Rosanoid diterpenoids: structural diversity, classification and biological activities
Sabrin R. M. Ibrahim, Hani Z. Asfour, Gamal A. Mohamed, Nabil A. Alhakamy, Hossam M. Abdallah, Hagar M. Mohamed

TL;DR
This paper reviews rosanoid diterpenoids, a unique class of natural compounds with diverse structures and biological activities, highlighting their potential for drug development.
Contribution
A comprehensive review of rosanoid diterpenoids from 1975 to 2025, focusing on their structural diversity and pharmacological potential.
Findings
Rosanoid diterpenoids are mainly found in Euphorbia species and show chemotaxonomic relevance to the Euphorbiaceae family.
They exhibit a wide range of biological activities, including anti-inflammatory, antimicrobial, and cytotoxic effects.
The compounds show promise for drug discovery but require further studies on pharmacokinetics and biosynthesis.
Abstract
Rosanoid diterpenoids, including ent-rosane and rosane diterpenoids, are structurally unique and bioactive subclass diterpenes characterized by a tricyclic carbon skeleton. This work aims to provide a comprehensive review of the literature on these diterpenoids from 1975. to September 2025., including their occurrence, structural diversity, and biological activities. An extensive literature search was conducted through scientific databases (ScienceDirect, PubMed, Scopus, Web of Science, and Google Scholar) and publishers’ webpages (Elsevier, Wiley, ACS, RSC, Taylor & Francis, Springer, Bentham, Thieme, and MDPI), covering reports from 1975 to September 2025. Rosanoid diterpenoids have been isolated from various natural sources, including fungi, liverworts, and higher plant families such as Euphorbiaceae, Lamiaceae, Alismataceae, Asteraceae, Velloziaceae, and Celastraceae. They are…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
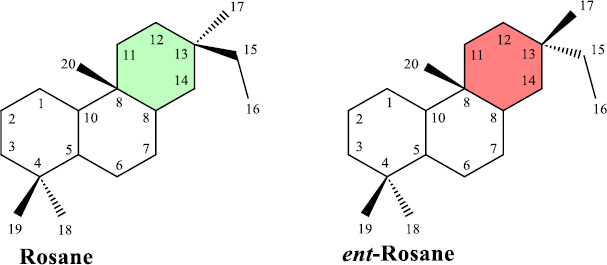 Figure 1
Figure 1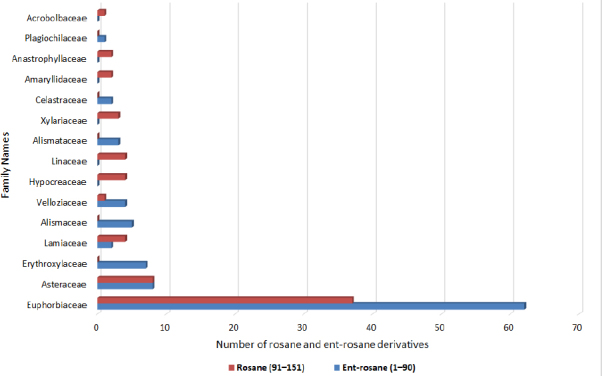 Figure 2
Figure 2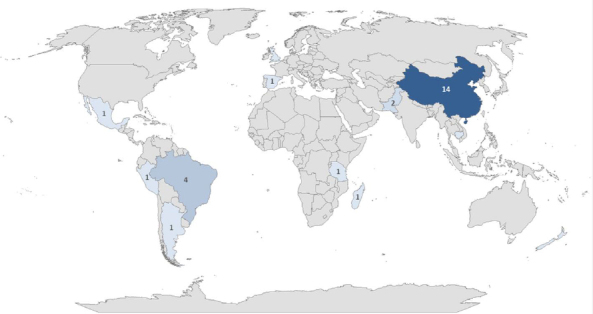 Figure 3
Figure 3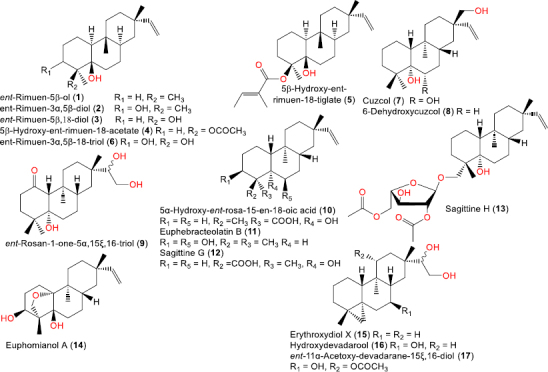 Figure 4
Figure 4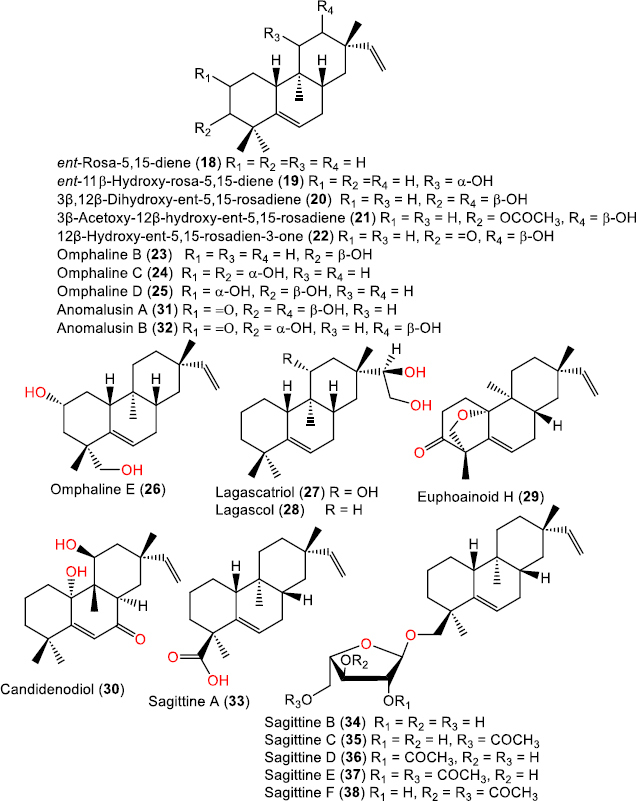 Figure 5
Figure 5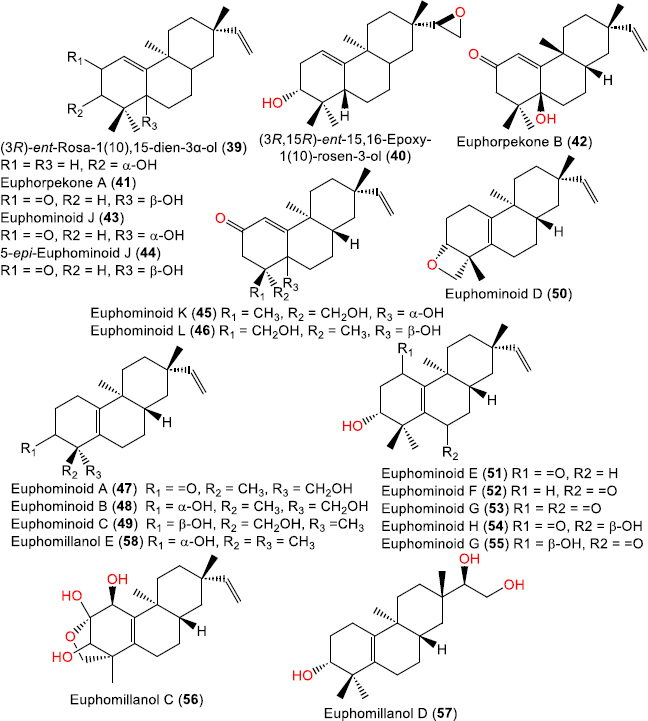 Figure 6
Figure 6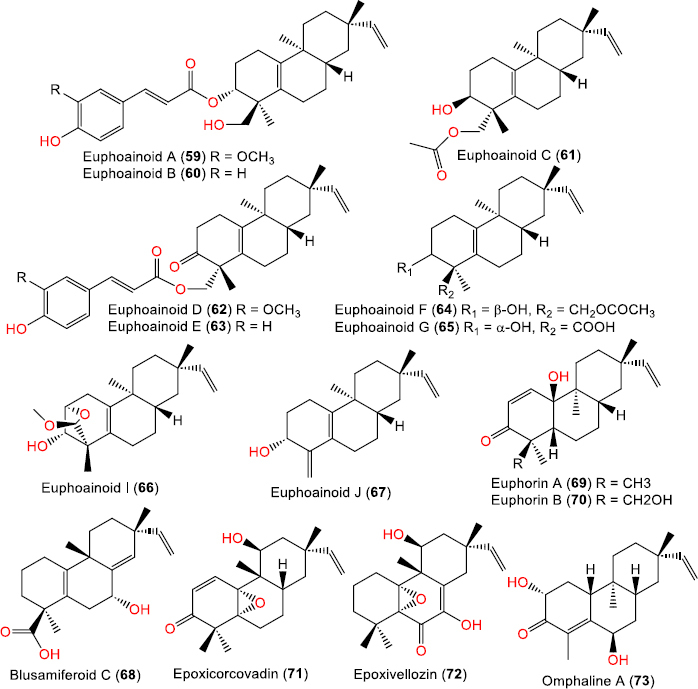 Figure 7
Figure 7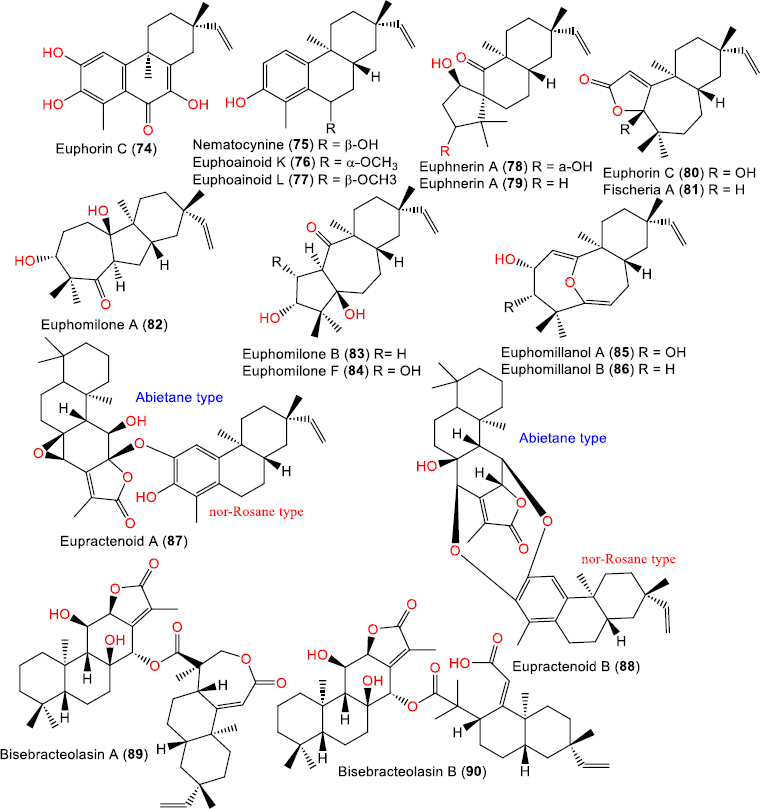 Figure 8
Figure 8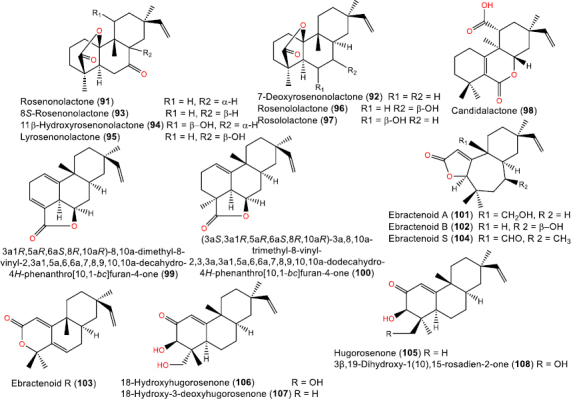 Figure 9
Figure 9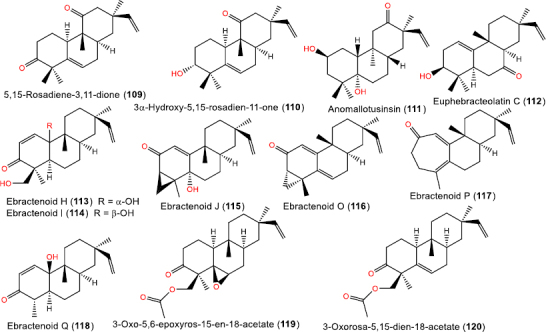 Figure 10
Figure 10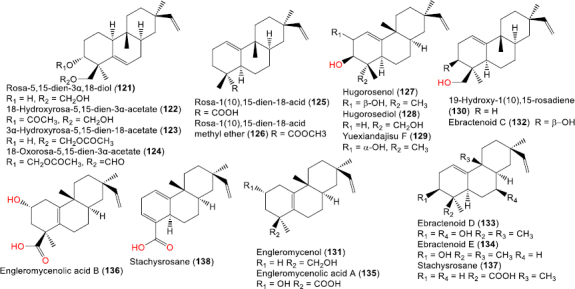 Figure 11
Figure 11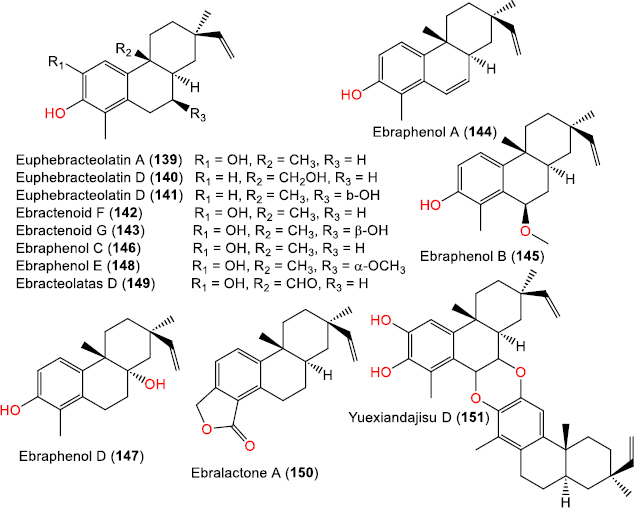 Figure 12
Figure 12| Compound name/class | Molecular weight | Molecular formula | Plant, family, part used and location | Ref. |
|---|---|---|---|---|
| Saturated ent-rosane diterpenoids | ||||
| ent-Rimuen-5β-ol ( | 290 | C20H34O | [ | |
| ent-Rimuen-5β,18-diol ( | 292 | C19H32O2 | ||
| ent-Rimuen-3α,5β-diol ( | 306 | C20H34O2 | ||
| 5β-Hydroxy-ent-rimuen-18-acetate ( | 334 | C21H34O3 | ||
| 5β-Hydroxy-ent-rimuen-18- tiglate ( | 374 | C24H38O3 | ||
| ent-Rimuen-3α,5β-18-triol ( | 308 | C19H32O3 | ||
| Cuzcol ( | 322 | C20H34O3 | [ | |
| 6-Dehydroxycuzcol ( | 306 | C20H34O2 | [ | |
| ent-Rosan-1-one-5α,15ξ,16-triol ( | 338 | C20H34O4 | [ | |
| 5α-Hydroxy-ent-rosa-15-en-18-oic acid ( | 320 | C20H32O3 | [ | |
| Euphebracteolatin B ( | 306 | C20H34O2 | [ | |
| [ | ||||
| Sagittine G ( | 320 | C20H32O3 | [ | |
| [ | ||||
| Sagittine H ( | 522 | C29H46O8 | [ | |
| Euphomianol A ( | 320 | C20H32O3 | [ | |
| Erythroxydiol X ( | 306 | C20H34O2 | [ | |
| Hydroxydevadarool ( | 322 | C20H34O3 | [ | |
| ent-11α-Acetoxy-devadarane--15ξ,16-diol ( | 364 | C22H36O4 | [ | |
| ent-Rosa-5,15-diene ( | 272 | C20H32 | - | [ |
| ent-11β-Hydroxy-rosa-5,15-diene ( | 288 | C20H32O | - | [ |
| 3β,12β-Dihydroxy-ent-5,15-rosadiene ( | 304 | C20H32O2 | [ | |
| 3β-Acetoxy-12β-hydroxy-ent-5,15-rosadiene ( | 346 | C22H34O3 | [ | |
| 12β-Hydroxy-ent-5,15-rosadien-3-one ( | 302 | C20H30O2 | [ | |
| Omphaline B ( | 288 | C20H32O | of | [ |
| Omphaline C ( | 304 | C20H32O2 | of | [ |
| Omphaline D ( | 304 | C20H32O2 | of | [ |
| Omphaline E ( | 304 | C20H32O2 | of | [ |
| Lagascatriol ( | 322 | C20H34O3 | [ | |
| Lagascol ( | 306 | C20H34O2 | [ | |
| Euphoainoid H ( | 300 | C20H28O2 | [ | |
| Candidenodiol ( | 318 | C20H30O3 | [ | |
| Anomalusin A ( | 318 | C20H30O3 | [ | |
| Anomalusin B ( | 318 | C20H30O3 | [ | |
| Sagittine A ( | 302 | C20H30O2 | [ | |
| Sagittine B ( | 420 | C25H40O | [ | |
| [ | ||||
| Sagittine C ( | 462 | C27H42O6 | [ | |
| Sagittine D ( | 462 | C27H42O6 | [ | |
| Sagittine E ( | 504 | C29H44O7 | [ | |
| [ | ||||
| Sagittine F ( | 504 | C29H44O7 | [ | |
| [ | ||||
| ent-Rosane diterpenoids with Δ1(10) unsaturation in rings A and B | ||||
| (3 | 288 | C20H32O | [ | |
| [ | ||||
| (3 | 304 | C20H32O2 | [ | |
| [ | ||||
| Euphorpekone A ( | 302 | C20H30O2 | [ | |
| Euphorpekone B ( | 302 | C20H30O2 | [ | |
| Euphominoid J ( | 302 | C20H30O2 | [ | |
| 5-epi-Euphominoid J ( | 302 | C20H30O2 | [ | |
| Euphominoid K ( | 318 | C20H30O3 | [ | |
| Euphominoid L ( | 318 | C20H30O3 | [ | |
| Δ5(10),Δ15(16)-unsaturated ent-rosane diterpenois | ||||
| Euphominoid A ( | 302 | C20H30O2 | [ | |
| [ | ||||
| Euphominoid B ( | 304 | C20H32O2 | [ | |
| Euphominoid C ( | 304 | C20H32O2 | [ | |
| Euphominoid D ( | 286 | C20H30O | [ | |
| Euphominoid E ( | 302 | C20H30O2 | [ | |
| [ | ||||
| Euphominoid F ( | 302 | C20H30O2 | [ | |
| Euphominoid G ( | 316 | C20H28O3 | [ | |
| Euphominoid H (54) | 318 | C20H30O3 | [ | |
| Euphominoid I ( | 318 | C20H30O3 | [ | |
| Euphomillanol C ( | 334 | C20H30O4 | [ | |
| Euphomillanol D ( | 322 | C20H34O3 | [ | |
| Euphomillanol E ( | 288 | C20H32O | [ | |
| Euphoainoid A ( | 480 | C30H40O5 | [ | |
| Euphoainoid B ( | 450 | C29H38O4 | [ | |
| Euphoainoid C ( | 346 | C22H34O3 | [ | |
| Euphoainoid D ( | 478 | C30H38O5 | [ | |
| Euphoainoid E ( | 448 | C29H36O4 | [ | |
| Euphoainoid F ( | 346 | C22H34O3 | [ | |
| Euphoainoid G ( | 318 | C20H30O3 | [ | |
| Euphoainoid I ( | 332 | C21H32O3 | [ | |
| Euphoainoid J ( | 272 | C19H28O | [ | |
| Other unsaturated ent-rosane diterpenoids | ||||
| Blusamiferoid C ( | 318 | C20H28O3 | [ | |
| Euphorin A ( | 302 | C20H30O2 | [ | |
| [ | ||||
| [ | ||||
| Euphorin B ( | 318 | C20H30O3 | [ | |
| [ | ||||
| Epoxicorcovadin ( | 316 | C20H28O3 | [ | |
| Epoxivellozin ( | 332 | C20H28O4 | [ | |
| Omphaline A ( | 304 | C19H28O3 | of | [ |
| Aromatic ent-rosane diterpenoids | ||||
| Euphorin C ( | 314 | C19H22O4 | [ | |
| [ | ||||
| Nematocynine ( | 286 | C19H26O2 | [ | |
| Euphoainoid K ( | 300 | C20H28O2 | [ | |
| Euphoainoid L ( | 300 | C20H28O2 | [ | |
| Modified skeletons ent-rosane diterpenoids | ||||
| Euphnerin A ( | 320 | C20H32O3 | [ | |
| Euphnerin B ( | 304 | C20H32O2 | [ | |
| Euphorin D ( | 304 | C19H28O3 | [ | |
| [ | ||||
| [ | ||||
| Fischeria A ( | 288 | C19H28O2 | [ | |
| [ | ||||
| [ | ||||
| Euphomilone A ( | 320 | C20H32O3 | [ | |
| Euphomilone B ( | 320 | C20H32O3 | [ | |
| Euphomilone F ( | 336 | C20H32O4 | [ | |
| Euphomillanol A ( | 318 | C20H30O3 | [ | |
| Euphomillanol B ( | 302 | C20H30O2 | [ | |
| Dimeric ent-rosane diterpenoids | ||||
| Eupractenoid A ( | 616 | C39H52O6 | [ | |
| Eupractenoid B ( | 600 | C39H52O5 | [ | |
| Bisebracteolasin A ( | 664 | C40H56O8 | [ | |
| Bisebracteolasin B ( | 666 | C40H58O8 | [ | |
| Compound name/class | Molecular weight | Molecular formula | Plant, family, part used and location | Ref. |
|---|---|---|---|---|
| Lactone-containing rosane diterpenoids | ||||
| Rosenonolactone ( | 316 | C20H28O3 | [ | |
| [ | ||||
| [ | ||||
| [ | ||||
| 7-Deoxyrosenonolactone ( | 302 | C20H30O2 | [ | |
| [ | ||||
| [ | ||||
| [ | ||||
| 8 | 316 | C20H28O3 | [ | |
| 11β-Hydroxyrosenonolactone ( | 332 | C20H28O4 | [ | |
| Lyrosenonolactone ( | 332 | C20H28O4 | [ | |
| Rosenololactone ( | 318 | C20H30O3 | [ | |
| [ | ||||
| Rosololactone ( | 318 | C20H30O3 | [ | |
| [ | ||||
| [ | ||||
| [ | ||||
| Candidalactone ( | 332 | C20H28O4 | [ | |
| (3a1 | 284 | C19H24O2 | [ | |
| (3a | 300 | C20H28O2 | [ | |
| Ebractenoid A ( | 304 | C19H28O3 | [ | |
| [ | ||||
| Ebractenoid B ( | 304 | C19H28O3 | [ | |
| [ | ||||
| [ | ||||
| Ebractenoid R ( | 286 | C19H26O2 | [ | |
| Ebractenoid S ( | 316 | C20H28O3 | [ | |
| Ketone-containing rosane diterpenoids | ||||
| Hugorosenone ( | 302 | C20H30O2 | [ | |
| [ | ||||
| 18-Hydroxyhugorosenone ( | 318 | C20H30O3 | [ | |
| [ | ||||
| 18-Hydroxy-3-deoxyhugorosenone ( | 302 | C20H30O2 | [ | |
| 3β,19-Dihydroxy-1(10),15-rosadien-2-one ( | 318 | C20H30O3 | [ | |
| [ | ||||
| [ | ||||
| 5,15-Rosadiene-3,11-dione ( | 300 | C20H28O2 | [ | |
| Unidentified Argentine liverwort | [ | |||
| 33-Hydroxy-5,15-rosadien-11-one ( | 302 | C20H30O2 | Unidentified Argentine liverwort | [ |
| Anomallotusinsin ( | 320 | C20H32O3 | [ | |
| Euphebracteolatin C ( | 302 | C20H30O2 | [ | |
| Ebractenoid H ( | 318 | C20H30O3 | [ | |
| [ | ||||
| Ebractenoid I ( | 318 | C20H30O3 | [ | |
| Ebractenoid J ( | 300 | C20H28O2 | [ | |
| [ | ||||
| Ebractenoid O ( | 282 | C20H26O | [ | |
| Ebractenoid P ( | 284 | C20H28O | [ | |
| Ebractenoid Q ( | 288 | C20H28O | [ | |
| 3-Oxo-5,6-epoxyros-15-en-18--acetate ( | 360 | C22H32O4 | [ | |
| 3-Oxorosa-5,15-dien-18-acetate ( | 344 | C22H32O3 | [ | |
| Rosadiene and ester derivatives | ||||
| Rosa-5,15-dien-3α,18-diol ( | 304 | C20H32O2 | [ | |
| 18-Hydroxyrosa-5,15-dien--3α-acetate ( | 346 | C22H34O3 | [ | |
| 3α-Hydroxyrosa-5,15-dien--18-acetate ( | 346 | C22H34O3 | [ | |
| 18-Oxorosa-5,15-dien-3α-acetate ( | 344 | C22H32O3 | [ | |
| Rosa-1(10),15-dien-18-acid ( | 302 | C20H30O2 | [ | |
| [ | ||||
| Rosa-1(10),15-dien-18-acid methyl ether ( | 316 | C21H32O2 | [ | |
| Hugorosenol ( | 304 | C20H32O2 | [ | |
| [ | ||||
| Hugorosediol ( | 304 | C20H32O2 | [ | |
| Yuexiandajisu F ( | 304 | C20H32O2 | [ | |
| [ | ||||
| [ | ||||
| [ | ||||
| 19-Hydroxy-1(10),15-rosadiene ( | 288 | C20H32O | [ | |
| Engleromycenol ( | 288 | C20H32O | [ | |
| Ebractenoid C ( | 304 | C20H32O2 | [ | |
| [ | ||||
| [ | ||||
| Ebractenoid D ( | 304 | C20H32O2 | [ | |
| [ | ||||
| [ | ||||
| Ebractenoid E ( | 304 | C20H32O2 | [ | |
| Engleromycenolic acid A ( | 318 | C20H30O3 | [ | |
| Engleromycenolic acid B ( | 318 | C20H30O3 | [ | |
| Stachysrosane ( | 302 | C20H30O2 | [ | |
| [ | ||||
| Stachysrosane (138) | 286 | C19H26O2 | [ | |
| [ | ||||
| Aromatic rosane diterpenoids | ||||
| Euphebracteolatin A ( | 286 | C19H26O2 | [ | |
| [ | ||||
| [ | ||||
| Euphebracteolatin D ( | 286 | C19H26O2 | [ | |
| Euphebracteolatin E ( | 286 | C19H26O2 | [ | |
| Ebractenoid F ( | 286 | C19H26O2 | [ | |
| [ | ||||
| [ | ||||
| Ebractenoid G ( | 302 | C19H26O3 | [ | |
| Ebraphenol A ( | 268 | C19H24O | [ | |
| Ebraphenol B ( | 300 | C20H28O2- | [ | |
| [ | ||||
| Ebraphenol C ( | 270 | C19H26O | [ | |
| Ebraphenol D ( | 286 | C19H26O2 | [ | |
| Ebraphenol E ( | 300 | C20H28O2 | [ | |
| Ebracteolatas D ( | 300 | C19H24O3 | [ | |
| Ebralactone A ( | 296 | C20H24O2 | [ | |
| Dimeric rosane diterpenoids | ||||
| Yuexiandajisu D ( | 568 | C38H48O4 | [ | |
| [ | ||||
| [ | ||||
- —Deanship of Scientific Research (DSR)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBiological Activity of Diterpenoids and Biflavonoids · Plant biochemistry and biosynthesis · Bioactive Natural Diterpenoids Research
Introduction
Sustainable healthcare practices that prioritize natural remedies and environmental stewardship provide an integrated approach to health by utilizing natural products. The use of natural products and their derived preparations is steadily growing in a variety of industries, including agriculture, food, cosmetics, and veterinary and human medicines [1]. Their growing significance arises from the fact that they primarily rely on renewable biological resources, which aligns with the principles of a sustainable and circular bioeconomy, reducing environmental impact while encouraging innovation in green chemistry [2,3].
For thousands of years, medicinal plants have been used as natural sources of treatment for various illnesses [4-6]. In addition, their bioactive constituents have been proven to have significant therapeutic applications and serve as lead molecules for drug discovery [7-9]. Terpenoids are among the vast and varied groups of natural metabolites derived from five-carbon isoprene building blocks through the condensation and subsequent modification of isoprene units in various ways, including cyclization and/or oxygenation [1,10]. These compounds have been reported from plants, animals, algae, fungi, coral, and other organisms, including different types such as kaurene, daphnane, abietane, tiglilane, pimarane, dolabellane, labdane, jatrophane, dolastane, casbane, tonantzitlolone, and miscellaneous [11,12]. They are crucial for the growth and development of plants. Terpenoids are not only essential for plant growth and ecological adaptation but also have been widely utilized in dietary supplements, illness prevention, and general human wellness [8,13].
Diterpenoids represent one of the largest classes of terpenoids, which are biosynthesized from four isoprene units via geranylgeranyl diphosphate (GGPP) [14]. Among them, rosanoid diterpenoids, including ent-rosane and rosane, are relatively uncommon subclasses of tricyclic compounds characterized by a 6/6/6 fused-ring system (Figure 1).
Basic skeletons of rosane and ent-rosane diterpenoids
Ent-rosane diterpenoids have been reported from various plant families, including Euphorbiaceae, Lamiaceae, Alismataceae, Asteraceae, Velloziaceae, and Celastraceae, with the genera Euphorbia, Sagittaria, Vellozia, Trichogonia, and Maytenus recognized as particularly rich sources (Figure 2).
Distribution of rosanoid diterpenoids (ent-rosane and rosane) across different families based on reported literature from 1975 to 2025
This distribution indicates the chemotaxonomic significance and high-yielding taxa for further chemical and biological research. These plants are distributed across tropical and subtropical regions such as China, Brazil, Peru, Madagascar and Spain [15-18]. While rosane diterpenoids have been identified in liverworts, higher plants, and fungi (Figure 3).
Geographical distribution of the number of rosanoid diterpenoid-producing species per country
These compounds exhibit structural diversity and a range of biological activities, including anti-inflammatory, antimicrobial, cytotoxic, antiviral, anti-osteoclastogenic, and antiadipogenic effects. Despite their structural diversity and pharmacological properties, no comprehensive review exists that examines their occurrence, structural variations, and biological relevance. Accordingly, this review provides a comprehensive overview of rosane-type diterpenoids, including their sources, structural classification, and biological activities. By integrating data reported up to 2025, this review aims to serve as a valuable reference for future studies on the chemistry and biological activities of this group of diterpenoids. This article also highlights the significant translation gaps, particularly in the areas of pharmacokinetics, metabolic stability, and toxicity and suggests integrating early ADMET/DMPK studies with bioactivity-based lead selection of rosanoid diterpenoid potential candidates.
Search methodology
Search strategy
An extensive literature search was conducted through scientific databases (ScienceDirect, PubMed, Scopus, Web of Science, and Google Scholar) and publishers’ webpages (Elsevier, Wiley, ACS, RSC, Taylor & Francis, Springer, Bentham, Thieme, and MDPI), focusing on rosanoid diterpenoids (rosane- and ent-rosane-type diterpenoids) reported in the literature up to September 2025. The search was done using the keywords: “Rosanoid diterpenoids”, “Rosane diterpenoids”, “ent-Rosane diterpenoids”, “Rosanoid diterpenoids + isolation”, “Rosanoid diterpenoids + structure elucidation”, “Rosanoid diterpenoids + biological activity”, “Rosanoid diterpenoids + pharmacology”, “Rosanoid diterpenoids + cytotoxicity”, “Rosanoid diterpenoids + anti-inflammatory”, “Rosanoid diterpenoids + antimicrobial”, “Rosanoid diterpenoids + anticancer”, and “Rosanoid diterpenoids + natural sources”.
Inclusion and exclusion criteria
Published articles, reviews, and book chapters available in the above scientific databases that reported the isolation, structural elucidation, occurrence, and biological activities of natural rosanoid diterpenoids were included. However, the reported studies from non-peer-reviewed journals, irrelevant reports, and papers written in non-English without an English abstract were excluded.
Classification of rosanoid diterpenoids
Rosanoid diterpenoids are biosynthetically derived from geranylgeranyl diphosphate through an ent-copalyl diphosphate (ent-CPP) intermediate, followed by a series of cyclization, oxidation, and rearrangement reactions [19]. In general, rosanoid diterpenoids have a tricyclic C20 framework that undergoes substantial cyclization, accounting for the considerable structural variation documented in the literature. In the present review, the classification is mainly based on fundamental skeletal features that can be uniformly identified throughout reported studies, particularly (i) the number and position of the double bonds (unsaturation pattern), (ii) the extent and placement of oxygenation (e.g. hydroxyl, carbonyl, carboxyl, and ester substituents), and (iii) the presence of ring modifications such as lactone formation, aromatization, epoxidation, dimerization, and rearrangement/seco frameworks. Accordingly, the reported ent-rosane diterpenoids are categorized into major subclasses based on their unsaturation patterns within the tetracyclic skeleton, providing a practical framework for comparing related analogues (Table 1; [Figures 4 to 8](#fig004 fig005 fig006 fig007 fig008)). In parallel, rosane diterpenoids are shown to be structurally diverse, including less common aromatic and dimeric derivatives, rosadiene/ester analogues, and lactone- and ketone-containing metabolites (Table 2; [Figures 9 to 12](#fig009 fig010 fig011 fig012)). Overall, this classification goal is to present a logical structural map of rosanoid diterpenoids and to highlight the main modification patterns.
Chemical structures of saturated ent-rosane diterpenes (1-17).
Chemical structures of ent-rosane diterpenoids with Δ4(5) unsaturation in rings A and B (18-38).
Chemical structures of Δ1(10)-unsaturated (39-46) and Δ5(10) and Δ15(16)-unsaturated ent-rosanes (47-58)
Chemical structures of Δ5(10), Δ15(16)-unsaturated (59-67) and other unsaturated ent-rosanes (68-73)
Chemical structures of aromatic (74-77) and modified skeletons (78-86), and dimeric ent-rosanes (87-90)
Chemical structures of lactone- (91-104) and ketone-containing (105-108) rosane derivatives
Chemical structures of ketone-containing (109-120) rosane diterpenoids
Chemical structures of rosadiene and ester derivatives (121-138)
Chemical structures of aromatic (139-150) and dimeric (151) derivatives
Ent-rosane diterpenoids
In this study, 90 ent-rosane diterpenoids were discussed and categorized into six major classes based on the unsaturation pattern of their skeletons (Table 1).
Saturated ent-rosane diterpenoids
This group comprises hydrogenated or polyoxygenated ent-rosanes bearing hydroxyl, acetoxy, carbonyl, or carboxyl groups but lacking olefinic bonds. Common compounds include ent-rimuen-5-ol, ent-rimuen-3α,5β-diol, ent-rimuen-5,18-diol, and their acetate/tiglate derivatives, which were separated from Trichogonia salviaefolia and Trichogonia villosa MeOH extract by TLC impregnated with AgNO_3_ and SiO_2_ CC and identified using MS and NMR tools [20]. Compounds 7 and 8 were isolated from the root bark of Maytenus cuzcoina by Sephadex LH-20, SiO_2_ (CH_2_Cl_2_:Et_2_O of increasing polarity), and preparative HPTLC developed with n-hexane: Et_2_O (4:6). Their stereo-structures were elucidated using spectroscopic analysis, computational data, and the Riguera ester procedure (Figure 4) [21].
Compound 10 was isolated from the herb of Sagittaria pygmaea by silica-gel chromatography [23]. Compound 11 was isolated from the roots of Euphorbia ebracteolata [24]. Compounds 12 and 13 were isolated from Sagittaria sagittifolia and S. trifolia var. sinensis [18,25]. Santos et al. [22] isolated compounds 9 and 15-17 from the roots and heartwood of E. barbatum together with other hydroxylated analogues.
Ent-Rosane diterpenoids with δ4(5) unsaturation in rings A and B
This class includes ent-rosane diterpenoids characterized by a double bond between C-5 and C-6. Most of these compounds were isolated from the Lamiaceae, Euphorbiaceae, and Alismataceae families. For example, compounds 18 and 19 were identified from Vellozia candida (Velloziaceae) and Sideritis serata (Lamiaceae) by chromatographic separation of CHCl_3_ extracts on Al_2_O_3_ and SiO_2_ columns [27]. Compounds 20-22 were obtained from the aerial parts of C. niveus [28] (Figure 5). Omphalines B-E (23-26) were isolated from the stems of Omphalea oppositifolia [29]. These compounds retain the Δ^5^,^15^ diene skeleton but differ in the number and orientation of hydroxyl groups [29]. Compounds 27 and 28 are oxygenated Δ^5^,^15^-dienes isolated from Sideritis serata aerial parts [15]. Compound 29 was isolated from the aerial parts of E. milii [30], retaining the Δ^5^,^15^ pattern and possessing an oxygen bridge forming a lactone or enone system across C-3 and C-1 [30]. Candidenodiol (30) was obtained from the leaves of V. candida by Valente et al. [31]. Compounds 31 and 32 were isolated from Mallotus anomalus roots [32]. Compounds 33-38 are Δ^5^,^15^-unsaturated diterpenes isolated from the rhizomes of Sagittaria sagittifolia and S. trifolia var. sinensis [18,25].
Ent-Rosane diterpenoids with Δ1(10) unsaturation in rings A and B
This structural subclass of ent-rosane diterpenoids is characterized by a double bond at C-1(10) in ring A. Members of this group have mainly been isolated from species belonging to the Plagiochilaceae and Euphorbiaceae families.
Chromatographic separation of the ether extract of Plagiochila deltoidea aerial parts collected in New Zealand using SiO_2_ CC (n-hexane/EtOAc)/ HPLC (n-hexane/EtOAc 9:1) afforded compounds 39 and 40 (Figure 6) [33]. Later, they were isolated from E. fischeriana and E. ebracteolata [24,34]. Two new ent-rosane diterpenoids, 41 and 42, containing α,β-unsaturated ketone moieties, were isolated from Euphorbia pekinensis Rupr. by SiO_2_ and Sephadex LH-20 CC and elucidated based on NMR, HRMS, X-ray diffraction analysis, and the CD method [35]. Four further Δ^1^(^10^)-unsaturated ent-rosanes, 43-46, were obtained from the aerial parts of E. milii collected in Baoshan, Yunnan Province (China) [36].
Δ5(10), Δ15(16)-Unsaturated ent-rosane (Rosen-1(10),15-diene) diterpenoids
Compounds 47-54, isolated from the aerial parts of E. milii [30,36], represent oxygenated tricyclic ent-rosane diterpenes with variable oxidation at C-3, C-6, C-18, and C-19, displaying Δ^5^,^10^ and Δ^15^,^16^-unsaturation. Their structures were elucidated by extensive NMR, chemical methods, experimental and calculated electronic circular dichroism (ECD), and confirmed by single-crystal X-ray diffraction for euphominoid A, establishing the ent-configuration [30,36]. Further phytochemical investigation of E. milii from Menglun (Yunnan) led to the identification of compounds 56-58 [38]. Compound 56 features a 1-methyl-6-oxabicyclo[3.2.1]oct-2-ene motif in ring A, which was assigned by NMR, X-ray, and ECD analyses [38] (Figure 7). A study by Peng et al. [30] reported compounds 59-67 from Euphorbia milii, which incorporate aromatic acyl substituents, tetrahydrofuran rings, and 18-nor derivatives.
Other unsaturated ent-rosane diterpenoids
This group features diverse double-bond positions in rings A, B, and C. For example, compound 68 was obtained from the aerial parts of Blumea balsamifera (Asteraceae) [39], which retains the ent-rosane 15-ene core with additional double bonds at C-1-C10 and C-8-C14. Compounds 69 and 70 were isolated from the roots of E. fischeriana, collected in Anhui, China [34] and later identified in E. ebracteolata Hayata from Bozhou and Chuzhou [19,40]. Compounds 71 and 72 were reported from the stem, roots, and leaf sheaths of Vellozia candida Mikan (Velloziaceae), growing on the hillsides of Corcovado Mountain and the rocky coastal regions of Rio de Janeiro [16]. They possess OH groups at C-7 and C-12, and an epoxide bridge between C-5 and C-10. Compound 73 was isolated from the leaves and twigs of Omphalea oppositifolia, collected in Moramanga, Alaotra-Mangoro, Madagascar [29]. This compound is a 19-nor-ent-rosane derivative, lacking the terminal C-20 methyl group, with OH at C-2 and C-6 and Δ^4^, Δ^15^ double bonds.
Aromatic, modified skeletons, and dimeric ent-rosane diterpenoids
These compounds are characterized by aromatic rings within their tetracyclic carbon skeletons, resulting from the oxidative dehydrogenation of the ent-rosane framework, which produces conjugated systems (Figure 8).
Compound 74 was isolated from the roots of E. fischeriana collected in Anhui, China. It possesses an aromatic A-ring, bearing oxygenated substituents. Its structure was elucidated by NMR and X-ray analyses [34]. Compound 75 was obtained from the roots of E. nematocypha and elucidated by NMR, HRSM, ECD, and optical rotation analyses [41]. Two additional aromatic ent-rosane diterpenes, compounds 76 and 77, were reported from the aerial parts of E. milii [30].
Some ent-rosane diterpenoids exhibit structural modifications or rearrangements of the parent tetracyclic skeleton through oxidation, migration of double bonds, and partial alteration of the ring system. Compounds 78 and 79, isolated from the stems of E. neriifolia L. collected in Guangxi, China, possess oxidatively modified ent-rosane backbones [37]. Compound 80, obtained from the roots of E. fischeriana and E. ebracteolata, displays a partially rearranged and oxygenated ent-rosane nucleus [19,34,40]. Similarly, compound 81 was isolated from E. fischeriana using SiO_2_ CC and elucidated by NMR and X-ray [17]. Compounds 82-84 were obtained from the aerial parts of E. milii in Yunnan, China [26,30]. Compound 84 is a rare ent-rosane-type diterpenoid with a 5/7/6 skeleton. Likewise, 85 and 86, also isolated from E. milii, are unprecedented 7/7/6-fused tricyclic 5,10-seco-ent-RDs and possess a unique 11-oxabicyclo[4.4.1]undeca-1(10),5-diene moiety [38].
Compounds 87 and 88 were isolated from the roots of E. ebracteolata collected in Bozhou, Anhui Province, China. Both possess dimeric frameworks linked through oxygen bridges between abietane lactone and nor-rosane units [42]. Likewise, compounds 89 and 90 are rare ent-abietane-rosane diterpenoid heterodimers with oxygen-bridged structures also obtained from the roots of E. ebracteolata [43].
Rosane diterpenoids
In this study, 61 rosane diterpenoids were listed. Structurally, these diterpenoids possess notable variations in their skeletons, including lactones, aromatic analogs, simple hydroxyl or keto derivatives, and even dimeric skeletons, highlighting the biosynthetic versatility of this class (Table 2).
Lactone-containing rosane diterpenoids
Rosenonolactone-type compounds were the first to be described, including 91, 92, 94, and 97 [44,45]. These lactone metabolites were separated from plants (Lycoris aurea and Jatropha curcas) and fungi (Trichothecium roseum) [46,47] (Figure 9).
Ketone-containing rosane diterpenoids
Several keto derivatives have been identified (Figure 10). These include different Hugonia metabolites such as 105-107 [52,53] and108 [54].
Rosadiene and ester derivatives
Trichogonia species yielded rosadiene-type analogs, such as 121 and its acetate derivatives; 122, 123, and their ester-modified derivatives (Figure 11) [20]. Additional hydroxylated derivatives (e.g., 127-129 and 136-138) illustrate modifications of the rosadiene scaffold.
Aromatic and dimeric rosane diterpenoids
Several unusual rosane diterpenoids feature aromatic A-rings or rearranged skeletons. For example, 139-141 possess an aromatic A-ring system [19], while 144-148 and 150 [62] are characterized by additional aromaticity or rare lactonic modifications [61,62]. A dimeric rosane derivative 151, was reported from Euphorbia ebracteolate (Figure 12) [64].
Biological activities
Rosanoid diterpenoids exhibit diverse bioactivities, including anti-inflammatory, cytotoxic, enzyme inhibitory, antimicrobial, and metabolic modulatory capacities. These bioactivities are directly linked to their structural variations. Their main biological properties are outlined, with an emphasis on structure-activity relationships (SAR) and, when available, mechanistic insights.
Anti-inflammatory activity
Several studies have examined the anti-inflammatory properties of rosane-type diterpenoids. de las Heras et al. [65] reported that compound 27 remarkably prohibited COX-1- and 5-LOX-mediated release of Prostaglandin E2 (PGE_2_) and leukotriene C4, respectively, with a COX-2/COX-1 selectivity ratio of 3.15, indicating selectivity toward COX-1. Compound 68 demonstrated anti-inflammatory activity by significantly suppressing TNF-α secretion in LPS-stimulated macrophages without inducing cytotoxicity and while having no effect on IL-6 and NO [39]. Compounds 78 and 79 inhibited NO production in LPS-induced murine microglial BV-2 cells. Compound 78 showed an IC_50_ of 22.4 μmol L^-1^, compared to 2-methyl-2-thiopseudourea sulfate (SMT) (IC_50_ = 2.0 μmol L^-1^) [37].
Compounds 101-102, 108, 115, 127, 129, 132-134, 142, and 143, isolated from E. ebracteolata roots, inhibited LPS-stimulated NO production in RAW 264.7 macrophages with IC_50_ values ranging from 1.02 to 7.50 μmol L^-1^, compared to indomethacin (IC_50_ = 16.67 μmol L^-1^) and hydrocortisone (IC_50_ = 54.23 μmol L^-1^). Among them, 108 demonstrated more powerful NO inhibition (IC_50_ = 1.02 μmol L^-1^) than indomethacin (IC_50_ = 16.67 μmol L^-1^). Additionally, compounds 115 (IC_50_ = 2.76 μmol L^-1^), 142 (IC_50_ = 3.33 μmol L^-1^), and 143 (IC_50_ = 2.44 μmol L^-1^) displayed marked inhibitory effects [51].
Mechanistic study by Chun et al. revealed that 142 remarkably inhibited NO production (IC_50_ = 2.39 μg mL^-1^) and NF-κB activity (IC_50_ = 4.01 μg mL^-1^) in LPS-stimulated RAW 264.7 macrophages. This compound markedly suppressed iNOS mRNA and protein expression, prevented IκB-α phosphorylation and degradation, and inhibited nuclear translocation of NF-κB p65 and p50 subunits. Additionally, it downregulated IL-6 and IL-1β expression and inhibited phosphorylation of JNK, ERK1/2, IKKα/β, and Akt. Thus, 142 exhibited anti-inflammatory efficacy by inhibition of the NF-κB/MAPK/ PI3K/Akt signaling cascades [66]. Compound 130 exhibited potent inhibitory activity against NO production (IC_50_ = 2.91 mg μL^-1^) in LPS-induced RAW264.7 cells. It (doses 10 and 20 mg mL^-1^) suppressed iNOS mRNA expression [58].
Anti-osteoclastogenic activity
Compounds 60-65, characterized by aromatic ester substitutions, showed potent inhibition of RANKL-induced osteoclastogenesis (IC_50_s = 4.6 to 9.8 μmol L^-1^), compared to alendronate sodium (IC_50_ = 4.4 μmol L^-1^), with compound 62 being the most active (IC_50_ 4.6 = μmol L^-1^) [30].
Anti-adipogenic and lipid-lowering activity
Compounds 56-58 demonstrated potent antiadipogenic capacity and reduced triglyceride levels in 3T3-L1 preadipocyte differentiation assays (EC_50_s = 3.92 to 18.30 μmol L^-1^). Compound 57 was the most potent compound, revealing that the C-15 and C-16 hydroxyl groups play a crucial role in enhancing antiadipogenic activity [38].
Antiviral activity
Compounds 47-49 and 43 showed potential to inhibit EBV lytic DNA replication in P3HR-1 cells (EC_50_s ranging from 5.4 to 29.1 μmol L^-1^), whereas compound 48 showed the strongest inhibition (EC_50_ = 5.4 μM), comparable to (+)-rutamarin (EC_50_ = 5.4 μmol L^-1^). It was found that compounds with a 5,10-double bond exhibited more potent inhibitory activity than those with α,β-unsaturated carbonyl groups against EBV lytic replication [36].
Anticancer and cytotoxic activities
Several investigations have reported the cytotoxic potential of rosane diterpenoids against various tumour cell lines. Compounds 20 and 21 were evaluated for their cytotoxic activity using sulforhodamine B (SRB) assay in vitro against human cancer cell lines: U251 (human glioblastoma), PC-3 (human prostatic adenocarcinoma), K562 (human chronic myelogenous leukemia), HCT-15 (human colo-rectal adenocarcinoma), and MCF-7 (human mammary adenocarcinoma). Compound 21 inhibited 91.05 % of PC-3 (prostate cancer), 83.46 % of HCT-15 (colon cancer) and 73.16 % of MCF-7 (breast cancer) cells, with IC_50_s = 34.76, 41.93, and 77.08 μmol L^-1^, respectively, whereas 20 produced 77.68, 52.11, and 49.17 % inhibition of PC-3, HCT-15, and MCF-7. Both compounds interacted with NF-κB and STAT-3, suggesting that ent-rosane diterpenes, particularly compound 21, have promising anticancer potential through inhibition of NF-κB and STAT-3 signalling [28].
Ribosomal S6 kinase (RSK) is a key regulator of tumour cell survival and growth. Molecular docking revealed that compound 75 exhibited strong binding affinity for RSK with a binding energy of -36.1 kJ mol^-1^, compared to the reference LJH685 (-33.3 kJ mol^-1^) by forming three hydrogen bonds with Asp211, Lys100, and Asp148. Also, it showed cytotoxic activity against HCC 1806, CT26, and HeLa cells (IC_50_s = 16.96, 52.04 and 52.70 μmol L^-1^, respectively) [41].
Compound 89 displayed potent activity against HL-60, SMMC-7721, and MCF-7 (IC_50_s = 2.61, 4.08, and 8.17 μmol L^-1^, respectively), while 90 was moderately active (IC_50_s = 10.64 to 16.05 μmol L^-1^) [43]. Meanwhile, 105, separated from Hugonia castaneifolia, exhibited cytotoxicity in the brine shrimp lethality bioassay [52]. A study by Ding et al. [19] stated that 112 displayed selective cytotoxic capacities against HepG2 cells (IC_50_ = 14.29 μmol L^-1^) compared to cisplatin (IC_50_ = 7.04 μmol L^-1^), while 140 and 141 were moderately active (IC_50_ = 23.69 and 40.85 μmol L^-1^). Interestingly, the compounds with an α,β-unsaturated ketone moiety were more potent than their aromatic analogs, indicating that the conjugated enone functionality played a significant role in cytotoxicity. The new rosane, 149, obtained from E. fischeriana, demonstrated moderate cytotoxic efficacy against A549 lung carcinoma cells (IC_50_ = 22.03 μmol L^-1^). Annexin V staining showed that this compound induced marked apoptosis [61]. Fu et al. [64] revealed that the 18-nor-rosane-type dimeric diterpenoid 151 from Euphorbia ebracteolata, exhibited marked cytotoxicity toward HCT-8 colon carcinoma and Bel-7402 hepatocellular carcinoma cells (IC_50_s = 2.66 and 3.76 μmol L^-1^, respectively), compared to adriamycin (IC_50_s = 0.21 and 0.48 μmol L^-1^, respectively), while it showed weak effecttiveness against KB, A549, and BGC-823 cell lines, suggesting the dimerization and the 2,3-dihydro-1,4-benzodioxine bridge might enhance the selectivity toward specific tumour targets.
Anti-Glucosidase and anti-tuberculosis activity
Compound 87 showed moderate α-glucosidase inhibition (IC_50_ = 7.94 μmol L^-1^), while 88 inhibited Mycobacterium tuberculosis GlmU acetyltransferase (IC_50_ = 41.85 μmol L^-1^), a key enzyme involved in bacterial cell wall biosynthesis and a promising target for tuberculosis therapy [42].
Antibacterial activity
These diterpenoids were assessed for their antimicrobial and antimycobacterial activities. Liu et al. [23] reported that 10 exhibited mild antibacterial activity against Streptococcus mutans and Actinomyces viscosus (MIC = 125.0 μg mL^-1^). Compound 13 showed weak inhibition against S. mutans and A. naeslundii (MIC = 62.5 μg mL^-1^) [18]. Compound 37 was active against Actinomyces naeslundiis ATCC 12104 (MIC = 62.5 μg mL^-1^) [25]. Compounds 105-107 exhibited antifungal activity against Cladosporium cucumericum (inhibition zone diameters 38.47, 63.59 and 12.56 mm^2^, respectively), with 107 demonstrating the highest activity (inhibition zone 12.56 mm^2^) [53]. Yu et al. [40] reported that 118 showed moderate inhibition of Mycobacterium tuberculosis H_37_Rv (MIC = 18 μg mL^-1^). Compound 118 moderately inhibited GlmU enzyme (IC_50_ = 12.5 μg mL^-1^). GlmU, a bifunctional N-acetylglucosamine-1-phosphate uridyltransferase/acetyltransferase enzyme essential for mycobacterial cell wall biosynthesis [40].
Enzyme inhibitory and other activities
Some rosane-type diterpenoids demonstrate enzyme-inhibitory properties. For example, compounds 91, 92 and 97, isolated from Trichothecium roseum, were found to inhibit cholesteryl ester transfer protein (CETP) in vitro (IC_50_s = 31, 65 and 60 μg mL^-1^, respectively) [45]. The aromatic derivatives 144-147 and 150, isolated from the roots of Euphorbia ebracteolata, demonstrated notable pancreatic lipase inhibition, a key enzyme involved in dietary fat absorption. Among them, 144 was the most potent (IC_50_ 1= 1.0 μg mL; Ki = 1.8 μg mL^-1^), compared to lovastatin (IC_50_ = 0.24 μmol L^-1^) [62]. Additionally, 148 showed a moderate lipase inhibitory effect (IC_50_ = 12.5 μmol L^-1^) [63]. Besides, 135 obtained from Engleromyces goetzii, significantly inhibited CETP activity (IC_50_ 7.55 μmol L^-1^) [48]. Compound 105 showed larvicidal activity against Anopheles gambiae mosquito larvae (LC_50_s = 0.3028, 0.0674 and 0.0582 mg mL^-1^ at 24, 48 and 72 h exposure time, respectively) [53]. From Stachys parviflora, 137 and 138 demonstrated pronounced antidiarrheal effects in castor oil-induced diarrhoea models in mice [59]. Also, 137 and 138 display sedative and skeletal muscle relaxant properties in behavioural assays. Docking studies revealed their potential interaction with the GABA-A receptor [60].
Critical perspective on biological activities
Overall, the reported bioactivities of rosanoid diterpenoids are mainly anti-inflammatory and cytotoxic/ /anticancer, with additional studies supporting enzyme inhibition, antimicrobial, antimycobacterial, antiviral, and metabolic modulatory (antiadipogenic/lipid-lowering and anti-osteoclastogenic) effects. In many cases, the activity was linked to specific structural features, including α,β-unsaturated carbonyl motifs, aromatic ester substitutions, and oxygenation patterns that affected potency across assays. Mechanistic support is available for selected examples, which strengthens the biological relevance of some lead structures.
Rosane-type diterpenoids exhibited promising anti-inflammatory effects in various experimental models, particularly by inhibiting NO production in LPS-stimulated macrophages and suppressing COX/LOX-related inflammatory mediators. Notably, compounds 101-102, 108, 115, 127, 129, 132-134, 142 and 143 were the most active, showing strong NO inhibition with IC_50_ values of 1.02 to 7.50 μmol L^-1^. Additionally, the compound 142’s effect was linked to NF-κB/MAPK/PI3K/Akt signalling pathways. In the anti-osteoclastogenic assay, compounds 60-65 with aromatic ester substitutions markedly inhibited RANKL-induced osteoclastogenesis (IC_50_ = 4.6 to 9.8 μmol L^-1^), supporting aromatic ester-bearing rosanoids could be promising for further optimization in bone-resorption-related models. Similarly, compounds 56-58 lowered accumulation of triglycerides in 3T3-L1 cells (EC_50_ = 3.92 to 18.30 μmol L^-1^), suggesting that C-15 and C-16 hydroxyls play a crucial role in enhancing antiadipogenic activity. In antiviral assays, compounds 47-49 and 43 suppressed EBV lytic DNA replication, demonstrating that the analogs with a 5,10-double bond were more potent than those with α,β-unsaturated carbonyl groups, suggesting that unsaturation can affect antiviral activity. In the cytotoxic and anticancer activities, the reported findings revealed that α,β-unsaturated ketone-containing rosanoids were more potent than aromatic analogs. Therefore, this underscores the need to interpret the above findings with deep mechanistic studies to confirm these conclusions.
Limitations of the reported studies on rosanoid diterpenoids
Most data arise from in vitro studies, which are usually carried out on a limited number of cell lines, microbes, or enzyme assays, with a narrow concentration range, and sometimes without direct comparison with reference drugs. Thus, it is difficult to assess the actual effectiveness, selectivity, and relevance of the described effects. Several studies have employed short-term models and single-cell protocols to investigate the anti-inflammatory, anti-osteoclastogenic, and anti-adipogenic/lipid-modulating effects. In many cases, the inhibition of mediators such as NO, cytokines, or RANKL-mediated pathways has been established; however, there is little or no detailed mechanistic investigation (e.g., at the protein and gene expression levels). Cytotoxic and anticancer effects are commonly observed with certain rosanoid diterpenoids, demonstrating IC_50_ values in the micromolar range. However, these findings typically arise from assays on a limited panel of cancer cell lines, with minimal comparison of toxicity towards healthy cells. In addition, there is a lack of in vivo studies, pharmacokinetic, and comprehensive toxicity investigations. The inconsistency in experimental designs across studies further complicates comparisons of outcomes. The current data are useful for identifying potential rosiglitazone frameworks; however, they are not yet reliable enough to support robust conclusions regarding their therapeutic applications or feasibility for drug development.
Conclusion and future research directions
Rosanoid diterpenoids are a structurally distinct but very small subgroup of diterpenes, which include ent-rosane and rosane derivatives, distinguished by a 6/6/6 tricyclic scaffold and a variety of oxidation patterns, ring modifications, and dimeric frameworks. This review summarizes the reported metabolites from liverworts, fungi, and higher plants, particularly from the family Euphorbiaceae plants. These compounds were assessed for multiple biological activities, including anti-inflammatory, anti-osteoclastogenic, anti-adipogenic, lipid-modulating, cytotoxic, antimicrobial, enzyme-inhibitory, and neuroactive effects. Accordingly, rosanoid diterpenoids may be considered promising scaffolds for further assessment as drug-like leads. Therefore, future research should primarily focus on expanding the sources of rosanoid diterpenoids by investigating understudied species and utilizing advanced analytical and metabolomics techniques; investigating their structure-activity relationships through standardized in vitro assays and semi-synthetic modification of key scaffolds; and conducting in vivo experiments to evaluate their pharmacokinetics, toxicity, and efficacy in relevant disease models. In parallel, integrating ADMET/DMPK profiling for the most active scaffolds would strengthen prioritization and reduce the risk of overinterpreting isolated assay results. In addition, research aimed at understanding molecular mechanisms will play a pivotal role in ascertaining the relevance of these compounds’ therapeutic potential.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Drasar P.B.Khripach V.A.. Growing importance of natural products research. Molecules. 25 (2020) 6. https://doi.org/10.3390/molecules 25010006 10.3390/molecules 25010006 PMC 698315331861374 · doi ↗ · pubmed ↗
- 2Mohamed G.A.Ibrahim S.R.M.. Natural occurring 2-(2-phenylethyl) chromones, structure elucidation and biological activities. Natural Product Research. 29 (2015) 1489-1520. https://doi.org/10.1080/14786419.2014.991323 10.1080/14786419.2014.99132325529202 · doi ↗ · pubmed ↗
- 3Mohamed H.M.Mohamed G.A.Abdallah H.M.Ibrahim S.R.M.. Harnessing Aspergillus fumigatus for sustainable development: biotechnological and industrial relevance. Journal of Pharmacy & Bioallied Sciences 17(17) (2025) 36-44. https://doi.org/10.4103/jpbs.jpbs_883_25 10.4103/jpbs.jpbs_883_2540859992 PMC 12373383 · doi ↗ · pubmed ↗
- 4Ibrahim S.R.M.Mohamed G.A.. Litchi chinensis: medicinal uses, phytochemistry, and pharmacology. Journal of Ethnopharmacology 174 (2015) 492-513. https://doi.org/10.1016/j.jep.2015.08.054 10.1016/j.jep.2015.08.05426342518 · doi ↗ · pubmed ↗
- 5Khayat M.T.Alharbi M.Ghazawi K.F.Mohamed G.A.Ibrahim S.R.M.. Ferula sinkiangensis (Chou-A Wei, Chinese Ferula): Traditional uses, phytoconstituents, biosynthesis, and pharmacological activities. Plants 12(12) (2023) 902. https://doi.org/10.3390/plants 12040902 10.3390/plants 1204090236840251 PMC 9958602 · doi ↗ · pubmed ↗
- 6Abdallah H.M.Mohamed G.A.Ibrahim S.R.M.. Lansium domesticum-A fruit with multi-benefits: Traditional uses, phytochemicals, nutritional value, and bioactivities. Nutrients. 14(14) (2022) 1531. https://doi.org/10.3390/nu 14071531 10.3390/nu 1407153135406144 PMC 9002712 · doi ↗ · pubmed ↗
- 7Ibrahim S.R.M.Fadil S.A.Fadil H.A.Hareeri R.H.Abdallah H.M.Mohamed G.A.. Ethnobotanical uses, phytochemical composition, biosynthesis, and pharmacological activities of Carpesium abrotanoides L. (Asteraceae). Plants 11(11) (2022) 1598. https://doi.org/10.3390/plants 11121598 10.3390/plants 1112159835736748 PMC 9230109 · doi ↗ · pubmed ↗
- 8Ibrahim S.R.M.Altyar A.E.Sindi I.A.El-Agamy D.S.Abdallah H.M.Mohamed. G.A. Mohamed S.G.A.. Kirenol: A promising bioactive metabolite from Siegesbeckia species: A detailed review. Journal of Ethnopharmacology 281 (2021) 114552. https://doi.org/10.1016/j.jep.2021.114552 10.1016/j.jep.2021.11455234438028 · doi ↗ · pubmed ↗