 (ZmDBB2) acts as a central regulator of SAS. Under reduced R : FR light conditions, ZmPIF4 upregulates ZmDBB2 expression, and ZmDBB2‐ZmPIF4 dimer synergistically promotes the expression of cell elongation‐related genes, such as EXPANSIN 1 (ZmEXPA1). Under high R : FR light conditions, ZmDBB2 interacts with ZmHY5 to repress its ability to induce the GAs catabolic gene ZmGA2OX4, thus positively regulating growth (X. Wang et al., 2025). By contrast, tomato SlBBX20 negatively regulates SAS by inhibiting SlPIF4 activity through dimerization, thereby dampening the expression of SlIAA29, whose encoded protein downregulates the auxin‐repressor SlIAA17 (Shiose et al., 2024).
Altogether, the accumulated evidence reveals the multifunctional role of BBX proteins in coordinating abiotic stress responses. By integrating hormonal signaling, antioxidant defense and gene expression regulation, BBXs emerge as promising targets for the genetic improvement of crops facing challenging environmental conditions.
2. Biotic stress response
Beyond abiotic stresses, plants constantly face pathogen infection and insect herbivory, which can strongly limit growth. These challenges activate phytohormone‐mediated defense pathways: salicylic acid (SA) primarily mediates immunity against biotrophic pathogens and sap‐sucking insects, whereas JA confers resistance to necrotrophic pathogens and chewing insects. Multiple molecular mechanisms regulate the crosstalk between light‐driven growth and defense, explaining why rapid growth often compromises immunity. This trade‐off is particularly problematic under the high‐density planting typical of extensive agriculture (Ballaré & Pierik, 2017). As BBX proteins become increasingly recognized for their roles in light signaling, their involvement in biotic stress responses is also gaining attention. (Fig. 3; Table 1).
In sweet potato (Ipomoea batatas), IbBBX24 enhances resistance to Fusarium oxysporum by upregulating and downregulating the JA‐responsive TF MYELOCYTOMATOSIS ONCOGENE 2 (IbMYC2) and the JA response repressor IbJAZ10 by directly binding to genes' regulatory regions. Additionally, IbBBX24‐IbJAZ10 heterodimerization alleviates the IbJAZ10‐mediated repression of IbMYC2, triggering a JA‐mediated response (Zhang et al., 2020). By contrast, tomato SlBBX25 reduces SlMYC2 activity by competing for binding to the transcriptional regulator MEDIATOR COMPLEX SUBUNIT 25 (SlMED25), thereby attenuating JA signaling and reducing the resistance to the necrotrophic fungus Botrytis cinerea (Luo et al., 2023). Conversely, SlBBX20 confers fruit resistance to this fungus by promoting the accumulation of protective compounds, such as flavonoids and steroidal glycoalkaloids (Shiose et al., 2024).
The overexpression of the hybrid bamboo (Bambusa pervariabilis × Dendrocalamopsis grandis) BDBBX21 led to elevated antioxidant enzyme activity and improved disease resistance against Arthrinium phaeospermum fungus (Y. Liu et al., 2024). Similarly, the cucumber (Cucumis sativus), CsCOL9 increases tolerance to the whitefly Bemisia tabaci by enhancing antioxidant enzyme activity and hydrogen peroxide levels (Xie et al., 2025).
BBXs also mediate defense through pathways other than JA signaling. For instance, in lily (Lilium pumilum), LpBBX28‐LpNAC48 dimer upregulates the expression of trichome formation‐associated TF GLABRA3 LIKE (LpGL3L) promoting trichome initiation and aphid resistance (Xin et al., 2025). Another case was described in chrysanthemum, where susceptibility to Alternaria sp. infection increased upon the silencing of CmBBX32, due to an attenuated SA response (B. Wang et al., 2025). Two BBX‐mediated resistance mechanisms against Phytophthora have been characterized. In potato, StBBX27 induces the expression of the defense‐related genes StWRKY8 and SOMATIC EMBRYOGENESIS RECEPTOR KINASE 2 (StSERK2) in response to P. infestans infection, thereby enhancing plant immunity. As a counter‐defense strategy, the oomycete effector Pi18609 interacts with StBBX27, increasing the affinity for the CONSTITUTIVE PHOTOMORPHOGENIC 1 (StCOP1) ubiquitin ligase, promoting StBBX27 ubiquitination and, subsequent, degradation (Sun et al., 2025). Finally, resistance against P. capsici in pepper (Capsicum annuum) is promoted by CaBBX14 through the upregulation of the SA response (Zhou et al., 2023). CaBBX14 silencing reduces the expression of the SA biosynthetic gene PHENYLALANINE AMMONIA LYASE 1 (CaPAL1) and, consequently, of the SA‐induced defense genes, such as PATHOGENESIS RELATED 1 (CaPR1), CaPR5 and SYSTEMIC ACQUIRED RESISTANCE 8.2 (CaSAR8.2).
Specialized metabolism
VI.
Given the stress‐mitigating roles of specialized metabolites, such as flavonoids, anthocyanins and carotenoids in plants (Nawkar et al., 2024), along with their well‐documented health benefits in humans (Simsek & Whitney, 2024), enhancing the accumulation of these compounds in fruits and vegetables has become a major research priority.
Because light perception and signaling play pivotal roles in regulating specialized metabolism, and BBX proteins are key regulators of light responses, these proteins are increasingly recognized as major modulators of specialized metabolite synthesis and accumulation. They act either by directly activating biosynthetic genes or by regulating, transcriptionally and/or post‐translationally, TFs such as HY5 and the MYELOBLASTOSIS (MYBs), well‐known inducers of the flavonoid and anthocyanin biosynthesis (Fig. 4; Table 1). Such is the case of AtBBX20 that promotes anthocyanin synthesis by inducing AtHY5 expression and by cooperatively upregulating the anthocyanin biosynthesis‐related target genes through dimerization with AtHY5 (Wei et al., 2016).
B‐box (BBX) genes that regulate specialized metabolism in crop species. The species are indicated by the genus and species initials. Artemisia annua (Aa), Cannabis sativa (Csa), Capsicum annuum (Ca), Chrysanthemum morifolium (Cm), Citrus hindsii (Ch), Citrus sinensis (Cs), Fragaria × ananassa (Fa), Glycine max (Gm), Lilium spp (Lv), Malus domestica (Md), Mangifera indica (Mi), Petunia hybrida (Ph), Prunus avium (Pav), Prunus persica (Pp), Pyrus pyrifolia × Pyrus communis (Py), Rosa chinensis (Rc), Solanum lycopersicum (Sl), Solanum melongena (Sm) and Vitis vinifera (Vv). Black lines and white boxes represent DNA and genes, respectively. E/LBG means early and late biosynthetic genes. BBX proteins are represented in purple, HY5 in pink, MYELOBLASTOSIS (MYBs) in orange, other proteins in blue and hormones in yellow boxes. Blue lines indicate transcriptional regulation, red lines indicate post‐translational regulation, and green lines indicate the effect of the BBX protein on the phenotype. Arrows represent positive regulation, and blunt‐ended arrows represent inhibition. Solid and dotted lines indicate direct or indirect effects, respectively. Both gene and hormone names are written in red or blue depending on whether they have a negative or a positive effect on the phenotype, respectively. Gene, protein and hormone abbreviations are detailed in the text.
In tomato, while SlBBX20 alone induces the expression of CHALCONE SYNTHASE 1 (SlCHS1), CHALCONE ISOMERASE (SlCHI), FLAVANONE 3‐HYDROXYLASE (SlF3H) and FLAVONOL SYNTHASE (SlFLS), its heterodimerization with SlHY5 intensifies flavonoid biosynthetic genes transcription during fruit ripening (Shiose et al., 2024). In vegetative organs, the direct binding of SlBBX25 to the promoter of the biosynthetic gene DIHYDROFLAVONOL 4‐REDUCTASE (SlDFR) induces the accumulation of anthocyanins in tomato plants. In this system, SlBBX25 protein stability is modulated by ubiquitination via COP9 SIGNALOSOME SUBUNIT 5–2 (SlCSN5‐2; Luo et al., 2021). SlCOL1 (SlBBX3) promotes anthocyanin accumulation to counteract ROS accumulation under short‐day and low‐temperature conditions in tomato leaves. It induces the ANTHOCYANIN 1 (SlAN1) MYB TF expression, and subsequently, the formation of the SlCOL1‐SlAN1‐SlAN2 complex upregulates anthocyanin biosynthetic genes (Liu et al., 2025). In both tomato seedlings and fruits, SlBBX24 promotes anthocyanin accumulation by interacting with the ANTHOCYANIN 2 (SlAN2) and SlAN2‐LIKE TFs (He et al., 2023).
In apple fruits, MdMYB1 and MYB10 are the main activators of anthocyanin biosynthetic genes. MdMYB1 is activated by MdBBX20‐MdHY5 (Fang et al., 2019b) and MdBBX21‐MdHY5 (Zhang et al., 2021) dimers, while MdMYB10 expression is upregulated by MdBBX22‐MdHY5 (An et al., 2019). Moreover, MdBBX21 directly activates the expression of MdBBX20, MdBBX22 and MdHY5 (Zhang et al., 2021). Additionally, MdBBX22 upregulates the miRNA mdm‐miR858 that post‐transcriptionally represses MdMYB9/11/12, which are TFs that enhance proanthocyanidin biosynthesis. Since anthocyanin and proanthocyanidin compete for the same precursors, MdBBX22 ultimately enhances anthocyanin accumulation (Zhang et al., 2022). Conversely, MdBBX37 diminishes anthocyanin synthesis by inhibiting the transcriptional activity of MdMYB1/9 over MdDRF, ANTHOCYANIDIN SYNTHASE (MdANS) and UDP‐GALACTOSE FLAVONOID 3‐O‐GALACTOSYLTRANSFERASE (MdUFGT) via heterodimerization (An et al., 2020). MdCOL4 together with MdHY5 also inhibit the expression of MdMYB1 and the late anthocyanin biosynthetic genes MdANS and MdUFGT (Fang et al., 2019a). Finally, MdCOL6 also induces the transcription of MdANS and MdUFGT in fruits (Wang et al., 2023b). Together, these data demonstrate a highly intricate BBX‐mediated network regulating anthocyanin accumulation in apple fruits.
The regulation of HY5 activity by BBX proteins has also been reported in other species. In strawberry (Fragaria × ananassa), FaBBX22 interacts with FaHY5 to synergistically promote the expression of anthocyanin biosynthetic genes (Liu et al., 2022). Likewise, anthocyanin accumulation is enhanced by the FaMYB5–FaBBX24 dimer, which strengthens FaMYB5 activation of its target genes in the fruits (Zhang et al., 2023). In citrus (Citrus hindsii), the ChBBX22‐ChHY5 dimer promotes fruit anthocyanin accumulation not only by activating the promoters of the biosynthetic genes FLAVONOID 3′‐HYDROXYLASE (ChF3′H), ChDFR and ChANS but also by inducing the expression of the MYB TF ChRUBY1 responsible for the induction of the anthocyanin biosynthetic genes (Fu et al., 2024). By a similar mechanism, during blood orange (Citrus sinensis) ripening, CsBBX24 also promotes the expression of CsRUBY1 (Fu et al., 2025).
In peach fruits, PpBBX32 promotes anthocyanin accumulation by directly upregulating the TF PpMYB10.1 and by enhancing its ability to activate anthocyanin biosynthetic genes through heterodimerization (Huang et al., 2024). Similarly, in mango (Mangifera indica) fruit peel, MiBBX24‐ and MiBBX27‐dependent activation of MiMYB11 mediates the blue light‐induced anthocyanin accumulation (Pan et al., 2025). In eggplant (Solanum melongena), SmBBX22 directly induces SmCHS and SmDRF to promote anthocyanin accumulation (J. Li et al., 2025); whereas the palm leaf raspberry (Rubus chingii) RcBBX26 was also reported as an inductor of this pathway during fruit ripening (Xu et al., 2024).
In grape (Vitis vinifera), VvBBX44 is a negative regulator of anthocyanin accumulation that represses the expression of VvMYBA1, which in turn induces VvUFGT expression. Although VvBBX44 and VvMYBA1 do not interact, they form a feedback inhibitory loop to prevent pigments overaccumulation in grape berries (W. Liu et al., 2023). By contrast, during veraison, VvBBX8, VvBBX11 and VvBBX32 physically interact and induce the expression of their direct target BRASSINAZOLE‐RESISTANT 1 (VvBZR1), enhancing the BR‐triggered accumulation of anthocyanins in the berries (Qiu et al., 2025). Another case of BBX‐mediated anthocyanin accumulation via hormonal pathways has been reported in sweet cherry (Prunus avium). PavBBX6 and PavBBX9 induce light and ABA‐mediated anthocyanin accumulation by directly binding to the promoters of PavUFGT and 9‐CIS‐EPOXYCAROTENOID DIOXYGENASE 1 (PavNCED1), which encode key enzymes in anthocyanin and ABA biosynthesis, respectively (Wang et al., 2023a).
In pear fruits, both PyBBX16‐PyHY5 and PyBBX18‐PyHY5 heterodimers upregulate PyMYB10, inducing anthocyanin accumulation (Bai et al., 2019a,b). Adding an extra layer of control, PyBBX21 interacts with both PyHY5 and PyBBX18, limiting the formation of the PyHY5‐PyBBX18 transcription activator complex (Bai et al., 2019b). Additionally, PyBBX24‐PyHY5 dimer represses PyUFGT, PyCHS and PyMYB10 expression. This negative regulatory effect is alleviated through the PLANT U‐BOX TYPE E3 UBIQUITIN LIGASE 59 (PyPUB59)‐mediated PyBBX24 degradation (Yang et al., 2024; Li et al., 2025a). Different PyBBX24 alleles with nonsense mutations were identified in several varieties. In contrast to the wild‐type PyBBX24, the truncated proteins promote anthocyanin biosynthesis (Yang et al., 2024). Similarly, in cotton, a mutant allele of GhBBX24 (GhDR) results in a shortened protein that fails to repress the expression of PRODUCTION OF ANTHOCYNIN PIGMENT 1D (GhPAP1D) MYB TF, upregulating anthocyanin biosynthetic genes. Additionally, GhBBX21 is a positive regulator of GhPAP1D expression, inducing the pigment accumulation (X. Wang et al., 2023; Li et al., 2025b).
An interesting splicing‐dependent regulatory module involving GmBBX22 and different MYB TFs is required for the blue light‐induced biosynthesis of anthocyanins and flavonols in soybean seedlings (Zhan et al., 2025). Under blue light, GmBBX22 transcripts are alternatively spliced, producing 3 isoforms: X1, X2 and X3. The full‐length X1 directly induces GmCHI and GmF3H transcription, while the truncated isoforms X2 and X3 induce the expression of GmMYB12 and GmMYB90, respectively. This specific regulation differentially impacts the metabolic flux partitioning between flavonols and anthocyanins as GmMYB12 and GmMYB90 activate GmFLS and GmDFR, respectively (Zhan et al., 2025). Moreover, in seeds, GmBBX4 dimerizes with blue light‐induced TFs SOYBEAN TGACG‐MOTIF BINDING FACTOR 1 (STF1) and STF2, impairing their transcriptional activation of flavonoid biosynthetic genes (Song et al., 2024c).
Relatively fewer studies have investigated the roles of BBXs in controlling specialized metabolism in other organs than fruits and leaves. In flowers of C. morifolium, CmBBX20 was implicated as a positive mediator of light‐triggered accumulation of flavonoids and chlorogenic acid (CGA; Lu et al., 2024). Conversely, CmBBX28‐CmMYB9a dimer reduces the ability of CmMYB9a to activate CmCHS, CmDFR and CmUFGT (L. J. Zhou et al., 2025). A more complex regulatory module was identified during anthocyanin accumulation in Lilium spp. flowers. Light‐induced LvBBX24 interacts with the bZIP TF LvbZIP44 to promote LvMYB5 expression, which subsequently activates anthocyanin biosynthetic genes. In darkness, this pathway is suppressed through LvCOP1‐mediated degradation of LvBBX24 (Gao et al., 2024). In wheat (Triticum aestivum) grains, TaBBX‐3B interacts with TaHY5‐7A to activate PURPLE PERICARP‐MYB 1 (TaPPM1) TF, leading to the accumulation of anthocyanins in the pericarp (Jiang et al., 2023). Finally, in sweet potato, overexpression of IbBBX29 increases flavonoid accumulation in both leaves and storage roots by directly activating the flavonoid biosynthetic genes IbCHS, IbCHI1 and IbF3203'H (Gao et al., 2023).
Besides promoting anthocyanin accumulation in the mango fruit peels, MiBBX24 and MiBBX27 mediate blue light‐induced carotenoid accumulation in the fruit flesh by directly activating the expression of the biosynthetic gene PHYTOENE SYNTHASE (MiPSY; Pan et al., 2025). Similarly, SlBBX25 and CaBBX10, both subjected to DE‐ETIOLATED 1‐ (SlDET1) and CaCOP1‐mediated ubiquitination, directly upregulate PSY, leading to carotenoid accumulation in tomato and pepper fruits, respectively (Xiong et al., 2019; J. Wang et al., 2025). In addition, CaBBX20 silencing reduced carotenoid content in pepper fruits due to the downregulation of the CAPSANTHIN‐CAPSORUBIN SYNTHASE (CaCCS; Ma et al., 2023). Interestingly, during citrus fruit ripening, ChBBX22 and CsBBX24 also activate the PSY1 promoter, and in the case of ChBBX22, this activation is further enhanced through dimerization with ChHY5 (Fu et al., 2024, 2025).
The influence of BBXs regulation on Chl biosynthesis has been explored in petunia (Petunia hybrida) flowers, where PhCOL16a overexpression activates Chl biosynthetic genes (Ohmiya et al., 2019). In addition, the interaction of SlCOL1 and SlBBX24 with SlGLK2 stabilizes this master regulator of chloroplast development, leading to enhanced Chl accumulation in immature tomato fruits. In the absence of SlCOL1, SlBBX24 is unable to interact with SlGLK2, facilitating ubiquitin‐mediated degradation of SlGLK2, limiting Chl biosynthesis in the fruits (Cui et al., 2025). CaBBX10 also promotes Chl biosynthesis in green pepper fruits by activating the transcription of the MAGNESIUM CHELATASE SUBUNIT D (CaCHLD; J. Wang et al., 2025).
The Artemisia annua AaBBX21 was the first BBX identified as a regulator of terpenoid biosynthesis. The accumulation of the sesquiterpene lactone artemisinin is enhanced by the activity of the AaBBX21‐AaHY5 dimer that synergistically upregulates the transcription of GLANDULAR TRICHOME‐SPECIFIC WRKY 1 (AaGSW1), OVEREXPRESSION OF REGULATORY ACTIVITY (AaORA) and AaMYB108 genes. The interaction between AaBBX21 and AaHY5 with the ubiquitin ligase AaCOP1 reduces the abundance of both TFs, explaining the reduction of artemisinin content in the dark (He et al., 2024).
Finally, cannabis (Cannabis sativa) CsaCOL1, CsaCOL5 and CsaCOL7 were identified as regulators of key cannabinoid biosynthetic genes. While CsaCOL1 and CsaCOL7 induce the expression of GERANYLDIPHOSPHATE:OLIVETOLATE GERANYLTRANSFERASE 4 (CsaPT4) and CANNABIDIOLIC ACID SYNTHASE (CsaCBDAS), CsaCOL5 upregulates OLIVETOL SYNTHASE (CsaOLS) and CsaPT4. Strikingly, CsaCOL7 represses ACYL‐ACTIVATING ENZYME (CsaAAE) and CsaOLS, indicating a dual role fine‐tuning cannabinoid biosynthesis (M. Gao et al., 2025).
Conclusions and perspectives
VII.
The knowledge about the function of BBX proteins has grown substantially in recent years. The state‐of‐the‐art on A. thaliana and crop BBXs reveals an evolutionarily conserved mode of action across species. As covered at length in the text, in addition to regulating gene expression by directly binding to the promoters of their target genes, BBXs modulate the activity of other TFs, including the core light‐signaling regulators HY5 and PIFs. Furthermore, ubiquitin ligase complex–mediated post‐translational degradation is another regulatory level of BBX proteins.
Although significant advances have been made, numerous aspects still require refinement to achieve a comprehensive understanding of the multilayer functions of the BBX proteins and to translate this knowledge into effective strategies for crop improvement.
The phenotypic information obtained from BBX overexpression lines must be interpreted with caution. For instance, in tomato, overexpression of SlBBX17 impairs plant growth (Xu et al., 2022a,b), SlBBX24 affects fruit size (Cui et al., 2022), whereas SlBBX25 alters fruit number (Luo et al., 2023), and the accumulation of carotenoids (Xiong et al., 2019) and anthocyanins (Luo et al., 2023). Notably, these phenotypes were detected only in overexpressing lines and were absent in the corresponding loss‐of‐function mutants. Such discrepancies may arise from ectopic or supraphysiological gene expression driven by constitutive promoters and/or functional redundancy among BBX paralogs.
The few reports that have investigated the function of individual BBXs across the entire plant life cycle have consistently revealed their pleiotropic effects. For instance, tomato Slbbx26 displays accelerated flowering and delayed fruit ripening (Moreira et al., 2025). Similarly, Citrus hindsii ChBBX22 inhibits internode elongation, while promoting anthocyanin and carotenoid accumulation in fruits (Fu et al., 2024). However, most reports characterize BBX function in the context of isolated traits or developmental stages. This fragmented information not only limits our understanding of BBX function, but also overshadows the potential existence of functional orthology within phylogenetic orthologous genes. Besides, the establishment of evolutionary relationships among BBXs from different species is hindered by phylogenetic analyses that rely on alignment and tree reconstruction algorithms lacking methodological rigor and branch support. The analysis of the available information for A. thaliana and tomato shows that, while the studies for AtBBXs are almost restricted to flowering time and light responses, SlBBXs were investigated in the context of other agronomically appealing processes, such as the regulation of fruit yield, stress responses and specialized metabolism (Fig. 1; Table 1). Based on current knowledge, only two cases of phylogenetic and functional orthology have been established between AtBBXs and SlBBXs: SlBBX25‐AtBBX20 (both positive regulators of specialized metabolism), and SlBBX1‐AtBBX1 (both flowering time regulators, albeit in opposite directions; Fig. 1; Table 1). In light of this scenario, comprehensively analyzing the roles of individual BBXs throughout the plant life cycle is paramount for assessing the impacts of their genetic manipulation for crop improvement. Based on this knowledge, the potential of BBX‐based breeding strategies can be further exploited by designing temporal‐ and organ‐specific manipulations to avoid undesirable trade‐offs (Fig. 5a).
Future perspectives. A roadmap for elucidating the B‐box (BBX) genetic diversity, regulatory mechanisms and multilayered functions to facilitate future BBX‐based breeding strategies. (a) Comprehensive analysis of BBX effects throughout the plant life cycle is essential to enable precise time‐ and organ‐specific manipulations and to prevent undesirable outcomes. (b) Current evidence indicates that individual BBX proteins mediate several stress responses and coordinate defense mechanisms with developmental programs by modulating the activity of regulatory factors. However, there remains a knowledge gap regarding the role of BBXs in the molecular signaling crosstalk triggered by multi‐stress stimuli. Protein abbreviations are detailed in the text. (c) The generation and functional characterization of mutant collections targeting single and multiple BBX genes (e.g. Clustered Regularly Interspaced Short Palindromic Repeats‐mediated mutants) together with the exploration of their natural allelic diversity will reveal underexplored genetic resources for the modulation of important agronomical traits in crops. (d) Although evidence suggests that BBX genes are regulated by and also regulate target genes through epigenetic mechanisms, a substantial knowledge gap remains in this area. HME indicates histone‐modifying enzymes. (e) Recently, it has been reported that the AtCO‐mediated induction of AtFT, which triggers the vegetative‐to‐reproductive program transition in A. thaliana, is not only driven by its interaction with AtNF‐YB and AtNF‐YC, but also by the proper stoichiometry of these three proteins, determining the formation of a functional biomolecular condensate within the cell nucleus. Further research on how BBX‐containing protein complexes are regulated through liquid–liquid phase separation will enhance our understanding of the regulatory landscape modulated by BBX proteins.
Moreover, BBXs are increasingly recognized as key regulators of responses to diverse environmental stresses. Notably, there are examples of individual BBX regulating more than one stress‐response pathway. For example, in tomato, SlBBX20 is required not only for the shade avoidance response but also for the resistance to Botrytis cinerea (Shiose et al., 2024). The current evidence derived from single‐stress experiments does not reflect field conditions in which several stresses occur simultaneously. Thus, further studies using multi‐stress experimental approaches are needed to elucidate BBX role within the molecular signaling crosstalk (Fig. 5b).
As mentioned above, recent studies indicate that natural allelic variation in BBX genes regulates key agronomic traits, representing an underexplored resource for biotechnological applications (Fig. 5c). However, the pleiotropic nature of BBX proteins, and the fact that several family members interact with the same regulatory partners (e.g. HY5 and PIFs), suggests that functional redundancy may occur within the family. While the roles of individual BBX genes were characterized through single‐gene interventions, relatively few studies have examined redundancy among paralogs. Notably, to date, all reported cases of redundancy correspond to ‘partial redundancy’, as the phenotypes of single mutants are enhanced when combined in double mutants, indicating additive effects. Examples include AtBBX24/25 in shade‐induced hypocotyl elongation (Gangappa et al., 2013), and AtBBX20/21/22 (Bursch et al., 2020) and AtBBX22/23 (X. Zhang et al., 2017) during seedling photomorphogenesis. In rice, OsCOL4/13 also exhibit partial redundancy flowering time repressors (Sheng et al., 2016).
Thus, developing and systematically characterizing single and multiple BBX mutant collections will elucidate the regulatory complexity of this protein family and be crucial for translating mechanistic insights into biotechnological strategies (Fig. 5c).
Epigenetic regulation is another aspect of BBX biology that has lagged behind. In pear, analysis of DNA methylation during light‐induced anthocyanin accumulation led to the identification of a BBX gene whose promoter undergoes light‐dependent demethylation (Liu et al., 2022). In A. thaliana, the positive regulator of photomorphogenesis AtBBX22 is transcriptionally repressed in darkness through HISTONE DEACETYLASE 19–mediated histone deacetylation (Jing et al., 2021). Moreover, transcription of the flowering inducer AtCO requires the removal of the repressive histone mark H3K9me2 by the demethylase JUMONJI 28 (AtJMJ28; Hung et al., 2021). Subsequently, AtCO interacts with the chromatin remodeling factor PICKLE (AtPKL), enhancing chromatin accessibility at the AtFT locus through increased histone H3 acetylation (Jing et al., 2019). AtCO also interacts with the histone‐methylation reader AtMRG2, which recognizes H3K4me and H3K36me, reinforcing AtCO binding to the AtFT promoter (Bu et al., 2014). Furthermore, AtCO promotes transcription of the floral integrator SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (AtSOC1) by recruiting NUCLEAR FACTOR Y (NF‐Y) to its promoter, leading to histone demethylation and consequent transcriptional activation (Hou et al., 2014). Collectively, these findings demonstrate that epigenetic regulation operates both upstream and downstream of BBX proteins, influencing their expression and shaping the transcriptional output of their target genes. This underscores the need for deeper investigation into how BBXs and epigenetic mechanisms jointly orchestrate gene expression (Fig. 5d).
Finally, a few reports have described the same BBX differentially regulating the same biological process or metabolic pathway in different physiological contexts or organs. In barley, HvCO1 and HvCO2 regulate flowering by inhibiting or inducing HvFT1 before or after vernalization, respectively (Mulki & von Korff, 2016). The tomato SlBBX26 impairs SlPIF4 activity by heterodimerization, preventing GAs accumulation by downregulating GA biosynthetic genes in internodes and upregulating GA catabolic genes in fruits (Moreira et al., 2025). At first glance, these observations may appear contradictory; however, the repertoire of TFs in the nucleus varies across different cell types and developmental stages, leading to the formation of diverse biomolecular complexes. These protein–DNA complexes aggregate into membraneless compartments known as biomolecular condensates, which regulate gene expression by modulating transcriptional activity, chromatin remodeling, RNA processing, protein transport and degradation. Droplet condensates are formed through liquid–liquid phase separation (LLPS) driven by the liquid‐like properties of some of their protein constituents. Several studies have linked LLPS‐derived condensates to the regulation of plant developmental processes and stress responses (Kim et al., 2021; Q. Liu et al., 2024). In this sense, it was recently characterized that AtCO undergoes LLPS, creating condensates together with AtNF‐YB, AtNF‐YC and AtFT promoter region. The proper formation of this cluster is essential for the AtCO‐mediated activation of AtFT expression and, consequently, floral transition (Huang et al., 2025; Fig. 5e). This is the only experimental evidence of a BBX acting in LLPS opens up a promising field of study for further research. In fact, we predicted the overall probability to undergo LLPS, revealing that 18 AtBBXs are high‐confidence droplet drivers capable of spontaneously undergoing LLPS, whereas 12 AtBBXs are droplet clients that assemble into biomolecular condensates through interactions with droplet drivers (Fig. 6). Notably, this predicted condensate‐forming capacity does not correlate with either protein domain topology or DNA‐binding activity (Table S3). It is worth mentioning that the proper condensate functionality depends on the correct stoichiometry of its constituents; thus, their abundance is tightly regulated. This finding unveils an uncharted area for deciphering the molecular mechanisms governing the function of this multigene family.
Probability of AtBBXs to undergo liquid–liquid phase separation. Graphical representation of the estimated probability of each amino acid residue to promote liquid–liquid phase separation (threshold = 0.6). The overall probability of spontaneous liquid–liquid phase separation (pLLPS) for each protein is indicated along with the corresponding protein name. Proteins whose chart is colored in blue are classified as droplet‐drivers (pLLPS ≥ 0.60), which can spontaneously undergo LLPS; those in orange are droplet‐clients (pLLPS < 0.6, but containing droplet‐promoting regions) that can compose biomolecular condensates through the interaction with droplet‐driver proteins, while in red are the proteins unlikely to undergo liquid–liquid phase separation. The gray areas indicate the domains (B‐box and CCT) within each protein. The prediction was performed with FuzDrop (Hatos et al., 2022). BBX, B‐box; CCT, CONSTANS, CONSTANS‐like and TIMING OF CAB1.
In conclusion, BBX proteins are key factors that enable plants to integrate environmental signals into developmental programs (Fig. 7a). Continued research will uncover the full extent of BBX functional diversity and the variety of molecular mechanisms underlying the multiple regulatory levels at which they operate, providing a foundation for the development of precise biotechnological strategies. Then, to improve crop performance and stress resilience under changing environmental conditions, specific BBX genes can be targeted in particular organs and/or physiological processes. Their activity can be attenuated or enhanced using modern genetic modification tools, such as promoter or gene sequence editing, gene silencing or overexpression (Fig. 7b).
B‐box (BBX) proteins bridge environmental stimuli and developmental reprogramming, being candidates for crop breeding strategies. (a) Upon perception of environmental cues, including light intensity and quality, temperature, water availability and biotic stress, BBX proteins initiate adaptive reprogramming of gene expression. This regulation occurs through the direct binding of BBX proteins to promoter regions of target genes and/or through the assembly of multiprotein complexes comprising BBX‐interacting proteins (BIPs), histone‐modifying enzymes (HMEs) and additional transcription factors (TFs). HY5 and PIFs have been extensively reported as BIPs, being key hubs in BBX mode of action. These interactions can be facilitated by the formation of biomolecular condensates whose composition and dynamics are specific to tissue type, environmental context and developmental stage. The abundance of individual condensate components is tightly controlled at both transcriptional and post‐translational levels (e.g. via ubiquitin‐mediated degradation), as proper condensate function depends on precise stoichiometric balance. Collectively, this BBX‐centered signaling framework enables the plant developmental program to be dynamically reconfigured in accordance with prevailing environmental conditions. (b) As the functions of BBX proteins are more precisely defined, biotechnological interventions can be developed using genetic engineering approaches. Genome editing tools, such as CRISPR/nuclease systems, can be employed to generate null alleles of BBX genes acting as negative regulators of desirable traits. In addition, advanced CRISPR‐based strategies, including base editing and prime editing, allow precise modification of specific gene regions. These approaches enable the fine‐tuning of BBX gene regulation through targeted alterations in promoter sequences, the modification of BBX‐coding regions to disrupt interactions with repressor proteins, or the engineering of BBX downstream target genes to enhance or reduce BBX binding to their promoters and/or protein products. Furthermore, transgenic strategies using cell‐type‐ or stimulus‐specific promoters can achieve precise overexpression or silencing of BBX genes, allowing spatially and temporally controlled modulation of BBX activity. Collectively, these biotechnological strategies hold substantial potential for improving plant yield, development, stress resilience and nutraceutical content. LB, T‐DNA left border; RB, T‐DNA right border; SP, specific promoter; SM, selective marker.
Competing interests
None declared.
Disclaimer
The New Phytologist Foundation remains neutral with regard to jurisdictional claims in maps and in any institutional affiliations.
Supporting information
Table S1 Functions of AtBBX loci. Table S2 Genome‐wide identification of BBX gene family. Table S3 AtBBXs DNA interaction.Please note: Wiley is not responsible for the content or functionality of any Supporting Information supplied by the authors. Any queries (other than missing material) should be directed to the New Phytologist Central Office.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alvarez‐Fernandez R , Penfold CA , Galvez‐Valdivieso G , Exposito‐Rodriguez M , Stallard EJ , Bowden L , Moore JD , Mead A , Davey PA , Matthews JSA et al. 2021. Time‐series transcriptomics reveals a BBX 32‐directed control of acclimation to high light in mature Arabidopsis leaves. The Plant Journal 107: 1363–1386.34160110 10.1111/tpj.15384 · doi ↗ · pubmed ↗
- 2An JP , Wang XF , Espley RV , Lin‐Wang K , Bi SQ , You CX , Hao YJ . 2020. An apple B‐Box protein Md BBX 37 modulates anthocyanin biosynthesis and hypocotyl elongation synergistically with Md MY Bs and Md HY 5. Plant and Cell Physiology 61: 130–143.31550006 10.1093/pcp/pcz 185 · doi ↗ · pubmed ↗
- 3An JP , Wang XF , Zhang XW , Bi SQ , You CX , Hao YJ . 2019. Md BBX 22 regulates UV‐B‐induced anthocyanin biosynthesis through regulating the function of Md HY 5 and is targeted by Md BT 2 for 26S proteasome‐mediated degradation. Plant Biotechnology Journal 17: 2231.31222855 10.1111/pbi.13196 PMC 6835122 · doi ↗ · pubmed ↗
- 4An JP , Wang XF , Zhang XW , You CX , Hao YJ . 2021 a. Apple B‐box protein BBX 37 regulates jasmonic acid mediated cold tolerance through the JAZ‐BBX 37‐ICE 1‐CBF pathway and undergoes MIEL 1‐mediated ubiquitination and degradation. New Phytologist 229: 2707–2729.33119890 10.1111/nph.17050 · doi ↗ · pubmed ↗
- 5An JP , Zhang XW , Liu YJ , Zhang JC , Wang XF , You CX , Hao YJ . 2021 b. Md ABI 5 works with its interaction partners to regulate abscisic acid‐mediated leaf senescence in apple. The Plant Journal 105: 1566–1581.33314379 10.1111/tpj.15132 · doi ↗ · pubmed ↗
- 6Armougom F , Moretti S , Poirot O , Audic S , Dumas P , Schaeli B , Keduas V , Notredame C . 2006. Expresso: automatic incorporation of structural information in multiple sequence alignments using 3D‐Coffee. Nucleic Acids Research 34: W 604–W 608.16845081 10.1093/nar/gkl 092PMC 1538866 · doi ↗ · pubmed ↗
- 7Atanasov V , Schumacher J , Muiño JM , Larasati C , Wang L , Kaufmann K , Leister D , Kleine T . 2024. Arabidopsis BBX 14 is involved in high light acclimation and seedling development. The Plant Journal 118: 141–158.38128030 10.1111/tpj.16597 · doi ↗ · pubmed ↗
- 8Bai S , Tao R , Tang Y , Yin L , Ma Y , Ni J , Yan X , Yang Q , Wu Z , Zeng Y et al. 2019 a. BBX 16, a B‐box protein, positively regulates light‐induced anthocyanin accumulation by activating MYB 10 in red pear. Plant Biotechnology Journal 17: 1985–1997.30963689 10.1111/pbi.13114 PMC 6737026 · doi ↗ · pubmed ↗