Vibrio natriegens is sensitive to its acidic fermentation products
Nicholas W. Haas, James B. McKinlay

TL;DR
Vibrio natriegens, a fast-growing bacterium used in biotechnology, is more sensitive to its own acidic fermentation byproducts than Escherichia coli, which affects its survival and productivity in anaerobic conditions.
Contribution
The study reveals that V. natriegens lacks key acid resistance genes and is more sensitive to its fermentation products compared to E. coli.
Findings
V. natriegens consumes less glucose and produces lower fermentation product titers than E. coli under anaerobic conditions.
V. natriegens dies rapidly after growth in fermentation, but this can be mitigated by increasing medium buffering.
The V. natriegens genome lacks most acid resistance genes found in E. coli and Vibrio cholerae.
Abstract
Vibrio natriegens is an emerging bacterial platform for a range of biotechnological applications due to its rapid growth and ease of genetic manipulation. Whereas much has been learned about V. natriegens’ aerobic physiology, comparatively little is known about its anaerobic fermentative physiology, despite its relevance to many industrial conditions. We compared the metabolic parameters of V. natriegens versus another biotechnologically relevant bacterium, Escherichia coli, under fermentative conditions. Both species excreted a similar array of fermentation products, but V. natriegens consumed less glucose and had a lower product titer. V. natriegens also exhibited rapid death, reaching extinction within 12 h after the growth phase, 3 days sooner than E. coli. Rapid V. natriegens death was avoided, and glucose consumption and product titers improved, by increasing the buffering…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
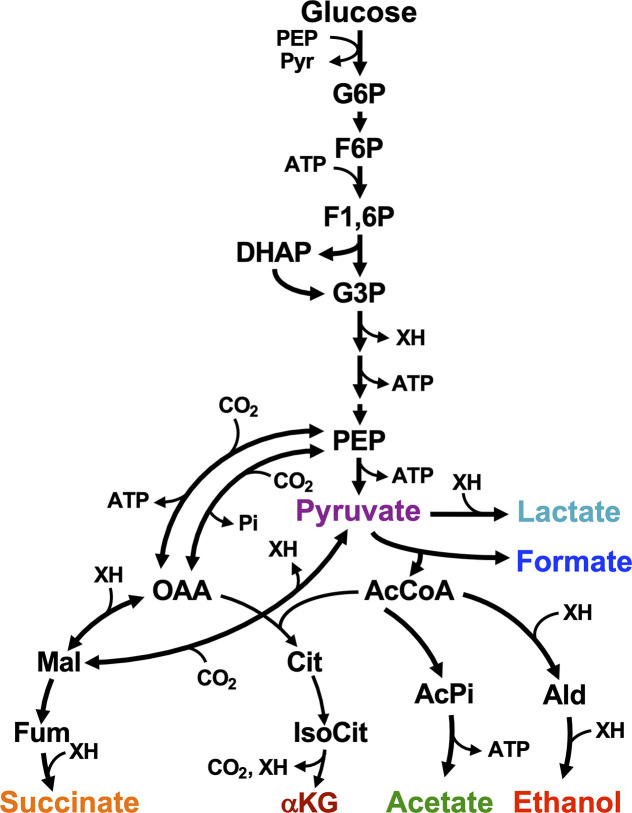 Fig 1
Fig 1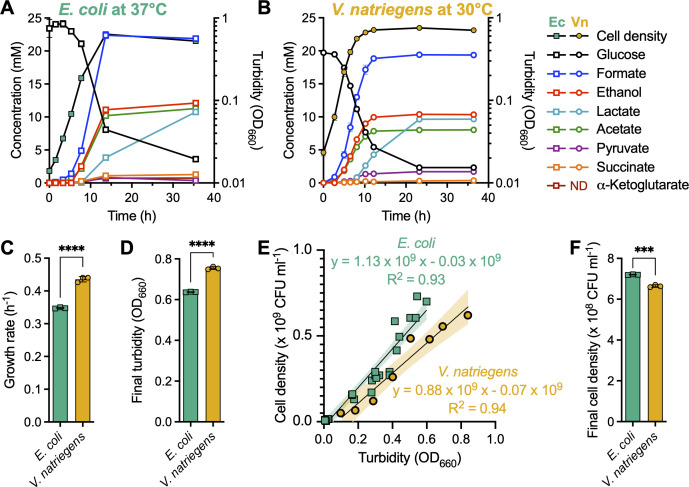 Fig 2
Fig 2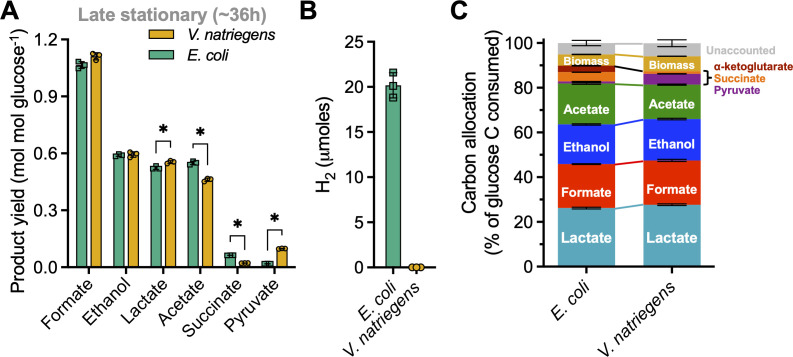 Fig 3
Fig 3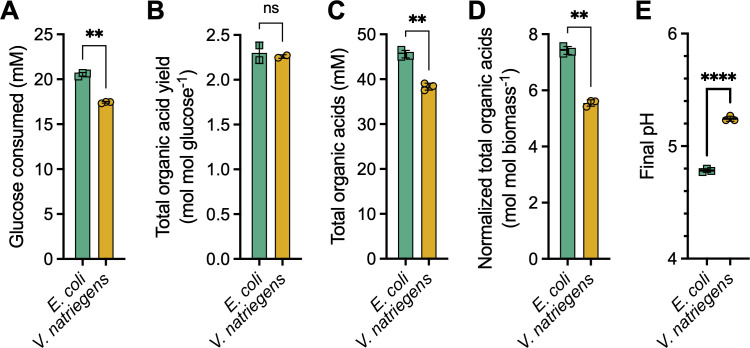 Fig 4
Fig 4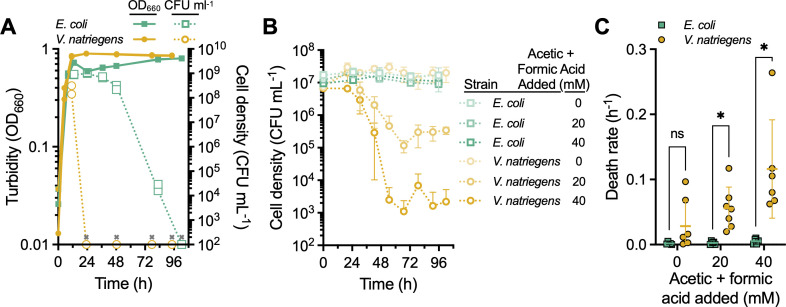 Fig 5
Fig 5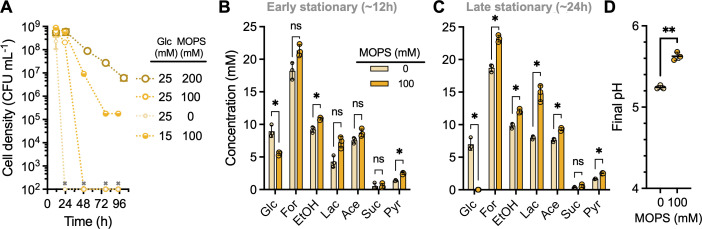 Fig 6
Fig 6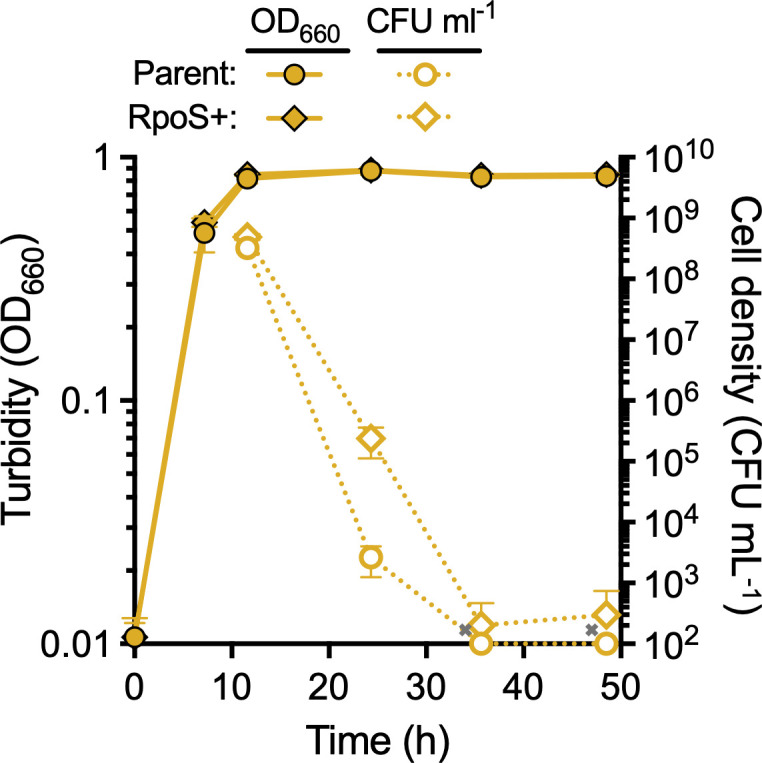 Fig 7
Fig 7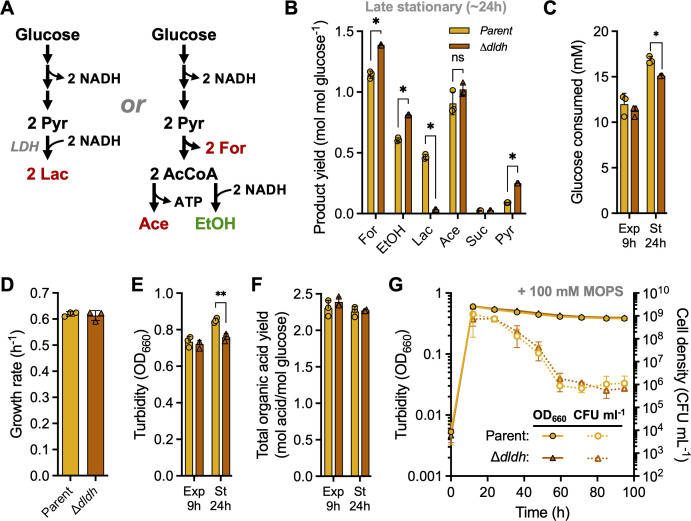 Fig 8
Fig 8| Gene | Description | Query NCBI RefSeq | Coverage (%) | Identity (%) | Locus tag | Protein ID |
|---|---|---|---|---|---|---|
| Cell envelope/general stress response | ||||||
| | σE/σ24/RpoE | 100 | 75 | PN96_RS01020 |
| |
| | Anti-σE | 96 | 41 | PN96_RS01025 |
| |
| | Anti-σE stabilizer | 96 | 44 | PN96_RS01030 |
| |
| | Regulatory protein | 81 | 34 | PN96_RS01035 |
| |
| | Transcriptional regulator | 98 | 61 | PN96_RS14495 |
| |
| | Sensor histidine kinase | 98 | 44 | PN96_RS14490 |
| |
| | Periplasmic stress protein | 71 | 31 | PN96_RS14500 |
| |
| | Cyclopropane FA synthase | 90 | 37 | PN96_RS07970 |
| |
| | Periplasmic chaperones | No homolog | ||||
| | Periplasmic chaperone | No homolog | ||||
| | Periplasmic endoprotease | No homolog | ||||
| | LPS modification | 96 | 55 | PN96_RS12400 |
| |
| | LPS modification | 98 | 63 | PN96_RS12375 |
| |
| | Transcriptional regulator |
| 100 | 53 | PN96_RS09525 |
|
| | Porin |
| 100 | 65 | PN96_RS01620 |
|
| | Phosphotransferase |
| No homolog | |||
| FHL | ||||||
| | Transcriptional regulator | 47 | 44 | PN96_RS02620 |
| |
| | Regulator of FhlA | No homolog | ||||
| | FHL subunit B | 93 | 29 | PN96_RS05940 |
| |
| | FHL subunit C | 55 | 27 | PN96_RS21860 |
| |
| | FHL subunit D, E | No homolog | ||||
| | FHL subunit F | 67 | 29 | PN96_RS05960 |
| |
| | FHL subunit G, assembly | No homolog | ||||
| | Hydrogenase Ni incorporation | No homolog | ||||
| | Hydrogenase Ni incorporation | 25 | 26 | PN96_RS01805 |
| |
| | Hydrogenase maturation | No homolog | ||||
| Amino acid decarboxylases/ ion balance | ||||||
| | Glutamate decarboxylase A | No homolog | ||||
| | Glutamate decarboxylase B | No homolog | ||||
| | GABA antiporter | No homolog | ||||
| | σS/σ38/RpoS | 96 (E71*) | 75 | PN96_RS01145 |
| |
| | Arginine decarboxylase | No homolog | ||||
| | Arginine:agmatine antiporter | 96 | 27 | PN96_RS21655 |
| |
| | Lysine decarboxylase | No homolog | ||||
| | Lysine:cadaverine antiporter | 72 | 26 | PN96_RS18150 |
| |
| | Ornithine decarboxylase | No homolog | ||||
| | Ornithine decarboxylase | No homolog | ||||
| | H+/Cl- antiporter | 93 | 56 | PN96_RS18465 |
| |
| | H+/Cl- antiporter | No homolog | ||||
| | Na+/K+ antiporter | 99 | 42 | PN96_RS13405 |
| |
| | K+ channel | 100 | 69 | PN96_RS13405 |
| |
| | K+ transporter (ATP) | No homolog | ||||
| | K+ transporter (ATP) | 75 | 29 | PN96_RS09780 |
| |
| | K+ transporter (ATP) | No homolog | ||||
| Acetoin and 2,3-butanediol production | ||||||
| | Acetolactate decarboxylase |
| No homolog | |||
| | Acetoin reductase |
| Multiple SDR oxidoreductase hits | |||
| Small proteins | ||||||
| | Acid stress small protein | No homolog | ||||
| | Acid stress small protein | No homolog | ||||
| | Acid stress small protein | No homolog | ||||
| | Acid stress small protein | No homolog | ||||
| Primer | Sequence (5′→ 3′) | Description |
|---|---|---|
| NH022 |
| F for amplifying Δ |
| NH025 |
| R for amplifying Δ |
| NH014 |
| F for confirming |
| NH015 |
| R for confirming |
| NH074 |
| Upstream F for Δ |
| NH075 |
| Upstream R for Δ |
| NH076 |
| Downstream F for Δ |
| NH077 |
| Downstream R for Δ |
| NH078 |
| F for confirming Δ |
| NH079 |
| R for confirming Δ |
| NH238 |
| F for amplifying |
| NH239 |
| R for amplifying |
| NH240 |
| F for confirming |
| NH241 |
| R for confirming |
- —Division of Molecular and Cellular Bioscienceshttp://dx.doi.org/10.13039/100000152
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEnzyme Production and Characterization · Microbial Metabolic Engineering and Bioproduction · Bacterial biofilms and quorum sensing
INTRODUCTION
Bioprocessing utilizes native or engineered microbes to generate value-added products from renewable resources, offering sustainable alternatives to petroleum-based counterparts (1–5). A diverse set of microbes has been proposed as platforms, or chassis, for bioprocessing (6–8). The marine bacterium Vibrio natriegens has increasingly gained attention as a chassis for bioprocessing primarily due to its rapid growth and metabolism (9–11) and the relative ease with which combinatorial genetics can be performed through natural transformation (12–15). Proof-of-concept studies have demonstrated that V. natriegens can be engineered to produce a variety of value-added products from a range of carbon substrates (11). However, whereas many bioprocesses use anoxic fermentative conditions to maximize product yields and minimize microbial growth, most V. natriegens studies have used oxic conditions, with a few exceptions (10, 16, 17).
Under anoxic conditions with glucose, V. natriegens carries out a mixed-acid fermentation, excreting ethanol and a variety of organic acids (Fig. 1) (10). The accumulation of organic acids acidifies the medium. In general, acid stress negatively impacts metabolism and viability. Negative impacts of an acidic environment are amplified when the low pH favors protonation of excreted organic acids, allowing them to diffuse into the cytoplasm where they then dissociate, acidifying the cytoplasm, weakening the proton motive force, and increasing the cytoplasmic anion concentration (18, 19). Even under aerobic conditions, V. natriegens growth is impacted by acidification from acetic acid that accumulates due to overflow metabolism (20). Thus, it is expected that these negative impacts would be exacerbated under anoxic fermentative conditions where there is a greater accumulation of organic acids. Understanding how V. natriegens responds to its fermentation products is important for designing future bioprocesses, as organic acids affect important production parameters including rate, fermentation time, yield, and associated costs for neutralization and waste treatment (21–23).
V. natriegens fermentative metabolism. Reactions are based on NCBI gene annotations. Acidic products are named here for their carboxylate forms. XH, electron carriers [e.g., NAD(P)H].
Whereas other Vibrios like V. cholerae can switch to neutral fermentation products in response to acidic conditions (24), production of neutral fermentation products by V. natriegens has not been observed (10). Fermentative bacteria like Escherichia coli have other mechanisms to deal with acid stress (23, 25). For example, E. coli can offset acidosis by converting formic acid into H_2_ and CO_2_ gases using formate hydrogen lyase (FHL) (26). In the presence of certain amino acids, E. coli and V. cholerae can also eliminate cytoplasmic protons via amino acid decarboxylation reactions (27, 28). As the medium acidifies during fermentation, E. coli also shifts its fermentation profile towards lactic acid (29), which might also offset acidosis by producing a single acid instead of the combination of acetic and formic acid (30) and avoiding accumulation of intracellular acetate anions (31). E. coli also has different global regulatory responses to organic acids like acetate, formate, and lactate (30, 32–34). The extent to which V. natriegens can use these or other acid resistance mechanisms during mixed acid fermentation is unknown.
Here, we compared the growth and metabolic parameters of V. natriegens versus E. coli under anoxic fermentative conditions. Whereas both organisms produced similar fermentation products, E. coli cultures consumed more glucose, accumulated more organic acids, and exhibited longer stationary-phase viability. We conclude that V. natriegens is comparatively sensitive to fermentative organic acids due to a lack of acid resistance mechanisms.
RESULTS AND DISCUSSION
V. natriegens and E. coli have similar fermentative product yields
V. natriegens is often compared to E. coli, a bacterium that is well-established in synthetic biology, as a potential platform for bioprocessing (9). However, a direct comparison of growth and metabolic parameters under fermentative conditions has not been reported. We grew E. coli MG1655 and V. natriegens ATCC14048 in their respective anoxic minimal media with 25 mM glucose as the sole carbon source. The media differed by a higher NaCl concentration for V. natriegens and by temperature; 37°C was used for E. coli versus 30°C for V. natriegens. Although V. natriegens was originally reported to grow ~1.06 times faster at 37°C (35), several groups grow V. natriegens at 30°C (12, 36–38), including in comparisons to E. coli where advantages have been noted at 30°C for V. natriegens protein production (39–42). Despite the lower growth temperature, V. natriegens grew 1.3 times faster than E. coli (Fig. 2A through C). Although V. natriegens reached a higher turbidity (OD_660_; Fig. 2D), E. coli has a higher colony-forming units (CFU) mL^−1^:OD_660_ ratio (Fig. 2E) and was thus estimated to achieve a 1.2-fold higher actual cell density (CFU mL^−1^) than V. natriegens (Fig. 2F).
*V. natriegens grows faster but to a lower cell density while producing a similar array of fermentation products as E. coli. E. coli (A) and V. natriegens (B) were grown in 60 mL of anoxic minimal media at 37°C or 30°C, respectively, and growth, glucose, and fermentation products were monitored. The same cultures were used to determine exponential growth rate (C) and final (highest) cell density (D). Standard curves to convert turbidity (OD660) measurements of cell density to CFU mL−1 (E) suggest that the difference in final cell densities between the two species is opposite when considering turbidity (D) versus CFU mL−1 (F). (C, D, F) Points, biological replicate values; bars, mean; error bars, SD; n = 3. Statistical differences were determined using an unpaired, two-tailed t-test; ***, P < 0.001; ***, P < 0.0001. (E) Points, biological replicate values; lines, fitting from simple linear regression analysis; shading, SD.
Late stationary-phase (36 h) fermentation product yields between E. coli and V. natriegens were either statistically similar or exhibited small differences for the most abundant fermentation products: formic acid, no difference; ethanol, no difference; lactic acid, 1.06-fold higher for V. natriegens; acetic acid, 1.20-fold higher for E. coli (Fig. 3A). Both species produced lactic acid as they exited exponential phase, with continued production into stationary phase (Fig. 2A and B). Greater fold differences were observed for minor fermentation product concentrations (Fig. 2A and B) and yields (Fig. 3A): (i) E. coli produced ~1.3 mM succinic acid whereas levels from V. natriegens were near the detection limit, (ii) V. natriegens produced ~1.7 mM pyruvate that was stable in the supernatant whereas E. coli transiently produced as much as 0.8 mM pyruvate, and (iii) E. coli produced ~0.8 mM α-ketoglutarate, whereas none was detected in V. natriegens supernatants (Fig. 2A and B). Another key difference was that V. natriegens did not produce H_2_, whereas E. coli is known to produce H_2_ via FHL as the pH drops below neutral (Fig. 3B) (43, 44); the V. natriegens genome does not encode FHL nor any hydrogenase (not shown). A carbon balance analysis revealed that only 5 ± 1% (E. coli) and 6 ± 2% (V. natriegens) of carbon was unaccounted for outside of fermentation products and biomass (Fig. 2C). Missing carbon could be explained in part by amino acids; Hoffart et al. found that extracellular alanine, valine, and glutamate could account for 4% of glucose carbon consumed by V. natriegens under fermentative conditions (10). Below, we focus on the major organic acid fermentation products.
*V. natriegens and E. coli major soluble fermentation product profiles are similar. Fermentation product yields (A), H2 accumulated in the headspace (limit of detection ~10 nmol) (B), and carbon allocations (C) were determined using late stationary-phase (36 h) samples. All samples were from the same cultures as for Fig. 2 except for H2. (A–C) Points, biological replicate values; bars, mean; error bars, SD; n = 3. (A) Statistical differences were determined using an unpaired, two-tailed t-test; , P < 0.05. For formic acid; EtOH, ethanol; Lac, lactic acid; Ace, acetic acid; Suc, succinic acid; Pyr, pyruvate. (C) Turbidity measurements were converted into moles of biomass carbon (C) as described in Materials and Methods.
V. natriegens is more sensitive to acid than E. coli
Neither strain fully consumed the glucose provided (Fig. 2A and B) but E. coli consumed 1.2 times more glucose than V. natriegens (Fig. 4A). A possible reason is that each species reached its acid tolerance threshold, and V. natriegens has a lower acid tolerance. Indeed, although each species had a similar total organic acid yield (Fig. 4B), E. coli accumulated 1.2-fold more organic acids (Fig. 4C) and 1.3-fold more organic acids per cell (Fig. 4D). As a result, the final E. coli pH was more acidic at 4.8 ± 0.0 compared to 5.2 ± 0.0 for V. natriegens (Fig. 4E).
*V. natriegens consumes less glucose and produces less organic acid than E. coli. Glucose consumed (A), total organic acid yield (B), total organic acid concentration (C), organic acid concentration normalized for biomass carbon (D), and supernatant pH (E) were determined using late stationary-phase samples (36 h, except for pH, taken at 24 h). All samples were from the same cultures as for Fig. 2 except for pH measurements. Points, biological replicate values; bars, mean; error bars, SD; n = 3. Statistical differences were determined using an unpaired, two-tailed t-test; ns, not significant; **, P < 0.01; ***, P < 0.0001. (D) Turbidity measurements were converted into moles of biomass carbon (C) as described in Materials and Methods.
To compare the acid tolerance between the two species, we tracked cell viability by CFUs during stationary phase (Fig. 5A). Herein, we assume that a decline in CFUs is equivalent to death, especially given the acidic conditions, but we acknowledge that a viable but non-culturable (VBNC) state is also possible, as has been described for other Vibrios (45). However, whereas the VBNC state is important for epidemiology, VBNC cells are of no practical use for value-added chemical production due to their lack of metabolic activity. Although death was not evident from turbidity measurements, CFU measurements indicated that both species ultimately went extinct. However, whereas viable E. coli cells were detected up to 82 h after inoculation, no viable V. natriegens cells were detected 24 h after inoculation (limit of detection = 10^2^ CFU mL^−1^), about 12 h after the end of the exponential growth phase (Fig. 4A).
*V. natriegens is sensitive to acidic fermentation products compared to E. coli. (A) Culture turbidity (solid lines; OD660) was tracked in minimal media with 25 mM glucose over the entire experiment. Viable cell density (dotted lines; CFU mL−1) was tracked from the end of the growth phase (~12 h). All data points from the duplicate cultures are shown. Symbols with an “x” were below the detection limit of 102 CFU mL−1. (B) Viable cell counts were monitored in cell suspensions inoculated in late exponential phase to defined media lacking glucose with different concentrations of a 1:1 solution of acetic and formic acid. Data points, mean; error bars, SD; n = 3. (C) Death rates determined from the slope of a semi-log plot fitted to the data from panel B and two other similar experiments that differed by starting cell densities (range: 0.05–2.0 × 107 CFU mL−1) and incubation time (range: 140–155 h). Horizontal line, mean; error bars, SD; n = 5–7. Statistical differences were determined using unpaired, two-tailed t-tests; ns, not significant; , P < 0.05. (A, B) The detection limit is 102 CFU mL−1.
To verify that organic acids were a primary driver of V. natriegens cell death, we inoculated late exponential phase cultures (V. natriegens, 0.65–0.69 OD_660_; E. coli, 0.82–0.87 OD_660_) to media lacking glucose but with different concentrations of a 1:1 mixture of acetic and formic acid. The cultures used to prepare these suspensions were grown with less glucose (15 mM) to ensure less acid exposure prior to the assay. Without added organic acids, both species showed little decline in cell viability (Fig. 5B and C). E. coli viability was also unaffected by 20 and 40 mM organic acids (Fig. 5B and C). In contrast, increasing the organic acid concentration progressively decreased V. natriegens viability (Fig. 5B) and increased the death rate (Fig. 5C).
We then verified that the V. natriegens sensitivity to organic acids is dependent on the pH by increasing the buffering capacity of medium with either 100 or 200 mM 4-morpholinepropanesulfonic acid (MOPS) buffer. In each case, viable cell counts exceeded 10^8^ CFU mL^−1^ at the onset of stationary phase (Fig. 6A). Unlike the rapid extinction observed when MOPS was excluded, 100 mM MOPS delayed extinction by 24 h and 200 mM MOPS avoided extinction within the 100 h time course (Fig. 6A). Even though cells still went extinct with 100 mM MOPS, glucose was fully consumed by 24 h, with continued production of most fermentation products, especially lactic acid (Fig. 6B and C). The final pH was also correspondingly higher when 100 mM MOPS was provided, as expected (Fig. 6D). Thus, we conclude that the rapid extinction of V. natriegens upon reaching stationary phase under fermentative cultures is due to acid sensitivity.
*Increasing the buffer concentration of the growth medium alleviates V. natriegens organic acid sensitivity. (A) V. natriegens viable cell density was tracked from the end of the growth phase (~12 h) in minimal media with different concentrations of glucose and MOPS (pH 7). The “0 mM MOPS” data are the same as in Fig. 5A. Data points, mean; error bars, range; n = 2; some error bars are smaller than the data point symbols. Symbols with an “x” were below the detection limit of 102 CFU mL−1. (B, C) Remaining glucose and accumulated fermentation products from V. natriegens cultures grown with 25 mM glucose in early stationary phase (B, ~ 12 h) and late stationary phase (C, ~ 24 h) with or without 100 mM MOPS (pH 7). Glc, glucose; For, formic acid; EtOH, ethanol; Lac, lactic acid; Ace, acetic acid; Suc, succinic acid; Pyr, pyruvate. (D) Late stationary-phase (24 h) pH values from cultures grown with 25 mM glucose with or without 100 mM MOPS (pH 7). Values without MOPS are the same as those in Fig. 4E. (B–D) Points, biological replicate values; bars, mean; error bars, SD; n = 3. Statistical differences were determined using unpaired, two-tailed t-tests; ns, not significant; *, P < 0.05; *, P < 0.01.
V. natriegens lacks acid resistance mechanisms
We sought an explanation for the stark difference between V. natriegens and E. coli survival in acidic conditions. Bacteria like E. coli and some well-characterized Vibrios, like V. cholerae, have mechanisms to deal with acid stress both during fermentative conditions and to transit the highly acidic stomach (23, 25, 28, 46). To determine the acid resistance repertoire of V. natriegens, we performed BLASTp searches (47) using known E. coli and V. cholerae acid resistance proteins as query sequences (Table 1). We focused on mechanisms involving cell envelope stress and specific acid response mechanisms. We excluded other proteins that emerge in functional genomic surveys but are commonly used by most bacteria across diverse conditions (30, 32–34, 48).
Based on our homolog threshold criteria of 25% identity over 50% of the query sequence, V. natriegens likely has several proteins to respond to cell envelope stress, which could include acid stress, but also other stressors, like heat. For example, V. natriegens has proteins with 53 and 65% amino acid identity to the V. cholerae transcriptional regulator ToxR and porin OmpU, respectively. These proteins are involved in V. cholerae tolerance to organic acids at low pH but can also respond to other stressors (46). V. natriegens also has proteins with 55 and 63% amino acid identity to the E. coli LPS-modification enzymes that were demonstrated to improve acid tolerance (48). We also identified a σ^E^ homolog and associated proteins that, in E. coli, activate a regulon in response to denatured outer membrane proteins, and the Cpx two-component system that responds to periplasmic and inner membrane stress signals (49). Among genes of the E. coli Cpx regulon, V. natriegens also has cfa, which encodes a protein that produces cyclopropane fatty acids that can protect against various stressors, including acidity (50). However, V. natriegens appears to lack many of the chaperones and proteases (i.e., degP, hdeABD, skp) that are crucial in E. coli for the proper folding or degradation of cell envelope proteins under stress conditions (51–53).
When considering specific responses to acid stress, the V. natriegens genome suggests that it is even less well-equipped. FHL, a complex of formate dehydrogenase (FDH) and hydrogenase, converts formic acid into CO_2_ and H_2_ gases (26, 43). Whereas V. natriegens has FDH protein homologs, V. natriegens does not have hydrogenase genes. In agreement with this bioinformatic result, no H_2_ was detected in V. natriegens fermentative cultures by gas chromatography (Fig. 3B).
To deal with anion accumulation from organic acids re-entering the cell, E. coli can accumulate K^+^ (31). V. natriegens has one homolog to an E. coli TrkG or TrkH K^+^ transporter (Table 1). However, it only has a protein with homology for one subunit of an ATP-dependent K^+^ transporter (Table 1); V. cholerae also lacks this transporter (not shown). V. natriegens also lacks homologs to several small proteins that respond to acute acid stress in E. coli (54) and a homolog to the V. cholerae regulatory phosphotransferase vc1080, which while not an acid tolerance protein itself, is downregulated to avoid formic acid toxicity (55).
V. natriegens also lacks amino acid decarboxylases that can eliminate cytoplasmic protons in both E. coli and V. cholerae (23, 25, 27, 28), but it has one of two H^+^/Cl^-^ antiporters that help prevent hyperpolarization of the membrane during the use of amino acid decarboxylases (56, 57). Amino acid decarboxylases for acid resistance are distinct from anabolism and require an external source of amino acids. We would thus not expect amino acid decarboxylases to be effective in our minimal medium that did not contain amino acids.
Amino acid decarboxylases can be regulated by a general stress response sigma factor RpoS in some pathogenic E. coli (25). Our V. natriegens strain has an rpoS mutation causing a premature stop codon (RpoS^E71*^). Given that the RpoS regulon is broad, we decided to investigate if the mutation affects V. natriegens acid tolerance in other ways. We thus repaired the stop codon to create an RpoS+ strain. We then compared the survival of the RpoS+ strain to that of the parent after entering stationary phase under fermentative conditions. The decline in RpoS+ viability was still severe upon reaching stationary phase, but the death rate was less pronounced than the parent strain, and the RpoS+ population did not reach extinction within 50 h (100–1,000 CFU mL^−1^; Fig. 7). Thus, RpoS likely plays a role in V. natriegens acid tolerance but cannot achieve the same level of acid tolerance as E. coli.
RpoS plays a minor role in V. natriegens acid tolerance. Culture turbidity (solid lines; OD660) was tracked in minimal media with 25 mM glucose over the entire experiment, whereas viable cell density (dotted lines; CFU mL−1) was tracked from the end of the growth phase (~12 h). Data points, mean; error bars, SD; n = 3. Some error bars are smaller than the data point symbols. Symbols with an “x” were below the detection limit of 102 CFU mL−1.
Lactic acid production prolongs growth and metabolism but does not improve survival
Organisms that produce multiple fermentation products can alter their fermentation profile to respond to environmental conditions. Unlike V. cholerae, V. natriegens lacks genes for producing neutral fermentation products (Table 1) (24). However, like E. coli, V. natriegens produced lactic acid in late exponential phase and into stationary phase (Fig. 2A and B). In E. coli, cytoplasmic lactate dehydrogenase (LDH) expression increases in response to acidic pH (29), suggesting that lactic acid production might decrease acid stress (30). A shift to lactic acid production over acetic acid production was also correlated with enhanced survival of pathogenic E. coli O157:H7 over an E. coli K-12 strain, likely by avoiding accumulation of cytoplasmic acetate anions (31). Shifting to lactic acid production would result in less ATP but also less acid overall to maintain electron balance (Fig. 8A) ; two pyruvate could either go to two lactic acid (pKa, 3.86) or to one ethanol + 2 formic acid (pKa, 3.75) + 1 acetate (pKa, 4.76). The latter pathway would generate more organic acids, one of which is acetic acid that would protonate more readily at low pH due to its high pKa. Acetic acid/acetate is also potentially more toxic than lactic acid/lactate in general (32).
*V. natriegens LDH extends fermentative metabolism but not viability. (A) Lactic acid production can hypothetically achieve electron balance while producing less acid (two lactic acid; left) via LDH compared to producing one ethanol, two formic acid, and one acetic acid (right). Comparison of late stationary-phase (24 h) fermentation product yields (B), glucose consumed (C), exponential growth rate (D), highest cell density measured as turbidity (E), and total organic acid yield (F) between the parent strain (WT Δdns::SpecR) and the LDH mutant (Δdldh Δdns::SpecR) in a minimal medium with 25 mM glucose without MOPS. For, formic acid; EtOH, ethanol; Lac, lactic acid; Ace, acetic acid; Suc, succinic acid; Pyr, pyruvate. (C, E, F) Exponential phase samples (9 h, Exp); Stationary-phase samples (24 h, St). (G) Turbidity (solid lines; OD660) and viable cell density (dotted lines; CFU mL−1) were tracked into stationary phase in a minimal medium with 25 mM glucose plus 100 mM MOPS. Data points, mean; error bars, SD; n = 3. The detection limit is 102 CFU mL−1. (B–F) Points, biological replicate values; error bars, SD; n = 3. Statistical differences were determined using unpaired, two-tailed t-tests; ns, not significant; *, P < 0.05; *, P < 0.01.
To test whether lactic acid production contributes to V. natriegens survival under fermentative conditions, we first created an LDH mutant. V. natriegens has three putative LDH genes, PN96_RS20515 (WP_020334824), PN96_RS17000 (dldh; WP_020334627), and PN96_RS17015 (lldh; WP_014233519). The latter two genes had been deleted by another group and had little effect on fermentative lactic acid production (10). However, the conditions in that study favored succinic acid production over lactic acid production, whereas we observed the opposite. Using the fermentative LDH gene from E. coli MG1655 (b1380) as a BLASTp query, we hypothesized that dldh was most likely the V. natriegens fermentative LDH (100% coverage, 62% identity); the other two candidates did not show significant sequence identity to the query sequence. Indeed, when we deleted V. natriegens dldh, the lactic acid yield was close to zero at 24 h (Fig. 8B). Compared to the parent strain, the Δdldh mutant produced 1.2 times more formic acid and 1.3 times more ethanol (Fig. 8B), a route that would replace the electron balancing role of lactic acid production (Fig. 8A). The Δdldh mutant also produced 2.8 times more pyruvate, suggesting that a bottleneck was created in the absence of LDH. The Δdldh mutant consumed less glucose than the parent (Fig. 8C), which did not affect the growth rate (Fig. 8D), but the final Δdldh mutant turbidity was 90% of that of the parent (Fig. 8E). Still, the total organic acid yield was not significantly different between the two strains, suggesting that even without lactic acid production, V. natriegens faced comparable acid exposure (Fig. 8F).
We thus examined the effect of the LDH deletion on V. natriegens stationary-phase survival under fermentative conditions. Anticipating that the rapid death rate would make a comparative analysis difficult, we added 100 mM MOPS to slow the death rate. Stationary-phase survival was similar between the parent and Δdldh strains (Fig. 8G). Thus, whereas, in the absence of MOPS, lactic acid production extends glucose consumption into stationary phase (Fig. 8C) and potentially allows for higher cell densities, at least as measured by turbidity (Fig. 8E), it does not improve stationary-phase survival under fermentative conditions with MOPS, wherein wild-type lactate production is elevated over other fermentation products (Fig. 6C).
Below we speculate why lactic acid might be produced during the transition out of exponential growth and into stationary phase. One possibility is that acetate and ethanol production becomes less available as an option. For example, as the pH drops and growth slows, the biosynthetic demand for ATP would also slow. This might limit ADP availability for acetate production and create a bottleneck at pyruvate (Fig. 8A). LDH offers a carbon and electron sink to maintain metabolic flow and help alleviate this bottleneck; a role for LDH in addressing this bottleneck would also explain the elevated pyruvate excretion in the Δdldh mutant (Fig. 8B). If we assume similar kinetic parameters to E. coli enzymes, it is possible LDH is poised to respond to pyruvate accumulation due to its increased expression and improved half-saturation constant for pyruvate relative to PFL at low pH (58–60). Characterization of the kinetic properties of these V. natriegens enzymes is needed to test this hypothesis.
Conclusion
As interest grows in V. natriegens as a bioprocessing strain, one must weigh its favorable attributes of rapid growth and genetic tractability against its less favorable attributes. Here we report that, relative to E. coli, V. natriegens is comparatively sensitive to the acidic conditions that arise during mixed acid fermentations. This difference in acid tolerance is likely due to V. natriegens lacking most of the acid resistance mechanisms that have been characterized in E. coli. Such acid-resistance mechanisms could potentially be engineered to improve V. natriegens acid tolerance. Increasing the buffering capacity of the fermentation medium would also avert acid sensitivity, but one would need to weigh the associated cost under industrial conditions. Thus, acid sensitivity will need to be considered when engineering V. natriegens for bioprocessing, unless the goal is to engineer V. natriegens to produce neutral products.
MATERIALS AND METHODS
Strains
Experiments used wild-type E. coli MG1655 (61) and the Vibrio natriegens ATCC 14048 (12) variant “Dalia, SAD1302, 2016,” which carries the following mutations: RpoS^E71*^, PN96_RS14445^S29P^ (putative cation solute symporter); Cultivarium Portal (https://portal.cultivarium.org/communities/vnat-sequencing?tab=data&dataset=Pilot%20study). V. natriegens mutants were made as described (12), except the transformation mixtures of cells and PCR products were incubated statically at 30°C for 12–16 h before outgrowth, and selection plates used 250 µg spectinomycin mL^−1^. The V. natriegens Δdldh and RpoS+ strains were made in V. natriegens TND1964, which is the wild-type strain with pMMB-tfoX that allows for IPTG-inducible competency (12). A Δdldh (ΔRS17000) construct was made by PCR amplifying (Q5 DNA polymerase; New England Biolabs) 3 kb upstream and downstream fragments. The two fragments were then combined by splicing-by-overlap extension PCR using the outermost primers. The RpoS+ construct was made using gDNA from a V. natriegens strain wherein rpoS was already repaired (Glasgo and Zinser, manuscript in prep). Each construct (1 µg) was then co-transformed into TND1964 with 50 ng of a DNA fragment containing a spectinomycin resistance cassette flanked by ~1 kb of homology to the upstream and downstream regions of the dns locus (PN96_RS00885) amplified from V. natriegens SAD1306 genomic DNA (12). The “parent” strain (NWH004) used as the control strain in experiments with the Δdldh mutant was derived from TND1964 by introducing a spectinomycin resistance cassette at the dns locus. Colonies were screened for RpoS activity by a positive colony bubble test using 30% H_2_O_2_ (62). Colonies were screened for Δdldh by PCR using OneTaq DNA polymerase (New England Biolabs). All mutations were confirmed by Sanger sequencing. Primers are listed in Table 2.
Growth conditions
Colonies were grown from 25% glycerol frozen stocks on agar plates of either lysogeny broth Miller medium (LB; E. coli), or LB supplemented with an additional 20 g NaCl L^−1^ (LB3; V. natriegens). LB3 was supplemented with either carbenicillin (100 µg mL^−1^), spectinomycin (250 µg mL^−1^), or 100 µM IPTG when appropriate. LB3 was also used for strain construction.
For all other experiments, anoxic minimal M9-derived coculture medium (MDC) (63) for E. coli or MDC with an additional 80 mM NaCl for V. natriegens (MDC80) was prepared by aliquoting 10 mL media into 27-mL anaerobic tubes or 60 mL media into 150-mL serum vials, bubbling with N_2_, then sealing with rubber stoppers and aluminum crimps before autoclaving. Media supplements were then added by syringe to the following final concentrations from anoxic 100× stock solutions: glucose, 25 mM; NH_4_Cl, 10mM; MgSO_4_, 1 mM; CaCl_2_, 0.1 mM. A 2 M MOPS solution was prepared by dissolving the free acid in ultrapure water and adjusting the pH to 7.0 using NaOH pellets. To make an anoxic stock, the solution was then filter-sterilized into a serum vial and flushed with filter-sterilized argon. This solution was added to cultures to the indicated concentration after autoclaving, with sterile ultrapure water also added where necessary to have common dilution effects between cultures. Starter cultures were inoculated into MDC or MDC80 from single colonies. Late-exponential phase starter cultures (0.65–0.85 OD_660_) were then used to inoculate test media to an initial cell density of ~0.01 OD_660_. Tubes were incubated horizontally at either 30°C (V. natriegens) or 37°C (E. coli) with shaking at 150 rpm (¾” stroke length).
Analytical procedures
A Genesys 20 spectrophotometer (Thermo Fisher) was used to measure cell density as turbidity at 660 nm (OD_660_) either directly in 27-mL culture tubes or in 1-mL samples in cuvettes when 150-mL serum vials were used; we chose 660 nm to be consistent with our previous studies involving V. natriegens and E. coli (38, 44). Growth rates were determined by fitting an exponential trendline to turbidity readings taken between 0.1 and 0.8 OD_660_ using GraphPad Prism v.6. Turbidity measurements were converted into molar biomass using a conversion factor of 270 (V. natriegens [64]) or 300 (E. coli [65, 66]) mg DCW L^−1^ OD_600_^−1^ and dividing by a molecular weight of 26.21 mg mmol^-1^ (V. natriegens [67]) or 24.97 (E. coli [68]) based on elemental compositions (67, 68).
Glucose and fermentation products were quantified in supernatant samples by HPLC (Shimadzu) as described (69). Temporal assessment of fermentations was performed in 60-mL cultures in 150-mL serum vials. Samples (1 mL) were taken at regular intervals; air contamination was minimized by flushing a 1-mL syringe and needle with filter-sterilized N_2_ before sampling. To quantify H_2_, 0.1 mL of headspace was sampled using a gas-tight syringe and injected into a Shimadzu GC-2014 gas chromatograph with a thermal conductivity detector as described (70). The H_2_ limit of detection was 12 nmol. To measure pH, cultures were centrifuged, and supernatants were assayed using a pH meter.
Viable cell counts were quantified as CFUs using the track plate method (71). Samples (0.4–0.5 mL) were removed from 10-mL cultures using an N_2_-flushed syringe and serially diluted in oxic MDC or MDC80 in a 96-well plate. The last six dilutions were spotted (10 µL) onto LB or LB3 agar using a multichannel pipette. A minimum of 1 colony was used as a cutoff, making the limit of detection 100 CFU mL^−1^ for an undiluted sample.
For experiments using nongrowing cells, late-exponential phase cultures (0.65–0.85 OD_660_) were inoculated into anoxic MDC or MDC80 with all supplements except glucose. The target inoculum was 10^7^ cells mL^−1^. Specified volumes of a solution containing 1 M acetic acid plus 1 M formic acid was then added by syringe. Cell suspensions were then sampled for CFU measurements. Death rates were determined from the slope of a semi-log equation (linear X, log Y) fitted to experimental data using GraphPad Prism v.6.
Bioinformatics
NCBI’s BLASTp (47) was used to identify putative acid-resistance genes in V. natriegens ATCC14048 (NCBI RefSeq: GCF_001456255.1) using amino acid sequences from E. coli MG1655 (NCBI RefSeq: GCF_000005845.2) or V. cholerae O1 biovar El Tor str. N16961 (GCF_000006745.1). V. natriegens proteins that had >25% identity with >50% query coverage were considered putative homologs.
Statistics
GraphPad Prism v.6 was used for all statistical analyses.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Epps TH III, Korley LTJ, Yan T, Beers KL, Burt TM. 2022. Sustainability of synthetic plastics: considerations in materials life-cycle management. JACS Au 2:3–11. doi:10.1021/jacsau.1c 0019135098218 PMC 8790729 · doi ↗ · pubmed ↗
- 2Liao JC, Mi L, Pontrelli S, Luo S. 2016. Fuelling the future: microbial engineering for the production of sustainable biofuels. Nat Rev Microbiol 14:288–304. doi:10.1038/nrmicro.2016.3227026253 · doi ↗ · pubmed ↗
- 3Mc Kinlay JB, Vieille C, Zeikus JG. 2007. Prospects for a bio-based succinate industry. Appl Microbiol Biotechnol 76:727–740. doi:10.1007/s 00253-007-1057-y 17609945 · doi ↗ · pubmed ↗
- 4Nielsen J. 2019. Cell factory engineering for improved production of natural products. Nat Prod Rep 36:1233–1236. doi:10.1039/c 9np 00005 d 30997457 · doi ↗ · pubmed ↗
- 5Xu X, Liu Y, Du G, Ledesma-Amaro R, Liu L. 2020. Microbial chassis development for natural product biosynthesis. Trends Biotechnol 38:779–796. doi:10.1016/j.tibtech.2020.01.00232029285 · doi ↗ · pubmed ↗
- 6Chi H, Wang X, Shao Y, Qin Y, Deng Z, Wang L, Chen S. 2019. Engineering and modification of microbial chassis for systems and synthetic biology. Synth Syst Biotechnol 4:25–33. doi:10.1016/j.synbio.2018.12.00130560208 PMC 6290258 · doi ↗ · pubmed ↗
- 7Volke DC, Orsi E, Nikel PI. 2023. Emergent CRISPR-Cas-based technologies for engineering non-model bacteria. Curr Opin Microbiol 75:102353. doi:10.1016/j.mib.2023.10235337413959 · doi ↗ · pubmed ↗
- 8Calero P, Nikel PI. 2019. Chasing bacterial chassis for metabolic engineering: a perspective review from classical to non-traditional microorganisms. Microb Biotechnol 12:98–124. doi:10.1111/1751-7915.1329229926529 PMC 6302729 · doi ↗ · pubmed ↗