Pecbloodin18-37: a promising antimicrobial peptide from Boleophthalmus pectinirostris with therapeutic potential against Edwardsiella tarda infection
Yuqi Bai, Wenbin Zheng, Weibin Zhang, Jingyuan Zhan, Fangyi Chen, Ke-Jian Wang

TL;DR
A new antimicrobial peptide from mudskippers shows strong potential to combat Edwardsiella tarda infections in aquaculture without promoting antibiotic resistance.
Contribution
Discovery and characterization of Pecbloodin18-37, a novel antimicrobial peptide with potent activity against Edwardsiella tarda and no resistance development.
Findings
Pecbloodin18-37 rapidly kills bacteria by damaging membranes and inducing reactive oxygen species.
The peptide inhibits biofilm formation and does not promote resistance even with prolonged exposure.
In mudskippers infected with E. tarda, Pecbloodin18-37 improved survival and restored immune balance.
Abstract
Edwardsiella tarda is a significant pathogen in aquaculture, causing severe systemic infections and considerable economic losses worldwide. The limited efficacy of current treatments and the emergence of multidrug-resistant strains underscore the urgent need for novel antimicrobial strategies. Antimicrobial peptides have garnered increasing interest due to their broad-spectrum activity and low risk of resistance development. In this study, we identified a novel immune-related gene from the mudskipper Boleophthalmus pectinirostris, named Pecbloodin, which encodes a mature peptide of 67 amino acids. A truncated peptide derived from this sequence, Pecbloodin18-37, exhibited broad-spectrum antibacterial activity, high thermal stability, and tolerance to sodium ions. Functional assays demonstrated that Pecbloodin18-37 exerts rapid bactericidal effects by compromising bacterial membrane…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
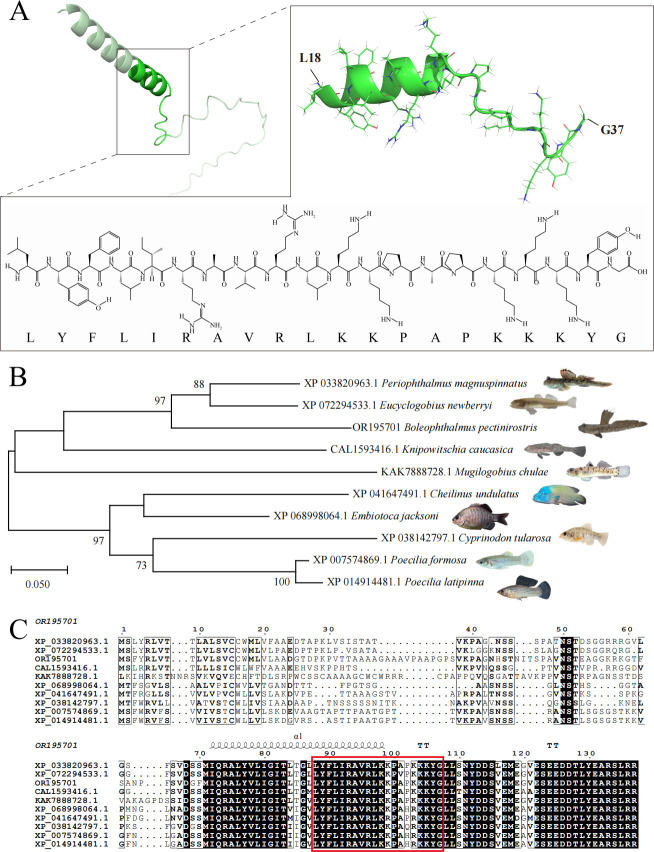 Fig 1
Fig 1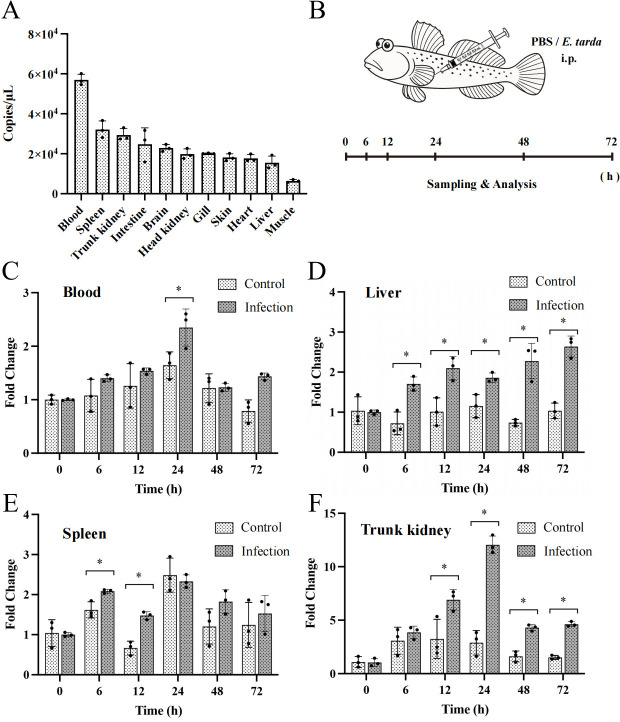 Fig 2
Fig 2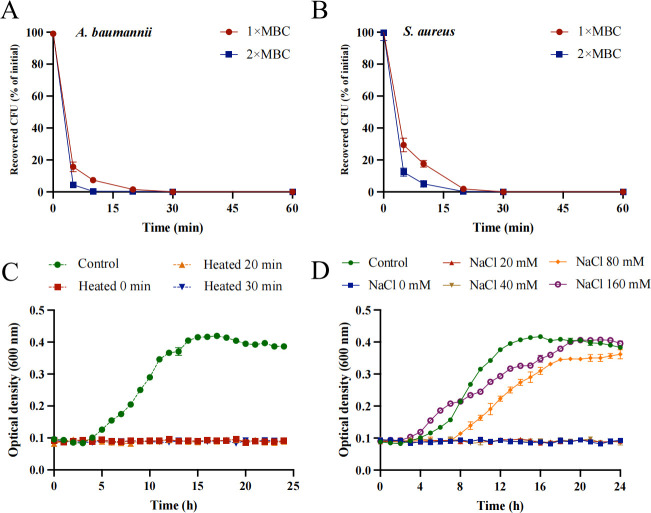 Fig 3
Fig 3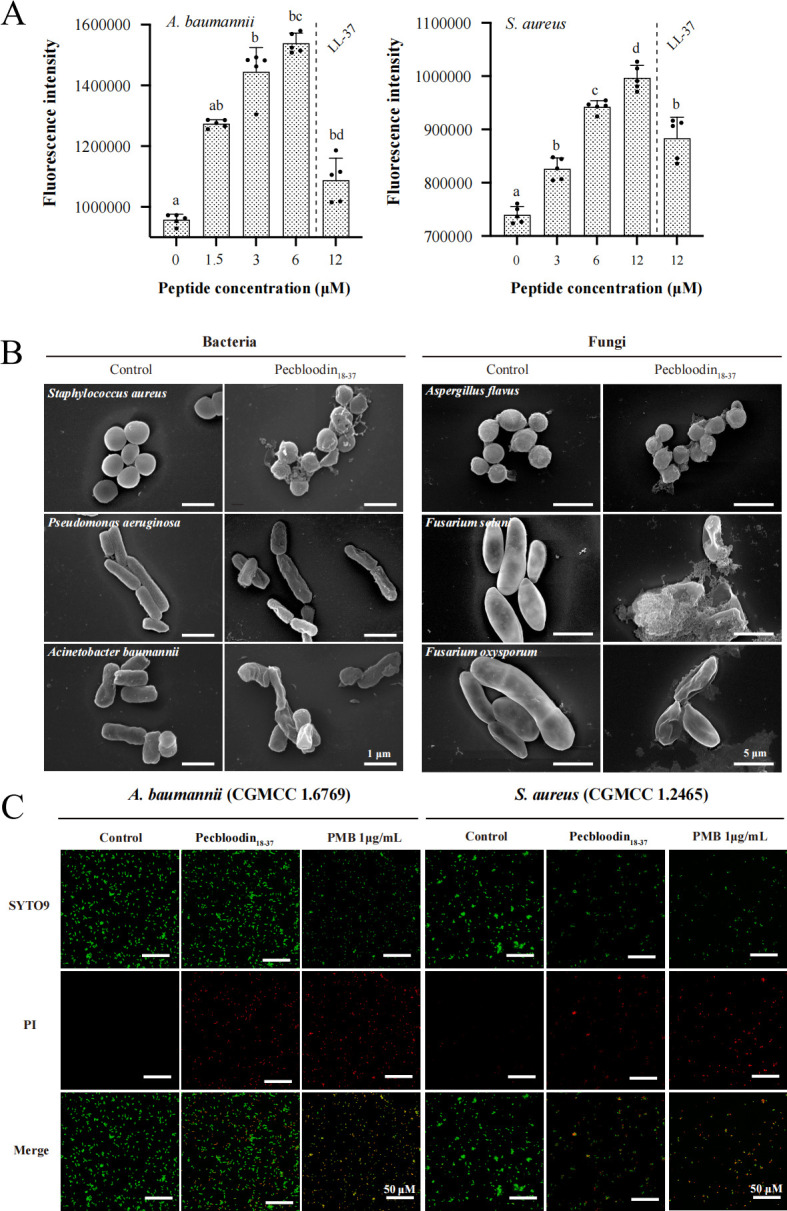 Fig 4
Fig 4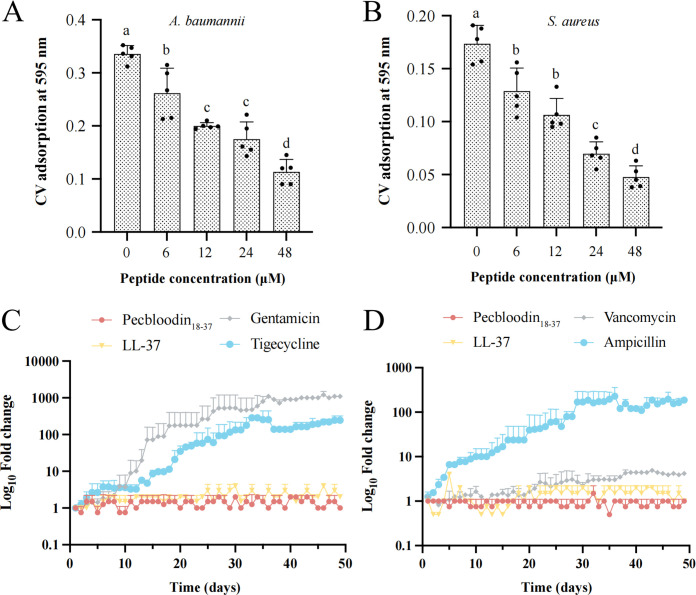 Fig 5
Fig 5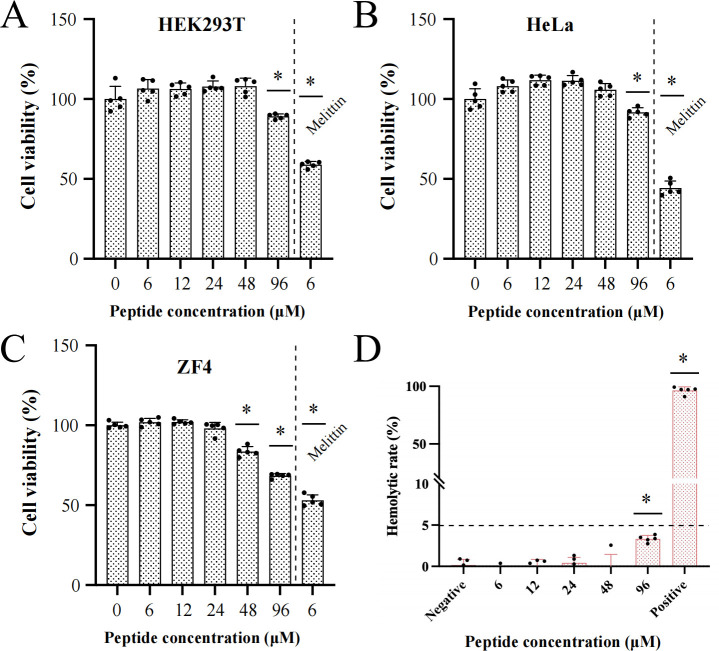 Fig 6
Fig 6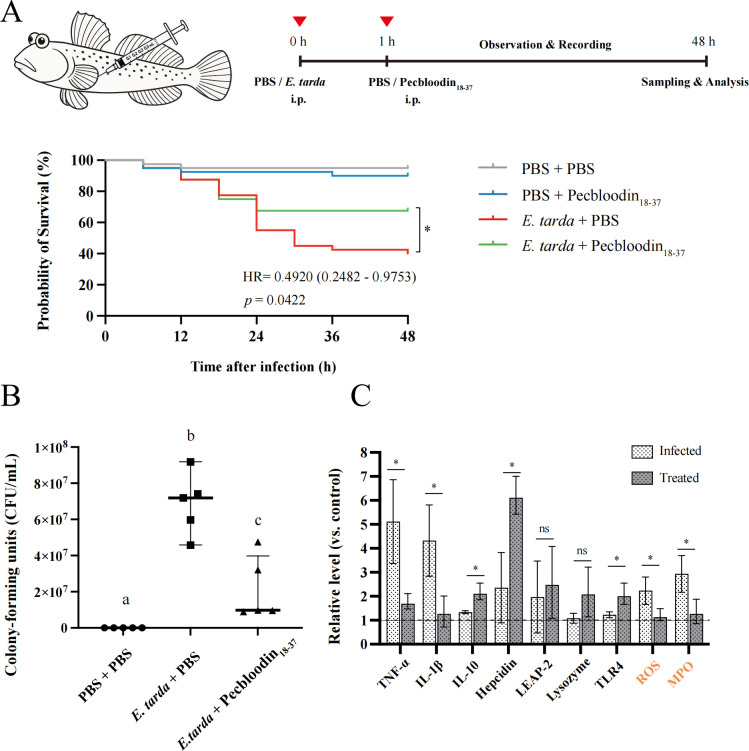 Fig 7
Fig 7| Physicochemical parameter |
| Pecbloodin18-37 | LL-37 |
|---|---|---|---|
| Number of amino acids (aa) | 67 | 20 | 37 |
| Molecular weight (Da) | 7,751.04 | 2,390.00 | 4,493.32 |
| Theoretical pI | 7.98 | 10.73 | 10.61 |
| Molecular formula | C349H570N92O102S2 | C116H193N31O23 | C205H340N60O53 |
| Total number of atoms | 1,115 | 363 | 658 |
| Grand average of hydropathicity | −0.219 | −0.410 | −0.724 |
| Total hydrophobic ratio (%) | 39 | 40 | 35 |
| Total net charge | +1 | +7 | +6 |
| Strain | Pecbloodin18-37 | LL-37 MIC (μM) | ||
|---|---|---|---|---|
| CGMCC no. | MIC | MBC | ||
| Gram-negative bacteria | ||||
| | 1.2385 | 24–48 | 24–48 | 12–24 |
| | 1.2421 | 1.5–3 | 3–6 | 6–12 |
| | 1.6769 | 1.5–3 | 1.5–3 | 3–6 |
| | 1.1872 | 12–24 | 24–48 | 48–96 |
| | 1.2017 | 12–24 | 24–48 | 48–96 |
| | 1.1833 | 3–6 | 6–12 | 12–24 |
| Gram-positive bacteria | ||||
| | 1.10753 | 1.5–3 | 1.5–3 | 1.5–3 |
| | 1.4260 | 12–24 | 12–24 | 3–6 |
| | 1.2465 | 3–6 | 3–6 | 6–12 |
| | 1.1886 | <1.5 | 1.5–3 | 1.5–3 |
| | 1.2135 | 3–6 | 3–6 | 6–12 |
| | 1.3760 | 3–6 | 3–6 | <1.5 |
| Fungi | ||||
| | 2.1563 | <1.5 | <1.5 | 1.5–3 |
| | 2.2411 | 3–6 | 6–12 | 6–12 |
| | 3.4410 | 24–48 | 48–96 | 12–24 |
| | 3.6785 | 12–24 | 12–24 | 12–24 |
| | 3.5840 | 6–12 | 6–12 | 6–12 |
| Multidrug-resistant strains | ||||
| MRSA-QZ19130 | – | 3–6 | 6–12 | 12–24 |
| MRSA-QZ19134 | – | 3–6 | 3–6 | 12–24 |
| | – | 3–6 | 6–12 | 6–12 |
| | – | 1.5–3 | 3–6 | 6–12 |
| | – | 1.5–3 | 1.5–3 | 3–6 |
| | – | 3–6 | 3–6 | 6–12 |
- —China Postdoctoral Science Foundationhttp://dx.doi.org/10.13039/501100002858
- —the Xiamen Ocean Development Bureau
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —the Fujian Ocean and Fisheries Bureau
- —the Fujian Ocean and Fisheries Bureau
- —the Postdoctoral Fellowship Program of CPSF
- —the Pingtan Research Institute of Xiamen University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial Peptides and Activities · Invertebrate Immune Response Mechanisms · Aquaculture disease management and microbiota
INTRODUCTION
Bacterial pathogens, such as Edwardsiella, Aeromonas, Vibrio, and Pseudomonas species, are widespread in aquaculture systems and can rapidly cause mass mortality events, leading to substantial economic losses estimated at approximately 6 billion USD annually (1). Among them, the genus Edwardsiella encompasses several important pathogens, including Edwardsiella tarda, Edwardsiella ictaluri, Edwardsiella piscicida, and Edwardsiella anguillarum, which pose significant threats to both aquaculture and public health on a global scale (2). Notably, E. tarda is the only species within this genus known to infect humans. It is ubiquitous in natural environments and is recognized as one of the primary pathogens affecting farmed freshwater and marine fish worldwide (3). Infections caused by E. tarda typically result in a systemic condition known as edwardsiellosis, which is characterized by ascites, severe internal organ lesions, exophthalmia, and herniation (4). Since its first reported outbreak in 1962, edwardsiellosis has inflicted severe economic damage across more than 20 commercially important fish species, including tilapia (Tilapia nilotica), freshwater catfish (Tandanus tandanus), Japanese eel (Anguilla japonica), largemouth bass (Micropterus salmoides), rainbow trout (Oncorhynchus mykiss), turbot (Scophthalmus maximus), and mullet (Mugil cephalus) (5, 6). Currently, antibiotics remain the primary strategy for controlling E. tarda infections. However, increasing antibiotic resistance in E. tarda, particularly to cephalosporins, aminoglycosides, and penicillin, has significantly compromised treatment efficacy (7). Consequently, the development of novel, effective antimicrobial agents with anti-E. tarda activity is urgently needed.
Antimicrobial peptides (AMPs) are key effectors of the innate immune system, capable of mounting rapid and broad-spectrum defense responses against a wide range of pathogens, including bacteria, viruses, fungi, and parasites (8). AMPs can be naturally isolated from various organisms, obtained as peptide derivatives, or synthesized artificially. They are typically characterized by structural features such as hydrophobicity, amphiphilicity, and a net positive charge, although a small subset of AMPs is anionic. In addition, AMPs often adopt α-helical or β-sheet conformations (9). These properties enable AMPs to interact with anionic components on microbial membranes, leading to membrane disruption and leakage of intracellular contents. In some cases, AMPs can also exert intracellular effects, ultimately resulting in microbial death (10). Due to their multiple modes of action and rapid bactericidal activity, the likelihood of pathogens developing resistance to AMPs is considerably lower than to traditional antibiotics (11). These advantages position AMPs as promising alternatives to antibiotics in aquaculture.
To date, more than 3,300 AMPs have been identified across a wide range of organisms. However, fish-derived AMPs account for less than 5% of these peptides (Antimicrobial Peptide Database). Marine fish, due to their adaptation to environments characterized by high salinity, fluctuating temperatures, and high microbial pressure, represent a valuable reservoir for novel AMP discovery (12). The mudskipper Boleophthalmus pectinirostris, a unique amphibious teleost, inhabits estuarine and intertidal zones where it is exposed to complex environmental stressors. This ecological niche likely necessitates an enhanced reliance on innate immune effectors. To date, only five AMPs have been identified from B. pectinirostris, including BpLEAP-2 (13), BpHep-1 and BpHep-2 (14), BpNKL (15), and Bolespleenin_334-347_ (16). Among them, the first four belong to previously characterized AMP families, whereas Bolespleenin_334-347_ represents a newly discovered AMP with distinct structural features. Further exploration of novel AMPs from B. pectinirostris is essential to better understand its immune defense strategies and provide a foundation for the development of new antimicrobial agents.
In the present study, we identified a novel immune-related gene from B. pectinirostris, designated as Pecbloodin. The full-length cDNA of Pecbloodin was obtained using rapid amplification of cDNA-PCR (RACE-PCR), and its tissue expression profile was analyzed via quantitative real-time PCR (qPCR). A predicted AMP derived from this gene, named Pecbloodin_18-37_, was chemically synthesized based on CAMP_R4_ prediction, and its antimicrobial activity was evaluated. We investigated its mechanism of action, assessed its ability to circumvent bacterial resistance, and tested its efficacy using a B. pectinirostris-E. tarda infection model. The expression levels of immune-related genes (TNF-α, IL-1β, IL-10, and TLR4) and AMP-related genes (Hepcidin, LEAP-2, and lysozyme) in the liver were examined. Additionally, reactive oxygen species (ROS) levels and myeloperoxidase (MPO) activity were measured. These findings are expected to provide theoretical and practical insights into the development of novel anti-E. tarda agents with low risk of resistance development.
RESULTS
Sequence analysis of Pecbloodin and design of the truncated peptide
The full-length cDNA sequence of Pecbloodin was successfully obtained and submitted to the GenBank database (accession No. OR195701). As shown in Fig. S1, the gene contains a 29-bp 5′ untranslated region (UTR) and a 366-bp 3′ UTR, with an open reading frame encoding a protein of 67 amino acids. The predicted molecular weight of the encoded protein is 7.75 kDa, and the theoretical isoelectric point (pI) is 7.98 (Table 1). Bioinformatic analysis identified a 20-residue peptide fragment within Pecbloodin, designated as Pecbloodin_18-37_ (H-LYFLIRAVRLKKPAPKKKYG-OH), with potential antimicrobial properties. As shown in Fig. 1A, the predicted tertiary structure of Pecbloodin_18-37_ adopts a typical α-helical conformation and contains seven basic amino acids, including lysine and arginine, which contribute to its cationic nature. Physicochemical property analysis revealed that Pecbloodin_18-37_ is an amphiphilic cationic peptide, with a net positive charge of +7 and a hydrophobicity of 40% (Table 1). Machine learning-based AMP prediction using the CAMP_R4_ platform indicated high confidence in its antimicrobial potential, with probability scores of 0.96 (Artificial Neural Network), 0.99 (Support Vector Machine), and 0.95 (Random Forest), collectively classifying Pecbloodin_18-37_ as an AMP.
Structural analysis of Pecbloodin and Pecbloodin18-37, phylogenetic tree construction, and sequence alignment with homologs. (A) Predicted three-dimensional structures of Pecbloodin and its truncated peptide Pecbloodin18-37 generated by AlphaFold. (B) Phylogenetic tree of Pecbloodin and its homologs constructed using the neighbor-joining method in MEGA, with 1,000 bootstrap replicates. (C) Multiple sequence alignment of Pecbloodin with homologous proteins from other species. The amino acid sequence of Pecbloodin18-37 is highlighted in red within the boxed region.
Sequence alignment showed that the Pecbloodin protein shares 98.51% amino acid identity with two uncharacterized proteins from Periophthalmus magnuspinnatus (XP_033820963.1) and Eucyclogobius newberryi (XP_072294533.1). Phylogenetic analysis further confirmed that these homologous sequences cluster within the same evolutionary clade, and the truncated peptide region is highly conserved among them (Fig. 1B and C).
Expression patterns of the Pecbloodin gene
The tissue distribution of Pecbloodin expression in B. pectinirostris was analyzed by qPCR (Fig. 2A). The gene was found to be most highly expressed in blood, while the lowest expression level was observed in muscle.
*Tissue-specific expression and temporal regulation of Pecbloodin in B. pectinirostris following E. tarda infection. (A) Expression levels of Pecbloodin in various tissues under basal conditions (n = 3). (B) Schematic diagram of E. tarda infection and sampling timeline. (C–F) Temporal expression of Pecbloodin in blood, liver, spleen, and trunk kidney after infection (n = 3). Asterisks indicate statistically significant differences compared to the control group (P < 0.05).
To investigate the immune response of Pecbloodin, its expression was further assessed in B. pectinirostris following E. tarda infection (Fig. 2B). Upon infection, Pecbloodin expression was significantly upregulated in the blood at 24 h post-infection (hpi) (Fig. 2C), in the liver at 6, 12, 24, 48, and 72 hpi (Fig. 2D), in the spleen at 6 and 12 hpi (Fig. 2E), and in the trunk kidney at 12, 24, 48, and 72 hpi (Fig. 2F). These results suggest that Pecbloodin is responsive to E. tarda infection and may play a role in the innate immune defense of multiple tissues.
Antimicrobial activity of Pecbloodin18-37
The antimicrobial activity of Pecbloodin_18-37_ was evaluated against a panel of microorganisms, as summarized in Table 2. The peptide exhibited broad-spectrum antibacterial activity. It effectively inhibited the growth of various gram-negative bacteria, including Pseudomonas aeruginosa, Acinetobacter baumannii, and Vibrio alginolyticus, as well as gram-positive bacteria, such as Listeria monocytogenes, Staphylococcus aureus, Corynebacterium glutamicum, Enterococcus faecalis, and Bacillus cereus, with minimum inhibitory concentration (MIC) values ≤6 µM and minimum bactericidal concentration (MBC) values ≤12 µM.
In addition to its antibacterial effects, Pecbloodin_18-37_ demonstrated antifungal activity against both yeast-like fungi and filamentous fungi, including Cryptococcus neoformans, Candida albicans, Fusarium oxysporum, and Fusarium solani, with MIC values ≤ 12 µM and MBC values ≤ 24 µM. The peptide also inhibited the conidial germination of filamentous fungi, such as F. solani, Fusarium graminearum, Aspergillus flavus, and F. oxysporum, as illustrated in Fig. S2.
Furthermore, the efficacy of Pecbloodin_18-37_ was assessed against clinically isolated multidrug-resistant (MDR) strains. Remarkably, the peptide showed potent inhibitory effects against MRSA (QZ19134), MDR P. aeruginosa (QZ19122), and MDR A. baumannii (QZ18050 and QZ18055), with MIC values as low as 6 μM. These findings suggest that Pecbloodin_18-37_ may serve as a promising candidate for combating antibiotic-resistant pathogens.
Bactericidal kinetics and stability of Pecbloodin18-37
Time-kill kinetic assays demonstrated the rapid bactericidal activity of Pecbloodin_18-37_. At 1× MBC, the peptide reduced the viability of both A. baumannii and S. aureus by over 99% within 30 min. At a higher concentration (2× MBC), this >99% reduction was achieved even faster—within 10 min for A. baumannii (Fig. 3A) and 20 min for S. aureus (Fig. 3B).
Bactericidal kinetics and stability of Pecbloodin18-37. (A and B) Time-dependent killing curves of Pecbloodin18-37 against A. baumannii and S. aureus at 1× and 2× MBC. (C) Thermal stability of Pecbloodin18-37 after treatment at 100°C for up to 30 min. (D) Antibacterial activity of Pecbloodin18-37 against A. baumannii under varying sodium ion concentrations, assessed by OD595. All data are expressed as mean ± SEM from three biological replicates.
The stability of Pecbloodin_18-37_ under thermal and ionic stress conditions was also evaluated. Thermal stability testing demonstrated that the peptide retained its antimicrobial activity against A. baumannii even after heat treatment at 100°C for up to 30 min (Fig. 3C), indicating strong heat resistance. Additionally, the effect of different sodium ion concentrations on its activity was examined. The results showed that Pecbloodin_18-37_ could not completely inhibit the growth of S. aureus at a sodium ion concentration of 80 mM (Fig. 3D).
Induction of endogenous ROS in bacteria by Pecbloodin18-37
AMPs are known to promote bacterial cell damage in part by inducing the accumulation of ROS (17). To assess whether Pecbloodin_18-37_ exerts similar effects, intracellular ROS levels were measured in A. baumannii and S. aureus following peptide treatment.
As shown in Fig. 4A, Pecbloodin_18-37_ significantly increased ROS production in both bacterial species in a dose-dependent manner. Higher peptide concentrations resulted in greater accumulation of endogenous ROS, suggesting that ROS generation may contribute to the bactericidal mechanism of Pecbloodin_18-37_.
Effects of Pecbloodin18-37 on ROS generation and microbial membrane integrity. (A) Intracellular ROS production in A. baumannii and S. aureus following Pecbloodin18-37 treatment, detected via DCFH-DA fluorescence (n = 5). Different lowercase letters denote statistically significant differences among groups (P < 0.05). (B) Scanning electron microscopy images of bacterial and fungal cells showing morphological changes after treatment with Pecbloodin18-37. (C) CLSM analysis of membrane permeability in A. baumannii and S. aureus stained with SYTO 9 and PI.
Morphological alterations in microorganisms induced by Pecbloodin18-37
Scanning electron microscopy (SEM) was employed to examine the morphological changes in bacterial (S. aureus, P. aeruginosa, and A. baumannii) and fungal (A. flavus, F. solani, and F. oxysporum) cells following treatment with Pecbloodin_18-37_.
As shown in Fig. 4B, treatment with Pecbloodin_18-37_ resulted in severe structural damage to both bacterial and fungal cells, including membrane disruption, surface collapse, and leakage of intracellular contents. In contrast, untreated cells displayed smooth, intact surfaces with no observable damage, indicating that Pecbloodin_18-37_ directly compromises microbial membrane integrity.
Pecbloodin18-37 increases bacterial inner membrane permeability
The effect of Pecbloodin_18-37_ on the inner membrane integrity of A. baumannii and S. aureus was evaluated using SYTO 9 and propidium iodide (PI) fluorescence staining. SYTO 9 stains all bacterial cells, while PI penetrates only those with compromised membranes, emitting red fluorescence.
As shown in Fig. 4C, untreated bacteria displayed uniform green fluorescence, indicating intact membranes. In contrast, Pecbloodin_18-37_-treated bacteria exhibited strong red fluorescence, comparable to that observed in the polymyxin B (PMB)-treated group, suggesting significant disruption of inner membrane integrity.
Inhibitory effect of Pecbloodin18-37 on biofilm formation
Biofilm formation is a common feature of bacterial infections and is a major contributor to antibiotic resistance (18), potentially increasing bacterial tolerance by 10- to 1,000-fold (19). To evaluate the anti-biofilm potential of Pecbloodin_18-37_, varying concentrations (6–48 μM) were tested against A. baumannii and S. aureus.
As shown in Fig. 5A and B, Pecbloodin_18-37_ significantly inhibited biofilm formation in both bacterial species in a dose-dependent manner, suggesting its potential utility in disrupting biofilm-associated resistance mechanisms.
Pecbloodin18-37 inhibits biofilm formation and does not induce bacterial resistance. (A and B) Quantification of biofilm formation by A. baumannii and S. aureus treated with varying concentrations of Pecbloodin18-37 using crystal violet staining and OD595 measurements (n = 5). Data are shown as mean ± SEM. Statistical significance was assessed using one-way ANOVA followed by the Dunnett test; different letters indicate significant differences. (C and D) Resistance induction assay over 48 days under sub-MIC exposure. MIC fold change is plotted on a log10 scale against time. LL-37 and antibiotics served as controls.
Pecbloodin18-37 exhibits antibacterial activity without inducing resistance
The rapid emergence of antibiotic resistance in clinical pathogens, particularly A. baumannii and S. aureus, poses a significant challenge to effective treatment (20). To assess the risk of resistance development, both bacterial species were subjected to 48 consecutive days of exposure to sub-MIC concentrations of two AMPs (Pecbloodin_18-37_ and LL-37) and four conventional antibiotics.
In A. baumannii, prolonged exposure to Pecbloodin_18-37_ and LL-37 did not result in any notable change in MIC values. In contrast, gentamicin resistance increased by nearly 1,000-fold, while tigecycline resistance increased by several hundred fold (Fig. 5C). Similarly, S. aureus showed no significant change in susceptibility to either AMP. However, the MIC for ampicillin increased by several hundred fold and that for vancomycin increased by less than 10-fold. Despite the relatively modest change, the increase in vancomycin resistance was statistically significant and may indicate a concerning trend (Fig. 5D). These findings underscore the low propensity of Pecbloodin_18-37_ to induce resistance compared to conventional antibiotics, highlighting its potential as a promising antimicrobial agent for long-term application.
Pecbloodin18-37 exhibits low cytotoxicity and hemolytic activity
The cytotoxicity of Pecbloodin_18-37_ was evaluated using two normal cell lines (HEK-293T and ZF4) and one cancer cell line (HeLa). As shown in Fig. 6A through C, Pecbloodin_18-37_ exhibited no significant cytotoxicity toward HEK-293T and HeLa cells at concentrations below 96 μM and no cytotoxic effects on ZF4 cells at concentrations below 48 μM. In contrast, Melittin (6 μM), used as a positive control, induced marked cytotoxicity in all three cell lines.
*In vitro cytotoxicity and hemolytic activity of Pecbloodin18-37. (A–C) Cell viability of HEK-293T, HeLa, and ZF4 cells after exposure to Pecbloodin18-37, measured by MTS assay. Melittin served as a cytotoxic control. (D) Hemolytic activity of Pecbloodin18-37 on mouse erythrocytes. Saline and 0.1% Triton X-100 were used as negative and positive controls, respectively. Data are presented as mean ± SEM. P < 0.05 indicates a statistically significant difference.
The hemolytic activity of Pecbloodin_18-37_ was assessed using freshly isolated mouse erythrocytes. Saline (0.9%) served as a negative control and caused no hemolysis, while 0.1% Triton X-100, used as a positive control, resulted in complete lysis of red blood cells. As shown in Fig. 6D, Pecbloodin_18-37_ caused no significant hemolysis at concentrations below 96 μM, indicating favorable hemocompatibility.
In vivo efficacy of Pecbloodin18-37 in an E. tarda-infected B. pectinirostris model
An in vivo infection model of B. pectinirostris was established to evaluate the antimicrobial, immunomodulatory, and antioxidant effects of Pecbloodin_18-37_. Fish were intraperitoneally injected with E. tarda, followed by administration of Pecbloodin_18-37_ 1 h post-infection.
At 48 h post-treatment, the survival rate in the Pecbloodin_18-37_-treated group was 67.5%, significantly higher than the 42.5% observed in the control group (P = 0.0422). The hazard ratio for the treatment group relative to the control was 0.4920 (95% CI: 0.2482–0.9753), indicating that Pecbloodin_18-37_ significantly reduced mortality risk (Fig. 7A).
*In vivo immunoprotective effects of Pecbloodin18-37 against E. tarda infection in B. pectinirostris. (A) Experimental scheme and Kaplan-Meier survival curves for mudskippers infected with E. tarda and treated with Pecbloodin18-37 (n = 40). (B) Liver bacterial load after treatment (n = 5). (C) Expression levels of immune-related genes (TNFα, IL-1β, IL-10, TLR4, and Hepcidin), ROS levels, and MPO activity in liver tissue (n = 5). Statistical comparisons were conducted using one-way ANOVA or t-tests as appropriate. Different letters or asterisks denote significant differences (P < 0.05).
Bacterial burden analysis showed a marked reduction in liver CFU counts in the treated group compared to the control (Fig. 7B). qPCR analysis of liver tissue revealed significant downregulation of pro-inflammatory cytokines (TNF-α and IL-1β) and upregulation of the anti-inflammatory cytokine IL-10, the antimicrobial peptide gene Hepcidin, and the pattern recognition receptor TLR4 in the Pecbloodin_18-37_-treated group. Moreover, Pecbloodin_18-37_ treatment significantly decreased hepatic ROS levels and MPO activity, indicating alleviation of infection-associated oxidative stress (Fig. 7C).
DISCUSSION
Intensive aquaculture practices have resulted in the widespread occurrence of various diseases, especially edwardsiellosis, which is notable for its high incidence rate (21). To combat bacterial infections in aquaculture, a significant quantity of antibiotics is used (22). Nevertheless, the rise of antibiotic resistance in aquaculture and its implications for public health and food safety have underscored the necessity for developing novel antibacterial drugs. The marine environment is a rich reservoir of bioactive molecules, where various immune-related substances play crucial roles in pathogen recognition and clearance (23–25). AMPs are regarded as a highly viable substitute for traditional antibiotics. They possess broad-spectrum antibacterial activity, fast bactericidal action, low potential for drug resistance, and are characterized by their safety, minimal toxicity, and lack of side effects (26). Mudskippers B. pectinirostris, which are amphibious fish inhabiting the tidal flats, have a series of unique physiological traits that enable them to thrive in the complex environment straddling land and sea (27). Owing to this intricate habitat, mudskippers are constantly exposed to various disease-causing microbes. Therefore, there is great potential to mine novel and effective antimicrobial peptides from this species. In our research, we discovered a novel and previously uncharacterized functional gene, Pecbloodin, from B. pectinirostris. Following bioinformatics examination and confirmation, a truncated peptide named Pecbloodin_18-37_ was identified from Pecbloodin. We investigated the physicochemical characteristics and antimicrobial activity of Pecbloodin_18-37_ to understand its antimicrobial efficacy against E. tarda infections and its underlying mechanism, offering new perspectives for developing antibiotic substitutes for future aquaculture applications.
Due to the immature adaptive immunity of fish, they primarily rely on their innate immune system to resist pathogenic microorganisms when faced with infection (28). The primary immune organs involved include the intestine, liver, spleen, and kidney (29). In this study, Pecbloodin was significantly upregulated in the blood, liver, spleen, and trunk kidney upon E. tarda challenge. These findings indicate that Pecbloodin may play a crucial role in the immune protection of mudskippers. Currently, it is commonly acknowledged that the net charge and hydrophobicity of AMPs are among the most significant factors influencing their antimicrobial activity (30). Pecbloodin_18-37_ possesses a substantial positive charge number of +7 and 40% hydrophobicity, similar to LL-37, and exhibits potent broad-spectrum antimicrobial efficacy against various bacterial and fungal pathogens. Similar to the binding properties of other cationic AMPs (31), the positive charge of Pecbloodin_18-37_ might enhance binding to negatively charged microbial surfaces through electrostatic interaction.
One of the bactericidal mechanisms of AMPs is the disruption of bacterial membrane structure, which typically relies on the peptide concentration at the membrane surface. Once a specific threshold is reached, the membrane structure is destroyed (32, 33). In this study, we examined the impact of Pecbloodin_18-37_ on the morphological structure of bacteria using SEM and verified its effect on bacterial membrane permeability through SYTO9 and PI staining. Pecbloodin_18-37_ was shown to interact with cell membranes, causing them to rupture and shrink on the surface, which leads to leakage of cellular contents and ultimately bacterial death. Furthermore, we found that the endogenous ROS in bacteria significantly rose after exposure to Pecbloodin_18-37_. This indicates that Pecbloodin_18-37_ simultaneously induces oxidative stress, and elevated levels of ROS may injure bacterial lipids, proteins, and DNA, leading to lethal bacterial damage (34). In conclusion, the disruption of membrane integrity and the surge in ROS levels may both contribute to bacterial killing.
Biofilms serve as a means for bacteria to withstand environmental stress and counteract the impact of drugs, including antibiotics (35). The development of bacterial biofilms generally includes three phases: (i) the planktonic stage, (ii) initial adherence to a surface, and (iii) the development of microcolonies and the release of extracellular polymeric substances (EPSs). When a thick EPS matrix is established, it restricts drug penetration, rendering it challenging to attain effective bactericidal concentrations within the biofilm, thereby conferring resistance (36). In this study, the bactericidal kinetics of Pecbloodin_18-37_ showed rapid killing of planktonic bacteria, thereby inhibiting further biofilm development. Moreover, at long-term sub-MIC concentrations, Pecbloodin_18-37_ did not induce resistance, which may be attributed to the multiple antimicrobial mechanisms of AMPs that limit bacterial resistance due to target mutations. This represents a significant edge of Pecbloodin_18-37_ over traditional antibiotics in preventing bacterial resistance.
Although AMPs have attracted widespread interest due to their myriad of advantages, the clinical application of AMPs is confronted with major obstacles, including poor stability, cytotoxicity, and production costs (10). Generally speaking, the activity of AMPs is notably influenced by elevated levels of sodium ions (37). The toxicity of certain cationic peptides also significantly limits their applications. For instance, Melittin, which possesses bactericidal properties, exhibits marked toxicity toward mammalian cells, although this property has also been harnessed for cancer studies (38). This study further evaluates the stability and safety of Pecbloodin_18-37_, with results indicating that high-temperature treatment has no significant impact on its activity. Moreover, the antibacterial activity of Pecbloodin_18-37_ remained largely unaffected when the sodium concentration was 40 mM, and although there was some impact at 80 mM sodium concentration, it can still inhibit bacterial growth within 7 h, demonstrating that Pecbloodin_18-37_ has a relatively high tolerance to sodium ions. In terms of safety assessment, while Pecbloodin_18-37_ exhibits cytotoxic effects on HEK-293T and HeLa cells at a concentration of 96 μM and on ZF4 cells at 48 μM, in comparison, the positive peptide control Melittin exhibits notable toxicity even at a low concentration of 6 μM. Additionally, the bactericidal concentrations of Pecbloodin_18-37_ for most bacteria or fungi are below 24 μM, rendering it relatively safe for practical applications. Subsequently, the hemolytic capacity of Pecbloodin_18-37_ in mouse red blood cells was further assessed, and no marked hemolysis was detected at a concentration of 48 μM. It is a well-established fact that cell membranes are enriched in neutral phospholipids like phosphatidylcholine and sphingomyelin, in contrast to bacterial membranes. The bulk of negatively charged phospholipids, such as phosphatidylserine, are predominantly situated in the inner layer of the bilayer (37). Thus, the selectivity of Pecbloodin_18-37_ might be significantly influenced by the variations in membrane composition and structure. In summary, Pecbloodin_18-37_ demonstrates excellent stability and biocompatibility, holding promising prospects for future applications.
The in vivo anti-infection efficacy evaluation of antimicrobial peptides is a crucial step toward their practical application. Prior research has demonstrated that AMPs have both direct antimicrobial properties and a substantial impact on modulating the immune response in vivo, which aids in controlling microbial infections in organisms (39). AMPs derived from the mudskipper, such as BpNKL (15), BpHep-2 (40), and BpLEAP-2 (13), have been shown to significantly boost the survival rate of mudskippers infected with E. tarda and to markedly lower the bacterial load in tissues. Additionally, BpLEAP-2 could also decrease the mRNA expression levels of pro-inflammatory factors in tissues. In this study, we found that Pecbloodin_18-37_ similarly enhanced the survival rate of mudskippers infected with E. tarda and effectively reduced the bacterial load in liver tissues. Meanwhile, Pecbloodin_18-37_ could significantly decrease the mRNA expression levels of pro-inflammatory factors TNF-α and IL-1β and increase the expression of IL-10, TLR4, and Hepcidin. As an anti-inflammatory agent, IL-10 can significantly diminish the synthesis of pro-inflammatory factors and holds a crucial position in the treatment of inflammatory and autoimmune diseases (41). The activation of TLR4 can bind to lipopolysaccharides in gram-negative bacteria and, together with the highly expressed AMPs such as Hepcidin, accelerate bacterial killing (42). In addition, Pecbloodin_18-37_ also reduces the levels of ROS and the enzyme activity of MPO in liver tissues. The decrease in MPO activity reflects the reduction in ROS levels, and reactive oxygen species are considered harmful to tissues through oxidative DNA damage to cells (43). In summary, Pecbloodin_18-37_ can not only directly kill bacteria in mudskippers but also inhibit E. tarda infection in mudskippers through immune regulation and antioxidant effects.
In this study, a novel functional gene, Pecbloodin, was identified from the mudskipper B. pectinirostris. Based on its physicochemical characteristics, a truncated antimicrobial peptide, Pecbloodin_18-37_, was screened and found to exhibit broad-spectrum antimicrobial activity and excellent thermal stability. Pecbloodin_18-37_ exerts its bactericidal effect by disrupting microbial membrane integrity, increasing membrane permeability, and inducing the accumulation of ROS, ultimately leading to cell death. In addition, Pecbloodin_18-37_ effectively inhibits biofilm formation and shows a low risk of inducing bacterial resistance. In vivo assays further demonstrated that Pecbloodin_18-37_ significantly improves the survival of E. tarda-infected mudskippers, reduces hepatic bacterial burden, and modulates immune and oxidative stress responses. Overall, Pecbloodin_18-37_ represents a promising antimicrobial agent with significant potential for therapeutic application in aquaculture, offering a viable alternative to conventional antibiotics.
MATERIALS AND METHODS
Animals, strains, and cell lines
Mudskippers (B. pectinirostris, 20 ± 5 g) were obtained from a local aquaculture facility in Xiapu, Fujian, China. Prior to experimentation, fish were acclimated for at least 7 days in a recirculating aquaculture system maintained at 25°C with 10‰ salinity. Fish were anesthetized using 200 mg/L ethyl 3-aminobenzoate methanesulfonate (MS-222; Sigma-Aldrich, USA) before injection or tissue sampling.
Standard microbial strains used in this study were acquired from the China General Microbiological Culture Collection Center (CGMCC), including E. coli (CGMCC 1.2389), P. aeruginosa (CGMCC 1.2421), A. baumannii (CGMCC 1.6769), E. tarda (CGMCC 1.1872), A. hydrophila (CGMCC 1.2017), V. alginolyticus (CGMCC 1.1833), L. monocytogenes (CGMCC 1.10753), S. epidermidis (CGMCC 1.4260), S. aureus (CGMCC 1.2465), C. glutamicum (CGMCC 1.1886), E. faecalis (CGMCC 1.2135), B. cereus (CGMCC 1.3760), C. neoformans (CGMCC 2.1563), C. albicans (CGMCC 2.2411), A. flavus (CGMCC 3.4410), F. oxysporum (CGMCC 3.6785), F. graminearum (CGMCC 3.349), and F. solani (CGMCC 3.5840). Clinically relevant multidrug-resistant strains, including MRSA QZ19130/QZ19134, MDR P. aeruginosa QZ19121/QZ19122, and MDR A. baumannii QZ18050/QZ18055, were kindly provided by the Second Affiliated Hospital of Fujian Medical University.
HEK-293T and HeLa cell lines were obtained from the Chinese Academy of Sciences Cell Bank and maintained in DMEM (Gibco, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, USA) at 37°C in a 5% CO_2_ incubator. The ZF4 cell line was purchased from the China Zebrafish Resource Center (Wuhan, China) and cultured in DMEM/F-12 (1:1) medium (Gibco, USA) with 10% FBS at 28°C under 5% CO_2_. Fresh mouse red blood cells were collected for hemolysis assays.
Cloning of the Pecbloodin gene
Specific primers were designed using Primer Premier 5.0 (Premier Biosoft) and synthesized by Sangon Biotech (Shanghai, China). Total RNA was extracted from various tissues using TRIzol Reagent (Invitrogen, UK) following the manufacturer’s protocol. RNA concentration and purity were evaluated with an Agilent 2100 Bioanalyzer (Agilent Technologies, USA), and integrity was confirmed by 1.0% agarose gel electrophoresis. Equal quantities of RNA from different tissues were pooled for downstream cDNA synthesis. First-strand cDNA was synthesized using the SMARTer RACE 5′/3′ Kit (Clontech, USA), and full-length cDNAs were amplified using LA Taq DNA polymerase (Takara, Japan), a high-fidelity enzyme optimized for long-fragment amplification (44). The resulting PCR products were ligated into the pMD18-T vector (Takara, Japan), transformed into E. coli, and subjected to bidirectional Sanger sequencing by Sangon Biotech Co., Ltd. (Shanghai, China).
Bioinformatic analysis, peptide design, and synthesis
Sequence alignment was conducted using DNAMAN 8.0. The 3D structure of Pecbloodin was predicted with AlphaFold (https://github.com/deepmind/alphafold) and visualized using PyMOL 2.5.8. Peptide chemical structures were drawn using ChemDraw (https://www.perkinelmer.com/category/chemdraw). Conserved domain analysis was performed via NCBI tools (NCBI; http://www.ncbi.nlm.nih.gov), and phylogenetic trees were constructed using MEGA 11.0. Sequence alignments were visualized with ESPript (https://espript.ibcp.fr/ESPript/ESPript/). Protein physicochemical properties were predicted with ProtParam (https://web.expasy.org/protparam/) and HeliQuest (https://heliquest.ipmc.cnrs.fr/cgi-bin/ComputParams.py).
The antimicrobial region of Pecbloodin was predicted using the CAMP_R4_ database (http://www.camp.bicnirrh.res.in/prediction.php). Based on this prediction and several well-established parameters for antimicrobial peptide design—including net positive charge, hydrophobicity, amphiphilicity, α-helical propensity, and an optimal sequence length—a 20-residue fragment, Pecbloodin_18-37_ (H-LYFLIRAVRLKKPAPKKKYG-OH), was chemically synthesized (97.8% purity) by GenScript (Nanjing, China) and stored at −20°C in lyophilized form.
Tissue distribution and expression analysis by qPCR
Tissues, including blood, spleen, trunk kidney, intestine, brain, head kidney, gills, skin, heart, liver, and muscle, were collected from healthy mudskippers. To assess immune-responsive expression, blood, liver, spleen, and trunk kidney were collected at 0, 6, 12, 24, 48, and 72 h post-infection with 1.0 × 10^4^ CFU/fish of E. tarda (13, 40), a sublethal dose that has been widely used to induce measurable innate immune responses without causing excessive mortality or severe tissue damage in mudskipper infection models.
Absolute qPCR was used to determine tissue distribution, while relative qPCR was used to assess temporal expression post-infection. Reactions were performed using a CFX384 real-time PCR system (Bio-Rad, USA) following previously established protocols (44). Relative expression was calculated using the 2^−ΔΔCt^ method (45).
Antimicrobial activity assay
Antibacterial activity was evaluated using a broth microdilution method (46). Bacterial suspensions (~10^6^ CFU/mL) were incubated with Pecbloodin_18-37_ (3–192 μM) in 96-well plates for 24 h at species-specific optimal temperatures. LL-37 (GL Biochem, Shanghai) was used as a positive control. MIC was defined as the lowest peptide concentration preventing visible growth, and MBC as the concentration causing ≤99.99% reduction in viable cells. All assays were performed in triplicate and repeated three times.
Time-kill kinetics assay
Time-dependent bactericidal activity of Pecbloodin_18-37_ was assessed against A. baumannii and S. aureus. Bacteria were prepared in Mueller-Hinton broth (MHB) at ~1 × 10^6^ CFU/mL and incubated with Pecbloodin_18-37_ at 1× and 2× MBC. At predetermined time points, aliquots were collected, serially diluted, and plated on nutrient agar (HKM, China). After incubation at 37°C for 18–24 h, colony counts were recorded. Bactericidal efficacy was expressed as percent CFU = (CFU at time point/initial CFU) × 100%. Each assay was performed in triplicate and repeated three times.
Thermal stability and sodium ion tolerance
To evaluate thermal stability, Pecbloodin_18-37_ was preheated at 100°C for 0, 20, or 30 min. The heated peptides were then incubated with A. baumannii (~1 × 10^6^ CFU/mL) in MHB at 37°C for 24 h. Bacterial growth was quantified by absorbance at 595 nm using a microplate reader.
For sodium ion tolerance, A. baumannii cultures were co-incubated with Pecbloodin_18-37_ (1× MIC) in MHB containing NaCl at final concentrations ranging from 20 to 160 mM. After 24 h at 37°C, absorbance at 595 nm was measured. All experiments were performed in triplicate and repeated at least three times.
Measurement of ROS
Intracellular ROS production was assessed using 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA; Jiancheng, China). Bacterial suspensions of A. baumannii and S. aureus (~1 × 10^8^ CFU/mL) were incubated with Pecbloodin_18-37_ (1.5–12 μM) or LL-37 (12 μM) for 30 min. After washing with phosphate-buffered saline (PBS), cells were treated with DCFH-DA (10 μM) for an additional 30 min. Fluorescence intensity was measured at 488/533 nm using a microplate reader. Each assay included five replicates and was repeated three times.
SEM analysis
The morphological effects of Pecbloodin_18-37_ were examined using SEM (Zeiss SUPRA 55, Germany) as previously described (47). Bacterial and fungal strains were suspended in 10 mM sodium phosphate buffer (NaPB) to final concentrations of ~1 × 10^8^ and ~1 × 10^7^ CFU/mL, respectively, and treated with Pecbloodin_18-37_ (1× MBC) for 20 min at room temperature. Cells were fixed in 2.5% glutaraldehyde overnight, dehydrated through an ethanol gradient, dried using a critical point dryer (EM CPD300, Leica), gold-coated, and visualized under SEM.
Membrane permeability assay
The effect of Pecbloodin_18-37_ on bacterial inner membrane integrity was evaluated using the LIVE/DEAD BacLight kit (Thermo Fisher, USA) as previously described (48). Log-phase cultures of A. baumannii and S. aureus were washed in 10 mM NaPB (pH 7.4) and adjusted to 1 × 10^7^ CFU/mL. Cells were incubated with Pecbloodin_18-37_ (1× MBC) at 37°C for 15 min. PMB (1 μg/mL) served as a positive control. Following staining with SYTO 9 and PI, samples were incubated in the dark for 15 min and analyzed using confocal laser scanning microscopy (Zeiss LSM780, Germany).
Biofilm inhibition assays
The anti-biofilm activity of Pecbloodin_18-37_ was assessed as previously described (49). Log-phase A. baumannii and S. aureus cultures (~1 × 10^6^ CFU/mL) were incubated in 96-well plates with varying concentrations of Pecbloodin_18-37_ (0–48 μM) at 37°C for 24 h. Formed biofilms were stained with 0.1% crystal violet (Sigma-Aldrich, Germany), and absorbance at 595 nm was measured. Each experiment included five replicates and was repeated three times.
Resistance development under long-term exposure
To assess the potential for resistance development, A. baumannii and S. aureus were serially passaged for 48 days in the presence of sub-MIC levels of Pecbloodin_18-37_, following a modified protocol (50). Conventional antibiotics (gentamicin, tigecycline, vancomycin, and ampicillin) and LL-37 were included for comparison. Each day, cultures were diluted 1:1,000 into fresh MHB containing the respective agent. MIC values were determined periodically. All experiments included three biological replicates and were repeated independently three times.
Cytotoxicity and hemolytic activity
The cytotoxicity of Pecbloodin_18-37_ toward HEK-293T, HeLa, and ZF4 cells was assessed using the MTS assay (49). Cells (~1 × 10^5^ cells/mL) were seeded into 96-well plates, incubated for 10 h, and then treated with Pecbloodin_18-37_ (6–96 μM) for 24 h. Melittin (6 μM) was used as a positive control. Each assay was performed in quintuplicate and repeated three times.
Hemolytic activity was assessed using freshly isolated mouse erythrocytes. Red blood cells were washed with 0.9% saline and resuspended to a 4% (vol/vol) cell suspension. Aliquots (100 μL) were incubated with 100 μL of Pecbloodin_18-37_ (various concentrations) at 37°C for 1 h. Following centrifugation (3,000 g for 4 min), supernatant absorbance was measured at 540 nm. Saline and 0.1% Triton X-100 served as negative and positive controls, respectively. Hemolysis (%) was calculated as:
In vivo protective efficacy in B. pectinirostris
The therapeutic efficacy of Pecbloodin_18-37_ was evaluated in B. pectinirostris following intraperitoneal challenge with E. tarda. Fish (n = 40 per group) were injected with 1.3 × 10^7^ CFU/fish of E. tarda (a dosage corresponding to the LD_50_). One hour post-infection, treatment groups received Pecbloodin_18-37_ (20 μg/fish, an optimal dose determined by preliminary efficacy and safety trials), while controls received PBS. Mortality was recorded for 48 h, and survival curves were plotted using GraphPad Prism 9.0.
At 48 h, livers were harvested, homogenized, serially diluted, plated on nutrient agar, and incubated at 28°C for CFU quantification. Relative qPCR was conducted to measure the expression of immune-related genes (TNF-α, IL-1β, IL-10, and TLR4) and AMP genes (Hepcidin, LEAP-2, and lysozyme), using RPL8 as the reference gene. ROS and MPO activity in liver tissue were quantified using commercial kits from Nanjing Jiancheng Bioengineering Institute (China) and Solarbio (China), respectively. These quantitative data were standardized by referencing the uninfected PBS control group. For each parameter, experimental group values were divided by the uninfected control group’s average and presented as fold changes. This uniform approach across all data sets enabled a consistent assessment of infection effects and peptide modulation relative to baseline.
Statistical analysis
All statistical analyses were conducted using SPSS (version 26.0) and GraphPad Prism (version 9.0). Data are expressed as mean ± standard deviation. Comparisons between two groups were performed using Student’s t-test. For multiple group comparisons, one-way ANOVA followed by Dunnett’s or Tukey’s post hoc test was applied as appropriate.
In two-group comparisons, statistical significance was indicated as follows: *P < 0.05 and **P < 0.01. For multiple comparisons, different lowercase letters (e.g., a, b, c, and d) were used to denote statistically significant differences between groups; groups sharing the same letter were not significantly different (P > 0.05).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sasikumar R, Saranya S, Lourdu Lincy L, Thamanna L, Chellapandi P. 2024. Genomic insights into fish pathogenic bacteria: a systems biology perspective for sustainable aquaculture. Fish & Shellfish Immunology 154:109978. doi:10.1016/j.fsi.2024.10997839442738 · doi ↗ · pubmed ↗
- 2Buján N, Toranzo AE, Magariños B. 2018. Edwardsiella piscicida: a significant bacterial pathogen of cultured fish. Dis Aquat Org 131:59–71. doi:10.3354/dao 0328130324915 · doi ↗ · pubmed ↗
- 3Xu T, Zhang X-H. 2014. Edwardsiella tarda: an intriguing problem in aquaculture. Aquaculture 431:129–135. doi:10.1016/j.aquaculture.2013.12.001 · doi ↗
- 4Du H-F, Zhang Y-H, Zhang M, Liu Q-A, Zhu H-J, Cao F. 2022. Marine fungal metabolites as a source of drug leads against aquatic pathogens. Appl Microbiol Biotechnol 106:3337–3350. doi:10.1007/s 00253-022-11939-635486178 · doi ↗ · pubmed ↗
- 5Park SB, Aoki T, Jung TS. 2012. Pathogenesis of and strategies for preventing Edwardsiella tarda infection in fish. Vet Res 43:1–11. doi:10.1186/1297-9716-43-6723035843 PMC 3479428 · doi ↗ · pubmed ↗
- 6Mohanty BR, Sahoo PK. 2007. Edwardsiellosis in fish: a brief review. J Biosci 32:1331–1344. doi:10.1007/s 12038-007-0143-818202458 · doi ↗ · pubmed ↗
- 7Algammal AM, Mabrok M, Ezzat M, Alfifi KJ, Esawy AM, Elmasry N, El-Tarabili RM. 2022. Prevalence, antimicrobial resistance (AMR) pattern, virulence determinant and AMR genes of emerging multi-drug resistant Edwardsiella tarda in Nile tilapia and African catfish. Aquaculture 548:737643. doi:10.1016/j.aquaculture.2021.737643 · doi ↗
- 8Baindara P, Kumari S, Dinata R, Mandal SM. 2025. Antimicrobial peptides: evolving soldiers in the battle against drug-resistant superbugs. Mol Biol Rep 52:432. doi:10.1007/s 11033-025-10533-z 40293554 · doi ↗ · pubmed ↗