Two ferric uptake regulator proteins in Ralstonia pseudosolanacearum strain OE1-1 function cooperatively in response to the extracellular iron level under ferrous iron-rich conditions
Sora Tateda, Yuki Terazawa, Akinori Kiba, Kouhei Ohnishi, Yasufumi Hikichi, Masayuki Tsuzuki

TL;DR
This study shows that two iron-regulating proteins in a plant pathogen work together to control iron uptake and disease-causing ability in tomato plants.
Contribution
The study reveals cooperative function of two Fur proteins in regulating iron response and virulence in Ralstonia pseudosolanacearum.
Findings
Fur1 and Fur2 repress siderophore-related genes under Fe2+-rich conditions.
The double deletion of Fur1 and Fur2 significantly reduces bacterial virulence in tomato plants.
Fur1 and Fur2 control extracellular Fe3+-chelating activity and growth under iron-rich conditions.
Abstract
Soil-borne Gram-negative bacteria in the Ralstonia solanacearum species complex (RSSC) cause bacterial wilt symptoms in diverse crop plants. Although iron is an important metal ion for most organisms, the regulatory effect of iron on soil-borne RSSC gene expression remains unknown. Thus, we identified two ferric uptake regulator (Fur) proteins, Fur1 and Fur2, in R. pseudosolanacearum strain OE1-1. All four RSSC phylotype strains had two Fur homologs, a conserved Fur (Fur1) and an alternative Fur (Fur2). We also analyzed the transcriptomes of a fur1 deletion mutant, fur2 deletion mutant, and fur1⁄fur2 double deletion mutant, revealing that Fur1 and Fur2 cooperatively repress siderophore-related gene expression under Fe2+-rich conditions. Furthermore, extracellular Fe3+-chelating activity was cooperatively controlled by Fur1 and Fur2 under Fe2+-rich conditions. Additionally, nitrate…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
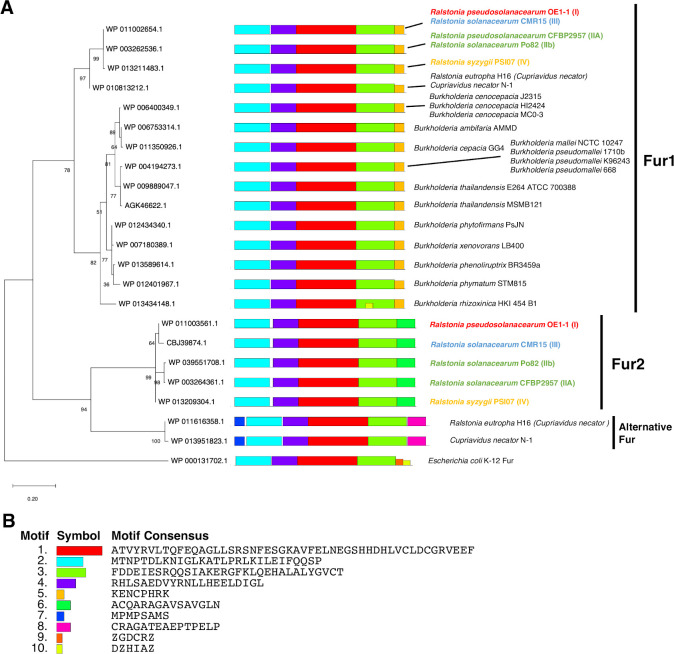 Fig 1
Fig 1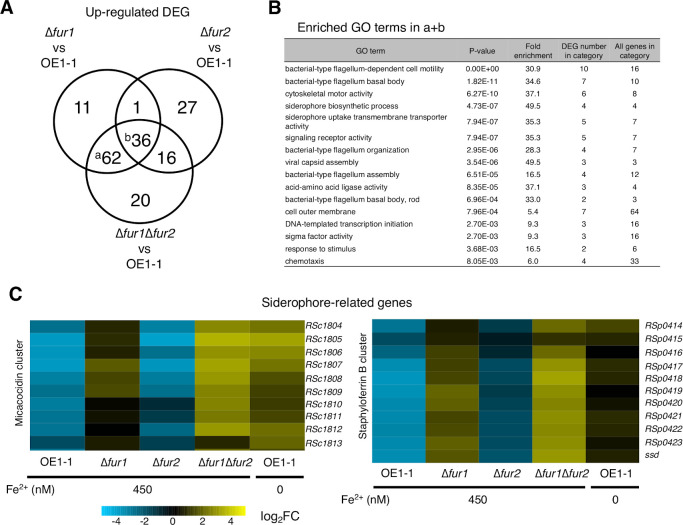 Fig 2
Fig 2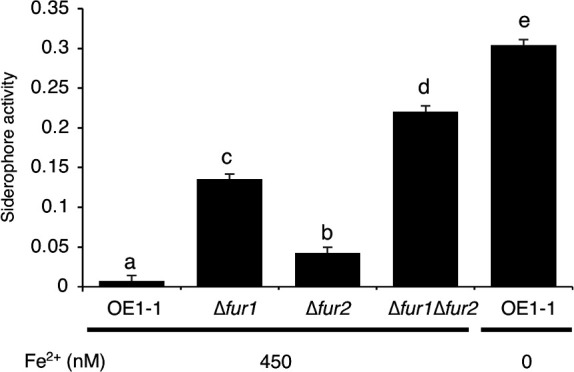 Fig 3
Fig 3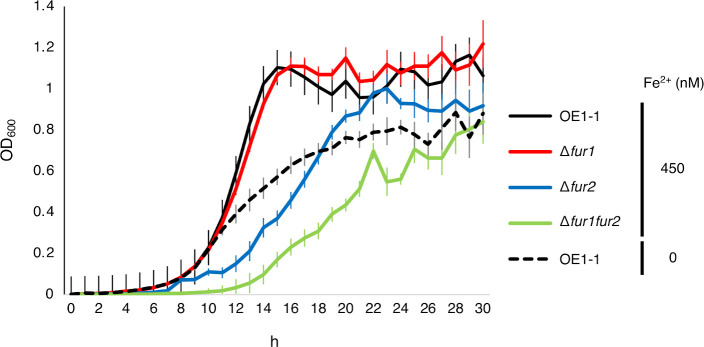 Fig 4
Fig 4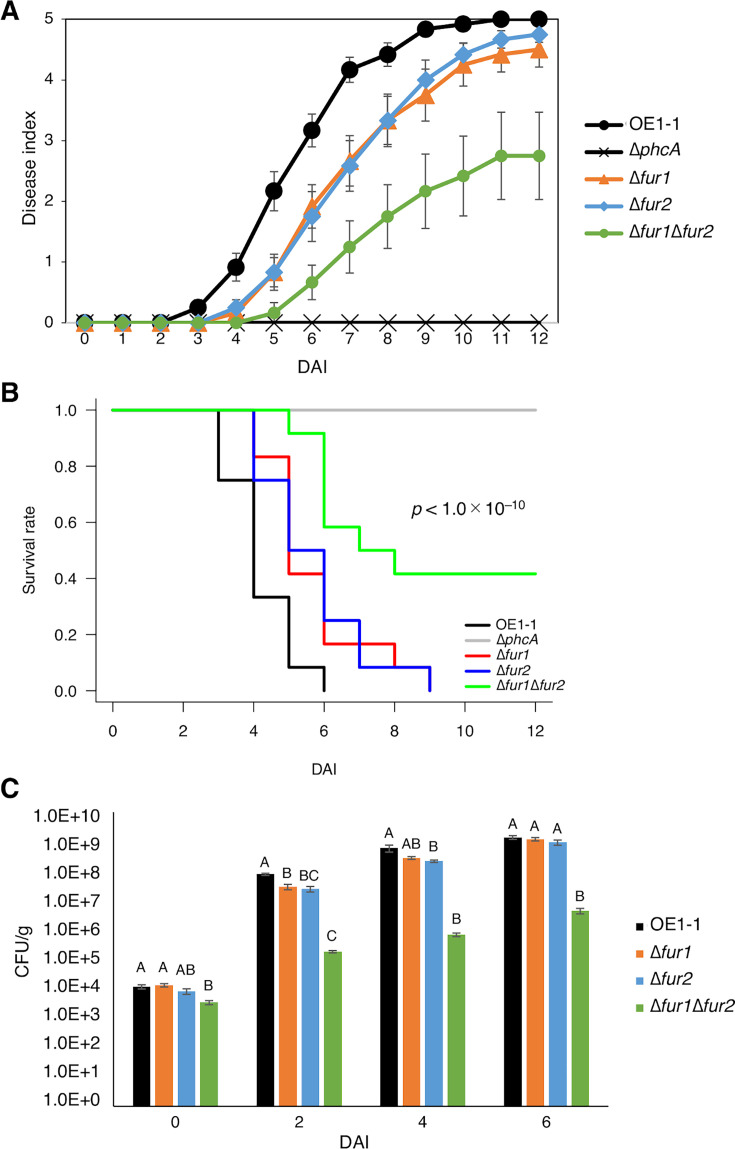 Fig 5
Fig 5- —Canon Foundation for Scientific Researchhttp://dx.doi.org/10.13039/100009851
- —Japan Society for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100001691
- —Japan Society for the Promotion of Sciencehttp://dx.doi.org/10.13039/501100001691
- —Japan Science Societyhttp://dx.doi.org/10.13039/501100007807
- —Japan Science and Technology Agencyhttp://dx.doi.org/10.13039/501100002241
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogenic Bacteria Studies · Infections and bacterial resistance · Plant-Microbe Interactions and Immunity
INTRODUCTION
The soil-borne, Gram-negative β-proteobacterium Ralstonia solanacearum species complex (RSSC) causes bacterial wilt in a wide range of host plant species, resulting in devastating crop losses worldwide (1). RSSC in soil infects host plants through the roots and then moves to vascular tissues, wherein bacteria proliferate extensively (2–5). RSSC reportedly responds to the substantial environmental changes occurring during the transition from nutrient-limited soil to nutrient-rich host plants (6). Elucidating how RSSC adapts and thrives under various environmental conditions has important implications for pathogen control.
Iron uptake is an important process in various organisms, including bacteria, because some enzymes require iron as a co-factor during redox reactions as well as proteins that bind to iron via metal-binding domains (7). Acquisition of abundant iron from hosts can be a reason why some bacteria infect other organisms (7). On the other hand, excessive amounts of intracellular iron may be harmful to bacteria because free Fe^2+^ leads to the production of hydroxyl radicals (8). Thus, bacterial cells must be able to sense extracellular iron levels and control intracellular iron levels. Iron has two different redox states with different characteristics: ferrous iron (Fe^2+^) and ferric iron (Fe^3+^). It has been estimated that iron redox state in the cytoplasm favors Fe^2+^ because of the solubility in the cytoplasm, and Fe^2+^ is generally the form incorporated into iron-dependent enzymes (9, 10). However, Fe^3+^, which is relatively unreactive and insoluble, is abundant under natural conditions. Also, in hosts, abundant iron is likely to be pooled in various ways as the labile iron pool, which requires bacteria to have specific mechanisms to obtain iron from hosts (7, 10). To take up environmental and/or host-derived Fe^3+^, bacterial cells produce siderophores, which are Fe^3+^-chelating secondary metabolites (11). In many bacterial species, siderophore production is controlled by a conserved transcriptional regulator family, ferric uptake regulator (Fur), whose members contain a DNA-binding domain and metal-binding motifs (12). The binding of Fe^2+^ to the metal-binding motif of Fur proteins alters the DNA-binding domain, thereby enabling Fur to bind to specific DNA motifs in the presence of Fe^2+^. In some bacterial species, the expression of siderophore-related genes is regulated by Fur proteins.
RSSC strains reportedly produce two siderophores: micacocidin (13) and staphyloferrin B (14). Micacocidin has a yersiniabactin-like structure and is biosynthesized by enzymes encoded at the mic locus in the RSSC genome (13). Staphyloferrin B is a polycarboxylate siderophore produced by enzymes encoded in the gene cluster that includes ssd, whose expression level increases under iron-deficient conditions (14). However, whether Fur proteins regulate siderophore production in RSSC remains unknown.
To elucidate the mechanism regulating RSSC gene expression in response to extracellular iron levels, we functionally characterized Fur proteins in R. pseudosolanacearum strain OE1-1. We determined that RSSC has an alternative Fur clade protein (Fur2) in addition to a conserved Fur (Fur1). A transcriptome analysis was performed to analyze the effects of Fur1 and Fur2 on gene expression. Moreover, we examined the effects of Fur1 and Fur2 on the Fe^3+^-chelating activity and growth of R. pseudosolanacearum strain OE1-1, while also investigating whether Fur1 and Fur2 contribute to the virulence of strain OE1-1.
RESULTS
The RSSC genome contains two genes encoding a conserved Fur and an additional non-conserved Fur exclusive to RSSC
To identify Fur proteins in RSSC, we first searched the R. pseudosolanacearum strain OE1-1 genome and identified two genes, fur1 (RSc2747) and fur2 (RSp0247), which encode Fur1 and Fur2, respectively. We obtained two putative Fur protein sequences from representative strains of four RSSC phylotypes. These sequences were aligned to compare domain structures. Fur1 and Fur2 in RSSC retained the N-terminal DNA-binding domain and two metal-binding sites conserved in the Fur proteins of the distantly related species Escherichia coli and Helicobacter pylori (15, 16), suggesting that Fur1 and Fur2 are Fur family members that bind to DNA and metal ions (Fig. S1).
Considering RSSC belongs to the family Burkholderiaceae (β-proteobacteria) (17), we obtained 22 putative Fur sequences from non-RSSC Burkholderiaceae species and one from E. coli as an outgroup for a phylogenetic analysis. In the resulting phylogenetic tree, Fur1 and Fur2 were clustered in different groups (Fig. 1A). Only four RSSC phylotypes and two strains of the closest species (Cupriavidus necator, which is also known as Ralstonia eutropha) had proteins in the additional clade containing R. pseudosolanacearum Fur2. By contrast, the other Burkholderiaceae species only had proteins belonging to the clade containing R. pseudosolanacearum Fur1 (Fig. 1A).
Ralstonia solanacearum species complex has two ferric uptake regulator homologous proteins in different clades. (A) A phylogenetic tree and putative amino acid motifs predicted by a MEME analysis revealed relationships between putative ferric uptake regulator (Fur) family proteins in Ralstonia solanacearum species complex (RSSC) and other closely related non-RSSC Burkholderiaceae species. The numbers written at each branch are those of the percentage of the result of a total of 1,000 times bootstrapping, supporting the branch. Protein IDs are provided along with strain name(s). Some protein IDs are shared by multiple species because the amino acid sequences were identical. For RSSC species, phylotype numbers are provided and distinguished by color: red: phylotype I, green: phylotypes IIA and IIB; blue: phylotype III; and yellow: phylotype IV. The Fur amino acid sequence in Escherichia coli strain K-12 (WP_000131702.1) was used as an outgroup. Different conserved motifs predicted by a MEME analysis are distinguished by differentially colored boxes. The sequences having the same amino acid motif are grouped as Fur1 or Fur2. (B) Motif IDs and motif consensus amino acid sequences shown in panel A, as provided by a MEME analysis, are displayed.
A MEME analysis conducted to compare domain structures identified 10 conserved amino acid motifs and revealed that Fur1 in RSSC and Fur in Burkholderia species have the same motif structures, suggesting that RSSC Fur1 and other Burkholderia Fur proteins have conserved functions, which is grouped as Fur1 (Fig. 1A and B). Fur2 in RSSC and two Fur proteins in C. necator (WP_01161616358.1 and WP_0135951823.1) differed in terms of their C-terminal motif(s) and additional N-terminal motif #7 in C. necator Fur proteins, suggesting that they are grouped into different clades: Fur2 and alternative Fur (Fig. 1A and B). The protein structures of Fur1 and Fur2 were predicted by AlphaFold 3, and it was suggested that C-terminal amino acids in Fur1 do not form a static structure, whereas those in Fur2 form α-helix (Fig. S2). The phylogenetic analysis and the MEME analysis combining AlphaFold analysis suggested that Fur1 and Fur2 are tentatively in a paralogous relationship with different functional characters, whose ancestor-encoding genes underwent the duplication before the divergence of RSSC phylotypes. These results suggest that the presence of an alternative Fur2 clade protein in addition to the conserved Fur1 protein is a feature of RSSC species (Fig. 1A and B).
Fur1 and Fur2 control siderophore-related gene expression under Fe2+-rich conditions
Fur proteins have a DNA-binding motif and regulate gene expression in several bacterial species (12). We investigated the potential regulatory effects of Fur1 and Fur2 on gene expression and analyzed the strain OE1-1 transcriptome by completing an RNA sequencing (RNA-seq) analysis. We also generated fur1 deletion, fur2 deletion, and fur1⁄fur2 double deletion mutants (∆fur1, ∆fur2, and ∆fur1Δfur2, respectively) in the R. pseudosolanacearum strain OE1-1 genetic background. For the RNA-seq analysis, total RNA was extracted from strain OE1-1, ∆fur1, ∆fur2, and ∆fur1Δfur2 grown in modified quarter-strength M63 medium supplemented with 450 nM Fe^2+^, as well as from strain OE1-1 grown in medium lacking Fe^2+^ as a control (i.e., Fe^2+^ deficiency). The RNA was extracted from the strains when OD_600_ reached 0.3 during the exponential phase. The Fe^2+^ concentration was selected because quarter-strength M63 medium is generally used as a minimal medium and is likely to supply the sufficient amount of usable iron to R. pseudosolanacearum strain OE1-1 (18). Also, it has been reported that R. solanacearum can survive in water without iron (19). Read count data for each gene were normalized, and relative expression levels were calculated. Genes with significant differences in expression (|log_2_(fold-change)| > 2, q-value < 0.05) relative to the corresponding expression in strain OE1-1 under Fe^2+^-rich conditions (450 nM Fe^2+^) were designated as differentially expressed genes (DEGs). A total of 116, 79, and 138 upregulated DEGs as well as 335, 53, and 398 downregulated DEGs were detected in ∆fur1, ∆fur2, and ∆fur1Δfur2, respectively (Table S1). The distribution of DEGs common to both conditions was analyzed to identify genes controlled by Fur1 and/or Fur2. A Venn diagram analysis showed that many upregulated (98 genes) and downregulated (300 genes) DEGs were shared by ∆fur1 and ∆fur1Δfur2, suggesting that these genes are controlled by Fur1, but are also partially controlled by Fur2 (Fig. 2A).
Fur1 and Fur2 cooperatively repress siderophore production-related gene expression in the presence of 450 nM Fe2+. (A) Venn diagram of the number of differentially expressed genes (DEGs) with a log2[∆fur1/OE1-1 (450 nM Fe2+)] expression level > 2, log2[∆fur2/OE1-1 (450 nM Fe2+)] expression level > 2, and/or log2[∆fur1Δfur2/OE1-1 (450 nM Fe2+)] expression level > 2, with FDR < 0.05. Lowercase letters refer to the gene sets in panel B. (B) Enriched Gene Ontology (GO) terms among the upregulated genes indicated by lowercase letters in panel A (P < 10−3). (C) Relative expression levels of genes in two siderophore production-related gene clusters are presented in heatmaps.
We performed a Gene Ontology (GO) enrichment analysis to functionally annotate DEGs. The upregulated DEGs shared by ∆fur1 and ∆fur1Δfur2 (a and b in Fig. 2A), which are likely controlled by Fur1, were annotated with the GO term “siderophore biosynthetic process” (4/4 genes) (Fig. 2B). The regulatory effects of Fur proteins on siderophore-related genes have been reported for other Gram-negative bacterial species (12), implying that Fur1 is a conserved Fur protein among Gram-negative bacteria. The R. pseudosolanacearum genome reportedly includes two gene clusters associated with the production of micacocidin (RSc1804–RSc1813) and staphyloferrin B (RSp0414–RSp0424) (13, 14). We analyzed transcriptome data to compare the relative expression levels of these two gene clusters. Under conditions with iron present, the expression levels of both gene clusters increased in ∆fur1 and ∆fur1Δfur2, with greater increases in the double mutant, similar to the corresponding expression in strain OE1-1 under Fe^2+^-deficient conditions. Thus, Fur1 and Fur2 may cooperatively repress the expression of these two siderophore-related gene clusters under Fe^2+^-rich conditions (Fig. 2C). The expression level of some staphyloferrin B cluster genes increased in ∆fur2, but that of micacocidin did not, suggesting that Fur2 alone has a repressive effect on staphyloferrin B cluster (Fig. 2C). Under Fe^2+^-rich conditions, Fur1 alone can repress the expression of siderophore-related genes, while Fur2 has repressive effects on these genes in the absence of Fur1 only for micacocidin cluster (Fig. 2B and C). Note that ∆fur1 and ∆fur1Δfur2 showed higher expression of staphyloferrin B cluster genes than strain OE1-1 in Fe^2+^ deficiency, suggesting that staphyloferrin B cluster regulation includes both transcription activation and repression: Fur1/2-pathway represses the expression of staphyloferrin B cluster, while other pathway(s) activate it, leading to the balanced expression level (Fig. 2C).
The upregulated DEGs shared by ∆fur1 and ∆fur1Δfur2 also included motility-related genes having the GO terms like “bacterial-type flagellum-dependent cell motility,” “bacterial-type flagellum basal body,” “bacterial-type flagellum organization,” “bacterial-type flagellum assembly,” “bacterial-type flagellum basal body, rod,” and “chemotaxis” (Fig. 2B). The terms related to flagella are also enriched in the DEGs only in ∆fur2 (b in Fig. S3A and C), ∆fur2 and ∆fur1Δfur2 (d), and ∆fur1, ∆fur2, and ∆fur1Δfur2 (f), suggesting that Fur1 and Fur2 contribute to the regulation of flagellum-related genes in cooperative manners (Fig. S3A and C). It was also found that virulence-related genes, such as eps operon and endo-1,4-β-glucanase-encoding egl, were downregulated significantly in ∆fur1 and ∆fur1Δfur2 and moderately in strain OE1-1 under iron-deficient conditions (Table S1). This result indicates that the expression of virulence-related genes is induced by Fur1 under the presence of Fe^2+^. Overall, Fur1 and Fur2 also have pleiotropic regulatory effects on various genes other than the siderophore-related genes in strain OE1-1.
Fur1 is the main regulator of extracellular Fe3+-chelating activity under Fe2+-rich conditions, but Fur2 is also involved
In certain bacterial species, Fur proteins control extracellular Fe^3+^-chelating activity, which is repressed in the presence of sufficient iron and de-repressed in response to iron deficiency. To determine whether Fur1 and Fur2 control extracellular Fe^3+^-chelating activity, an in vitro extracellular Fe^3+^-chelating activity assay (18, 20) was conducted using ∆fur1, ∆fur2, and ∆fur1Δfur2 as well as strain OE1-1. The extracellular Fe^3+^-chelating activity of strain OE1-1 under Fe^2+^-rich conditions was lower than that in the absence of Fe^2+^ (Fig. 3). The extracellular Fe^3+^-chelating activity was higher for ∆fur1 and ∆fur2 than for strain OE1-1 under Fe^2+^-rich conditions, with higher activity detected for ∆fur1 than for ∆fur2 (Fig. 3). Notably, the extracellular Fe^3+^-chelating activity was higher for ∆fur1Δfur2 than for ∆fur1 and ∆fur2 under Fe^2+^-rich conditions, suggesting that Fur1 and Fur2 redundantly repress the extracellular Fe^3+^-chelating activity under Fe^2+^-rich conditions (Fig. 3). However, Fur1 seems to have a stronger repressive effect than Fur2.
Low extracellular Fe3+-chelating activity in the presence of 450 nM Fe2+ is mainly maintained by Fur1, with assistance from Fur2. Mean Fe3+-chelating activity levels [(A630 Control – A630 Sample)/A630 Control] of different samples are presented. Fe2+ concentrations in the growth medium are indicated at the bottom. Strain OE1-1 grown under 0 nM Fe2+ conditions was used as a positive control. Eight biological replicates were used (standard deviations are presented as error bars). Mean extracellular Fe3+-chelating activity levels were analyzed for significant differences between R. pseudosolanacearum strains via analysis of variance followed by Tukey–Kramer’s honestly significant difference test. Significant differences are indicated by different lowercase letters (P < 0.01).
Fur1 and Fur2 redundantly affect the expression of various metabolism-related genes
Our transcriptome analysis also revealed the redundant regulatory effects of Fur1 and Fur2 on gene expression. A total of 20 and 102 genes were identified as upregulated and downregulated DEGs, respectively, only in ∆fur1Δfur2 under Fe^2+^-rich conditions (Fig. 2A). Accordingly, Fur1 and Fur2 may redundantly regulate the expression of these genes under Fe^2+^-rich conditions (Fig. 2A; Fig. S3A and B). Enriched GO terms among DEGs specific to ∆fur1Δfur2 were related to nitrate metabolism. More specifically, “nitrate reductase activity” (4/5 genes) and “nitrate assimilation” (4/8 genes) were GO terms assigned to the upregulated DEGs exclusive to ∆fur1Δfur2. Hence, Fur1 and Fur2 redundantly repress the expression of these nitrate metabolism-related genes (Fig. S3C). The GO terms “integral component of membrane” (42/1,164 genes) and “oxidoreductase activity” (7/112 genes) were enriched among the downregulated DEGs; however, the specific functions of these genes in terms of bacterial metabolism were not obvious because the enrichment was not strong (Fig. S3D).
Deleting fur2 decreased growth, but deleting both fur1 and fur2 resulted in a much greater decrease
According to the transcriptome analysis, Fur1 and Fur2 affect the expression of various metabolism-related genes and siderophore production-related genes. We also analyzed the effects of Fur1 and Fur2 on the growth levels in response to different extracellular Fe^2+^ levels. Bacterial growth (per hour) was measured based on the optical density at 600 nm (OD_600_) values of cultures of strain OE1-1, ∆fur1, ∆fur2, and ∆fur1Δfur2. The bacterial growth of ∆fur2 was lower than that of strain OE1-1, whereas ∆fur1 had a similar growth level to that of strain OE1-1 (Fig. 4). Notably, bacterial growth was much lower for ∆fur1Δfur2 than for ∆fur2, suggesting that Fur1 and Fur2 redundantly affect the growth of strain OE1-1 (Fig. 4). The early growth level of strain OE1-1 in medium without Fe^2+^ was the same as that of strain OE1-1 in medium with 450 nM Fe^2+^, but after that, the growth slowed down as a result of iron deficiency (Fig. 4). The observed bacterial growth indicated that Fur2 may be needed to maintain an appropriate strain OE1-1 growth level, but Fur1 has redundant effects.
Deleting fur2 alone and deleting both fur1 and fur2 decreased the in vitro growth level. Strain OE1-1, ∆fur1, and ∆fur2 growth levels at 30°C under aerobic conditions were monitored hourly. Seven biological replicates per strain were grown under the indicated Fe2+ conditions. Averages and standard error bars are presented.
Fur1 and Fur2 contribute to the full virulence of OE1-1 in tomato plants
Fur family members are important for the virulence of many bacterial pathogens (12). To clarify the effects of Fur1 and Fur2 on virulence, we conducted virulence assays involving tomato plants, with a previously reported non-pathogenic strain (∆phcA) serving as a negative control (20–22). The disease index was markedly lower for plants inoculated with either ∆fur1 or ∆fur2 than for plants inoculated with strain OE1-1 at 4–7 days after inoculation (DAI), reflecting the effects of Fur1 and Fur2 on the virulence of OE1-1 (Fig. 5A). Notably, the disease index was lower for plants inoculated with ∆fur1Δfur2 than for plants inoculated with ∆fur1 and/or ∆fur2 at 6 DAI (Fig. 5A). Furthermore, the survival rate of plants inoculated with ∆fur1 and ∆fur2 decreased more slowly than that of plants inoculated with strain OE1-1, while that of plants inoculated with ∆fur1Δfur2 decreased much more slowly, with five of 12 ∆fur1Δfur2-inoculated plants surviving until 12 DAI (Fig. 5B).
Fur1 and Fur2 cooperatively maintain the full virulence of strains in tomato plants. (A) Virulence of R. pseudosolanacearum strains. Inoculated plants were examined using the following disease index scale: 0, no wilting; 1, 1%–25% wilting; 2, 26%–50% wilting; 3, 51%–75% wilting; 4, 76%–99% wilting; and 5, dead. For each bacterial strain, six replicate plants were analyzed twice. Data were combined and presented as the mean ± SE of 12 replicates. (B) Survival rates of plants inoculated with R. pseudosolanacearum strains, with the Chi-squared test conducted to determine P-values. (C) R. solanacearum strain populations in the roots of tomato plants inoculated using a root-dipping method were analyzed using Hara–Ono medium. CFU, colony-forming unit. SE is indicated by an error bar. Mean abundances were analyzed for significant differences between R. pseudosolanacearum strains via an analysis of variance followed by Tukey–Kramer’s honestly significant difference test. Significant differences are indicated by different lowercase letters (P < 0.05).
We also analyzed R. pseudosolanacearum strain populations in tomato plant roots. At 2 DAI, ∆fur1 and ∆fur2 were significantly less abundant than strain OE1-1 in tomato roots (P < 0.05) (Fig. 5C). Additionally, ∆fur1Δfur2 was considerably less abundant than ∆fur1 and ∆fur2 at 6 DAI (P < 0.05). Note that ∆fur1Δfur2 showed a significant decrease in the population even at 0 DAI, which is possibly because of the lower attachment rate in tomato plant roots. Accordingly, Fur1 and Fur2 cooperatively maintain the bacterial population (Fig. 5C). The observed decrease in the disease index and the slow decrease in the survival rate of plants inoculated with ∆fur1 and ∆fur2 imply that both Fur1 and Fur2 contribute to the virulence of strain OE1-1. Furthermore, based on the results for ∆fur1Δfur2, Fur1 and Fur2 likely cooperatively influence the virulence of strain OE1-1.
DISCUSSION
Considering that RSSC exists under both iron-limited conditions in rhizosphere soil and iron-rich conditions within host plants, the mechanism through which bacterial cells perceive and respond to extracellular iron conditions must be thoroughly characterized (2, 3, 23). In the current study, we revealed that RSSC has two Fur proteins (Fur1 and Fur2). Furthermore, a transcriptome analysis indicated that Fur1 and Fur2 cooperatively control siderophore-related gene expression (Fig. 2). Deleting fur1 alone leads to increased siderophore-related gene expression, which is repressed under Fe^2+^-rich conditions, but deleting both fur1 and fur2 results in even higher siderophore-related gene expression (Fig. 2C). Thus, Fur1 may be a major repressor of siderophore-related gene expression, with Fur2 providing secondary repressive effects. The rank order of extracellular Fe^3+^-chelating activity among R. pseudosolanacearum strains under Fe^2+^-rich conditions was as follows: OE1-1 < ∆fur2 < ∆fur1 < ∆fur1Δfur2 (Fig. 3). Therefore, Fur1 and Fur2 may cooperatively maintain low extracellular Fe^3+^-chelating activities under Fe^2+^-rich conditions by repressing siderophore-related gene expression in RSSC. In addition, deleting both fur1 and fur2 decreased the expression of nitrate metabolism-related genes, which was in contrast to the lack of significant changes following the deletion of *fur1 *or fur2 (i.e., single deletions). These results suggest that Fur1 and Fur2 redundantly control the expression of nitrate metabolism-related genes (Fig. S3A and C). R. solanacearum exploits nitrogen metabolism for respiration in addition to oxygen metabolism under the low-oxygen conditions typically found in xylem vessels (24). The deletion of denitrification-related genes alters iron-related gene expression, indicating that denitrification influences iron metabolism (25). Thus, nitrogen metabolism and iron-dependent gene regulation are tightly linked.
The transcriptome analysis in this study also revealed that Fur1 and Fur2 have pleiotropic regulatory effects on various genes. It was revealed that Fur1 and Fur2 cooperatively contribute to the repression of motility-related genes involved in flagellum assembly or chemotaxis (Fig. 2B; Fig. S3A and C). The transcriptome data showed that strain OE1-1 under the iron-deficient condition did not show the increased expression of flagellum components such as flg genes, suggesting that there are also induction pathways dependent on the presence of Fe^2+^, in addition to repression pathways by Fur1 and Fur2 (Table S1). The transcriptome analysis also revealed that some virulence genes, including eps operon and egl, are upregulated by Fur1 (Table S1). It has been reported using Escherichia coli that Fur also has direct inducive effects on some genes, as well as repressive effects (26). Although it is unclear whether the virulence genes such as eps operon and egl are directly regulated by Fur1, it is possible that Fur1 and Fur2 have inducive effects on target gene expression in strain OE1-1.
An analysis of virulence in tomato plants showed that deleting fur1 or fur2 decreased virulence, but deleting both fur1 and fur2 resulted in an even greater decrease (Fig. 5A and B). Furthermore, we could observe the decrease of bacterial population and virulence on tomato plants in ∆fur1Δfur2 compared to ∆fur1 and ∆fur2 (Fig. 5C). In the case of ∆fur1 and ∆fur2, the bacterial population decreased at 2 DAI but recovered to the same level as that in strain OE1-1 by 6 DAI, while the disease index also became close to that in strain OE1-1 by 12 DAI, suggesting that the single deletion caused the delay in both population increase and disease progression (Fig. 5). On the other hand, the bacterial population increase and the disease progression of ∆fur1Δfur2 did not reach the same level of those in strain OE1-1, suggesting that Fur1 and Fur2 have cooperative effects on the maintenance of both bacterial population and virulence during infection (Fig. 5). However, it is still unclear how the virulence of ∆fur1Δfur2 decreased in tomato plants because the virulence of R. pseudosonalacearum is likely determined as a result of interaction with host plants. It is possible that ∆fur1Δfur2 did not reach the threshold of population to accomplish the full virulence, or that ∆fur1Δfur2’s phenotypic changes reduced its virulence. In addition, an increase in extracellular Fe^3+^-chelating activity possibly explains the decreased virulence of ∆fur1, ∆fur2, and ∆fur1Δfur2 (Fig. 5A and B). Earlier research demonstrated that host plants and microbes compete for available iron (27, 28). A previous in planta transcriptome analysis of Pseudomonas syringae in Arabidopsis thaliana leaves indicated that an iron acquisition pathway is a major target of plant immunity-related mechanisms (29). It is possible that the increased Fe^3+^-chelating activities of ∆fur1 and ∆fur1Δfur2 lead to induced plant immunity. Additionally, excessive amounts of Fe^2+^ may be harmful to bacteria because the Fenton reaction produces hydroxyl radicals in the presence of free Fe^2+^ (8). Excessive intracellular Fe^2+^ levels in ∆fur1 and ∆fur1Δfur2 may lead to decreased virulence. However, further studies are needed to address the effect of inner host plant environment on the bacteria and the bacterial behaviors in response to it. These studies will also address the mechanisms by which the virulence of fur1- and/or fur2-deletion strains resulted in the delay and/or decrease of virulence.
A simple working model for Fur proteins has been developed. Specifically, homodimeric Fe^2+^-binding Fur proteins bind to a cis-element in the target promoter to prevent the binding of RNA polymerase (12). On the basis of the transcriptome data in the current study, both Fur1 and Fur2 may function as repressors (Fig. 2C). In the case of single deletion, the effect of Fur1 was stronger than that of Fur2; however, considering the case of double deletion, Fur2 was also important for siderophore regulation, and there seems to be mechanisms controlling siderophore production through the cooperation of Fur1 and Fur2. The mechanism is unknown, and further biochemical analyses are needed (Fig. 2C). According to our phylogenetic analysis, Fur1 and Fur2 belong to different groups (Fig. 1; Fig. S2). Fur1 has a motif structure that is conserved among Fur proteins in closely related non-RSSC Burkholderia species, whereas Fur2 appears to be a different clade protein with a different C-terminal motif sequence and the predicted structure, suggesting that Fur1 and Fur2 are possibly paralogs with different functions (Fig. 1). The presence of two Fur proteins is a characteristic feature of RSSC strains and possibly mediates the virulence of RSSC. However, differences in the mechanisms underlying Fur1 and Fur2 functions will need to be determined in future studies.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions
Bacterial strains used in this study are listed in Table S2. All R. pseudosolanacearum strains were routinely grown at 30°C in a modified quarter-strength M63 medium [3.75 mM (NH_4_)2_SO_4, 667 µM MgSO_4_, 25 mM KH_2_PO_4_, and 450 nM FeSO_4_] with 20 mM sodium L-glutamate as the carbon source. For control growth conditions (i.e., without Fe^2+^), quarter-strength M63 medium was prepared without 450 nM FeSO_4_. For the siderophore activity assay and growth analysis, 450 nM FeSO_4_ was replaced by 450 nM FeCl_2_. E. coli strains were grown at 37°C in lysogeny broth medium (30). To determine the growth level of R. pseudosolanacearum strains, the OD_600_ value was recorded hourly using a TVS062CA Biophotorecorder (Advantec, Tokyo, Japan).
Phylogenetic analysis
Putative Fur amino acid sequences were obtained from the NCBI database following a BLASTP search using R. pseudosolanacearum strain OE1-1 Fur1 and Fur2 protein sequences as queries. Protein sequences in RSSC strains for all four phylotypes and closely related species (31) were used as references. Finally, we obtained eight sequences from RSSC strains and 14 sequences from closely related species. Additionally, the Fur protein in E. coli strain K-12 served as an outgroup for the phylogenetic analysis, which was performed using MEGA11 (32). The MUSCLE program was used for aligning multiple sequences. A phylogenetic tree was constructed according to the maximum likelihood method, with 1,000 bootstrap replicates and the following options: Test of Phylogeny: None; Model/Method: WAG model; Rates Among Sites: Gamma distributed with invariant sites (G+I); Number of Discrete Gamma Categories: 5; Gaps/Missing Data Treatment: Complete deletion; ML Heuristic Method: Nearest-neighbor interchange; Initial Tree for ML: Make initial tree automatically; Branch Swap Filter: Very strong. To compare domain structures, the Clustal Omega web server of EMBL was used to align multiple sequences (https://www.ebi.ac.uk/Tools/msa/clustalo/). Previously determined domain structures (16) were used for the analysis. To obtain the identity scores of full-length amino acid sequences, a FASTA file was first generated using the Clustal Omega web server, with Fur1 and Fur2 amino acid sequences from all four RSSC phylotypes as inputs. The FASTA file was analyzed using the Ident and Sim function of the Sequence Manipulation Suite (https://www.bioinformatics.org/sms2/ident_sim.html) to calculate the amino acid sequence identity (33).
Conserved motif analysis
Fur protein amino acid sequences used for the phylogenetic analysis were screened for conserved motifs using MEME (version 5.5.7; https://meme-suite.org/meme/tools/meme), with the number of motifs set to 10 (34).
Prediction of protein structures by AlphaFold 3
Structures of Fur1 and Fur2 were predicted using the online AlphaFold server with AlphaFold 3 (https://alphafoldserver.com/; [35]) by inputting each amino acid sequence of Fur1 and Fur2 with default parameters. The predicted models were colored with the same colors of the MEME analysis outputs and represented by ChimeraX (36).
General DNA manipulations
Genomic DNA was isolated, plasmid DNA was manipulated, and PCR amplifications were completed using standard techniques (37). R. pseudosolanacearum strain OE1-1 was transformed via electroporation (38). Double-stranded DNA sequencing templates were prepared using the GenElute Plasmid Miniprep Kit (Sigma Chemical Co., St. Louis, MO, USA). Sequences were determined using the ABI Prism 3100-Avant Genetic Analyzer (Applied Biosystems, Tokyo, Japan). DNA sequencing data were analyzed using DNASIS-Mac software (Hitachi Software Engineering, Yokohama, Japan).
Generation of fur1 and fur2 deletion mutants
Plasmids used in this study are listed in Table S2. A 697 bp DNA fragment (delta-fur1-1) was amplified by PCR using strain OE1-1 genomic DNA (NCBI Reference Sequence: NZ_CP009764.1) as the template and primers delta-fur1-1-FW (5′-CGgga tccAC GGAAT CGACG GAGCG C-3′), which includes a BamHI site (lowercase letters), and delta-fur1-1-RV (5′-CTGTT GCGTC ACATG CGCCT GGCTC CCTA-3′). A 706 bp DNA fragment (delta-fur1-2) was amplified by PCR using strain OE1-1 genomic DNA and primers delta-fur1-2-FW (5′-AGGCG CATGT GACGC AACAG CCGCC TCAT-3′) and delta-fur1-2-RV (5′-CCCaa gcttA GCTGG ACCTG GCGTT CTC-3′) with a HindIII site (lowercase letters). Amplified delta-fur1-1 and delta-fur1-2 sequences, along with primers delta-fur1-1-FW and delta-fur1-2-RV, were used for a PCR amplification of a 1,384 bp DNA fragment, which was then digested with BamHI (Takara Bio, Ohtsu, Japan) and HindIII (Takara Bio). The resulting 1.4 kb fragment was inserted into the pK18mobsacB vector digested with BamHI and HindIII (39) to produce the pdelta-fur1 recombinant plasmid. A 769 bp DNA fragment (delta-fur2-1) was amplified by PCR using strain OE1-1 genomic DNA and primers delta-fur2-1-FW (5′-CGgga tccAA AAGGT CAACG GCCTG CGC-3′), which includes a BamHI site (lowercase letters), and delta-fur2-1-RV (5′-TGCAA CGCCT ACATG GTGTT GGGGC GAGAA G-3′). A 706 bp DNA fragment (delta-fur2-2) was amplified by PCR using strain OE1-1 genomic DNA and primers delta-fur2-2-FW (5′-CCAAC ACCAT GTAGG CGTTG CACGC GCTTC A-3′) and delta-fur2-2-RV (5′-CCCaa gcttG AGAAG GAAGA CCCGG TGG-3′) with a HindIII site (lowercase letters). Amplified delta-fur2-1 and delta-fur2-2 sequences, along with primers delta-fur2-1-FW and delta-fur2-2-RV, were used for PCR amplification of a 1,444 bp DNA fragment, which was then digested with BamHI and HindIII. The obtained 1.4 kb fragment was inserted into the pK18mobsacB vector, digested with BamHI and HindIII, to produce the pdelta-fur2 recombinant plasmid. An electroporation-based method was used to insert recombinant plasmids into strain OE1-1 competent cells, which were prepared as previously described (40). Kanamycin-sensitive, sucrose-resistant recombinants, Δfur1 or ∆fur2 (Table S2), were selected. To verify the deletion of *fur1 *(RSc2747), a 1,636 bp DNA fragment (WT: 2,061 bp) was amplified by PCR using primers delta-fur1-SQ-FW (5′-CTCGC TGGTG GTCAT GCTG-3′) and delta-fur1-SQ-RV (5′-GCTTC CTGCT GCGCG TC-3′). The fragment sequence was analyzed using primers delta-fur1-SQ-FW and delta-fur1-SQ-RV. To confirm *fur2 *(RSp0247) was deleted, a 1,826 bp DNA fragment (WT: 2,276 bp) was amplified by PCR using primers delta-fur2-SQ-FW (5′-ATGGT CCGCT GTTCA AGGGC-3′) and delta-fur2-SQ-RV (5′-TGGCC GAGCA GCCGA TC-3′). The fragment sequence was analyzed using primers delta-fur2-SQ-FW and delta-fur2-SQ-RV. To delete both fur1 and fur2, fur2 was deleted from ∆fur1.
RNA-seq
R. pseudosolanacearum strains were grown in quarter-strength M63 medium or modified quarter-strength M63 medium without Fe^2+^ until the OD_600_ value reached 0.3. The growth level was checked in a timely manner by recording OD_600_ hourly using a TVS062CA Biophotorecorder (Advantec, Tokyo, Japan) in the same way as bacterial growth levels were analyzed. Total RNA was extracted using a High Pure RNA Isolation kit (Roche Diagnostics, Mannheim, Germany). Ribosomal RNA was eliminated from the extracted RNA using a Ribo-Zero rRNA Removal kit (Gram-negative bacteria) (Illumina, Madison, WI, USA), as previously described (41). An oriented, paired-end RNA-seq analysis (2 × 100 bp) was performed using an Illumina HiSeq 2500 system or a DNBSEQ-G400 system. Generated reads were trimmed using Cutadapt (version 1.1; http://code.google.com/p/cutadapt/ [42]) and then mapped to the GMI1000 strain genome using Bowtie2 (version 2.4.2) (43). Read count data were obtained using featureCounts (version 2.0.0) (44). Three independent biological replicates were analyzed per strain.
Data analysis
RNA-seq data were analyzed using R cran (45). Genes with no RNA-seq reads in at least one sample in the raw count data set were excluded. RNA-seq read counts of the remaining genes were normalized using calcNormFactors (trimmed mean of M-values normalization) in edgeR (46). To extract genes with significant changes in transcript levels, the following thresholds were applied: q-value < 0.05 and |log_2_(fold-change)| ≥ 2. FDR (q-value) was calculated using P-values estimated by edgeR according to the Benjamini–Hochberg method (47). Heatmaps were created using the R package pheatmap.
Gene ontology enrichment analysis
A GO enrichment analysis was performed using the R package GoSeq (48), with GO terms obtained from QuickGO (https://www. ebi.ac.uk/Quick GO/). Fold enrichment was calculated as follows: (number of DEGs annotated with the term/number of all genes annotated with the term)/(number of DEGs/number of all genes).
Extracellular Fe3+-chelating activity analysis
The extracellular Fe^3+^-chelating activity of R. pseudosolanacearum strains was analyzed as previously described (18, 20). Briefly, R. pseudosolanacearum strains were grown at 30°C in modified quarter-strength M63 medium containing 450 nM FeCl_2_ or 0 nM FeCl_2_ (control) for 18 h, and then the culture concentration was adjusted to 2.0 × 10^9^ CFU/mL using 0.1 M PIPES buffer (pH 6.5). After a 6-hour incubation, each culture was passed through a 0.2 μm pore filter. Next, a 100 µL aliquot of each culture or PIPES buffer (i.e., reference) was added to 100 μL chromazurol S (CAS) solution (2.4 mM hexadecyl-trimethyl ammonium bromide, 0.06 mM FeCl_3_, 0.6 mM HCl, and 0.6 mM CAS in PIPES buffer). After a 30-minute incubation at 30°C, the absorbance at 630 nm was measured. To calculate the Fe^3+^-chelating activity, the absorbance of the reference was subtracted from the total absorbance. Seven or eight biological replicates were used per assay. Mean values were analyzed by analysis of variance with Tukey–Kramer’s honestly significant difference test to assess the significance of any differences among R. pseudosolanacearum strains (strain OE1-1 as the reference).
Virulence assay
Three-week-old tomato plants (Solanum lycopersicum ‘Ohgata-Fukuju’) were inoculated with R. pseudosolanacearum strains (1.0 × 10^8^ CFU/mL) using a published root-dip method (49). Plants were monitored daily for wilting symptoms, which were rated according to the following disease index scale: 0, no wilting; 1, 1%–25% wilting; 2, 26%–50% wilting; 3, 51%–75% wilting; 4, 76%–99% wilting; and 5, dead. For each bacterial strain, two independent groups were tested, with six biological replicates per group.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mansfield J, Genin S, Magori S, Citovsky V, Sriariyanum M, Ronald P, Dow M, Verdier V, Beer SV, Machado MA, Toth I, Salmond G, Foster GD. 2012. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol Plant Pathol 13:614–629. doi:10.1111/j.1364-3703.2012.00804.x 22672649 PMC 6638704 · doi ↗ · pubmed ↗
- 2Inoue K, Takemura C, Senuma W, Maeda H, Kai K, Kiba A, Ohnishi K, Tsuzuki M, Hikichi Y. 2023. The behavior of Ralstonia pseudosolanacearum strain OE 1-1 and morphological changes of cells in tomato roots. J Plant Res 136:19–31. doi:10.1007/s 10265-022-01427-336427093 · doi ↗ · pubmed ↗
- 3Rivera-Zuluaga K, Hiles R, Barua P, Caldwell D, Iyer-Pascuzzi AS. 2023. Getting to the root of Ralstonia invasion. Semin Cell Dev Biol 148–149:3–12. doi:10.1016/j.semcdb.2022.12.00236526528 · doi ↗ · pubmed ↗
- 4Tsuzuki M, Inoue K, Kiba A, Ohnishi K, Kai K, Hikichi Y. 2023. Infection route in tomato roots and quorum sensing of Ralstonia pseudosolanacearum strain OE 1-1. Physiol Mol Plant Pathol 125:101995. doi:10.1016/j.pmpp.2023.101995 · doi ↗
- 5Vasse J, Frey P, Trigalet A. 1995. Microscopic studies of intercellular infection and protoxylem invasion of tomato roots by Pseudomonas solanacearum. Mol Plant-Microbe Interact 8:241. doi:10.1094/MPMI-8-0241 · doi ↗
- 6de Pedro-Jové R, Puigvert M, Sebastià P, Macho AP, Monteiro JS, Coll NS, Setúbal JC, Valls M. 2021. Dynamic expression of Ralstonia solanacearum virulence factors and metabolism-controlling genes during plant infection. BMC Genomics 22:170. doi:10.1186/s 12864-021-07457-w 33750302 PMC 7941725 · doi ↗ · pubmed ↗
- 7Ratledge C, Dover LG. 2000. Iron metabolism in pathogenic bacteria. Annu Rev Microbiol 54:881–941. doi:10.1146/annurev.micro.54.1.88111018148 · doi ↗ · pubmed ↗
- 8Fenton HJH. 1894. LXXIII.—Oxidation of tartaric acid in presence of iron. J Chem Soc Trans 65:899–910. doi:10.1039/CT 8946500899 · doi ↗