Genomic characterisation of extensively drug-resistant Acinetobacter baumannii isolates from a tertiary hospital in Ghana
Isabella E. Ashley-Agbedor, Samiratu Mahazu, Fleischer C. N. Kotey, Jennifer Amedior, Bright Adu, Emmanuel Darko, Paul Kwao, Nicholas T. K. D. Dayie, Anthony S. Ablordey, Japheth A. Opintan

TL;DR
This study characterizes drug-resistant Acinetobacter baumannii in Ghana, revealing the presence of multiple antibiotic resistance genes and high-risk clones.
Contribution
The study reports the first detection of specific high-risk clones (ST2 and ST164) producing carbapenemase genes in Ghana.
Findings
18 out of 65 A. baumannii isolates were carbapenem resistant.
blaNDM-1 was the most common resistance gene, often co-occurring with blaOXA-23 or blaOXA-58.
Sequence types ST2 and ST164 were identified as high-risk clones in clinical isolates.
Abstract
Acinetobacter baumannii (A. baumannii) is an emerging “superbug” whose infections have become extremely difficult to treat due to its diverse antimicrobial resistance mechanisms and resistance to last-resort antibiotics including carbapenems. Despite this, data on genetic determinants and genomic context of carbapenemase genes in A. baumannii are scarce in Ghana. This study investigated the genetic determinants of carbapenem resistance in clinical isolates of A. baumannii (Ab) and explored the genetic contexts of carbapenemase-encoding genes in extensively drug-resistant A. baumannii (XDR-Ab). We analysed 65 archived clinical A. baumannii isolates. Identification and antimicrobial susceptibility profiles of the isolates were determined using a MALDI-TOF-MS and the Microscan device, respectively. Carbapenem resistant A. baumannii (CR-Ab) isolates were screened for carbapenemase-encoding…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
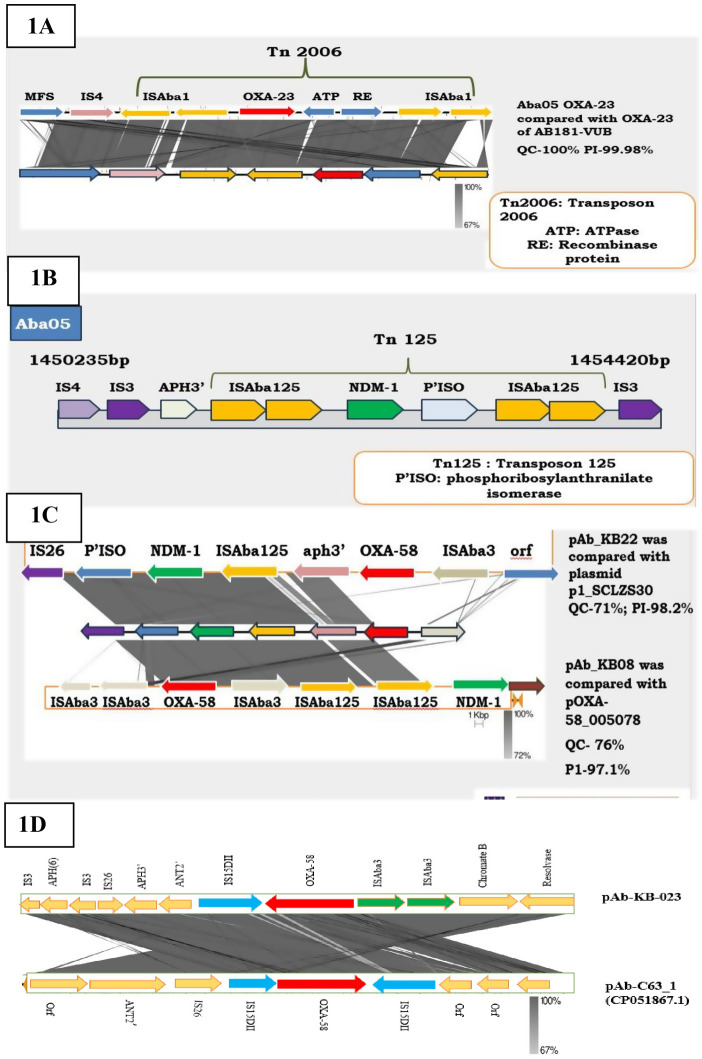 Fig 1
Fig 1 Fig 2
Fig 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Infections and bacterial resistance · Antibiotic Use and Resistance
Introduction
Acinetobacter baumannii is an emerging notorious nosocomial pathogen that has frequently been associated with hospital-acquired infections such as septicemia, urinary tract infections, wound infections, ventilator-associated pneumonia, and surgical site infections, with 20–72.5% of these infections occurring in intensive care units (ICUs) [1–6]. However, the management of these infections continues to be exceedingly difficult because of the wide spectrum of antimicrobial resistance of the organism [7]. Recently, multidrug-resistant A. baumannii (resistance to at least one antimicrobial agent in three or antimicrobial categories) (MDR-Ab), and/or extensively drug-resistant A. baumannii (XDR-Ab) strains (resistance to one agent in all but two or less antimicrobial class) have been implicated in several infections in healthcare facilities worldwide [1,8–17]. Whereas in 2017, CR-Ab was the Number one critical pathogen, in 2024, CR-Ab remains one of the critical pathogens on World Health Organization (WHO) priority lists for which novel therapeutic antibiotics are urgently needed [18–20]. CR in A. baumanni is chiefly mediated by the acquisition of carbapenemases through horizontal gene transfer, mostly facilitated by mobile genetic elements (MGEs) [21–23]. Several studies have reported the production of Class D beta-lactamases (CHDLs) OXA-23-like, OXA24/40-like, OXA-58-like, OXA-143-like, and OXA-235-like enzymes as common genetic determinants of CR in A. baumannii, although the carriage of blaNDM-1, blaVIM, and blaIMP genes has also been extensively described in CR-Ab strains worldwide [24–28]. Across Africa, NDM-1, OXA-23*,* and OXA-58 are the most prevalent carbapenemases reported [11,29–34]. In Ghana, since 2019, when the first report of NDM-1-producing A. baumannii was released [19], limited studies have also described clinical A. baumannii strains harboring bla_OXA-23, blaNDM-1_ and blaOXA-58 genes from Ghanaian hospitals [12,35,36]. Molecular characterisation of CR-Ab isolates has demonstrated the significant role of MGEs (conjugative plasmids, insertion sequences, transposons, and integrons) in the global spread of carbapenemases [22,37–42]. Nevertheless, in Ghana, the paucity of molecular data highlighting the resistance and dissemination mechanisms of CR*/XDR-Ab* undermines efforts to develop effective and efficient strategies for combating infections caused by these strains. In this study, we investigated the genetic determinants of CR as well as the genetic context of carbapenemase-encoding genes and their association with MGEs in whole genome sequenced XDR-Ab isolates from KBTH.
Methods
Study design and setting
The study analyzed a total of 65 archived non-duplicate A. baumannii isolates recovered from in-patients’ specimens processed for culture and susceptibility testing at. the KBTH Central Laboratory between June and December 2022. The KBTH is the largest referral centre in Ghana and has 21 specialised clinical and diagnostic departments. The isolates were stored in nutrient broth with 10% glycerol and kept in a −80 ^0^C freezer. Anonymized data and bacterial isolates were accessed on the July 20, 2023, to obtain information on sample types from which the isolates were recovered and the wards of origin of isolates solely for analysis purpose. Authors had no access to information that could identify individual patient.
Identification of isolates
The stored cultures were brought to room temperature to thaw and subsequently sub-cultured on nutrient agar (Beckton, Dickinson and Company, USA) and incubated aerobically for 24 hours at 37 ^0^C. Discrete colonies were selected and identified using the MALDI-TOF Biotyper (Bruker Daltonics, Karlsruhe, Germany).
Antibiotic Susceptibility Testing
The susceptibility profiles of isolates to meropenem (10 µg), imipenem (10 µg), and doripenem (10 µg) (BD BBL, USA) were determined by the Kirby-Bauer disc diffusion method as described in the CLSI [43]. For quality control, a known carbapenem susceptible strain E. coli ATCC 25922 and carbapenem producing A. baumannii NCTC 13304 were used. Isolates that were found to be resistant to imipenem, doripenem and meropenem (≤15 mm zone of inhibition) were further tested to determine their MICs for 14 antibiotics including meropenem, doripenem and imipenem using the Microscan (Beckman Coulter, USA). Interpretation of MIC results was done following the CLSI guidelines (2023) [43]. A. baumannii isolates that were resistant to all the carbapenems tested (meropenem, doripenem and imipenem) were selected for genotypic characterization.
Genotypic detection of carbapenemase genes
Genomic DNA extraction of all CR isolates was performed using the MagAttract High Molecular Weight (HMW) DNA Kit (Qiagen, Hildon, Germany), according to the manufacturer’s instructions [44]. LAMP reaction was used to screen for carbapenemase-encoding genes blaNDM-1, blaKPC, blaVIM, blaOXA-48, blaOXA-23, blaOXA-58 and blaIMP using primers described by Lahiri and colleagues [45].
Whole-genome sequencing and multi-locus sequence typing
Whole-genome sequencing was performed on six XDR-A. baumannii isolates. Genomic DNA was extracted from the isolates using a NucleoSpin tissue kit according to the manufacturer’s instruction (Macherey-Nagel, Düren, Germany) [46]. The Native Barcoding Kit EXP-NBD104 and the ligation sequencing kit SQK-LSK109 enabled the preparation of gDNA libraries for long-read sequencing [47]. Sequencing was performed using the Nanopore MinION (Oxford Nanopore Technologies, United Kingdom) [48] and sequences were assembled with Flye v2.6 [49]. MLST of the isolates was performed using the online tool PubMLST Pasteur scheme [50].
Bioinformatics, phylogenetic, and statistical analyses
The online system RAST and BLAST tools were used to annotate and compare the assembled sequences [35]. Resfinder v4.3.3 (Center for Genomic Epidemiology) [51] was used to detect AMR-encoding genes. Mobile genetic elements (MGEs) were identified using mge_finder 1.1.2, Uniprot BLAST and ISfinder (www-is.biotoul.fr) [52]. The SnapGene viewer v4.1.9 and Easyfig v2.2.5 were used to visualise the genetic contexts of carbapenemase-encoding genes and compare sequences [35,53]. To determine the genetic relatedness of isolates, a single nucleotide polymorphism (SNP) based phylogenetic tree was constructed using the CSIPhylogeny tool (Center for Genomic Epidemiology). A. baumannii reference strain GCF_0086326351.1 was used as rooting genome [54]. Figtree V1.4.4 enabled the visualisation and annotation of phylogeny trees [54,55]. The isolates chromosomes and plasmid sequences have been deposited with GenBank (SAMN43561363, SAMN43561408, SAMN43561092, SAMN43800676 and SAMN43303015).
Statistical Products and Services Solutions (SPSS), version 22.0 for Microsoft Windows [56], was used to analyze data, which involved descriptive statistics (such as means, tables, and graphs) and Chi-square tests at 95% confidence interval (CI). A p-value of ≤ 0.05 was considered statistically significant.
Ethical approval
The study protocol was reviewed and approved by the Korle Bu Teaching Hospital Scientific and Technical Committee/Institutional Review Board (KBTH-STC/1RB/00071/2023). Data and bacterial isolates used for the study were anonymized, hence informed consent was not required.
Results
Antimicrobial resistance profile of CR isolates
The 65 A. baumannii isolates, were recovered from wound swabs (n = 26), urine (n = 23), blood (n = 7), aspirates (n = 4), catheter tip (n = 2), HVS (n = 2) and eye swab (n = 1). Overall, CR was observed in 18/65 (27.7%) isolates. Table 1 shows the MICs of CR-isolates. CR isolates were mostly isolated from surgical wounds (n = 7, 44.4%), urine (n = 7, 38.9%), blood (n = 2, 11,1%), ulcer wounds (n = 1, 5.5%) and aspirate (n = 1, 5.5%).
Table 1: Antimicrobial resistance profile of CR-A. baumannii isolates.
Genotypic detection of carbapenemase genes in CR-A-baumannii isolates
All 18 CR-Ab isolates screened for carbapenemase-encoding genes harbored a corresponding blaOXA-51 carbapenemase gene. Additionally, the Metallo beta-lactamase (MBL) blaNDM-1 (85.7%) and CHDL blaOXA-23 (71.4%) were the most prevalent genes, followed by blaOXA-58 (42.9%) and blaOXA-420 (14.3%) was the least prevalent gene detected. Four (22.2%) isolates lacked any acquired carbapenemases.
Whole genome sequencing of XDR-A. baumannii isolates
Table 2 shows the genomic characteristics of XDR-A. baumannii isolates. The most predominant gene was blaNDM-1 (66.7%) detected in isolates Aba05, Aba08, Aba20 and Aba22 and coexisted with either blaOXA-23 (Aba05 and Aba20) or blaOXA-58 (Aba08 and Aba22). Besides, the isolates harbored other AMR encoding genes that conferred resistance against aminoglycosides (aac (6’) aadA2, aph (3’)-19, 16SrRNAmethythransrase), fluoroquinolones (A. baumannii ParC/gyrA), sulfonamides (Sul1/Sul2), disinfectants/antiseptics (qacE delta1/qacG) and macrolides (macrolide phosphor-transferase, mphE). Multidrug efflux pumps, including members of the major facilitator superfamily (MFS) and resistance–nodulation division family (RND) were also identified in the isolates’ genomes.
Table 2: Molecular characteristics of XDR- A. baumannii isolates.
Genetic contexts of blaOXA-23, blaNDM-1, blaOXA-58 and blaOXA-420
Fig 1 (A–D) depicts the genetic environments of carbapenemase genes detected in the study isolates. The blaOXA-23 in Aba05 and Aba20 was chromosomally embedded and was flanked upstream by TetB-IS4-element-ISAba1-ISAba1 and downstream by ISAba1-ISAba1. Furthermore, composite transposon Tn2006 was detected in both isolates (Fig 1A).
(A–D). The genetic contexts of blaOXA-23, blaNDM-1, blaOXA-58 and blaOXA-420 in CR-A. baumannii. (A) Shows blaOXA-23 genetic context in Aba05 and Aba20 compared with A. baumannii strain AB181-VUB, blaOXA-23 gene was embedded in transposon Tn2006. (B) In isolates Aba05 and Aba20, the blaNDM-1 genes were identified between chromosomes sequences (1450235 bp - 145440 bp) and (838,300 bp – 843,418 bp) respectively, captured by transposon Tn125. (C). The whole plasmid sequence of Aba08 and Aba22 encoding blaOXA-58 and blaNDM-1 genes compared with plasmids pOXA-58_005078 (D) shows blaOXA-420 surrounding regions identified in Aba23 plasmid compared with plasmid pAb-c631_1.
The blaNDM-1 was also detected on the chromosomes of Aba05, Aba11 and Aba20. In Aba20, the blaNDM-1 was flanked by two copies of ISAba125 upstream and, a sequentially arranged ISAba125-ISAba125-IS3-ISAba14 elements downstream, embedding in a composite transposon Tn125 as described by Poirel et al (2012) (fig 1B). A further screening of the upstream region of blaNDM-1 showed an array of aph3-ISAba33-IS4-IS3-element. However, in Aba20, only one copy of ISAba125 was detected upstream of bla_NDM-1_ and ISAba34 downstream. Also, IS4 and aph3’ were located upstream of ISAba125. In Aba11, an entirely different genetic environment was observed for blaNDM-1 in which insertion sequence ISAde1 bracketed the gene. Flanking ISAde1 downstream*,* were the sequentially arrayed aph3’, ISAba14 and IS3 transposase elements.
Plasmid-borne (pAb_KB08 and pAb_KB22) blaNDM-1 and blaOXA-58 was carried by isolates Aba08 and Aba22. In the genetic environment of blaNDM-1 and blaOXA-58 on plasmid pAb_KB08, the presence of msr, mph2’, resolvase-invertase-type recombinase ISAba3, ISAba3, blaOXA58, ISAba3, ISAba125, ISAba125, blaNDM-1, bleMBLs, and ISAba32 were detected. A nucleotide BLAST analysis of the whole plasmid showed it shared 98.17% similarities with plasmid p1_SCLZS30, (query cover 71%; accession number: CP090385.1) and 97.1% identity with plasmid pOXA-58_005078 (query cover 76%; accession number: CP033131.1) (Fig 1C). Although the blaOXA-66 gene was detected in both isolates, no insertion sequences were detected upstream or downstream of the gene. However, the genetic environment of blaOXA-58 in isolate Aba11 varied from that of Aba08 and Aba22. For Aba11, insertion sequence IS15DII and ISAba3-aph3’ were found upstream and downstream of blaOXA-58 respectively. A BLAST search of this plasmid showed 63% query coverage and 99.93% percentage identity with plasmid pNDM_SCLZS86 (accession number: CP041590.1).
The blaOXA-420 (an OXA-58-like enzyme) was also plasmid-encoded carried by isolate Aba23 (pAb_KB23). Insertion sequences IS15DII and ISAba3 flanked the blaOXA-420 upstream and downstream respectively. In the immediate upstream region of IS15DII, an array of IS26, aph(3’)-la, aph(3”)-b, aph(6)-Id and ant(2”)-la sequentially arranged, whilst in the downstream of blaOXA-420, mph(E), Sul2 and tet (39) were detected. In addition, other genetic elements such as ISAlw27, IS1006 and ISAcsp12 were also identified in the upstream region of IS26. A nucleotide BLAST analysis of plasmid pAb_KB23 showed, its 99.74% identity with plasmid pAb-C63_1 (100% query cover; accession number CP051867.1) (Fig 1D).
Genetic diversities of XDR-A. baumannii isolates
Fig 2 shows the phylogeny of XDR isolates. The isolates were genetically diverse belonging to sequence types, ST 2 (Aba20 and Aba05), ST 164 (Aba22), ST 2146 (Aba23), ST 52 (Aba11), and ST 16 (Aba08). Nevertheless, isolates Aba05 and Aba20 recovered from two patients admitted to the surgical and stroke units respectively, belonged to the same sequence type ST 2,
Genetic relation of XDR-A. baumannii isolates recovered from different in-patient specimens and their sequence types (STs).
Discussion
MDR*-A. baumannii* infections continue to pose a major threat in healthcare settings, yet much remains to be understood and discovered about this nosocomial pathogen. In the current study, we report approximately a 28% prevalence of CR-Ab in a major referral hospital in Ghana. Comparatively, this prevalence is higher than the 5.6% reported for the same country by Ayibieke et al [35], indicating a potential increase in the prevalence of CR-Ab. Our findings aligns with CR-Ab prevalence of 20% and 22.4% reported from some studies conducted in the USA and Canada respectively [57,58]. Elsewhere in Africa, higher prevalence of 61% and 49% have been reported in Ethiopia and Algeria respectively [29,59]. The rising prevalence of CR-Ab across the Sub-Saharan African region could further worsen the disease burden and mortality associated or attributed with antimicrobial resistance (AMR) [60].
In A. baumannii, the genetic basis of carbapenem-resistance has widely been associated with the production of carbapenem-hydrolyzing class D β-lactamases and Metallo β-lactamases [61]. This observation is consistent with our findings where most of the isolates harbored Class B (NDM-1) and Class D (OXA-23, OXA-58 and OXA-420) carbapenemases.
We observed the co-existence of blaNDM-1 with either blaOXA-23 or blaOXA-58 in XDR-Ab isolates. The emergence of A. baumannii strains co-harboring blaOXA-23 and blaNDM-1 was first reported in India, a sub-continent considered to be the reservoir for OXA-23 and NDM-1-producing A. baumannii strains [62]. Subsequently, Nepal, Egypt, Pakistan and Libya have also reported the co-existence of blaOXA-23 and blaNDM-1 in A. baumannii [16,28,61,63]. Our observation of the co-existence of blaNDM-1 and blaOXA-23 is consistent with a previous study in Ghana that highlighted the predominance of CR-Ab co-harboring blaOXA-23 and blaNDM-1 [12]. Studies in South Africa, however, show that CR-Ab isolates predominantly carry only the blaOXA-23 gene. [33,64,65]
Analysis of the genetic environment of blaNDM-1 and blaOXA-23 revealed the presence of ISAba1 and ISAba125 upstream of both genes respectively. The insertion of ISAba1 and ISAba125 upstream of blaOXA-23 and blaNDM-1 respectively, provided strong sequence promoters and resulted in increased carbapenem resistance among A. baumannii isolates [25,66–68]. More importantly, blaOXA-23 and blaNDM-1 resided within composite transposons Tn2006 and Tn125 respectively which are major dissemination vehicles for blaOXA-23 and blaNDM-1 genes worldwide [24,25,67]. The detection of Tn2006 and Tn125 in our isolates raises concern since these MGEs could further facilitate the spread of blaNDM-1 and blaOXA-23 genes among A. baumannii strains and other Gram-negative pathogens across the hospital.
The study also identified four distinct plasmids (pAbKB_08, pAbKB_11, pAbKB_22 and pAbKB_23) encoding blaOXA-58, blaNDM-1 and blaOXA-420 genes. The carriage of plasmid-borne carbapenemase-encoding genes by isolates recovered from patients admitted to different wards, buttresses the disperse nature of these transferable MGEs among A. baumannii isolates across the hospital. This confirms A. baumannii and other Gram-negative organisms’ propensity to easily acquire and accumulate AMR-genes [69,70]. In 2003, infection outbreaks caused by OXA-58-producing A. baumannii was first reported from France, and are currently on a global scale [71–75]. However the occurrence of CR-Ab harboring plasmid-encoded blaOXA-58 and blaNDM-1, is scarce [70], limited studies from Thailand, Malaysia and China, have described plasmid-borne blaOXA-58 and blaNDM-1 in A. baumannii, A. pitti and A. nosocomialis [76–78]. In Ghana, whereas a plasmid-borne blaOXA-58, was described in CR-Ab [35], to the best of our knowledge this study is the first to report plasmids co-harboring blaOXA-58 and blaNDM-1 in this country. Moreover, plasmid pAbKB_22 99.80% similarities with plasmid GD03255 identified in A. bereziniae and its 98.2% identity with plasmid pl_SCLZS30 carried by A. towneri, as well as plasmid pAbKB_08 97.61% identity with plasmid pOXA-58_010062 harbored by A. wuhouensis strain*,* (all detected in China, unpublished), may seem to point to the fact that the occurrence of plasmid-borne blaOXA-58 and blaNDM-1 in Acinetobacter species may be endemic in Asia. Similar to our findings, a recent report from Brazil also identified plasmid-borne blaOXA-58 and blaNDM-1 in A. baumannii clinical isolates [70]. This gives indication of the possible steady spread of these MGEs across the nations.
CR-Ab carrying blaOXA-420 was first detected in Nepal [78,79] and subsequent studies in Nigeria and Kenya have also reported on OXA-420-producing A. baumannii isolates [80,81]. In Ghana, two studies have described OXA-420 A. baumannii producers [12,35] and this study is the third to report on these strains. Interestingly, plasmid pAbKB_23, detected in the current study share 100% sequence identity with plasmid pAb-c63, previously described by Ayibieke et al in Ghana [35]. The detection of blaOXA-420 in three separate studies conducted in three different regions (Accra, Takoradi and Eikwe) in Ghana affirms the suspicion of Ayibieke et al of the probable circulation of blaOXA-420 in hospitals in Ghana requiring urgent infection control and prevention measures to stop their spread.
Globally, outbreaks of infections due to diverse populations of A. baumannii strains have been reported and the majority of these infections are driven by global clone 1 (GC1) and global clone 2 (GC2), represented as ST1 and ST2 respectively. ST2 is the dominant sequence type of A. baumannii in Thailand and most widespread clones, influencing the spread of carbapenem-resistance genes around the world. [25,82,83]. ST2 A. baumannii strains are also noted for XDR and treatment difficulties or failures due to their higher virulence [84]. In our study, the two ST2 isolates (Aba05 and Aba20) were genetically related, indicating a possible clonal spread. In Ghana, whereas ST1 A. baumannii isolates have been previously described in two separate studies [35,36], to the best of our knowledge, this is the first detection of ST2 A. baumannii isolates in Ghana. Studies conducted in Lebanon, Greece, Italy, Turkey and Denmark have associated a series of nosocomial infection outbreaks with ST2 A. baumannii isolates carrying blaOXA-23 and blaNDM-1 [85–87] and therefore, their detection in the KBTH in Ghana is of concern and calls for intensive surveillance and infections control measures to avert a possible outbreak. Also, the study report for the first time, the detection of ST164 OXA-58, NDM-1-producing A. baumannii strain in Ghana. ST164, NDM-1 positive A. baumannii strains are endemic in Thailand [83], nevertheless, this strain has also been reported in China, Egypt, Sudan and recently, in Denmark [54,87–89]. It is worth noting that the isolates described in these previous studies lacked the blaOXA-58 gene which was present in our isolate. ST164 A. baumannii is an emerging high-risk clone that needs to be controlled to avoid their spread. This study also identified ST2146 OXA-420 positive A. baumannii strain (Aba23). ST2146 is a novel sequence type, newly assigned to a CR-A. baumannii isolate carrying blaOXA-420 and blaNDM-1 in a previous study in Ghana. [12] and its detection in this present study indicates ST2146 may be an emerging clone steadily spreading in Ghana. Similarly, the detection of ST16 (isolate Aba08) and ST52 (Aba11) sequence types frequently associated with Asian countries such as Myanmar, Thailand and Vietnam [90,91] may suggest a possible emergence of these strains in Ghana.
Although only a limited number of genomes were studied due to financial constraints, the study nevertheless has revealed significant findings, that may shed light on the dynamics of CR and XDR in A. baumannii as well as added to the limited number of genomic characterisation studies of A. baumannii in Ghana and Africa.
Conclusion
Our study highlights the occurrence of multiple carbapenemase-encoding genes, other AMR encoding genes such as aac(6’) aadA2, aph(3’)-19, 16SrRNAmethythransrase, ParC/gyrA, Sul1/Sul2, qacE delta1/qacG, mphE, and different efflux pumps as major antimicrobial mediating factors in XDR clinical A. baumannii isolates. The study also reveals the close association of carbapenemase-encoding genes with diverse transferable MGEs that could facilitate their spread among A. baumannii and other Gram-negative pathogens. We reported to the best of our knowledge, the first detection of high-risk clones ST2 OXA-23, NDM-1 producing A. baumannii and ST164 OXA-58, NDM-1-producing A. baumannii strains in Ghana. Our findings further emphasize the need for more genomic epidemiological studies and continuous surveillance to ensure the effective control of the spread of resistant organisms and high risk clones especially in hospitals.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cai B, Echols R, Magee G, Arjona Ferreira JC, Morgan G, Ariyasu M, et al. Prevalence of Carbapenem-Resistant Gram-Negative Infections in the United States Predominated by Acinetobacter baumannii and Pseudomonas aeruginosa. Open Forum Infect Dis. 2017;4(3):ofx 176. doi: 10.1093/ofid/ofx 176 29026867 PMC 5629822 · doi ↗ · pubmed ↗
- 2Hsueh P-R, Teng L-J, Chen C-Y, Chen W-H, Yu C-J, Ho S-W, et al. Pandrug-resistant Acinetobacter baumannii causing nosocomial infections in a university hospital, Taiwan. Emerg Infect Dis. 2002;8(8):827–32. doi: 10.3201/eid 0805.020014 12141969 PMC 2732518 · doi ↗ · pubmed ↗
- 3Levin AS, Levy CE, Manrique AEI, Medeiros EAS, Costa SF. Severe nosocomial infections with imipenem-resistant Acinetobacter baumannii treated with ampicillin/sulbactam. Int J Antimicrob Agents. 2003;21(1):58–62. doi: 10.1016/s 0924-8579(02)00276-5 12507838 · doi ↗ · pubmed ↗
- 4Wisplinghoff H, Paulus T, Lugenheim M, Stefanik D, Higgins PG, Edmond MB, et al. Nosocomial bloodstream infections due to Acinetobacter baumannii, Acinetobacter pittii and Acinetobacter nosocomialis in the United States. J Infect. 2012;64(3):282–90. doi: 10.1016/j.jinf.2011.12.008 22209744 · doi ↗ · pubmed ↗
- 5Fragkou PC, Poulakou G, Blizou A, Blizou M, Rapti V, Karageorgopoulos DE, et al. The Role of Minocycline in the Treatment of Nosocomial Infections Caused by Multidrug, Extensively Drug and Pandrug Resistant Acinetobacter baumannii: A Systematic Review of Clinical Evidence. Microorganisms. 2019;7(6):159. doi: 10.3390/microorganisms 7060159 31159398 PMC 6617316 · doi ↗ · pubmed ↗
- 6Richet H, Fournier PE. Nosocomial infections caused by Acinetobacter baumannii: a major threat worldwide. Infect Control Hosp Epidemiol. 2006;27(7):645–6. doi: 10.1086/505900 16807835 · doi ↗ · pubmed ↗
- 7Ramirez MS, Bonomo RA, Tolmasky ME. Carbapenemases: Transforming Acinetobacter baumannii into a Yet More Dangerous Menace. Biomolecules. 2020;10(5):720. doi: 10.3390/biom 10050720 32384624 PMC 7277208 · doi ↗ · pubmed ↗
- 8Labarca JA, Salles MJC, Seas C, Guzmán-Blanco M. Carbapenem resistance in Pseudomonas aeruginosa and Acinetobacter baumannii in the nosocomial setting in Latin America. Crit Rev Microbiol. 2016;42(2):276–92. doi: 10.3109/1040841 X.2014.940494 25159043 · doi ↗ · pubmed ↗