The T-cell receptor repertoire of wild mice
Jacob A Cohen, Simon Hunter-Barnett, Gayathri Nageswaran, Suzanne Byrne, Gemma Freeman, Matthew V Cowley, Benny Chain, Mark Viney

TL;DR
This study explores how the T-cell receptor repertoires of wild mice vary with age, sex, and location, revealing insights into their immune system's adaptability.
Contribution
The study provides a novel description of T-cell receptor repertoires in wild mice, linking immune diversity to environmental exposure.
Findings
Wild mice maintain large TCR repertoires with consistent richness across age and sex.
TCR diversity is influenced by a sex-by-age interaction, indicating more abundant clones in older mice.
The findings suggest wild mice balance broad immune response capacity with strong responses to persistent pathogens.
Abstract
Wild animals live in a pathogen-rich environment and are normally infected with a wide range of micro- and macro-parasites. Wild animals’ T cells are central to the effectiveness of their adaptive immune response in ameliorating the effect of these infections. Here, we have investigated the T-cell receptor (TCR) repertoire of wild mice to investigate how it varies in animals of different ages and sex and from different sites. We sequenced the TCR alpha and beta chains of CD4+ and CD8+ T cells of 65 wild Mus musculus domesticus from two UK sites. We analysed repertoire richness and diversity finding that wild mice have large TCR repertoires. Repertoire richness, which measures the breadth of the repertoire, was not significantly affected by mouse age or sex, suggesting that wild mice maintain the capacity to respond to novel antigens throughout their lives. In contrast, repertoire…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
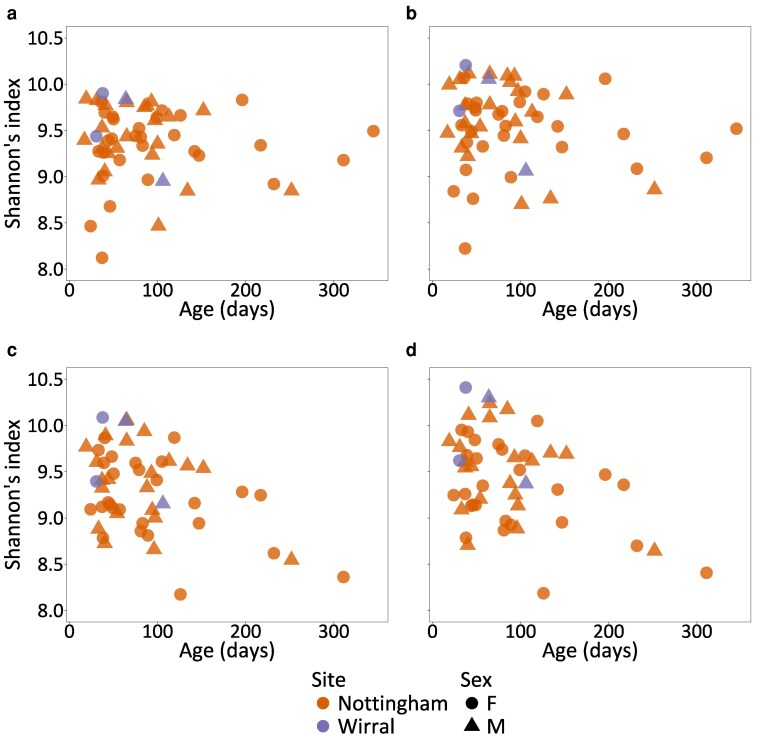 Figure 1
Figure 1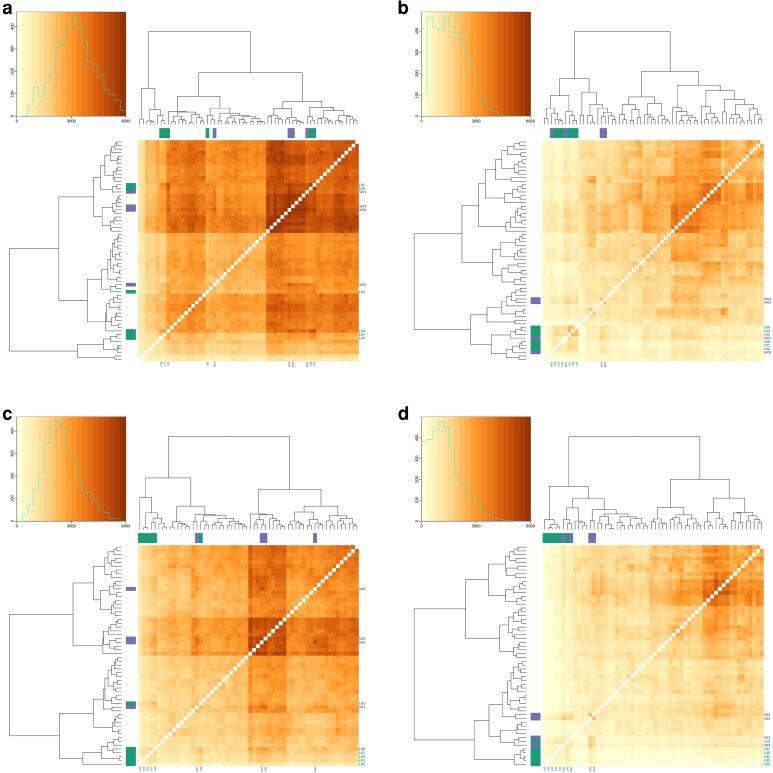 Figure 2
Figure 2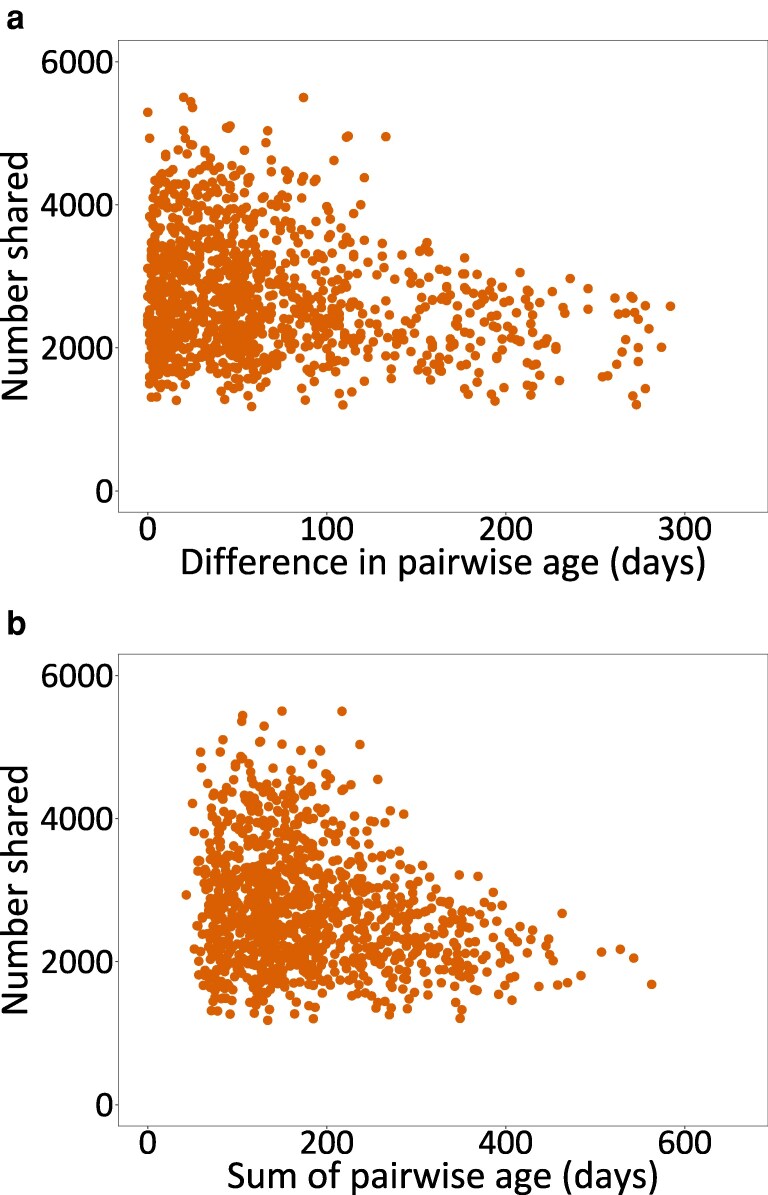 Figure 3
Figure 3- —NERC Exploring the Frontiers
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsT-cell and B-cell Immunology · Parasitic Infections and Diagnostics · Immunotoxicology and immune responses
Introduction
The vertebrate immune system protects individuals from infection and disease. Most of our description and understanding of this system comes from study of the laboratory mouse, but laboratory mice are not exposed to the infections and other threats of the natural environment, which is the context in which the immune system and its function have evolved. There has been recent interest in studying the immune system and immune state of wild mice and other rodents [1], finding that in wild mice the immune system is in an altered, often more activated, state compared with that of laboratory animals [2–4]. Other studies have used rewilded mice, that is laboratory mice released into outdoor enclosures, and have found that they have an altered immune response that, for example, makes them susceptible to a parasitic nematode infection, while laboratory-maintained mice are resistant to it [5]. Transplanting laboratory mice with wild mouse microbiomes also brings about profound immunological changes, reducing inflammation and, for example, increasing susceptibility to infection with influenza virus [6]. The unifying thread from these studies is that laboratory mice are underexposed to infections compared with wild/rewilded/wild microbiome-exposed mice, and that greater infection stimulates the immune system into a different functional state, compared with laboratory mice.
B and T cells are the principal players of the adaptive immune response and the dominant cell types in the lymphatic system, together (in humans) accounting for 40% of immune cells (∼25% T cells and ∼15% B cells) [7]. T cells bind to antigenic peptide molecules that cause the T cells to proliferate and then to act directly against the antigenic source. Antigens are perceived by heterodimeric (in mice and humans principally α and β chains) antigen-specific T-cell receptors (TCRs), coded for by somatically recombined TCR genes. Each pathogen antigen is recognized by a small number of specific clones of T cells, so that an individual animal contains a large, diverse repertoire of T-cell clones. T-cell responses to individual pathogens are idiosyncratic [8]; for example, in people infected with Mycobacterium tuberculosis, individuals’ CD4^+^ T cells recognized an average of 24 different epitopes, but individuals varied extensively in the epitopes that they recognized [9].
Next generation parallel sequencing of TCR-coding genes and transcripts has been used to investigate the TCR-coding sequences present among circulating T cells in people and in laboratory mice. Theoretically, the TCR repertoire is enormous: in people potentially up to 10^15^ alpha and beta TCRs, though reduced to some 10^13^ after thymic selection, and then absolutely limited by the number of T cells present in an individual, some 10^12^ in a person and 10^8^ in a laboratory mouse [10, 11]. Sequence-based analyses are therefore only sampling a small fraction of the actual T cells present and so only sampling a small sub-fraction of the TCR repertoire [12].
To date, most analyses of the TCR repertoire have been of people, though with some study of laboratory mice. Many studies have confirmed that people and laboratory mice have large TCR repertoires. The clone size distribution of the repertoire is approximated well by a power law, with the vast majority of TCRs found at very low frequency (typically observed once in a sample), but a proportion of large clones taking up a disproportionate part of the overall repertoire [13, 14]. These distribution profiles can vary quantitatively between CD4^+^ and CD8^+^ cells, between naive and memory cells, and as individuals age [13, 15]. Specifically, as people age there is a reduction in overall TCR diversity, driven in part by a decrease in the number of TCR-defined clones in naive, but not in memory CD4^+^ cells, with similar changes in CD8^+^ cells, though they have a comparatively smaller TCR repertoire [8, 16 ]. The decrease in diversity of memory cells is driven by an increase in the relative abundance of some clones; indeed, in some individuals, individual clones specific to a chronic virus such as Cytomegalovirus can occupy a significant proportion of the total repertoire [17, 18].
Here, we sought to investigate the TCR repertoire of wild mice to complement other studies that have investigated wild mouse immune state. Animals’ exposure to pathogens and to other antigens drives the expansion of specific T-cell clones, such that individuals may vary in their TCR repertoire because of differences in their lifetime of exposure, so that the TCR repertoire may vary in mice of different ages. Male and female mice also live differently, with females living and breeding communally, but males being more solitary [19, 20], and this may affect exposure to infection such that there are sex differences in the TCR repertoire. Therefore, here we have investigated if, and how, the TCR repertoire varies among wild mice of different sex and age and among those obtained from different sites. We present these data as a first analysis of wild mouse TCR repertories that is particularly relevant to eco-immunology, but also to laboratory mouse immunology.
Materials and methods
Animals
We live-trapped mice, Mus musculus domesticus between May and September 2023 at two sites: (i) a pig farm in Nottinghamshire and (ii) a dairy farm on the Wirral peninsula, both in the UK. Mice were killed by cervical dislocation, sexed, and weighed. We aged mice by determining the mass of their dried eye lenses after Rowe et al. 1985 [21]; this method has been validated against mice of known age. We also analysed the repertories of six female CD-1 laboratory mice.
Splenocytes, fluoresence-activated cell sorting (FACS), and isolation of RNA
From each mouse, we aseptically isolated the spleen, determined its mass, and then passed it through a 70 μm sieve to create a single-cell suspension. We resuspended the splenocytes in blood cell lysis buffer (Invitrogen) held at room temperature for 5 min, supplemented with an excess of phosphate buffered saline (PBS), sedimented the cells by centrifugation, and washed them in PBS by resuspension and sedimentation, after which the cells were resuspended in 5 ml of staining buffer (BioLegend). We determined the number of splenocytes using a haemocytometer. We removed a portion of the cells as the FACS negative control. To the remainder we blocked Fc receptors using TruStain FcX™ PLUS on ice for 10 min and then stained the cells by addition of the following conjugated antibodies: anti-CD3-PE, anti-CD4-FITC, and CD8-APC for 20 min in the dark, after which the cells were washed twice in staining buffer. Immediately before FACS the cells were additionally stained with 4′,6-diamidino-2-phenylindole (DAPI).
Samples were analysed and separated using a FACSAria III Cell Sorter (BD Bioscience, Oxford, UK). The gating strategy was incorporated from dot plots with the following hierarchy: scatter gates (side scatter/ forward scatter [SSC/FCS]) to identify mammalian cell clusters, distinct from debris with low particle size and granularity that were expected to be excluded from the gate. A singlet gate was applied (FSC-height/FSC-area [FSC-H/FSC-A]) to remove doublets/multiplets that deviated from cluster linearity. From the singlet population, particles with a negative signal towards the DAPI stain were considered membrane-intact and hence gated as live cells. The CD3^+^ population was used to identify T cells, and finally CD4^+^ (negative to CD8) or CD8^+^ (negative to CD4) to detect and sort corresponding T-cell subsets (Supplementary Material S1). On-site data visualization, gating, and sort commands were performed using FACSDiva software, version 8.1.
The resulting CD4^+^ and CD8^+^ sorted cells were concentrated by centrifugation and the supernatant was removed and RLT buffer (QIAGEN) with 1% (v/v) 2 mercaptoethanol was added to the pellet of cells and mixed by pipetting to form a lysate, which was then processed through a QIAshredder homogenizer spin column following the manufacturer’s methods, with the resultant RNA stored at −80°C.
TCR sequencing and analysis
From the isolated RNA, TCR alpha and beta genes were sequenced using an established quantitative sequencing pipeline, which uses unique molecular barcodes to correct for polymerase chain reaction (PCR) bias and sequencing errors [13, 22]. The FASTQ files were processed and annotated using an in-house, open source computational pipeline Decombinator V4 [23, 24]. We excluded any TCR nucleotide sequences that did not contain a CDR3 sequence.
Data analysis
Each TCR nucleotide sequence defines a different T-cell clone, where the number of occurrences of an identical nucleotide sequence is a measure of the abundance of that T-cell clone. Different TCR nucleotide sequences can code for the same TCR amino acid sequence. In the analyses that follow we therefore refer to, and differentiate, nucleotide-defined TCRs and amino acid–defined TCRs. Amino acid–defined TCR sequences were the predicted translation of the concatenation of the nucleotide sequences of the variable gene segment (V), joining gene segment (J), and CDR3 region.
We quality controlled the amino acid–defined TCR sequence data by examining the number of sequences obtained for alpha and beta chains of CD4^+^ and CD8^+^ cells and then excluding data for a chain where there were fewer than 5000 TCR molecules that passed our quality control for that chain. We also examined the frequency distribution of amino acid sequences and fitted a power law distribution to it to generate an alpha coefficient and then examined the relationship of this with wild mouse age. To test whether the frequency distribution of amino acid sequences for individual mice matched the power law distribution, we used the Kolmogorov–Smirnov test with 500 simulations. Mice were classified into three categories based on the resulting P values: fits power law (P > 0.1), borderline fit to power law (0.1 > P > 0.05), and does not fit power law (P < 0.05). Results were summarized for each receptor type. The slope of the distribution did not vary with age.
We define TCR repertoire size as the sum of the number of occurrences of all amino acid–defined TCR sequences, separately for alpha and beta genes from CD4^+^ and CD8^+^ cells. We also calculated the number of unique amino acid–defined TCR sequences for each of the alpha and beta chains of CD4^+^ and CD8^+^ cells.
We calculated the richness of the TCR repertoire as the number of unique amino acid–defined TCRs as a proportion of the repertoire size, which corrects for the depth of sequencing of the samples. For the wild mice, we tested for differences in the richness of the TCR repertoire between the alpha and beta chains of CD4^+^ and CD8^+^ cells using a gamma-distributed generalized linear model (GLM) with TCR richness as the response variable and receptor type (i.e. CD4^+^ alpha, CD4^+^ beta, CD8^+^ alpha, and CD8^+^ beta), Sample site (Nottingham vs. Wirral), Age, Sex, and an Age × Sex interaction as explanatory variables. We calculated estimated marginal means (EMMs), which corrects for multiple comparisons, from the GLMs to test post hoc for differences between receptor types.
We calculated two diversity indices of the wild mouse TCRs: Shannon’s diversity index and Simpson’s diversity index. These metrics have recently been identified as being appropriate to quantify richness and evenness in TCR repertories [25]. For both, we down-sampled each mouse’s repertoire to that of the smallest, relevant repertoire (i.e. CD4^+^ alpha, CD4^+^ beta, CD8^+^ alpha, and CD8^+^ beta) to account for differences in repertoire size between mice. To do this, we first replicated all TCR amino acid–defined sequences by their number of occurrences, then randomly sampled this list without replacement to the size of the smallest, relevant repertoire, and then re-compiled the TCR amino acid–defined sequences with their total number of occurrences. This down-sampling process was done separately for the two diversity measures.
Shannon’s diversity index was calculated as:
where R is the repertoire size and the proportional abundance of the TCR amino acid–defined sequence.
Simpson’s diversity index was calculated as:
where R is the repertoire size and is the proportional abundance of the TCR amino acid–defined sequence. Analyses were conducted as for Shannon’s diversity index. We log transformed Simpson’s diversity index as:
Thus, larger values indicate higher diversity.
We tested for differences in Shannon’s and Simpson’s diversity among the four receptor types (above) using a GLM (Gaussian distribution) with Shannon or Simpson’s index as the response variable and Receptor type, Sample site, Age, Sex, and an Age × Sex interaction as explanatory variables. We calculated EMMs from the GLMs to test post hoc for differences between (i) receptor types and (ii) diversity indices in male and female mice.
To investigate the degree of sharing of amino acid–defined TCR sequences between all pairwise combinations of the wild and laboratory mice, we first down-sampled each mouse’s TCR amino acid–defined repertoire to that of the smallest, relevant repertoire, as described above. We then compiled these repertoires into lists of unique amino acid–defined TCRs and performed pairwise comparisons between mice to determine the number of shared amino acid sequences between mouse pairs. From this, we generated heatmaps of the degree of sharing, supplemented with dendrograms calculated from distance matrix data, which then arranged more similar individual mice together. We also examined these data using a principal coordinate analysis (PCoA) by calculating a re-scaled pairwise dissimilarity (with the most similar pair having a dissimilarity of 0), which was used in a weighted PCoA separately for alpha and beta chains of CD4^+^ and CD8^+^ cells.
To investigate whether mouse sex and age affected the extent to which mice shared amino acid–defined TCR sequences, we generated pairwise sharing data for all combinations of mice; we restricted this to only the Nottingham-derived mice to avoid any potentially effects of sample site. We then tested the effects of (i) pairwise mouse sex (same sex vs. different sex), (ii) pairwise difference in mouse age, and (iii) pairwise sum of mouse age on the number of pairwise shared amino acid–defined TCR sequences with a GLM (Gaussian distribution). The response variable for the GLM was the number of pairwise shared amino acid–defined TCR sequences, while the explanatory variables were Receptor type, pairwise Sex difference, pairwise Age difference, pairwise Age sum, a pairwise Age difference × receptor type interaction, and a pairwise Age sum × Receptor type interaction. We calculated slope estimates for the explanatory variables from the GLM using EMMs to test post hoc for differences between receptor types.
We conducted all data analysis in R (R Core Team 2025 [26]). Model diagnostics were carried out using the ‘DHARMa’ package [27]. EMMs post hoc testing was carried out using the ‘emmeans’ package [28]. PCoAs were conducted using the ‘vegan’ package [29]. In addition, we used the ‘dplyr’ [30], ‘stringr’ [31], ‘purrr’ [32], ‘tidyr’ [33], ‘poweRlaw’ [34], ‘here’ [35], and ‘modeest’ [36] packages. Plots were generated in R using packages ggplot2 [37], ‘gplots’ [38], ‘showtext’ [39], ‘patchwork’ [40], ‘ggpubr’ [41], and ‘ggbeeswarm’ [42].
Results
Animals
We caught 69 mice (30 male and 39 female), 65 from the Nottingham site and 4 from the Wirral site. The average age of the wild mice was 13 weeks [standard deviation (SD) 10, median 9, range 2–49]; average body mass was 15.5 g (SD 4, median 11, range 7–27). Among the wild mice, the spleens had an average mass of 53 mg (SD 33, range 10–170), yielding an average of 16 million splenocytes (SD 13, range 8.7 × 10^4^–5.1 × 10^7^]. There was a positive relationship between mouse age and spleen mass (R^2^ = 0.3, P < 0.0001) and number of splenocytes (R^2^ = 0.13, P < 0.0025) and between spleen mass and number of splenocytes (R^2^ = 0.49; P, 0.0001) (Supplementary Material S2). Previous comparisons of wild and laboratory mice have found that wild mice are smaller than laboratory mice, and that their spleens are absolutely smaller (approximately one third of the mass) and with commensurate fewer cells, but also proportionally smaller (compared with body mass), compared with those of laboratory mice [3].
The FACS of splenocytes resulted in an average of 8 million CD3^+^ T cells per wild mouse (SD 8 million, range 1.1 × 10^4^–4.9 × 10^7^). There were proportionally more CD4^+^ cells than CD8^+^ cells, with an average of 62% (SD 12, range 34–82), giving an average CD4^+^:CD8^+^ ratio of 2.4 (SD 1.4, range 0.2–6.9). For four female mice from the Nottingham site insufficient cells were collected for TCR sequence analysis, refining our sample size to 65 (30 male and 35 female) wild mice and 6 lab mice.
TCR sequences
To quality control the data, we examined the number of alpha and beta chain coding sequences for CD4^+^ and CD8^+^ cells separately and excluded the data for one cell type if there were fewer than 5000 TCR molecules of either the alpha or beta chain, which resulted in the exclusion of 9 and 13 mice, respectively. This resulted in final sample sizes of 62 (56 wild and 6 lab) for CD4^+^ alpha and beta, and 58 (52 wild and 6 lab) CD8^+^ alpha and beta chain TCR sequences that were analysed further.
After our data quality control, the minimum TCR repertoire size among the wild mice was 54 908, 58 772, 66 881 and 64 224, and for the laboratory mice, the minimum TCR repertoire size was 68 637, 79 052, 49 379, and 75 640 for CD4^+^ alpha and beta and CD8^+^ alpha and beta chains, respectively. We examined the frequency distribution of the amino acid–defined TCR sequences, which showed that many of these followed a power law distribution, which has frequently been observed on other TCR datasets (Supplementary Material S3).
TCR repertoire richness and diversity
We investigated the richness of the TCR repertoire, finding that among the wild mice beta chain richness was greater than that of the alpha chain, for both within and between CD4^+^ and CD8^+^ cell comparisons (Supplementary Material S4). The comparatively larger repertoire of beta chains, observed previously in human T cells [13], occurs because they are encoded by recombination of V, D, and J genes, unlike the V and J recombination of alpha chain coding genes.
There was no significant effect of wild mouse Age, Sample site, Sex, or Age × sex on the TCR repertoire richness (all |z| < 1.2, P > 0.26; Supplementary Material S4). However, the TCR diversity measured by Shannon’s index (after down-sampling each mouse’s repertoire to that of the smallest, relevant repertoire) differed significantly between the CD4^+^ beta and the CD8^+^ alpha chains (Fig. 1; Supplementary Material S5), and there were significant effects of Sex (t = 2.79, P = 0.0058), Sample site (t = 2.76, P = 0.0063), Sex × Age (t = −2.07, P = 0.0396), and a marginally non-significant effect of Age (t = −1.78, P = 0.0762) with older mice tending to have a lower diversity. The difference between Shannon’s index of female and male mice was marginally non-significant (effect size = −0.0996, P = 0.0859). We obtained broadly similar results using Simpson’s diversity index though with more significant differences in diversity between receptor types (Supplementary Material S6), significant effects of Sex (t= 2.02, P = 0.0452), and Age (t = −3.97, P = 9.91 × 10^−5^), with older mice having lower diversity (Supplementary Material S7). The difference between Simpson’s index of female and male mice was also marginally non-significant (effect size = −0.187, P = 0.0884).
Alpha diversity measured by Shannon’s index of wild mouse, amino acid–defined TCRs for (a) CD4+ alpha, (b) CD4+ beta, (c) CD8+ alpha, and (d) CD8+ beta chains.
TCR networks
We investigated how much different wild and laboratory mice shared the same amino acid–defined TCR sequence, which we did by counting the number of common sequences between every pairwise combination of mice. This showed that there was considerable sharing of TCR sequences, with this greatest for alpha chains (5.9 and 5.2% for CD4^+^ and CD8^+^, respectively) than for beta chains (2.9 and 2.2% for CD4^+^ and CD8^+^, respectively) (Fig. 2; Supplementary Material S8). We asked how many TCR sequences were public, which we defined as those shared among ≥75% of the mice (both wild and laboratory). We found that this ranged from 157 (for CD8^+^ beta) to 1113 (for CD4^+^ alpha), but that this was always <0.25% of unique TCRs (Supplementary Material S8). Previous studies of public TCR sequences in 28 laboratory mice found that ∼0.5% of sequences were shared among 75% of them [43]. A PCoA analysis showed that there were differences in the sharing of sequences among mice from the different wild sampling sites, and between wild mice and laboratory mice (Supplementary Material S9).
The sharing of amino acid–defined TCR sequences among wild and laboratory mice, with the colour scale showing the number of shared sequences for (a) CD4+ alpha, (b) CD4+ beta, (c) CD8+ alpha, and (d) CD8+ beta chains. Against the plots, purple indicates wild mice from the Wirral site and green laboratory mice; mice where no colour is shown are wild mice from the Nottingham site. In all plots, the brown scale is the same. In each, the inset plot shows a frequency distribution of the number of shared sequences; note that the y-axis is a different scale in these four plots, though the x-axis is consistent.
We investigated whether mouse sex and age affected the extent to which mice shared amino acid–defined TCR sequences. Mindful of the differences in sharing among mice from the different wild sites, we focussed this analysis on only the mice from the Nottingham site, and used a GLM that analysed the number of shared sequences by (i) same sex vs. different sex pairs, (ii) sum of age of mouse pairs, and (iii) difference in age of mouse pairs. We used these sum and difference in age metrics to see if individuals’ TCR repertoires converged or diverged as mice age. This showed that overall there was no difference in the number of shared TCR sequences between same sex and different sex pairs (t = 0.398, P = 0.69) and that the sum of mouse pair age was not significant (t = 0.180, P = 0.47), nor for the difference in mouse pair age (t = −0.541, P = 0.122) (Fig. 3; Supplementary Material S10). However, post hoc analysis of the four different receptor types showed that for CD8^+^ alpha and beta receptors there were consistent negative relationships of the number of shared sequences and the sum or difference of age (Supplementary Material S11). This was also observed for CD4+ beta chains, but not alpha.
The number of shared amino acid–defined TCR sequences for CD8+ alpha between all pairwise combinations of wild mice from the Nottingham site against (a) the difference in pairwise age and (b) the sum of the pairwise age, where age is in days. The results for CD4+ alpha and beta and CD8+ beta chains are shown in Supplementary Material S10.
Discussion
The aim of this work was to describe—for the first time, as far as we are aware—the TCR repertoire of wild mice and to investigate if, and how, individuals’ age, sex, and the site from which they were obtained affected their TCR repertoire. This work is directly relevant to the fields of eco-immunology and mouse immunology more generally. We successfully sampled wild mice and obtained TCR sequences from CD4^+^ and CD8^+^ cells. Despite wild mice being smaller than laboratory mice and having absolutely and proportionally smaller spleens with fewer splenocytes [4], we successfully obtained sufficient CD4^+^ and CD8^+^ cells from 65 wild mice for TCR sequence analysis.
Wild mice have large TCR repertoires and their richness was not affected by individuals’ sex, age, or the site from which the mice were obtained. Richness (which is the normalized number of unique TCRs in a repertoire) measures the breadth of the TCR repertoire. We hypothesize, therefore, that both male and female mice maintain the capacity to respond to a wide range of antigens in both CD4^+^ and CD8^+^ T-cell compartments throughout their natural lifespan in the wild. This contrasts with some studies in humans that report declining richness with age [8]. These differences between wild mice and people may reflect some immunological differences between wild mouse and human populations. But in addition to this (i) our sampling of wild mice will have under-sampled very young mice (which typically will not leave their nests), so that we may not have been able to account for TCR richness in very young individuals, and (ii) the maximum lifespan of wild mice is not equivalent to that of elderly people included in these studies and/or the relative rarity of old mice limits our statistical power.
In contrast to TCR repertoire richness, measures of diversity (such as Shannon and Simpson’s indices) assess both the number of different TCRs and their relative abundance, such that when there are more abundant clones then the diversity is lower. The two diversity indices give slightly different results, but concur in showing an effect of sex on TCR diversity, and generally agree in showing an effect of age; Shannon’s index also shows an effect of an interaction between age and sex. In the context of constant richness, decreased diversity must reflect an increase in the abundance of some clones, which we hypothesize may arise from chronic exposure to a limited set of infections prevalent in wild mouse populations. An influence of sex on diversity has been reported previously in humans, with greater diversity in females [44]. The mechanism behind these effects remains unclear, but could reflect hormonal differences leading to differences in thymic selection [44]. Decreased diversity with age has also been described repeatedly in humans [15]. In wild mice the effects of sex and age may reflect differences among mice in their exposure to infection, with higher diversity a consequence of less exposure to infection. Thus, young mice will not yet have had time to experience many infections unlike older mice. Male house mice may have more limited social interactions because they typically disperse from the site of their birth [20] leading to comparatively less intense or less chronic exposure to infection in their environment. In contrast, females breed communally [19], which may expose them to greater levels of infection and antigenic challenge. Antigenic sources beyond infection—including self-antigens and tumours—will also result in T-cell responses, and these may contribute to the age and sex effects on TCR diversity. However, the antigenic assault from infection is particularly strong in wild animals, especially compared to laboratory animals, and we suspect likely to be the major antigen source across wild animal populations. Taken together, our results suggest that wild mice maintain a broad TCR repertoire that allows them to react to a breadth of antigens but, particularly in older, female mice, this is evidence of strong, life-long immune responses to a limited set of persistent infections in their environment.
We found that individual mice shared a considerable number of TCR sequences, specifically ∼5% of alpha chains and ∼2% of beta chains. The differences in the amount of TCR sharing between alpha and beta chains is, presumably, because beta chains are comparatively more diverse, making the chance of them being shared less likely compared with alpha chains. We found a pattern of greater sharing between mice from within the same group (from Nottingham, from the Wirral, or laboratory mice) than between groups, which is suggestive of diverse antigenic experience of mice—leading to different TCR sequences—in the different environments of the groups of mice. This is consistent with previous work that found immunological differences between wild mice from different sample sites [4].
We also found that wild mouse age affects the number CD8^+^ TCR sequences that are shared among mice. We suggest that this occurs because as mice age they become more idiosyncratic in their amino acid–defined TCR sequences, so that older mice share fewer sequences with younger mice (the effect of the difference in mouse age) and that older mice share fewer sequences with other, older mice (the effect of the sum of mouse age). Idiosyncratic responses of laboratory mice to protein antigens and viral infection have been reported previously [14, 45]. Study of wild mouse (M. musculus and Apodemus sylvaticus) gut microbiomes show that they differ significantly among individuals [46, 47], and this may result in idiosyncratic antigenic experience in the gut, which may contribute to the individuality of wild mouse TCRs that we have observed.
Our study has a number of limitations. One, we obtained our samples from live-trapped mice, and this method does not effectively sample very young animals, so that these are under-represented in our study. Two, we caught wild mice at only two sample sites, and most of the caught mice came from one site. This did not give us much statistical power to investigate how TCR repertoires may vary among individuals from different sample sites. This could be addressed by further work in the future, which could be extended to analyse both spatial and temporal aspects of TCR repertoire diversity. Three, we analysed only a small number of control, laboratory mice. Surprisingly, given the central role of laboratory mouse models in the development of modern immunology, there appear to be a dearth of publicly available data on the TCR repertoires of the common strains of laboratory mice. We have therefore been unable to make a comprehensive comparison of TCR repertoires between wild and laboratory mice. Given the already established differences in immune state between wild and laboratory mice, a future, in-depth comparison between the TCR repertoires of wild and laboratory mice would be of interest.
Immunological analyses of non-standard model species is often hampered by the paucity of immunological reagents that are available. However, because the TCR repertoire can be investigated using sequence-based analyses this approach can be widely applied to a whole range of species. In so doing, this may allow a broad, comparative study of wild animals’ TCR repertoires.
In summary, here we describe the TCR repertoire of wild mice and investigated how this is affected by individuals age, sex, and site of sampling. Our analyses of the wild mouse TCR repertoire find some patterns that are similar to human TCR repertories, though we find differences between wild male and female mice that may reflect their different lives and exposure to infection.
Supplementary Material
kyag002_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Viney M, Riley EM. The immunology of wild rodents: current status and future prospects. Front Immunol 2017, 8, 1481.29184549 10.3389/fimmu.2017.01481 PMC 5694458 · doi ↗ · pubmed ↗
- 2Beura LK, Hamilton SE, Bi K, Schenkel JM, Odumade OA, Casey KA, et al Normalizing the environment recapitulates adult human immune traits in laboratory mice. Nature 2016, 532, 512–6.27096360 10.1038/nature 17655 PMC 4871315 · doi ↗ · pubmed ↗
- 3Abolins S, King EC, Lazarou L, Weldon L, Hughes L, Drescher P, et al The comparative immunology of wild and laboratory mice, Mus musculus domesticus. Nat Commun 2017, 8, 14811.28466840 10.1038/ncomms 14811 PMC 5418598 · doi ↗ · pubmed ↗
- 4Abolins S, Lazarou L, Weldon L, Hughes L, King EC, Drescher P, et al The ecology of immune state in a wild mammal, Mus musculus domesticus. P Lo S Biol 2018, 16, e 2003538.29652925 10.1371/journal.pbio.2003538 PMC 5919074 · doi ↗ · pubmed ↗
- 5Leung JM, Budischak SA, Chung The H, Hansen C, Bowcutt R, Neill R, et al Rapid environmental effects on gut nematode susceptibility in rewilded mice. P Lo S Biol 2018, 16, e 2004108.29518091 10.1371/journal.pbio.2004108 PMC 5843147 · doi ↗ · pubmed ↗
- 6Rosshart SP, Vassallo BG, Angeletti D, Hutchinson DS, Morgan AP, Takeda K, et al Wild mouse gut microbiota promotes host fitness and improves disease resistance. Cell 2017, 171, 1015–1028.e 13.29056339 10.1016/j.cell.2017.09.016PMC 6887100 · doi ↗ · pubmed ↗
- 7Sender R, Weiss Y, Navon Y, Milo I, Azulay N, Keren L, et al The total mass, number, and distribution of immune cells in the human body. Proc Natl Acad Sci U S A 2023, 120, e 2308511120.37871201 10.1073/pnas.2308511120 PMC 10623016 · doi ↗ · pubmed ↗
- 8Weng NP . Numbers and odds: TCR repertoire size and its age changes impacting on T cell functions. Semin Immunol 2023, 69, 101810.37515916 10.1016/j.smim.2023.101810 PMC 10530048 · doi ↗ · pubmed ↗