Reply to Roske and Yeeles: Mismatch correction by a replicative polymerase constrained on DNA by a ring
Feng Wang, Qing He, Michael E. O’Donnell, Huilin Li

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
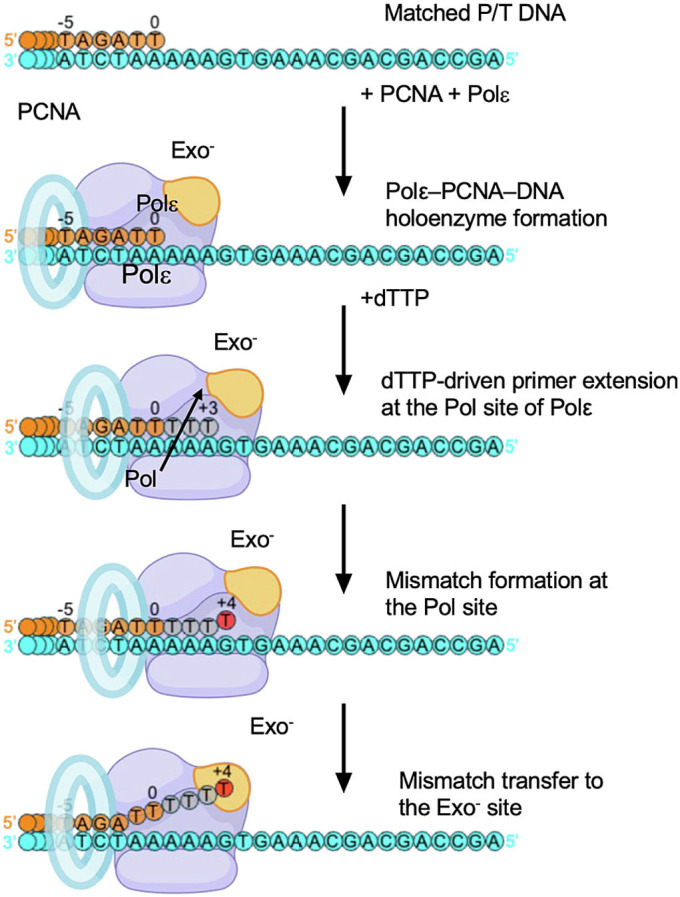 Figure 1
Figure 1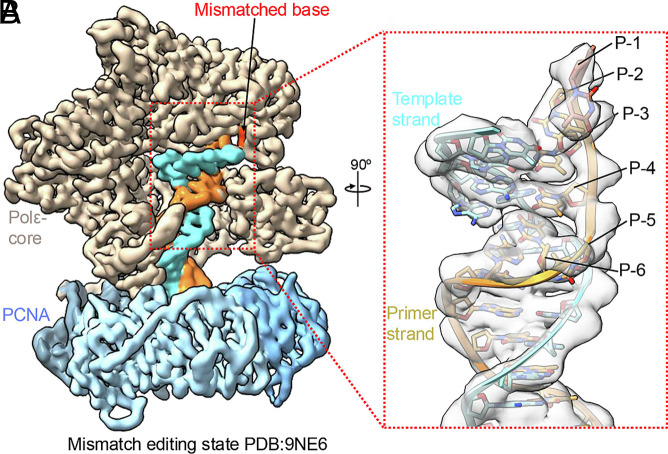 Figure 2
Figure 2- —HHS | NIH | National Institute of General Medical Sciences (NIGMS)100000057
- —HHS | NIH | National Institute of General Medical Sciences (NIGMS)100000057
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDNA Repair Mechanisms · Genetic factors in colorectal cancer · Bacterial Genetics and Biotechnology
Roske and Yeeles comment on our study of 3′ mismatch correction by Polε held on DNA by proliferating nuclear antigen (PCNA) (Fig. 1, figure 2 in ref. 1). Our goal was to observe the constraints that PCNA places on Polε during mismatch correction. Mismatch excision studies generally add a polymerase to a preformed mismatch; the typical result is 3 bp of unwinding for 3′ DNA to enter the exo site. Indeed, addition of Polε to a preformed mismatch (with PCNA added later) yields the 3 bp result, but PCNA is not fully engaged with Polε (SI Appendix, figure S5B in ref. 2). These reaction schemes do not require 3′ DNA to first enter the pol site. Therefore, we developed an in situ method that starts with a flush 3′ DNA in the pol site of Polε fully engaged with PCNA; 0.5 mM deoxythymidine-5’-triphosphate (dTTP) is then added to facilitate incorporation of three dTTP’s and a 3′ dT–dC mismatch (+4 dT total), followed by switching of the mismatched 3′ dT to the exo site (Fig. 1) (3). This scheme results in 6 bp unwinding of the 3′ terminus to enter the exo site of a fully engaged Polε-PCNA due to constraints on DNA movements by PCNA (Fig. 2 A and B). In their recapitulation of our work, Roske and Yeeles show (Fig. 2B, lanes 5 to 7 in ref. 1) that the single 3′ mismatch (+4 band) is the major species at 3 min, validating our use of 3-min incubation before grid preparation. A minor +9 band that would require 3 mismatches (figure 2 B and C in ref. 1) is rapidly formed in 10 s, even before the single +4 mismatch. We suggest the rapid formation may be due to undetectable dNTP contaminants largely consumed within 10 s. Some readthrough is eventually expected for exo-deficient Pols (4, 5). We did not use the alternative substrates shown in figure 2D of ref. 1. Importantly, it cannot be known which proteins produce different gel bands, and thus we focused on cryo-electron microscopy (cryo-EM) particles that contained both PCNA and Polε. All three mismatch-proofreading intermediates show density supporting incorporation of the +4 nucleotide, followed by 6 bp unwinding with PCNA fully engaged (Fig. 2).
The other difference between the studies (figure 1 in ref. 1) centers on a Polε–PCNA blunt-end structure, which we do not observe under our conditions. There are numerous differences in the conditions used in the two studies (e.g., T–G vs. T–C mismatch, order of protein addition, buffers, salts, ddATP) (2, 3). While blunt-end binding is not a physiological pathway to proofreading, we agree with their assertion that biotinylation of 5′ ends should specify polymerase binding to a 3′ end.
Reactions in cells are highly dynamic, and proofreading may follow multiple paths. Our study addresses only constraints imposed by PCNA on the exo process. We do not exclude that Polε may associate with a mismatch before PCNA, as in ref. 2. In conclusion, our Polε–PCNA was preassembled on DNA, ensuring mismatch formation in the pol site followed by transition to the exo site. The structural and biochemical data support our original conclusions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1J. J. Roske, J. T. Yeeles, Cryo-EM maps of human DNA polymerase ε should be re-evaluated in light of its unexpected behaviour in vitro. Proc. Natl. Acad. Sci. U.S.A. 123, e 2533320123 (2026).41790928 10.1073/pnas.2533320123 · doi ↗ · pubmed ↗
- 2J. J. Roske, J. T. Yeeles, Structural basis for processive daughter-strand synthesis and proofreading by the human leading-strand DNA polymerase pol ε. Nat. Struct. Mol. Biol. 31, 1921–1931 (2024).39112807 10.1038/s 41594-024-01370-y PMC 11638069 · doi ↗ · pubmed ↗
- 3F. Wang, Q. He, M. E. O’Donnell, H. Li, The proofreading mechanism of the human leading-strand DNA polymerase ε holoenzyme. Proc. Natl. Acad. Sci. U.S.A. 122, e 2507232122 (2025).40440070 10.1073/pnas.2507232122 PMC 12146725 · doi ↗ · pubmed ↗
- 4V. Khare, K. A. Eckert, The proofreading 3’→ 5’ exonuclease activity of DNA polymerases: A kinetic barrier to translesion DNA synthesis. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 510, 45–54 (2002).10.1016/s 0027-5107(02)00251-812459442 · doi ↗ · pubmed ↗
- 5M. W. Frey, N. G. Nossal, T. L. Capson, S. J. Benkovic, Construction and characterization of a bacteriophage T 4 DNA polymerase deficient in 3’–> 5’exonuclease activity. Proc. Natl. Acad. Sci. U.S.A. 90, 2579–2583 (1993).8464864 10.1073/pnas.90.7.2579 PMC 46138 · doi ↗ · pubmed ↗