Controlling GRF4‐GIF1 expression for efficient, genotype‐independent transformation across wheat cultivars
Sadiye Hayta, Mark A. Smedley, Meltem Bayraktar, Macarena Forner, Anna Backhaus, Clare Lister, Martha Clarke, Cristobal Uauy, Simon Griffiths

TL;DR
A new wheat transformation system enables efficient gene editing across different wheat types without causing harmful side effects.
Contribution
A genotype-independent transformation system with controlled GRF4-GIF1 expression minimizes pleiotropy while enabling broad cultivar compatibility.
Findings
GRF4-GIF1 expression improved transformation efficiency across multiple wheat cultivars.
Tissue-specific promoters and heat-inducible excision reduced pleiotropic effects like reduced fertility.
The system supports functional genomics and gene editing for improved crop traits.
Abstract
Wheat is a staple crop critical for global food security, and its continuous genetic improvement is essential to meet the demands of a growing population. Efficient, genotype‐independent transformation is a major bottleneck in wheat functional genomics and gene editing. The growth regulating factor (GRF)–GRF‐interacting factor (GIF) fusion technology enhances regeneration efficiency and broadens the range of transformable cultivars, but constitutive expression can reduce fertility and spikelet number. Here, we present an optimised Agrobacterium‐mediated wheat transformation protocol incorporating GRF4‐GIF1, tested across multiple tetraploid and hexaploid cultivars. Transformation efficiency was improved through adjustments in selection pressure, zeatin concentration, and promoter choice, with GRF4‐GIF1 consistently enabling successful transformation across genotypes. Tissue‐specific…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1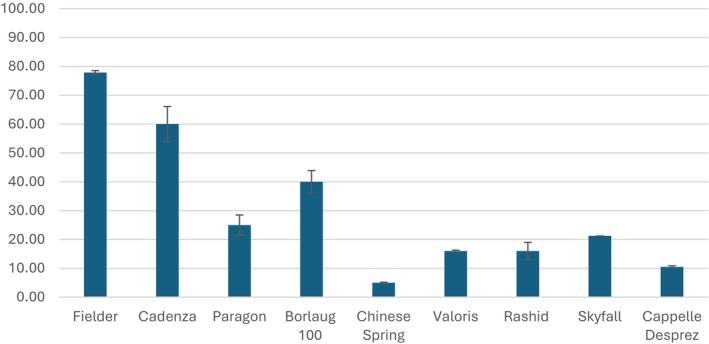 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4| Genotype name | Type of wheat | Full genome sequence | Transformed previously |
|---|---|---|---|
| Fielder | US spring wheat | Yes | Yes |
| Cadenza | UK alternative wheat | Yes | Yes |
| Paragon | UK spring wheat | Yes | No |
| Reedling (Borlaug 100) | CIMMYT variety | Yes | No |
| Chinese Spring | Chinese spring wheat variety | Yes | No |
| Skyfall | Group 3 winter wheat | No | No |
| RGT Rashid | Group 3 winter wheat | No | No |
| Valoris | French winter wheat | No | No |
| Capelle Desprez | UK winter wheat | No | No |
| OmRabi5 | ICARDA spring durum wheat | No | No |
| CIRNO‐C | CIMMYT spring durum wheat | No | No |
| Svevo | Spring durum wheat | Yes | No |
| Kronos | Spring durum wheat | Yes | Yes |
| Wheat cultivar | GRF4‐GIF1 | S1 Hygromycin (mg/L) | S2 Hygromycin (mg/L) | Regeneration hygromycin (mg/L) | Zeatin (mg/L) |
|---|---|---|---|---|---|
| Fielder | − | 20 | 30 | 20 | 2.5 |
| Fielder | + | 15 | 15 | 15 | 1.0 |
| Kronos | − | 15 | 15 | 15 | 2.5 |
| Kronos | + | 15 | 15 | 15 | 2.5 |
| Cadenza | − | 10 | 10 | 10 | 1.0 |
| Cadenza | + | 10 | 10 | 10 | 1.0 |
| Skyfall | + | 10 | 10 | 10 | 2.5 |
| Reedling | + | 10 | 10 | 10 | 1.0 |
| Cappelle Desprez | + | 10 | 10 | 10 | 1.0 |
| Paragon | + | 5 | 5 | 5 | 1.0 |
| Valoris | + | 5 | 5 | 5 | 1.0 |
| Chinese Spring | + | 5 | 5 | 5 | 2.5 |
| Construct name | Promoter driving GRF4‐GIF1 | Reason for choosing | Marker gene | Addgene number |
|---|---|---|---|---|
| pGGG ZmUbi GRF4‐GIF1 | ZmUbi:: | Strong promoter | OsUbi:: | # 246478 |
| pGGG OsAct GRF4‐GIF1 | OsAct:: | Medium promoter | OsUbi:: | # 246479 |
| pGGG 35S GRF4‐GIF1 | CaMV35S:: | Weak promoter | OsUbi:: | # 246480 |
| pGGG HSCre GRF4‐GIF1 | Zm | Excisable via Heat Shock | OsUbi:: | # 246481 |
| pGGG PLTP GRF4‐GIF1 | Zm | Tissue‐specific promoter | OsUbi:: | # 246482 |
| pGGG GRF5‐GIF1 | Zm | Strong promoter | OsUbi::G | # 246483 |
| pGGG GUS | Non‐GRF4‐GIF1 Control | Control | OsUbi:: | # 165418 |
| pSoup HSCre GRF4‐GIF1 | Zm | Excisable via Heat Shock | N/A | # 246484 |
| Variety | Construct | Main seeds ± SE | Weight(g) ± SE | TGW(g) ± SE | ØArea ± SE | ØWidth ± SE | ØLength ± SE | Tillers ± SE | Height (cm) ± SE | Spike number ± SE |
|---|---|---|---|---|---|---|---|---|---|---|
| Fielder | Control | 448.24 ± 25.5a | 21.44 ± 1.2a | 48.48 ± 0.7b | 16.25 ± 0.2b | 3.44 ± 0.02b | 6.49 ± 0.06c | 14.67 ± 0.8b | 85.52 ± 1.1a | 14.62 ± 0.8b |
| HS Cre | 324.61 ± 19.2b | 16.05 ± 0.8b | 50.54 ± 0.9b | 17.16 ± 0.3b | 3.49 ± 0.03b | 6.75 ± 0.05b | 16.44 ± 0.7ab | 81.42 ± 1.2a | 16.12 ± 0.7ab | |
| Wox5 | 231.79 ± 27.9c | 14.11 ± 1.5b | 63.00 ± 2.0a | 20.44 ± 0.5a | 3.90 ± 0.06a | 7.09 ± 0.07a | 11.26 ± 0.7c | 84.84 ± 2.4a | 11.00 ± 0.7c | |
| PLTP | 369.13 ± 18.4ab | 17.28 ± 0.7b | 48.25 ± 1.0b | 16.41 ± 0.3b | 3.42 ± 0.03b | 6.62 ± 0.06bc | 18.50 ± 0.6a | 83.19 ± 1.0a | 18.22 ± 0.6a | |
| Kronos | Control | 157.00 ± 11.7a | 8.70 ± 0.6a | 56.22 ± 1.4a | 19.59 ± 0.3a | 3.59 ± 0.04b | 7.70 ± 0.05a | 15.33 ± 0.9b | 66.88 ± 0.9a | 15.04 ± 0.9bc |
| HS Cre | 45.50 ± 13.0b | 2.48 ± 0.7b | 54.87 ± 1.1a | 20.41 ± 0.5a | 3.66 ± 0.04ab | 7.85 ± 0.11a | 15.10 ± 11.15b | 61.25 ± 1.34bc | 14.70 ± 1.3c | |
| Wox5 | 24.23 ± 6.2b | 1.41 ± 0.4b | 52.73 ± 2.4a | 20.77 ± 0.6a | 3.76 ± 0.05a | 7.83 ± 0.13a | 11.92 ± 1.1b | 60.15 ± 1.78c | 10.31 ± 1.1d | |
| PLTP | 64.05 ± 14.8b | 3.88 ± 0.9b | 57.87 ± 1.2a | 20.19 ± 0.3a | 3.61 ± 0.03ab | 7.80 ± 0.08a | 23.95 ± 1.3a | 65.15 ± 1.0ab | 23.75 ± 1.3a |
- —Biotechnology and Biological Sciences Research Council10.13039/501100000268
- —the Scientific and Technological Research Council of Turkey10.13039/501100004410
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant tissue culture and regeneration · Wheat and Barley Genetics and Pathology · Plant Molecular Biology Research
INTRODUCTION
Bread wheat (Triticum aestivum L.), also known as common wheat, is one of the “big three” cereal crops worldwide Shewry (2009), with its extensive cultivation range and vital role as the primary source of cereal‐based processed products. Wheat significantly contributes to global food security, providing approximately 20% of the world's caloric intake and 25% of daily protein consumption. Furthermore, wheat is an important source of essential minerals for human nutrition and contributes up to 20% of essential dietary minerals in the UK (Sigalas et al., 2024). Durum wheat (Triticum turgidum L. ssp. durum), known for its high protein content, golden colour, and firm texture, serves as an important food source in certain regions; its production accounts for approximately 7% of total global wheat production, and about 75% of worldwide durum wheat production is used by the pasta industry (Grosse‐Heilmann et al., 2024).
Improving wheat genetics to enhance its nutritional value, disease resistance, and resilience to climate change will have a profound impact on sustainable food production and global nutritional security. The advent of advanced technologies, including the development of multiple reference‐quality genome assemblies, has ushered in a new era for wheat research, equipping researchers and breeders with vital tools to enhance wheat and address future food security challenges (Walkowiak et al., 2020). These genome assemblies provide a foundation for functional gene discovery and breeding, facilitating the development of the next generation of modern wheat cultivars. The availability of these genomic sequences, together with new open‐access transformation protocols (Hayta et al., 2019), videos, and plasmid repositories, will enable the powerful application of molecular tools in future wheat research and breeding. Effective transgenic methodologies are now crucial for conducting gene functional studies, enhancing traits, and integrating precision breeding techniques.
Wheat transformation is highly genotype‐dependent and typically relies on a few cultivars, such as Fielder and Bobwhite, which exhibit a strong tissue culture response (Hayta et al., 2019). Despite the first successful reports of Agrobacterium‐mediated wheat transformation in the 1990s (Cheng et al., 1997), transformation efficiencies remained low, around 5%, for many years. An in planta method was reported in 2009 (Risacher et al., 2009), but it was not widely adopted. Later, Japan Tobacco Inc. demonstrated improved transformation efficiency in the model wheat genotype Fielder using a patented, licensed system called PureWheat, which requires specialist training and specific vectors, thereby limiting its accessibility (Ishida et al., 2015).
Our first wheat CRISPR study, on the ZIP4‐B2 (Ph1 locus), a meiotic gene involved in crossover within wheat (Rey et al., 2018), led to the publication of our freely available, efficient, and reproducible transformation protocol in 2019, achieving transformation efficiencies averaging around 25% in Fielder, and also Kronos and Cadenza with lower efficiencies of 10% and 4%, respectively (Hayta et al., 2021). Kronos and Cadenza are important reference varieties, with extensive TILLING resources available for gene discovery and complementation (Krasileva et al., 2017). The key factors influencing wheat transformation include the cultivar used, the quality and health of the donor material, the developmental stage of the immature embryos, the handling of the material, and the media component composition; transformation efficiencies are drastically affected by multiple factors, with these narrow optimal windows (Hayta et al., 2021).
The genotype independence of transformation techniques further broadens the potential for genetic improvements, making wheat breeding more versatile and accessible. Debernardi et al. (2020) developed a growth regulator fusion technology involving growth regulating factor 4 (GRF4) and its cofactor GRF‐interacting factor 1 (GIF1). Overexpressing GRF4‐GIF1 in wheat significantly boosts regeneration efficiency and broadens the range of transformable wheat genotypes. We tested this technology using our published wheat transformation method. Fielder plants transformed with the GRF4–GIF1 chimera increased dramatically to 77.5% efficiency (Debernardi et al., 2020). This technology accelerates the transformation pipeline and enables high transformation efficiencies by overcoming narrow optimal windows, such as specific growing conditions and embryo developmental stages, which are critical for successful transformation in wheat.
Low transgene copy number GRF4‐GIF1 transgenic plants exhibited normal development and fertility in Fielder; however, constitutive expression of other plant developmental regulators such as the maize Baby boom (ZmBbm), the maize Wuschel2 (ZmWus2) (Johnson et al., 2023), and a Wuschel homologue TaWox5 (Wang et al., 2022) can lead to negative pleiotropic effects, necessitating their removal from transgenic plants and limiting their practical application (Wang et al., 2020). In an effort to minimise the negative pleiotropic effects associated with developmental regulators, Lowe et al. (2016) demonstrated a strategy for successful plant regeneration using Cre recombinase, which excised morphogenic gene sequences flanked by loxP sites. This approach was critical in maize transformation, removing unwanted expression cassettes (Bbm and Wus2) linked to morphological abnormalities including reduced root elongation and ensuring normal plant development. Further advancements were made by Lowe et al. (2018) who identified the ZmPLTP (maize phospholipid transferase protein gene), a maize promoter with strong expression in leaves, embryos, and callus, but downregulated in roots, meristems, and reproductive tissues. This promoter was used to drive Bbm expression in maize, enabling efficient somatic embryo formation without the callus phase and rapid plant regeneration. The ZmPLTP allowed for uniform transformation and successful regeneration of maize plantlets. Co‐transformation has been widely used because it is a simple and clean technique, leaving no residual DNA sequences, such as inverted repeats and recombination sites, in transgenic plants from which the selectable marker gene has been eliminated with high frequency. Co‐transformation involves the simultaneous integration of a selectable marker gene and a gene of interest from different T‐DNAs, followed by their subsequent recombination and segregation in the progeny, provided the two genes are integrated into unlinked loci (Liu et al., 2020).
Debernardi et al. (2020) demonstrated that transgenic plants overexpressing the GRF4‐GIF1 chimera under the control of the ZmUbi promoter maintained fertility and morphology. However, these plants exhibited a 23.9% reduction in grain number per spike and a 13.7% increase in grain weight, suggesting that GRF4‐GIF1 can modulate physiological traits without drastically impairing plant development. In our study, high transgene copy numbers of GRF4‐GIF1 were associated with fertility depending on the cultivar but also showed a modest positive effect on grain area and grain length. For effective gene function characterisation, it is critical to develop transgenic plants that exhibit desirable traits while minimising adverse effects linked to morphological gene overexpression. The GRF4–GIF1 technology addresses genotype dependence and expands the applicability of transformation methods, including gene editing tools, across elite wheat cultivars. Moreover, the ability to segregate or remove GRF4–GIF1 after transformation provides enhanced flexibility for downstream applications, such as functional genomics, gene editing, and precision breeding. Importantly, GRF4–GIF1 technology, together with open protocols, further democratises gene editing by making these approaches increasingly accessible to research centres and wheat breeders lacking high‐end facilities.
Despite the efficiency gains offered by GRF4–GIF1, its constitutive expression can lead to pleiotropic effects, including reduced fertility, spikelet number, and altered plant architecture, which vary across genotypes. Promoter strength, temporal activity, and construct design, as well as media composition such as zeatin and hygromycin levels, can influence both transformation efficiency and phenotypic outcomes. Additionally, we propose strategies to limit the temporal activity of GRF4–GIF1, such as using a PLTP tissue‐specific promoter, excision of GRF4–GIF1, and co‐transformation with two Agrobacterium plasmids, one carrying the GRF4–GIF1 expression cassette and the other carrying both the selectable marker and the gene of interest (GOI). These strategies were tested across multiple wheat cultivars to mitigate undesirable pleiotropic effects, such as reduced grain number. Together, these refinements enhance the accuracy and reliability of functional analyses for trait‐associated genes, enabling efficient wheat transformation while maintaining developmental stability.
RESULTS AND DISCUSSION
GRF transcription factors and physiological impact
GRF transcription factors are highly conserved across angiosperms, gymnosperms, and mosses, encoding proteins with conserved QLQ (Gln, Leu, and Gln) and WRC (Trp, Arg, and Cys) domains essential for protein–protein and protein–DNA interactions. In many plant lineages, GRFs are regulated by microRNA miR396, which downregulates GRF expression in mature tissues (Debernardi et al., 2014).
We first tested the ZmUbi GRF4‐GIF1 construct with nine hexaploid wheat cultivars (Table 1), including several that had previously been considered recalcitrant to transformation. A construct with GUS reporter driven with OsUbi promoter used as the control. For GRF4‐GIF1 transformations, 70–100 immature embryos were used per cultivar, and 50–80 embryos were used for controls. Calli transformed with ZmUbi::GRF4‐GIF1, successfully produced somatic embryos and regenerated plantlets (Figure 1; Figure S1). using ZmUbi::GRF4‐GIF1, which successfully produced somatic embryos and regenerated plantlets (Figure 1; Figure S1). These results are consistent with Debernardi et al. (2020), who reported that the GRF4‐GIF1 chimera promotes embryogenesis, shoot proliferation, or both in wheat, thereby improving transformation efficiency, particularly in recalcitrant cultivars.
Regeneration of wheat calli using the ZmUbi GRF4‐GIF1 construct across different cultivars.Representative regeneration responses are shown for the wheat cultivars Fielder, Reedling (Borlaug 100), Cadenza, Chinese Spring, and Skyfall, respectively.
In wheat, in vitro regeneration can proceed via two main pathways: somatic embryogenesis, in which a single somatic cell from embryos develops into complete plants, and organogenesis, where shoots or roots are initiated from callus or directly from explants. Somatic embryogenesis is considered the more fundamental mode of regeneration because it produces progeny with high genetic stability and reduced somaclonal variation. This pathway is particularly important in wheat, as it enables the transformation and regeneration of genotypes that are otherwise recalcitrant to in vitro culture (Kowalik et al., 2026).
In our system, transformation with ZmUbi::GRF4‐GIF1 primarily promotes somatic embryogenesis, generating multiple embryos from calli delivered from immature embryo explants which subsequently develop shoots and fully regenerated plantlets. Enhancing embryogenic competence with GRF4‐GIF1 is critical for cultivars that normally exhibit low regeneration capacity, as it increases the number of transformable cells and improves overall transformation efficiency.
Our construct carries both the hygromycin selectable marker and the GRF4‐GIF1 chimera on the same T‐DNA. Therefore, the transgene copy number, as measured by Hyg integration, reflects the number of GRF4‐GIF1 copies present in each transformed line. Data from mature plants indicated that the degree of fertility loss associated with GRF4‐GIF1 expression varied among cultivars (Figure S2). Lines with higher transgene copy numbers exhibited a slightly increased proportion of sterile plants, although the severity of this effect depended on the specific cultivar. These results demonstrate a clear relationship between the number of integrated T‐DNA copies and developmental outcomes, providing a mechanistic explanation for observed differences in fertility across genotypes.
Among the cultivars tested, Kronos had the lowest seed set, indicating the greatest sensitivity to GRF4‐GIF1 whereas Paragon and Cadenza were less affected (Figure S1). GRF4‐GIF1 had a slightly positive effect on grain area and grain length across all cultivars, consistent with findings by Debernardi et al. (2020) who reported a 23.9% reduction in grain number per spike and a 13.7% increase in grain weight. A significant increase in the number of spikes was observed in Fielder (26.1 spikes per plant) compared with the control (11.8 spikes per plant).
Transformation efficiency and phenotypic effects in hexaploid wheat
We first optimised wheat transformation using pGGG constructs containing the PvUbi promoter driving the hygromicin selection gene with the CAT1 intron, and the rice ubiquitin promoter driving a GUS reporter gene containing two introns in Fielder, Reedling (Borlaug 100), Cadenza, Chinese Spring, and Skyfall. Using the same selection system, we tested a construct in which the ZmUbi promoter drives GRF4‐GIF1 to assess its effect on wheat transformation efficiency. After the initial callus induction stage, cultures grown under light conditions initiated a significantly greater amount of shoot formation (Figure 1) compared with those transformed with the control construct. Both shoot and root development appeared normal, producing an average of 3.2 plantlets per embryo. All regenerated plantlets tested positive for the transgene.
Transformation efficiencies with this construct reached 77.5% in Fielder, 60% in Cadenza, and 40% in Borlaug 100. In winter, wheat cultivars such as Valoris and Skyfall, efficiencies were 16 and 20%, respectively (Figure 2). Notably, multiple transgenic plants were often regenerated from a single embryo, and approximately 80% of these plants exhibited a copy number difference greater than three compared with each other (73 out of 91 plants). This indicates that the plantlets likely arose from independent transformation events. This suggests that the actual transformation efficiency may be even higher than reported. When the calculated on the basis independent regenerants per embryo, transformation efficiency in Fielder reached 98.3%. Consistent with the findings of Johnson et al. (2023), who reported that 84% of transgenic plantlets regenerated from the same embryos originated from independent transformation events. Only Fielder and Cadenza were amenable to transformation without the GRF4‐GIF1 fusion, albeit with substantially lower efficiencies, 25 and 5% respectively.
Transformation efficiencies in various wheat cultivars.Transformation efficiencies achieved using different GRF4‐GIF1 constructs across multiple wheat cultivars, including elite and commercial lines.
This advancement has enabled successful transformation across a broader range of wheat genotypes, including elite and commercial cultivars, such as Borlaug 100, RGT Rashid, Skyfall, and Valoris. Among these, Skyfall is the most widely cultivated hexaploid bread making wheat in the UK, Valoris is a French variety notable for its high pentosan content, and Borlaug 100, developed by CIMMYT, serves as a key international breeding line (Table 1). Across all tested wheat cultivars, GRF4‐GIF1 constructs consistently enabled successful transformation, although efficiency varied depending on the genotype and construct used. Skyfall, RGT Rashid, Valoris, and Cappelle Desprez are winter wheat varieties, and as such, are traditionally considered more recalcitrant to transformation than spring varieties; these varieties all showed lower transformation efficiencies (Figure 2). Given its consistent ability to enhance regeneration across diverse cultivars that could not be transformed without GRF4–GIF1, it can serve as a robust positive selection tool in wheat transformation experiments.
The growth of donor material is one of the main bottlenecks in wheat transformation. Donor materials are grown in controlled environment rooms (CERs). When plants are not healthy, transformation efficiency decreases across all cultivars. The GRF4–GIF1 system enables transformation of all tested wheat cultivars regardless of factors, such as donor material quality, plant health, or the developmental stage of immature embryos. Additionally, it shortens the overall transformation timeline around 2 weeks as reported by Debernardi et al. (2020).
We observed high transgene copy numbers (greater than 10), which were associated with the phenotypic effects such as reduced seed number (Figure S1) when GRF4‐GIF1 was driven by the ZmUbi promoter; any escapes were observed scarcely. Accordingly, we reduced the hygromycin concentration to promote reduced transgene copy number and lowered zeatin levels when GRF4‐GIF1 was included in the construct (Table 2).
Effects on fertility and spike morphology
Analysis of mature plants revealed that GRF4–GIF1 insertion significantly reduced fertility across all cultivars (P < 0.001) (Figure S1). Lines with higher transgene copy numbers exhibited slightly greater sterility, with severity varying by cultivar. Among the cultivars tested, Kronos exhibited the lowest seed number, indicating the highest sensitivity to GRF4–GIF1, while Paragon and Cadenza were less affected.
Rudimentary basal spikelets (RBS) were more frequent in Cadenza and Skyfall lines carrying GRF4–GIF1, whereas other cultivars were largely unaffected. Total spikelet number and total spike number were significantly reduced across genotypes (P < 0.001) (Figure S2).
Effects on yield‐related traits
The effect of GRF4–GIF1 on thousand‐grain weight (TGW) was genotype‐dependent. Kronos experienced a negative impact, Chinese Spring showed a slight reduction, and Fielder displayed a positive effect. Across all cultivars, average grain area was slightly increased (P < 0.001) (Figure S2), consistent with previous observations (Debernardi et al., 2020).
Effects on tiller number and plant architecture
GRF4–GIF1 significantly increased tiller number (10 versus 15) in Fielder (P = 0.0014), with other genotypes showing similar, though non‐significant, trends. The number of spikes per plant (10 versus 15) was significantly increased in Fielder (P = 0.0015). Conversely, plant height was negatively affected across all genotypes (P < 0.001), with the magnitude of reduction varying among cultivars (Figure S2).
GRF4–GIF1 expression induces a trade‐off between reproductive and vegetative traits. While spikelet fertility and total spikelet number are reduced, certain cultivars, such as Fielder, Cadenza, Paragon, and Reedling exhibit increased grain size, tiller number, and spike number, indicating cultivar‐specific compensatory effects (Figure S2). These findings underscore the critical role of promoter selection and genetic background in balancing high transformation efficiency with phenotypic stability.
In gene editing applications, segregation of the T‐DNA in subsequent generations typically eliminates these pleiotropic effects, enabling recovery of normal, edited lines. For transformation experiments targeting sensitive developmental traits, careful control of transgene expression is necessary to minimise unintended phenotypic consequences.
Extending GRF4‐GIF1 to tetraploid wheat using different promoters
Based on our initial observations indicating increased sensitivity of the tetraploid wheat cultivar Kronos to ZmUbi‐driven GRF4‐GIF1 overexpression, manifested by partial sterility and recognising that ZmUbi is a strong constitutive promoter, we developed a series of constructs designed either to modulate GRF4‐GIF1 expression using alternative promoters or to facilitate its excision following transformation (Table 3). These constructs allowed us to investigate strategies for minimising phenotypic changes while maintaining high transformation efficiency, thereby tailoring the system to the specific requirements of tetraploid wheat.
To broaden the scope of our analysis beyond Kronos, for which a TILLING mutant population is available, we evaluated these constructs in several additional tetraploid wheat cultivars, including CIRNO‐C (developed by CIMMYT), Om Rabi 5 (from ICARDA), and Svevo, an Italian durum wheat cultivar with full genomic sequence available.
Transformation efficiencies varied markedly among the tetraploid wheat cultivars tested. The Italian durum wheat variety Svevo exhibited the highest efficiency (55%) when transformed with the ZmUbi::GRF4‐GIF1 construct. Kronos, CIRNO‐C, and Om Rabi showed lower efficiencies of 50, 11, and 4%, respectively. In the absence of GRF4‐GIF1, only Kronos and Svevo could be transformed, and at much lower frequencies (2 and 15%, respectively).
The OsAct::TaGRF4‐GIF1 construct produced a more moderate effect (approximately 4%) but still performed better than the weak CaMV35S promoter and than the excisable and tissue‐specific promoter systems in both Kronos and CIRNO‐C. In contrast, Om Rabi and Svevo were successfully transformed using tissue‐specific and excisable promoters, reaching efficiencies of 4 and 2% in Om Rabi, and 15 and 19% in Svevo, respectively.
Overall, transformation outcomes depended on both the cultivar and the construct used. Notably, the highest efficiencies across all tetraploid lines were consistently obtained when the GRF4‐GIF1 fusion was driven by the strong ZmUbi promoter, highlighting the importance of promoter strength in optimising GRF4‐GIF1‐mediated transformation.
Zeatin concentration and regeneration
In many plant transformation systems, cytokinins are essential for shoot regeneration, with zeatin being the preferred cytokinin in wheat transformation. Previous high‐efficiency protocols have typically used zeatin at 5 mg L^−1^ (Ishida et al., 2015). In our study, Svevo, which exhibited greater regeneration capacity among all tested tetraploid wheat cultivars, achieved a transformation efficiency of 55% using the GRF4–GIF1 construct with only 1.0 mg L^−1^ zeatin, indicating that the GRF4–GIF1 fusion can promote shoot initiation and/or regeneration without the need for high zeatin concentrations. This finding aligns with Debernardi et al. (2020), who also reported high transformation efficiencies in Fielder without the addition of exogenous cytokinin. Svevo maintained a moderate transformation efficiency of 15% when transformed with the control construct that does not contain GRF4–GIF1, while Kronos showed 8.5% efficiency under the same conditions. In contrast, cultivars such as CIRNO‐C and Kronos demonstrated improved transformation outcomes when cultured with a higher zeatin concentration of 2.5 mg L^−1^, with Kronos reaching 50% efficiency. Notably, CIRNO‐C could not be transformed without the GRF4–GIF1 construct; = All cultivars produced more plantlets per immature embryo when cultured with 2.5 mg L^−1^ zeatin (11–21 plantlets/embryo) compared with lower zeatin levels (1–17 plantlets/embryo). In particular, CIRNO‐C (21 plantlets/embryo) and Om Rabi 5 (11 plantlets/embryo) benefited most from the higher zeatin concentration, as their transformation efficiencies were lower; the additional zeatin helped increase the number of transgenic plantlets. In contrast, Kronos (2%–50% transformation efficiency) and Svevo (3%–55% transformation efficiency) already exhibited high regeneration rates, so culturing them with 1.0 mg L^−1^ zeatin was sufficient to produce a large number of plantlets from each embryo.
These results highlight the genotype‐specific hormonal requirements and underscore the critical role of GRF4‐GIF1 in enabling the transformation of otherwise recalcitrant tetraploid wheat lines.
Further controlling
GRF4‐GIF1 expression to minimise phenotypic effects
To mitigate undesirable phenotypic effect described above, particularly sterility associated with high transgene copy numbers, we considered the regulatory role of microRNA miR396, which naturally downregulates GRF expression in mature tissues (Debernardi et al., 2020). This effect may be more pronounced in tetraploid wheat, which could have a lower miR396‐mediated regulatory capacity compared with hexaploid cultivars.
We hypothesise that a high copy of GRF4–GIF1 gene can overwhelm the endogenous miR396 pool, resulting in excessive GRF activity in mature plants and contributing to pronounced phenotypic effects. Vandeputte et al. (2024) observed a similar phenomenon in maize, where male sterility correlated with higher copy number variation in ZmGRF4–GIF1 and TaGRF4–GIF1 T_0_ transgenic plants.
To address this, we employed two complementary strategies. First, tissue‐specific expression using the non‐constitutive ZmPLTP promoter restricts GRF4–GIF1 activity to embryos and callus during transformation, minimising expression in mature plants. This promoter has been successfully used in maize for similar purposes (Lowe et al., 2018). Second, we applied a heat‐inducible Cre/lox excision system to remove the GRF4–GIF1 cassette post‐regeneration. This allows excision of GRF4–GIF1 after plant regeneration, reducing expression during later developmental stages and thereby mitigating strong phenotypic effects. Similar Cre/lox‐based excision strategies with ZmBbm and ZmWus2 have been reported in maize (Wang et al., 2020).
These strategies were tested in both the hexaploid cultivar Fielder and the tetraploid cultivar Kronos, demonstrating that controlled expression can maintain high transformation efficiency while substantially reducing deleterious phenotypes. The need for construct redesign in tetraploid wheat arises from the combined effects of promoter choice, copy number, and differences in miR396‐mediated regulation between hexaploid and tetraploid genotypes. By explicitly linking these factors, we provide a mechanistic rationale for the observed genotype‐specific phenotypic outcomes and for the modifications made to optimise the system across wheat ploidy levels.
Tissue‐specific expression using the non‐constitutive
ZmPLTP promoter driven GRF4‐GIF1
In Fielder, the ZmUbi::GRF4‐GIF1 construct delivered the highest transformation efficiency at 62% calculated as 1 plant per embryo, and producing an average of 3.1 plantlets per embryo. By contrast, the ZmPLTP GRF4‐GIF1 construct achieved a transformation efficiency of 20%, generating 1.7 plantlets per embryo with a low escape rate of 0.4%. The ZmUbi::GRF4–GIF1 construct resulted in a 33.6% transformation efficiency, whereas ZmPLTP::GRF4–GIF1 yielded a 23% efficiency and was associated with improved seed number (Table 4). Kronos was most sensitive to the ZmUbi::GRF4–GIF1 construct, which resulted in 46% sterility. In contrast, expression of GRF4–GIF1 under the PLTP promoter caused only a moderate reduction in main seed number, an increase in spike number, and tiller number while TGW remained comparable to the control (57.87 g vs. 56.22 g in non‐transgenic plants), while maintaining fertility in approximately 85% of the transgenic plants, highlighting its suitability for minimising phenotypic disruption in sensitive genotypes. Also in Fielder, main seed number and TGW were improved.
Grain size parameters remained largely unchanged, indicating that PLTP‐driven expression supports phenotypic stability while enhancing transformation efficiency. This highlights PLTP as a suitable promoter for minimising adverse developmental effects in both Fielder and Kronos.
These results demonstrate that the promoters driving GRF4–GIF1 expression strongly influence transformation efficiency. While stronger promoters enhance transformation efficiency, they also result in more undesirable phenotypic effects (Figure 3). In contrast, tissue‐specific expression driven by the ZmPLTP promoter enables efficient transformation while minimising associated fertility problems.
Phenotype of Kronos PLTP GRF4‐GIF1 and non‐transformed control plants (right).Transgenic Kronos plants expressing the ZmPLTP GRF4‐GIF1 construct showed phenotypes closely resembling non‐transformed controls, indicating minimal pleiotropic effects from transgene expression.
Excision of
GRF4‐GIF1 via heat‐induced Cre‐lox system
Continued expression of morphogenic genes can interfere with downstream functional analysis and characterisation of trait genes. HS Cre‐Lox excision system can reduce the number of morphogenic genes inserted and improve plant development and fertility.
Using the HS Cre‐Lox system, transformation efficiencies reached 30% in Fielder and 15% in Kronos. Successful GRF4–GIF1 excision was achieved in 11 of 15 plants (73%) in Kronos, whereas excision in Fielder occurred in 8 of 20 plants (13%). In both Fielder and Kronos, the HS Cre‐Lox system reduced seed number and total seed weight compared with non‐transgenic controls (Table 4), with the effect more pronounced in Kronos than in Fielder; however, TGW remained stable, indicating the production of fewer but normally sized grains. Extending the heat shock to 24 h slightly improved TGW but did not restore seed number. Notably, fertility in Kronos was markedly increased to 87%. Overall, the HS Cre‐Lox system helped restore fertility in both cultivars, with a more pronounced effect in Kronos.
Plant height was slightly reduced in Kronos, while in Fielder it remained similar to the control. Spike number increased moderately in Fielder but showed minimal change in Kronos. These results suggest that the HS Cre‐Lox system can achieve transgene excision but may still induce moderate fertility effects, especially in sensitive genotypes.
These outcomes are consistent with excision‐based improvements previously reported in maize (Wang et al., 2020), potentially reducing the copy number of ZmBBM and ZmWUS2 from the maize genome and thus eliminating the detrimental ectopic expression even in plants still retaining some morphogenic gene sequences (Aesaert et al., 2022; Johnson et al., 2023). This technology has been extended to wheat where a higher copy number of ZmBbm and ZmWus2 resulted in developmental defects. When using the heat shock inducible moCRE gene (Zm‐Hsp17.7 _ pro _ :moCRE), all the transgenic plants were fertile, further supporting the utility of this system in mitigating GRF4‐GIF1 phenotypic associated effects.
Co‐transformation
We explored co‐transformation as a method to introduce both GRF4‐GIF1 and other GOIs into wheat. By using a two‐Agrobacterium system with separate T‐DNAs for GRF4‐GIF1 and the GOI, we achieved transformation efficiencies comparable to those obtained with the ZmUbi GRF4‐GIF1 construct. This approach offers greater flexibility and the potential for segregation of GRF4‐GIF1 and GOI in subsequent generations.
We applied this co‐transformation strategy to several elite wheat cultivars, including Borlaug 100, Skyfall, Kronos, and Cadenza. These cultivars could not be transformed or showed very low transformation efficiencies when GRF4‐GIF1 was not included. However, when constructs containing both the GOI with a selectable marker and a separate GRF4‐GIF1 expression cassette were introduced, regeneration efficiency increased markedly, resulting in a substantial improvement in overall transformation efficiency.
This strategy was applied to several elite wheat cultivars (Borlaug 100, Skyfall, Kronos, and Cadenza) that were previously recalcitrant or showed very low transformation efficiency. Inclusion of GRF4‐GIF1 significantly increased regeneration efficiency and overall transformation success, similar in concept to the “altruistic transformation” system described in maize, where transient expression of Wus2 promotes somatic embryo formation while the selectable marker T‐DNA integrates stably (Hoerster et al., 2020).
Comparison with other morphogenic gene
We also tested the wheat homologue of Arabidopsis WUSCHEL‐RELATED HOMEOBOX 5 (WOX5) based on previous reports indicating that WOX5 can enhance wheat transformation efficiency and reduce genotype dependence (Wang et al., 2022). WOX5 was evaluated in both Fielder and Kronos cultivars. Selection of transformed plants was performed using the GOI‐linked selectable marker, and phenotypic effects were systematically recorded in mature T_0_ plants, including fertility (seed set), plant height, tiller number, spike number, and grain traits (area, length, and weight).
Fielder, ZmUbi::WOX5 yielded ~15% transformation efficiency, producing an average of 1.04 plantlets per line. In Kronos, transformation efficiency was slightly higher with GRF4‐GIF1 constructs (~24%), but WOX5 expression caused severe fertility penalties (~41% sterility) and morphological alterations, including wider, spiralled leaves (Figure 4). Grain area, weight, and length increased, whereas tiller and spike number decreased (Table 4).
Phenotype of Fielder plants transformed with Wox5.Fielder plants expressing Wox5 exhibited distinct morphological changes compared with non‐transformed controls. The mature plants displayed a shorter stature, larger grain size, and wider, spiralled leaf shapes, indicating significant developmental effects associated with Wox5 overexpression.
In contrast, GRF4‐GIF1 consistently produced higher transformation efficiencies with morphologically normal plants. In Kronos, PLTP‐ and HS Cre‐Lox‐driven GRF4‐GIF1 lines restored fertility relative to ZmUbi‐driven GRF4‐GIF1, while WOX5 lines showed strong pleiotropic effects. This comparison highlights GRF4‐GIF1 as a balanced morphogenic regulator, capable of enhancing regeneration without severe developmental penalties.
In summary, GRF4‐GIF1 consistently achieved higher transformation efficiencies while producing morphologically normal plants, highlighting its advantage as a balanced morphogenic regulator for wheat transformation. In both Fielder and Kronos, all constructs reduced seed number relative to controls, but WOX5 caused the strongest fertility penalty in Kronos, accompanied by a marked increase in TGW and grain size. In contrast, PLTP‐ and HS Cre‐Lox‐driven GRF4‐GIF1 lines restored fertility in Kronos. While the HS Cre‐Lox system increased tiller and spike number, it did not fully restore overall yield. Among Kronos transgenic lines, ZmUbi‐driven GRF4‐GIF1 exhibited reduced plant height and high sterility, whereas lines with HS Cre‐mediated excision or PLTP‐driven GRF4‐GIF1 showed substantially recovered fertility. WOX5 expression primarily impaired fertility and produced pronounced morphological changes, including widened, spiralled leaves and thickened stems, despite increases in grain size.
CONCLUSION
GRF4–GIF1 enhances wheat transformation efficiency across diverse cultivars but induces a trade‐off between reproductive and vegetative traits, with high transgene copy numbers leading to reduced fertility, spikelet number, and seed set. These pleiotropic effects are genotype‐dependent, with sensitive cultivars such as Kronos being most affected, while others like Fielder can exhibit compensatory increases in tiller number, spike number, and grain size.
Promoter choice strongly influences the balance between transformation efficiency and phenotypic stability. Constitutive expression using ZmUbi drives high efficiency but exacerbates developmental defects, whereas tissue‐specific expression via the ZmPLTP promoter limits GRF4–GIF1 activity to early developmental stages, maintaining high fertility and normal grain traits while still supporting efficient transformation. Optimisation of GRF4–GIF1 expression, driven by promoters with differing strengths and temporal activities, in combination with varying levels of zeatin and hygromycin, further improves transformation outcomes across multiple tetraploid and hexaploid wheat cultivars.
Complementary strategies, such as the heat‐inducible Cre‐lox excision system, allow post‐regeneration removal of GRF4–GIF1, restoring fertility and mitigating phenotypic abnormalities, particularly in sensitive genotypes. Together, these approaches demonstrate that careful regulation of GRF4–GIF1 expression through promoter selection, controlled excision, and media optimisation can optimise wheat transformation while minimising unintended developmental effects. These findings provide a framework for generating high‐efficiency, phenotypically stable transgenic and gene edited wheat lines.
MATERIALS AND METHODS
Plant material and growth conditions
In this study, we utilised selected hexaploid bread wheat (Triticum aestivum L.) cultivars, as well as a few tetraploid durum wheat (Triticum durum Desf.) cultivars (Table 1). Small batches of wheat seeds from different cultivars were sown at weekly intervals to ensure a continuous supply of immature embryos. Donor plants were grown under controlled environmental conditions at 18 ± 1°C (day) and 15 ± 1°C (night), with 65% relative humidity and light levels of 600 μmol m^−2^ s^−1^ provided by a combination of fluorescent and tungsten lighting in controlled environment growth rooms as described in Hayta et al., 2021. Winter wheat cultivars were exposed to a 4‐week vernalisation period at temperature 5°C, under a short‐day, 8 h light:16 h dark photoperiod, 70 μmol m^−2^ s^−1^.
Construct design and preparation
The hypervirulent Agrobacterium tumefaciens strain AGL1 (Lazo et al., 1991) was used in all plant transformation experiments. All transformation vectors were based on the binary vector pGoldenGreenGate (pGGG) (Smedley et al., 2021) and were assembled using MoClo golden gate assembly (Weber et al., 2011). Each pGGG vector contained the hygromycin selectable marker (hpt) and Cat1 intron driven by the switchgrass ubiquitin promoter (PvUbi) (Addgene # 246468), the GRF4‐GIF1 gene fusion driven by various promoters or excisable, and the β‐glucuronidase gene (GUS) with two introns (GUS2Int) under the control of the rice ubiquitin promoter (Addgene # 246471). The pGGG vectors were electroporated into the Agrobacterium AGL1 competent cells with the helper plasmid pAL155 (Addgene # 246472), which contains an additional VirG gene as previously described in Hayta et al. (2019).
Single colonies of AGL1 containing the desired constructs were inoculated into 10 mL LB medium supplemented with appropriate antibiotics and incubated at 28°C, shaking at 200 rpm for approximately 65 h. A modified version of the Tingay et al. (1997) protocol was used to prepare standard Agrobacterium inocula, following Bartlett et al. (2008). Equal volumes of sterile 30% glycerol and bacterial culture were mixed and aliquoted (400 μL) into 0.5 mL tubes. These inocula were stored at −80°C until use.
The GRF4‐GIF1 gene fusion was synthesised as a MoClo golden gate level 0 component, internal Eco31I and BpiI restriction enzyme sites were removed by silent mutation and standardised MoClo overhangs and Eco31I sites added for assembly. The level 0 GRF4‐GIF1 CDS was assembled into expression cassettes in the Level 1 Position 2 pICH47742 (Addgene # 48001) with the promoters listed in Table 3 and the Nos terminator. All the regular promoters listed were at our laboratory's disposal; however, the Zea mays phospholipid‐transfer protein (ZmPLTP) promoter was designed and synthesised as a MoClo golden gate Level 0 component, internal Eco31I and BpiI restriction enzyme sites removed and standardised MoClo overhangs and Eco31I sites added for assembly. The pGGG GRF4‐GIF1 GUS (Addgene # 246478) vector containing the maize ubiquitin promoter (ZmUbi) driving the GRF4‐GIF1 and the GUS marker gene served as a control.
The inducible site‐specific recombinase (Cre) recombinase CDS under the control of the barley heat‐shock inducible promoter 17 (HvHsp17) was deployed for conditional excision of the ZmUbi::GRF4‐GIF1::NosT and self‐excision of the HvHsp17::Cre::HSTerm. A loxP acceptor cassette was designed and synthesised (Genewiz), which enabled the insertion via BpiI of Level 1 expression cassettes flanked by loxP sites. The cassette in turn could be isolated via Eco31L and cloned into any MoClo L1 acceptor plasmid. The L1P2 HvHsp17::CreInt::HvHsp17::Term and L1P1 ZmUbi::GRF4‐GIF1::Nos were assembled into the loxP cassette, and then the whole assembly was inserted into pICH47811 (AddGene #48008); this vector was deemed L1P2 Rev Cre/LoxP heat shock excisable GRF4‐GIF1 (Addgene # 246486), before cloning in the reverse orientation upstream of the hpt selection cassette in the final Level 2 pGGG binary vector.
Constructs with two separate T‐DNAs provide the opportunity to either use “old” constructs in elite wheat cultivars without having to re‐clone to insert GRF4‐GIF1 or to obtain GRF4‐GIF1‐free plants through segregation in later generations. A T‐DNA cassette was designed and synthesised which was flanked by left and right border repeat sequences and was classically cloned into the pSoup helper plasmid (Addgene # 165419) via the KpnI and HindIII restriction enzyme sites. The L1P2 Cre loxP GRF‐GIF Cassette (Addgene # 246486) was cloned into the pSoup's T‐DNA via Eco31L using a golden gate assembly. The resulting plasmid known as pSoup HSCre GRF4‐GIF1 (Addgene # 246484) can be co‐electroporated into AGL1 with pGGG‐based plasmids or an Agrobacterium culture containing pSoup HSCre GRF4‐GIF1 can be mixed with an Agrobacterium culture containing a suitable plasmid of interest and co‐transformed into wheat.
Various GRF4‐GIF1 constructs, driven by different promoters, were designed for testing in tetraploid wheat, which is more sensitive to phenotypic changes than hexaploid bread wheat (Table 3). Additionally, GRF5‐GIF1 was included as an alternative construct to assess its potential in tetraploid wheat, alongside TaWOX5, which was tested in Kronos and Fielder.
Agrobacterium‐mediated transformation of different wheat cultivars
Immature embryo isolation and transformation procedure
Immature embryos were isolated from wheat grains harvested approximately 14 days post‐anthesis (early milk stage, GS73), when embryos measured 1–1.5 mm in diameter. Kernels from florets 1 and 2 of central spikelets were selected for transformation. Spikes were trimmed and surface‐sterilised with 70% (v/v) ethanol for 1 min, followed by 10% (v/v) sodium hypochlorite for 7 min, and rinsed three times with sterile distilled water.
Under aseptic conditions, immature embryos were excised using fine forceps under a stereomicroscope and placed in wheat inoculation medium (WIM) containing Murashige and Skoog (MS) salts, glucose, MES buffer, Silwet L‐77, and 100 μM acetosyringone (AS).
Agrobacterium preparation and co‐cultivation
A. tumefaciens cultures were prepared 1 day prior to transformation and grown overnight in MG/L medium at 28°C, shaken at 200 rpm. Bacterial cells were pelleted and resuspended in WIM to an optical density of OD_600_ = 0.5, supplemented with 100 μM AS, and incubated for 4–6 h at room temperature before inoculation.
Immature embryos were inoculated with the Agrobacterium suspension and co‐cultivated on solidified WIM containing AgNO_3_, CuSO_4_, and agarose at 24 ± 1°C in darkness for 3 days.
Callus induction and selection
After co‐cultivation, embryogenic axes were excised and transferred to wheat callus induction medium (WCI) containing 2 mg L^−1^ Picloram, 0.5 mg L^−1^ 2,4‐D, and Timentin. Calli were sequentially selected on media containing two levels of hygromycin: Selection 1 (S1) and Selection 2 (S2), each for 2 weeks. The specific hygromycin concentrations used at each selection stage varied depending on the wheat cultivar (Table 2).
Regeneration and rooting
After the first week of S2, embryogenic calli were transferred to light conditions (100 μmol m^−2^ s^−1^, 16 h photoperiod) to initiate shoot regeneration. Regeneration was performed on wheat regeneration medium (WRM) containing MS salts, sucrose, MES buffer, Timentin, and cultivar‐specific concentrations of hygromycin and zeatin riboside (Table 2), at 24 ± 1°C under a 16 h photoperiod. Shoots (1–2 cm) exhibiting root primordia were subsequently transferred to hormone‐free MS medium to promote root development.
Plant acclimatisation
Fully rooted plantlets were gently washed and transferred to soil in cell trays covered with propagator lids for acclimatisation. Plants were maintained under controlled environmental conditions (18 ± 1°C day / 15 ± 1°C night, 65% relative humidity, 400–600 μmol m^−2^ s^−1^ light intensity).
Co‐transformation system
To evaluate co‐transformation efficiency, Agrobacterium containing the GOI construct was co‐inoculated with Agrobacterium harbouring the pSoup HSCre GRF4‐GIF1 construct. Overnight cultures were adjusted to OD_600_ = 0.4 and mixed at a 3:2 ratio of GOI:GRF4‐GIF1 prior to embryo inoculation. Alternatively, pGGG vectors lacking GRF4‐GIF1 were electroporated into Agrobacterium strains already containing pSoup HSCre GRF4‐GIF1, and inocula were prepared and used for transformation following the standard procedure described by Hayta et al. (2019, 2021).
Heat‐shock treatment for inducible site‐specific recombinase (Cre) to excise
GRF4‐GIF1
Plant material transformed with constructs containing the heat‐shock inducible Cre/lox‐mediated excision system for GRF4‐GIF1 was subjected to a 40°C heat treatment for 16 h (overnight) during the second‐regeneration phase. Only calli that had developed green shoots were exposed to heat‐shock prior to sub‐culturing onto fresh media for further development.
DNA extraction and copy number analysis
Leaf samples 0.5–0.7 cm were collected, and DNA extracted using the protocol adapted from (Pallotta et al., 2003). Transgene presence and copy number were determined by quantitative real‐time PCR (qPCR) using TaqMan assays targeting the hygromycin resistance gene (hpt) and the endogenous CO_2_ (Constans‐like, AF490469) gene as described by Hayta et al. (2019).
Primers and probes were designed using the “TaqMan Probe and Primer Design” module in Primer Express (Applied Biosystems). Reactions were performed using the Absolute qPCR Mix (Low ROX version; Catalogue No. AB1318B, ThermoScientific) in a multiplex format detecting both hpt and CO2 targets simultaneously.
Each reaction contained 200 nM of each primer and probe, and 5 μL of DNA template adjusted to a final concentration between 1.25–10 ng μL^−1^ (6.25–50 ng total DNA per reaction). Amplifications were carried out on an Applied Biosystems QuantStudio 5 system.
Thermal cycling conditions were as follows: initial enzyme activation at 95°C for 15 min, followed by 40 cycles of denaturation at 95°C for 15 s and annealing/extension at 60°C for 60 s.
The values obtained were used to calculate transgene copy number using the 2^–ΔΔCT^ calculation as described in (Livak & Schmittgen, 2001).
Transformation efficiency calculation
The transformation efficiency throughout this article was calculated as single events occurring from each embryogenic line (i.e., one regenerated plant derived from one embryo) and displayed as the percentage of positive transgenic plants produced from the total number of immature embryos isolated and inoculated with Agrobacterium in an experiment. Transgenesis was confirmed by qPCR.
Data collection
At maturity, full phenotypic traits, including plant height, tiller number, spike number, TGW, and grain length, width, and area across were measured. Grain traits were quantified using a MARVIN Seed Analyser (GTA Sensorik GmbH; hereafter referred to as “MARVIN”).
Statistical analysis
Data were analysed using ANOVA on phenotypic data in RStudio (v1.3.1056). Means were compared using Tukey's HSD test with a significance level of P < 0.05.
Supporting information
Figure S1. Somatic embryos produced in Fielder and Kronos on R1 medium.
Figure S2. In wheat varieties transformed with the ZmUbi::GRF4‐GIF1 construct, (a) spike number, tiller number, and average grain area increased, whereas plant height, spikelet number, and spike fertility were reduced. Changes in thousand‐grain weight (TGW) depended on the cultivar. (b) Transgene copy number and seed number.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aesaert, S. , Impens, L. , Coussens, G. , Van Lerberge, E. , Vanderhaeghen, R. , Desmet, L. et al. (2022) Optimized transformation and gene editing of the B 104 public maize inbred by improved tissue culture and use of morphogenic regulators. Frontiers in Plant Science, 13, 883847.35528934 10.3389/fpls.2022.883847 PMC 9072829 · doi ↗ · pubmed ↗
- 2Bartlett, J.G. , Alves, S.C. , Smedley, M. , Snape, J.W. & Harwood, W.A. (2008) High‐throughput Agrobacterium‐mediated barley transformation. Plant Methods, 4, 22.18822125 10.1186/1746-4811-4-22PMC 2562381 · doi ↗ · pubmed ↗
- 3Cheng, M. , Fry, J.E. , Pang, S. , Zhou, H. , Hironaka, C.M. , Duncan, D.R. et al. (1997) Genetic transformation of wheat mediated by agrobacterium tumefaciens. Plant Physiology, 115, 971–980.12223854 10.1104/pp.115.3.971PMC 158560 · doi ↗ · pubmed ↗
- 4Debernardi, J.M. , Mecchia, M.A. , Vercruyssen, L. , Smaczniak, C. , Kaufmann, K. , Inze, D. et al. (2014) Post‐transcriptional control of GRF transcription factors by micro RNA mi R 396 and GIF co‐activator affects leaf size and longevity. The Plant Journal, 79, 413–426.24888433 10.1111/tpj.12567 · doi ↗ · pubmed ↗
- 5Debernardi, J.M. , Tricoli, D.M. , Ercoli, M.F. , Hayta, S. , Ronald, P. , Palatnik, J.F. et al. (2020) A GRF‐GIF chimeric protein improves the regeneration efficiency of transgenic plants. Nature Biotechnology, 38, 1274–1279.10.1038/s 41587-020-0703-0PMC 764217133046875 · doi ↗ · pubmed ↗
- 6Grosse‐Heilmann, M. , Cristiano, E. , Deidda, R. & Viola, F. (2024) Durum wheat productivity today and tomorrow: a review of influencing factors and climate change effects. Resources, Environment and Sustainability, 17, 100170.
- 7Hayta, S. , Smedley, M.A. , Clarke, M. , Forner, M. & Harwood, W.A. (2021) An efficient agrobacterium‐mediated transformation protocol for Hexaploid and tetraploid wheat. Current Protocols, 1, e 58.33656289 10.1002/cpz 1.58 · doi ↗ · pubmed ↗
- 8Hayta, S. , Smedley, M.A. , Demir, S.U. , Blundell, R. , Hinchliffe, A. , Atkinson, N. et al. (2019) An efficient and reproducible agrobacterium‐mediated transformation method for hexaploid wheat (Triticum aestivum L.). Plant Methods, 15, 121.31673278 10.1186/s 13007-019-0503-z PMC 6815027 · doi ↗ · pubmed ↗