Revision of the genus Splanchospora (Pleosporales, Neohendersoniaceae)

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
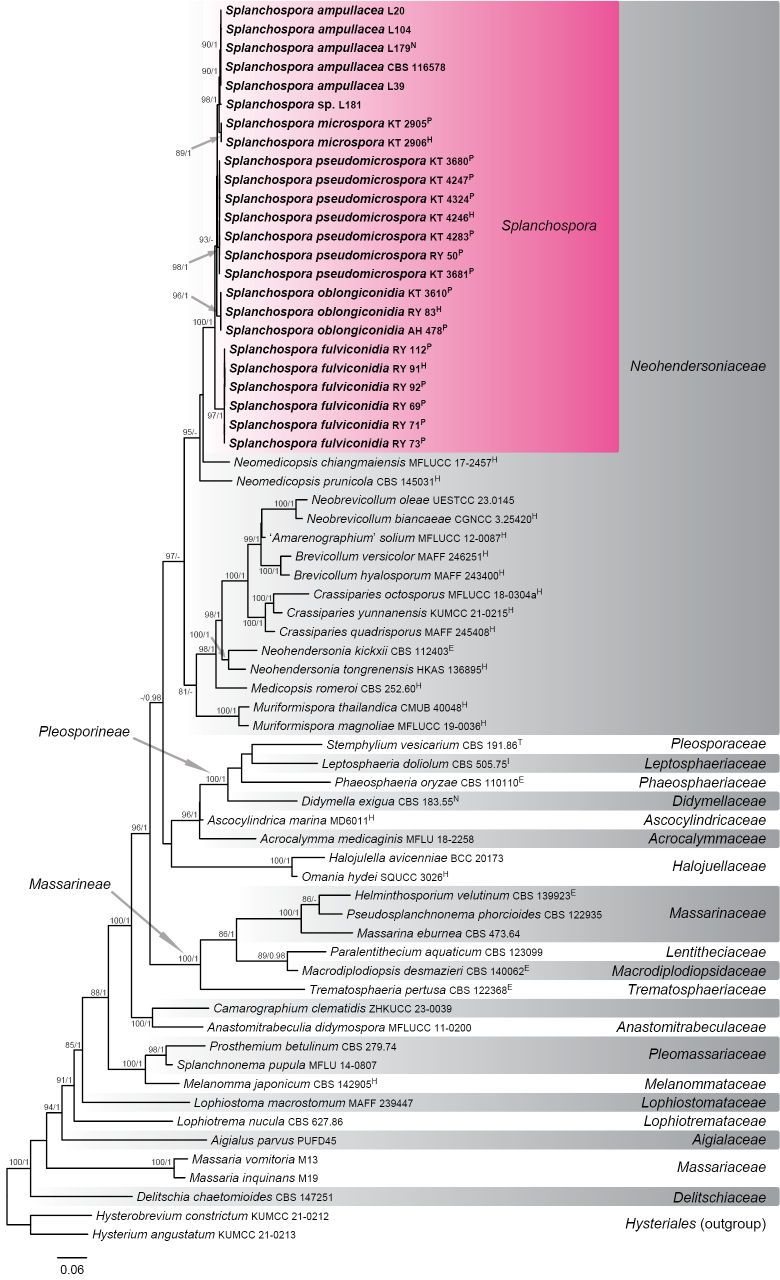 Figure 1
Figure 1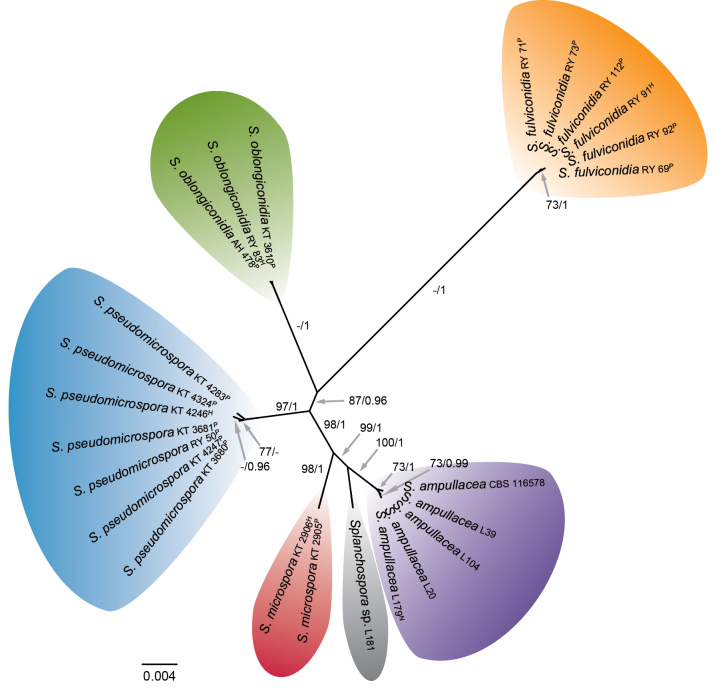 Figure 2
Figure 2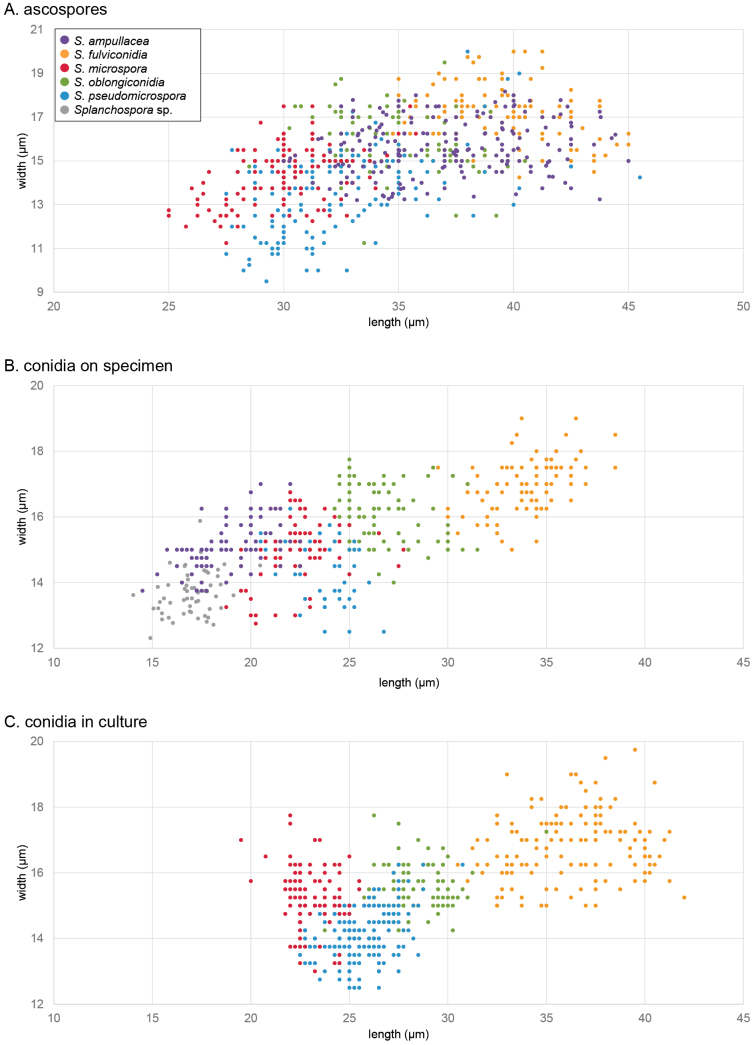 Figure 3
Figure 3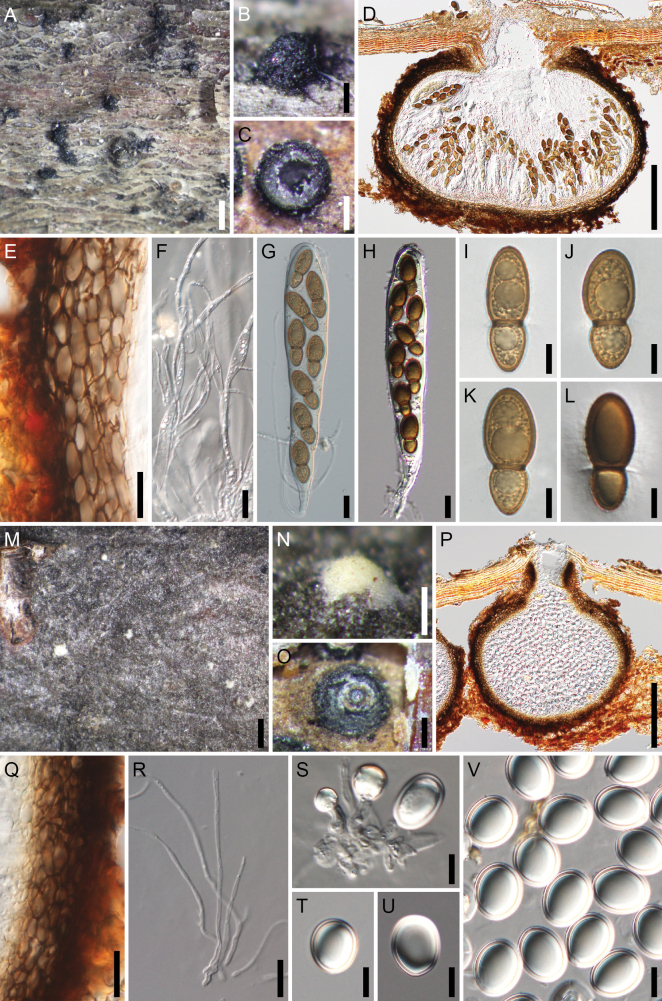 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6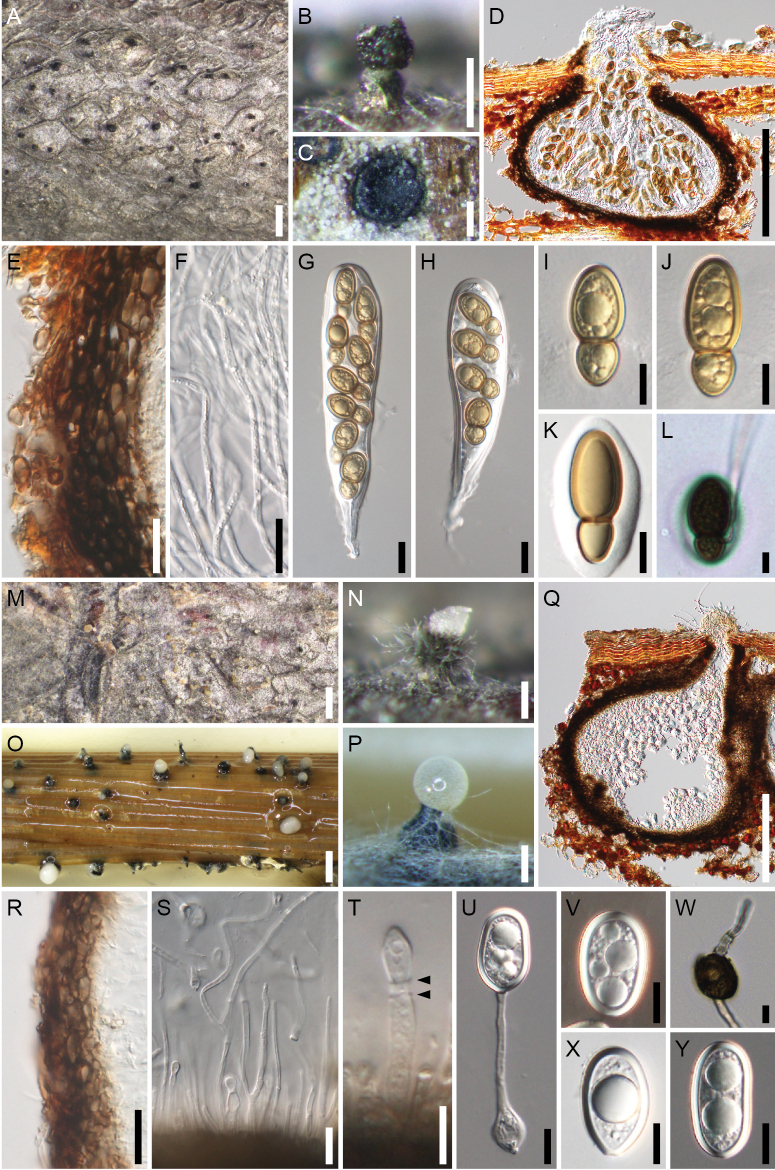 Figure 7
Figure 7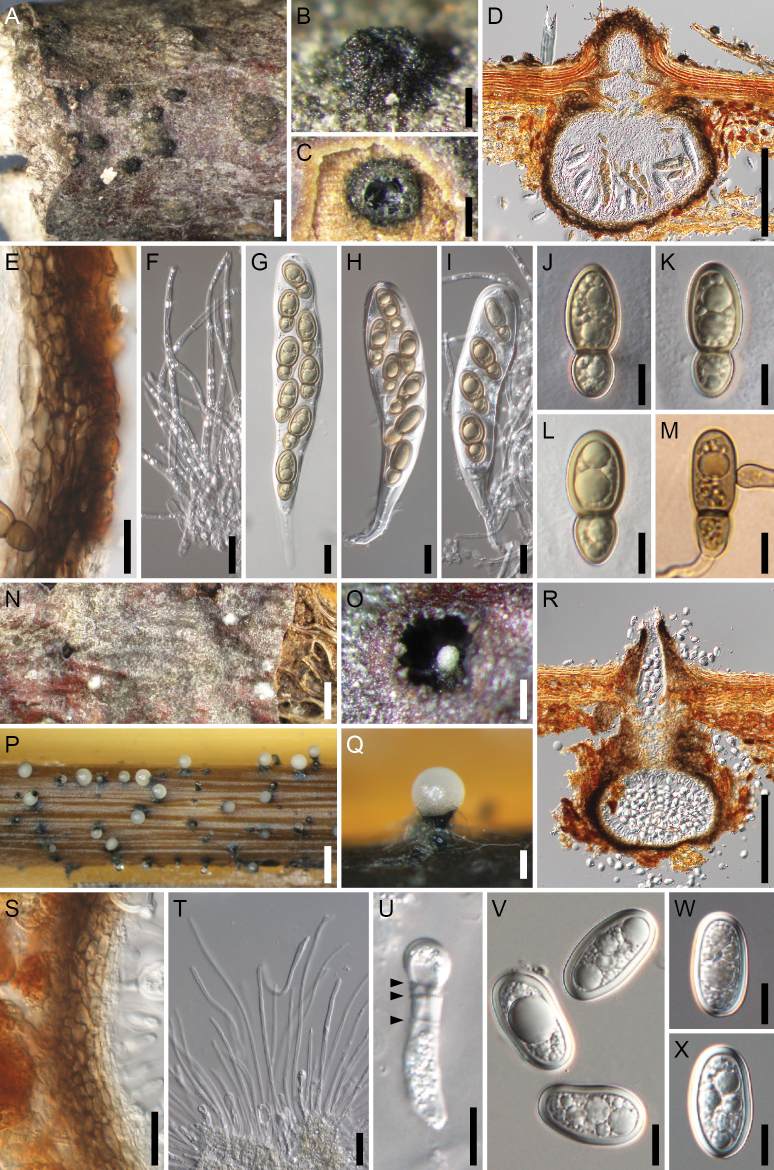 Figure 8
Figure 8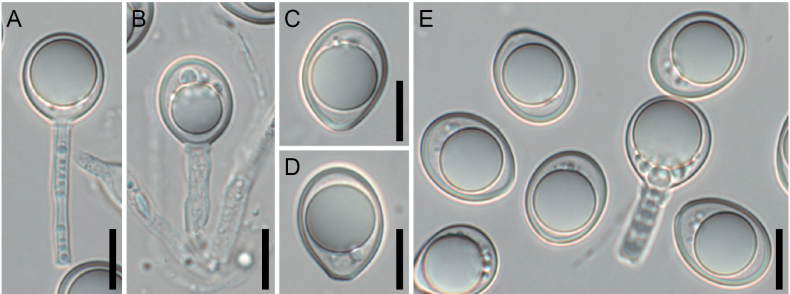 Figure 9
Figure 9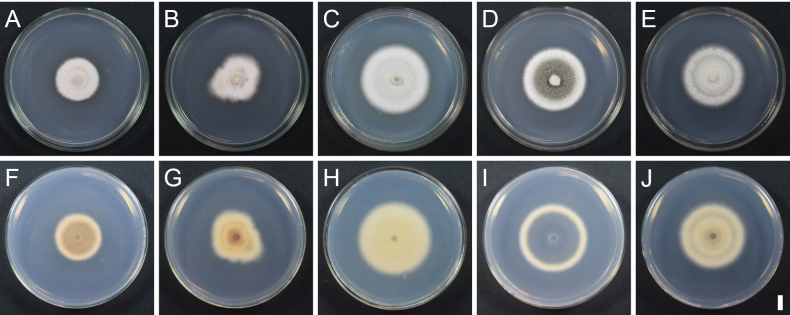 Figure 10
Figure 10| Taxon | Culture/voucher no. | Type† | GenBank accession no. | References | ||||
|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
| ||||
|
| MFLU 18-2258 |
|
|
|
|
|
| |
|
| PUFD45‡ |
|
|
|
|
|
| |
| ‘ | MFLUCC 12-0087 | H |
| – |
| – | – |
|
|
| MFLUCC 11-0200 |
|
|
|
|
|
| |
|
| MD6011‡ | H |
| – |
| – | – |
|
|
| H |
|
|
|
|
|
| |
|
| H |
|
|
|
|
|
| |
|
| ZHKUCC 23-0039 |
|
|
| – |
|
| |
|
| MFLUCC 18-0304a | H | – |
|
| – |
|
|
|
| H |
|
|
|
|
| ||
|
| KUMCC 21-0215 | H |
|
|
|
|
|
|
|
|
|
|
|
|
|
| ||
|
| N |
|
|
|
| genome¶ | ||
|
| BCC 20173 |
| – |
|
|
|
| |
|
| E |
|
|
|
|
|
| |
|
| KUMCC 21-0213 |
|
|
|
|
|
| |
|
| KUMCC 21-0212 |
|
|
|
|
|
| |
|
| I§ |
|
|
|
|
| ||
|
|
|
|
|
|
| |||
|
|
|
|
|
|
| |||
|
| E | – |
|
| – | – |
| |
|
| M19 |
|
|
|
|
|
| |
|
| M13 |
|
|
|
|
|
| |
|
|
|
|
|
|
| |||
|
| H |
|
|
|
|
| ||
|
| H |
|
|
|
|
|
| |
|
| MFLUCC 19-0036 | H |
|
|
|
|
|
|
|
| CMUB 40048 | H |
|
|
|
|
|
|
|
| CGNCC 3.25420 | H |
|
|
|
|
|
|
|
| UESTCC 23.0145 |
|
|
|
|
|
| |
|
| E | – |
|
| – | – |
| |
|
| HKAS 136895 | H | – |
|
| – | – |
|
|
| MFLUCC 17-2457 | H |
|
|
| – | – |
|
|
| H | – |
|
|
| – |
| |
|
| SQUCC 3026 | H |
|
|
| – |
|
|
|
| H |
|
|
|
|
| ||
|
| E |
|
|
|
|
| ||
|
|
|
|
|
|
| |||
|
|
|
|
|
|
|
| ||
|
| MFLU 14-0807 | – |
|
| – | – |
| |
|
|
|
|
|
|
| This study | ||
| L20 |
|
|
|
|
| This study | ||
| L39 | – |
|
|
|
| This study | ||
| L104 | – |
|
|
|
| This study | ||
| L179 | N | – |
|
|
|
| This study | |
|
| RY 69 | P |
|
|
|
|
| This study |
| RY 71 | P |
|
|
|
|
| This study | |
| RY 73 | P |
|
|
|
|
| This study | |
| RY 91 | H |
|
|
|
|
| This study | |
| RY 92 | P |
|
|
|
|
| This study | |
| RY 112 | P |
|
|
|
|
| This study | |
| RY 152 | P | – |
| – | – | – | This study | |
| RY 153 | P | – |
| – | – | – | This study | |
| RY 161 | P | – |
| – | – | – | This study | |
|
| KT 2905 | P |
|
|
|
|
| This study |
| KT 2906 | H |
|
|
|
|
| This study | |
|
| AH 478 | P |
|
|
|
|
| This study |
| KT 3610 | P |
|
|
|
|
| This study | |
| RY 83 | H |
|
|
|
|
| This study | |
|
| KT 3680 | P |
|
|
|
|
| This study |
| KT 3681 | P |
|
|
|
|
| This study | |
| KT 4246 | H |
|
|
|
|
| This study | |
| KT 4247 | P |
|
|
|
|
| This study | |
| KT 4283 | P |
|
|
|
|
| This study | |
| KT 4324 | P |
|
|
|
|
| This study | |
| RY 50 | P |
|
|
|
|
| This study | |
| RY 154 | P | – |
| – | – | – | This study | |
| RY 158 | P | – |
| – | – | – | This study | |
| L181 | – |
|
|
|
| This study | ||
|
| T| |
|
|
|
|
| ||
|
| E |
|
|
|
|
| ||
| ITS1 (1–143) | ITS2 (303–452) | |||||||||||||||||||||||||
| site number | 49 | 53 | 79 | 82 | 132 | 135 | 136 | 141 | 340 | 341 | 342 | 343 | 344 | 362 | 363 | 379 | 387 | 404 | 414 | 419 | 422 | 444 | ||||
| C | C | C | A | A | A | C | G | C | G | C | C | T | C | T | C | C | C | T | A | G | T | |||||
| G |
| C | A | C/- |
| T |
|
|
| -† | -† | -† | T | T | C | C | A | A | G |
|
| |||||
| G | C | C | A | - | A | T | G | C | G | C |
| T | T | C | T | C | C | T | A | G | T | |||||
| G | C | C |
| A | A | C | G | C | G | C | C | T | C | C | C | C | C | T | A | A | T | |||||
| G | C | C | A | A | A | C | G | C | G | C | C | T | T/C | C/T | C | C | C | T | A | G | T | |||||
| C | C |
| A | A | A | C | G | C | G | C | C | T | C | T | C | T | C | T | A | G | T | |||||
| site number | 490–807 | 884 | 885 | 887 | 935 | 969 | 977 | 978 | 979 | 980 | 988 | 56 | 58 | 60 | 99 | 371 | 373 | 410 | 526 | 624 | 655 | 662 | 1026 | 1248 | 1267 | 1285 |
|
| 318 sites‡ | T‡ | G‡ | G‡ | T‡ | T‡ | G‡ | -‡ | G‡ | A‡ | C‡ | A | T | T | T | G | G | C | T | G | C | G | C§ | C§ | G§ | C§ |
|
| - |
|
|
| -† |
|
|
|
|
|
|
|
|
| C |
|
|
|
|
|
| G |
|
|
|
|
|
| - | T | G | G | T | T | G | - | G | A | C | A | T | T | C | G | G | C | T | G | C |
| C | C | G | C |
|
| - | T | G | G | T | T | G | - | G | A | C | A | T | T | T | G | G | C | T | G | C | G | C | C | G | C |
|
| 318 sites | T | G | G | T | T | G | - | G | A | C | A | T | T | T | G | G | C | T | G | C | G | C | C | G | C |
| n/a¶ | n/a¶ | n/a¶ | n/a¶ | n/a¶ | n/a¶ | n/a¶ | n/a¶ | n/a¶ | n/a¶ | n/a¶ | A | T | T | T | G | G | C | T | G | C | G | n/a¶ | n/a¶ | n/a¶ | n/a¶ | |
| site number | 18 | 45 | 81 | 84 | 87 | 90 | 120 | 127 | 132 | 139 | 147 | 150 | 153 | 189 | 192 | 198 | 219 | 291 | 294 | 297 | 303 | |||||
|
| T | C | T | C | C | C | T | C | A | T | G | C | T | G | C | A | C | G |
| G | C | |||||
|
|
|
| C | C |
| C |
|
| G |
| G | C | T |
| C | A |
| G | A | G |
| |||||
|
| T | C | T | C | C | C | T | C | A | T | G | C | C | G | C | A | C | G | A | G | C | |||||
|
| T | C | C |
| C |
| T | C | A | T |
| C | C | G |
| A | C |
| A | G | C | |||||
|
| T | C | C | C | C | C |
| C | G | T | G |
| C | G | C |
| C | G | A | G | C | |||||
| T | C | T | C | C | C | T | C | A | T | G | C | C | G | C | A | C | G | A |
| C | ||||||
| site number | 306 | 309 | 312 | 333 | 336 | 360 | 366 | 372 | 378 | 396 | 399 | 456 | 465 | 480 | 483 | 492 | 501 | 504 | 510 | 522 | 537 | |||||
|
| C | T | C | T | G | C | C | C | G | T | C |
| T | T | A | C | C | C | T | T |
| |||||
|
| C |
|
| C |
| T | T |
| A |
|
| T | T | T | A | C |
| C |
| T | A | |||||
|
| C | T | C | T | G | C | C | C | G | T | C | T | T | T | A | C | C | C | T | T | A | |||||
|
|
| T | C | C | G | T | T | C | A | T | C |
| C | T |
|
| C |
| T |
| A | |||||
|
| C | T | C | C | G | C | C | C | G | T | C | T | C |
| A | C | C | C | T | T | A | |||||
| C | T | C | T | G | C | C | C | G | T | C | T | T | T | A | C | C | C | T | T | A | ||||||
| site number | 570 | 579 | 609 | 618 | 624 | 636 | 642 | 657 | 666 | 667 | 675 | 684 | 687 | 708 | 711 | 714 | 726 | 729 |
| 741 | 765 | |||||
|
| C | G | T | T | A | T | G | C | C | C | T | C | T | T | C | C | A | G | C | G | C | |||||
|
|
| G | A | T | A | C | A |
| T | T | T |
| T |
| C | C | G | A |
| G | C | |||||
|
| C | G | T | T | A | C | G | C | C | C | T | C | T | T | C | C | A | G | C | G | C | |||||
|
| C |
| A |
| A | C | A | C | T | C | T | C | C | T | C | T | G | A | C | G |
| |||||
|
| C | G | A | T |
| C | A | C | T | T | T | C | C | T |
| T | G | A | C |
| C | |||||
| C | G | T | T | A | T | G | C | C | C |
| C | T | T | C | C | A | G | C | G |
| ||||||
| site number | 768 | 780 | 783 | 789 | 795 | 831 | 861 | 873 | 918 |
| 930 | 936 | 942 | 963 |
| 975 | 1020 | 1026 | 1041 | 1047 | 1050 | 1056 | ||||
|
| G | C | T | A | A | T | G | T | G | A | T | T | C | T | C | T | A | T | T | C | C | C | ||||
|
|
| C | T | A | A | T | A |
| G | A | C | T | C | T | C | T | G |
|
|
| T | T | ||||
|
| G | C |
| A | A | T | G | T | G | G | C | T | C | T | C | T | A | T | T | C | C | T | ||||
|
| G |
| T | G | A | T | G | T |
| G | C |
|
|
| C |
| G | T | T | C | T | T | ||||
|
| G | C | T | G |
|
| A | T | G | G | C | T | C | T |
| T | G | T | T | C | T | T | ||||
| G | C | T | A | A | T | G | T | G | G | T | T | C | T | C | T | A | T | T | C | C | C | |||||
| site number | 5 | 17 | 28 | 33 | 45 | 46 | 114 | 117 | 128 | 157 | 163 | 164 | 168 | 169 | 173 | 174 | 182 | |||||||||
|
| T | T | C | A | - | - | T | C | G | T | A | T | A | T | G | T | A | |||||||||
|
| T | A |
|
|
|
| G | C |
| T |
|
| A | T |
|
|
| |||||||||
|
|
| T | C | A | - | - | G | C | G |
| A | T | A | T | G | T | A | |||||||||
|
| T | A | C | A | - | - | G | C | G | T | A | T | C |
| G | T | A | |||||||||
|
| T | T | C | A | - | - | G/T | C | G | T | A | T | A | T | G | T | A | |||||||||
| T | T | C | A | - | - | G |
| G | T | A | T | C | T | G | T | A | ||||||||||
| exon 1 (271–409) | ||||||||||||||||||||||||||
| site number | 183 | 186 | 201–221 | 222 | 227 | 228 | 229 | 231 | 242 | 246 | 248 | 249 | 250 | 258 | 262 | 263 | 264 | 396 | ||||||||
|
| T/C | A | - | A | T | T | G | C |
| C | C | A | A | A |
| C | T | T | ||||||||
|
| T | A |
| C |
|
|
|
| G |
|
| A |
| A | T | C | T |
| ||||||||
|
| T |
| - | A | T | T | G | C | G | C | C | A | A | A | T | C | T | T | ||||||||
|
| T | A | - | C | T | T | G | C | G | C | C |
| A | A | T | C | C | T | ||||||||
|
| C | A | - | A | T | T | G | C | G | C | C | A | A | A | T | C | C | T | ||||||||
| T | A | - | A | T | T | G | C | G | C | C | A | A |
| T |
| C | T | |||||||||
| exon 2 (460–1,253) | ||||||||||||||||||||||||||
| site number | 412 | 417 | 418 | 422 | 423 | 424 | 425 | 426 | 427 | 431 | 437 | 438 | 439 | 446 | 467 | 518 | 635 | 638 | 641 | 659 | 677 | 704 | 752 | |||
|
| A | A | T | T | A | A | T | G | C | G/C | G | G | T | T | C | C | T | C | C | C | T | C | T | |||
|
| A |
|
| T | C | A | - | G | T |
| G |
| C | T | C |
| C/T | C | C |
|
| C |
| |||
|
| A | A | T | -† | -† | -† | - |
| T | G | G | G | C | T | C | C | T | C |
| C | T |
| T | |||
|
|
| A | T | T | C | A | T | G | C | G | G | G | C |
|
| C | C |
| C | C | T | C | T | |||
|
| A | A | T | T |
| A | T | G | C | G | G | G | T | T | C | C | T | C | C | C | T | C | T | |||
| A | A | T | T | A | A | T | G | C | G |
| G | C | T | C | C | T | C | C | C | T | C | T | ||||
| site number | 758 | 767 | 776 | 788 |
|
| 872 | 893 | 923 | 986 | 1058 | 1070 | 1073 |
| 1097 | 1103 | 1106 | 1160 | 1178 | 1196 | 1211 | 1226 | 1235 | 1253 | ||
|
| C | T | T | C | A | C | T | G | G | T | G | G | G | C | C | C | T | C | G | C | T |
| G | T | ||
|
| C | T |
| T | A | C | C | C |
|
| G |
| G |
| C | C | T | T |
|
| T | C | G |
| ||
|
|
| C | T | T | A | C | T | G | G | T |
| G | G | C |
| C | C | C | G | C | T | C |
| T | ||
|
| C | C | T | C |
| C | C | C | G | T | G | G | G | C | C | C | C | T | G | C | T | C | G | T | ||
|
| C | T | T | C | A |
| T | G | G | T | G | G | G | C | C |
| C | T | G | C |
| C | G | T | ||
| C | T | T | C |
| C | T | G | G | T | G | G |
| C | C | C | C | C | G | C | T | C | G | T | |||
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Fungal Plant Pathogen Control · Infectious Diseases and Mycology
Introduction
The monotypic genus Splanchospora Lar.N. Vassiljeva was established in 1998 to rearrange Splanchnonema ampullaceum (Basionym: Sphaeria ampullacea Pers.), inhabiting twigs of Tilia spp. L. native to Europe. The genus is characterised solely by 2-celled ascospores with a strongly submedian primary septum (Vasilyeva 1998; Tanaka et al. 2005). Although a detailed description of the sexual morph of the type species, Splanchospora ampullacea, was provided by Barr (1982) and Cannon and Minter (2014), additional characterisation of the genus on the basis of the asexual morph has not yet been performed. Tonolo (1956) carried out a culture study of Massariella curreyi, a synonym of S. ampullacea, and observed a phoma-type pycnidial asexual morph. However, Barr (1982) pointed out that this may be a spermatial state. Due to the ambiguous morphological circumscription of Splanchospora, Cannon and Minter (2014) suggested that this species should be retained in the genus Splanchnonema rather than Splanchospora. According to Vasilyeva (1998), Splanchospora belongs to Massariaceae; however, this classification could so far not be verified by phylogenetic analysis owing to the absence of S. ampullacea sequence data. Therefore, to date, the genus remains as Ascomycota genus incertae sedis (Hyde et al. 2024).
Peck (1894) described Massariella curreyi var. americana from Tilia americana L. in the USA as being different from the typical M. curreyi found in Europe. However, Barr (1982) subsequently re-examined the holotype of this North American variety and found that it did not differ sufficiently, on the basis of the sexual morph alone, from its European counterpart and synonymised it with M. curreyi. Splanchospora ampullacea is considered a semi-cosmopolitan species which colonises various linden species and has even been reported in the Russian Far East (Vasilyeva 1998).
In our ongoing taxonomic study of dothideomycetous fungi (e.g., Yoshioka et al. 2025), several fresh specimens of sexual morphs of Splanchospora spp. were collected from the twigs of lindens. In most of these specimens, coelomycetous asexual morphs were consistently observed to be associated with the sexual morphs. Under culture conditions, single ascospore isolates formed asexual morphs matching those observed on the original specimens. In this study, we aimed to revise the generic concept of Splanchospora by incorporating information from the asexual morphs, to clarify the phylogenetic position of the genus, and to identify the species found in Japan.
Materials and methods
Specimens and isolates
Twenty-eight fresh fungal specimens were collected from dead twigs of Tilia spp. (Malvaceae), Carpinus cordata Blume, and Ostrya japonica Sarg. (both Betulaceae) in Japan and Europe from 2006 to 2024. Twenty-six of these specimens were collected from linden trees, most planted in botanical gardens or parks. All specimens were deposited at the Fungaria of Hirosaki University (HHUF) and/or the Department of Botany and Biodiversity Research, University of Vienna (WU). Single-spore isolates were obtained from the fresh specimens, and representative isolates were deposited at the NARO Genebank (MAFF), Tsukuba, Japan; or the Westerdijk Fungal Biodiversity Institute (the CBS Culture Collection), Utrecht, The Netherlands. Additionally, a strain of S. ampullacea (CBS 116578) was purchased from the Westerdijk Fungal Biodiversity Institute for comparative analysis. For detailed information on these specimens and isolates, see Suppl. material 1: table SS1.
Colony characteristics were recorded from growth on potato dextrose agar (PDA) at 20 °C in the dark for 2 weeks. Colony colours were determined by referring to Rayner (1970). Several pieces of mycelial agar were placed on water agar containing sterilised banana leaves, Japanese beech twigs, and rice straws to observe sporulation in vitro. After the mycelia had colonised the substrates at 20 °C for 2 weeks, the plates were incubated at 20 °C under blacklight blue illumination for 2–4 weeks to induce sporulation.
Morphological observations and comparison of spore sizes
Macroscopic morphological observations were made using an Olympus SZX10 stereomicroscope (Olympus, Tokyo, Japan). Slide preparations were typically mounted in distilled water. Shear’s mounting medium was used to mount sections of ascomata/conidiomata. Morphological characteristics were observed using an Olympus BX 51 or 53 biological microscope equipped with an Olympus DP21 digital microscopy camera (Olympus), a Zeiss Axio Imager.A1 compound microscope equipped with a Zeiss Axiocam 506 colour digital camera (Zeiss, Jena, Germany), or a Leitz Ortholux microscope equipped with a Nikon 995 colour digital camera. The position of the primary septum in the ascospores was determined using a decimal system, expressed as the ratio of the upper hemisphere length to the total ascospore length according to the method described by Shoemaker (1984). Means ± standard deviation (SD) were calculated for sizes of asci, ascospores, conidiogenous cells, and conidia; length of ascus stalk; l/w ratios of ascospores and conidia; and ratios of the upper hemisphere length to the total ascospore length. Scatter plots of the measured spore sizes were created to confirm the morphological differences.
DNA extraction, PCR amplification, and sequencing
DNA was extracted from the cultures using the ISOPLANT II Kit (Nippon Gene, Tokyo, Japan) according to the manufacturer’s instructions, the DNeasy Plant Mini Kit (QIAgen GmbH, Hilden, Germany) according to Voglmayr and Jaklitsch (2011) and Jaklitsch et al. (2012), or the GeneJET Plant Genomic DNA Purification Kit (Thermo Fisher Scientific Inc., Waltham, MA, USA).
The following loci were amplified and sequenced: the small subunit (SSU) and large subunit (LSU) of the nuclear ribosomal DNA, the internal transcribed spacer (ITS) region, the second largest RNA polymerase II subunit (RPB2) gene, and the translation elongation factor 1-alpha (TEF1) gene. The primers and annealing temperatures used for the polymerase chain reaction (PCR) are listed in Suppl. material 1: table SS2. Most PCR products were purified using the FastGene Gel/PCR Extraction Kit (Nippon Gene), according to the manufacturer’s instructions and sequenced using the analysis services of SolGent (Daejeon, Korea). The PCR products from strains L20, L39, L104, L179, and L181 were purified using enzymatic PCR clean-up (Werle et al. 1994), as described by Voglmayr and Jaklitsch (2008). The DNA from the five strains was cycle-sequenced using the ABI PRISM Big Dye Terminator Cycle Sequencing Ready Reaction Kit v.3.1 (Applied Biosystems; Warrington, UK). Sequencing was performed using an automated DNA sequencer (ABI 3730xl Genetic Analyzer, Applied Biosystems). The newly generated sequences were deposited into the GenBank database (Table 1).
Phylogenetic and barcode analyses
The first phylogenetic analysis of the SSU, ITS, LSU, RPB2, and TEF1 sequences from 64 ingroup strains/specimens of Pleosporales was performed to clarify the familial placement of Splanchospora. Except for those of Splanchospora, the other 40 members of Pleosporales were retrieved from the GenBank database (see Table 1 for details on the sources of the sequences). Two members of Hysteriales were selected as outgroups. For the second phylogenetic analysis, single-gene trees based on the ITS, RPB2, and TEF1 sequences as well as a combined tree of these three loci were generated to assess the species boundaries of the 24 strains (29 strains in the ITS tree) within Splanchospora. The FASTA-format sequence alignments used in these analyses are available in the Suppl. materials 1–9.
The multiple sequence alignment program MUSCLE, implemented in MEGA v.7.0 (Kumar et al. 2016), was used to align the SSU, ITS, LSU, RPB2, and TEF1 sequences. Maximum-likelihood (ML) and Bayesian methods were used for the phylogenetic analyses. Kakusan4 software (Tanabe 2011) was used to estimate the optimum substitution models for each dataset, using the Akaike information criterion (Akaike 1974) or the corrected Akaike information criterion (AICc; Sugiura 1978) for the ML analysis and the Bayesian information criterion (BIC; Schwarz 1978) for the Bayesian analysis. The TreeFinder Mar 2011 program (http://www.treefinder.de) was used for the ML analysis based on the models selected using the AICc4 parameter. The ML bootstrap support (MLBS) values were obtained using 1,000 bootstrap replicates. The Bayesian analysis program MrBayes v.3.2.6 (Ronquist et al. 2012) was executed using substitution models selected on the basis of the BIC4 parameter. Two simultaneous and independent Metropolis-coupled Markov chain Monte Carlo (MCMCMC) runs were performed for 3,000,000, 2,000,000, and 1,500,000 generations for the first, second combined, and second single-gene analyses, respectively, with trees sampled every 1,000 generations. The convergence of the MCMCMC procedure was assessed on the basis of the effective sample size scores (all >100) using MrBayes and Tracer v.1.7.2 (Rambaut et al. 2018). The first 25% of trees were discarded during the burn-in phase, and the remainder were used to calculate the 50% majority-rule trees and determine the Bayesian posterior probability (BPP) values for individual branches. Statistical support was defined as full (100% MLBS/1.00 BPP) or high (≥90% MLBS/ ≥0.95 BPP). Values lower than 70% MLBS and 0.95 BPP were considered to be of low support and not indicated in phylogenetic trees.
Differences in SSU, ITS, LSU, RPB2, and TEF1 sequences amongst the various species of Splanchospora were calculated using MEGA v.7.0. Table 2 illustrates the variation in intraspecific conserved sites amongst the Splanchospora species.
Results
Molecular phylogenetic and barcode analyses
In the first analysis, ML and Bayesian phylogenetic trees were generated using an aligned sequence dataset comprising SSU, ITS, LSU, RPB2, and TEF1 sequences. The dataset and statistics resulting from the phylogenetic analyses (e.g. the number of characters and selected substitution models) are provided in Suppl. material 1: table S3. The ML tree with the highest log-likelihood (InL = −39,388.245) is shown in Fig. 1. The phylogram provided a higher confidence topology and values for familial classification than the individual gene trees did, with 20 families reconstructed in Pleosporales. The topology recovered by the Bayesian analysis was almost identical to that of the ML tree, except for the branching order between two Neomedicopsis species. Members of Splanchospora formed a monophyletic and fully supported clade (100% MLBS/1.0 BPP) within the family Neohendersoniaceae. Splanchospora was found to be closely related to the genus Neomedicopsis. However, their relationships were not fully supported, and Neomedicopsis appeared to be polyphyletic in the ML and Bayesian trees.
Maximum-likelihood (ML) tree of Pleosporales based on combined SSU, ITS, LSU, RPB2, and TEF1 regions. ML bootstrap support (MLBS) values above 70% and Bayesian posterior probability (BPP) values above 0.95 are presented at the nodes as MLBS/BPP. A hyphen (‘-’) indicates values lower than 70% MLBS or 0.95 BPP. The newly obtained sequences are shown in bold. The scale bar represents nucleotide substitutions per site. Ex-epitype, ex-holotype, ex-isotype, ex-neotype, ex-paratype, and ex-type strains are indicated as E, H, I, N, P, and T, respectively.
In the second analysis, ML and Bayesian phylogenetic trees based on the ITS, RPB2, and TEF1 sequences were generated using an aligned sequence dataset of members of Splanchospora. The statistics and models used for these datasets are listed in Suppl. material 1: table S3. The ML trees with the highest log-likelihoods (InL = −771.13881 in ITS, −1,919.8091 in RPB2, −2,353.4835 in TEF1, and −5,099.5852 in ITS-RPB2-TEF1) are shown in Fig. 2 and Suppl. material 1: fig. S1. The topology recovered by the Bayesian analyses was almost identical to that of the ML trees, except for the branching order amongst species in the TEF1 tree. The combined tree revealed six distinct lineages, indicating the existence of six species. These were the type species S. ampullacea and the cryptic species (Splanchospora sp., L181), both from Europe, and four newly identified species from Japan (Splanchospora fulviconidiasp. nov., Splanchospora microsporasp. nov., Splanchospora oblongiconidiasp. nov., and Splanchospora pseudomicrosporasp. nov.). Splanchospora ampullacea, S. microspora, and S. pseudomicrospora formed a highly supported monophyletic clade (≥97% MLBS/1.0 BPP). Splanchospora fulviconidia and S. oblongiconidia had low MLBS values (<70% MLBS/1.0 BPP; Fig. 2), whereas the single-gene trees based on RPB2 and TEF1 formed fully supported clades (100% MLBS/1.0 BPP) of the two species (Suppl. material 1: fig. S1B, C) and the ITS tree formed a fully supported clade of S. fulviconidia and a highly supported clade of S. oblongiconidia (91% MLBS/1.0 BPP) (Suppl. material 1: fig. S1A).
Maximum-likelihood (ML) tree of Splanchospora based on combined ITS, RPB2 and TEF1 regions. ML bootstrap support (MLBS) values above 70% and Bayesian posterior probability (BPP) values above 0.95 are presented at the nodes as MLBS/BPP. A hyphen (‘-’) indicates values lower than 70% MLBS or 0.95 BPP. The scale bar represents nucleotide substitutions per site. Ex-holotype, ex-neotype, and ex-paratype strains are indicated as H, N, and P, respectively.
Amongst the examined loci, interspecies variation was observed at 22 of 452 sites in ITS, 10 of 1,346 sites in SSU, 15 of 1,320 sites in LSU, 85 of 1,065 sites in RPB2, and 102 of 1,253 sites in TEF1 (Table 2). Of the 5,436 sites in total, the numbers of sites with distinct bases or gaps in each species compared with those in the other species were as follows: S. ampullacea, 6; S. fulviconidia, 117; S. microspora, 17; S. oblongiconidia, 28; S. pseudomicrospora, 14; and Splanchospora sp., 11. All the SSU sequences of S. ampullacea (CBS 116578 and L20) and S. pseudomicrospora (7 strains) contained a 318 bp insertion.
Taxonomy
Neohendersoniaceae
Taxon classificationFungiPleosporalesNeohendersoniaceae
Giraldo & Crous, Mycol. Progr. 16 (4): 343 (2017)
BC95C543-D81A-55C2-BE14-EF05EEACA462
Type genus.
Neohendersonia.
Description.
Sexual morph. Ascomata pseudoperithecial, scattered, immersed, erumpent at the ostiolar neck, globose to depressed globose or ampulliform, ostiolate. Ostiolar neck central to excentric. Ascomatal wall composed of brown cells. Pseudoparaphyses septate, branched and anastomosed. Asci bitunicate, fissitunicate, cylindrical to clavate or pyriform, pedicellate. Ascospores broadly ellipsoidal to clavate-ellipsoid or broadly fusiform, 1-septate to multi-septate or muriform, hyaline or pigmented, smooth or faintly roughened.
Asexual morph. Conidiomata pycnidial, solitary or gregarious, immersed, globose to depressed globose or ampulliform. Ostiolar neck central. Conidiomatal wall composed of brown cells. Paraphyses absent or present, septate, branched. Conidiophores absent. Conidiogenous cells discrete, determinate or indeterminate, annellidic or phialidic, cylindrical, lageniform, doliiform or ampulliform, hyaline, smooth. Conidia globose to ellipsoid, obovoid, cylindrical, clavate or fusiform, septate or aseptate, hyaline or pigmented. Spermatia subglobose to ellipsoidal.
Notes.
Neohendersoniaceae accommodates eight genera, Brevicollum, Crassiparies, Medicopsis, Muriformispora, Neobrevicollum, Neohendersonia, Neomedicopsis, and Splanchospora. ‘Amarenographium’ solium is also a member of this family, but its generic placement remains uncertain (Tanaka et al. 2017). Crassiparies (Li et al. 2016; Tanaka et al. 2017), Medicopsis (Devadatha et al. 2020), Muriformispora (de Silva et al. 2022; Wu et al. 2024), Neomedicopsis (Hyde et al. 2018; Crous et al. 2019), and Splanchospora (this study) are known to have both sexual and asexual morphs. On the other hand, Brevicollum (Tanaka et al. 2017) and Neobrevicollum (Lu et al. 2024) are known only from their sexual morphs, whereas Neohendersonia (Giraldo et al. 2017) and ‘Amarenographium’ solium (Hodhod et al. 2012) are known only from their asexual morphs. In this study, we show that the presence of paraphyses in the conidiomata of Splanchospora expands the morphological concept of the asexual stage in Neohendersoniaceae.
Splanchospora
Taxon classificationFungiPleosporalesNeohendersoniaceae
Lar.N. Vassiljeva, Nizshie Rasteniya, Griby i Mokhoobraznye Dalnego Vostoka Rossii 4: 237 (1998)
BF552B1D-3B7E-50B6-937F-1D066A97827C
Type species.
Splanchospora ampullacea *
Description.
Sexual morph. Ascomata pseudoperithecial, scattered, immersed, erumpent at the ostiolar neck, subglobose to depressed globose or ampulliform, ostiolate. Ostiolar neck central, developed or less developed. Ascomatal wall composed of brown cells. Pseudoparaphyses septate, branched and anastomosed. Asci bitunicate, clavate, short-stalked, with 8, rarely 2–7 ascospores. Ascospores clavate-ellipsoid, 1-septate, with a submedian septum, strongly constricted at the septum, thick-walled, pale to dark brown, smooth or faintly roughened, with an entire sheath.
Asexual morph. Conidiomata pycnidial, scattered, immersed, subglobose to depressed globose or ampulliform. Ostiolar neck central. Conidiomatal wall composed of brown cells. Paraphyses septate, branched. Conidiophores absent. Conidiogenous cells annellidic, cylindrical to ampulliform. Conidia globose to ellipsoid, aseptate, hyaline or yellowish brown, smooth, thick-walled.
Notes.
Splanchospora formed a monophyletic clade that was distinctly separated from any genera in the family Neohendersoniaceae (Fig. 1). Splanchospora is similar to the genera with sexual morphs in having immersed ascomata with a central ostiolar neck, except for Brevicollum, which has an excentric to ostiolar neck (Tanaka et al. 2017). However, Splanchospora differs from Crassiparies, Medicopsis, Muriformispora, Neobrevicollum, and Neomedicopsis in having clavate-ellipsoid, thick-walled ascospores strongly constricted at the submedian septum. Amongst the members of Neohendersoniaceae with asexual morphs, Neohendersonia and Neomedicopsis are more similar to Splanchospora in having annellidic conidiogenous cells (those of Neomedicopsis were described as ‘proliferating percurrently’ and illustrated in fig. 34B in Crous et al. 2019) and thick-walled conidia. Notably, Neomedicopsis shares long conidiogenous cells and aseptate conidia with Splanchospora and is sister to the latter genus. However, Splanchospora is distinguished from Neohendersonia and Neomedicopsis by the presence of paraphyses in the conidiomata.
Splanchospora
ampullacea
Taxon classificationFungiPleosporalesNeohendersoniaceae
(Pers.) Lar.N. Vassiljeva, Nizshie Rasteniya, Griby i Mokhoobraznye Dalnego Vostoka Rossii 4: 238 (1998)
6EF5D60F-95CB-568F-8092-BD1A2F2006B1
≡ Splanchnonema ampullaceum (Pers.) Shoemaker & P.M. LeClair, Canad. J. Bot. 53(15): 1570 (1975). = Massaria curreyi Tul. & C. Tul. (as ‘curreii’), Selecta Fungorum Carpologia, Tomus Secundus. Xylariei - Valsei - Sphaeriei 2: 231 (1863). ≡ Massariella curreyi (Tul. & C. Tul.) Sacc., Syll. Fung. 1: 717 (1882).
Basionym.
Sphaeria ampullacea Pers., Synopsis methodica fungorum: 41 (1801).
Scatter plot comparisons of spore size amongst Splanchospora species. A ascospores; B conidia on specimen; C conidia in culture. The vertical and horizontal axes indicate width and length, respectively. The purple, orange, red, green, blue, and gray circles represent spore sizes of S. ampullacea, S. fulviconidia, S. microspora, S. oblongiconidia, S. pseudomicrospora, and Splanchospora sp., respectively.
Splanchospora ampullacea. A–L Sexual morph. M–V Asexual morph. A Ascomata in face view. B Slimy mass of ascospores exuded though the ostiole of ascoma. C Ascoma in lateral section. D Ascoma in longitudinal section. E Ascomatal wall at side. F Pseudoparaphyses. G, H Asci. I–L Ascospores. M Conidiomata in face view. N Slimy mass of conidia exuded though the ostiole of conidioma. O Conidioma under bark. P Conidioma in longitudinal section. Q Conidiomatal wall at side. R Paraphyses. S Conidiogenous cells and immature conidia. T–V Conidia. Specimen: L179 (A–V). Scale bars: 1 mm (A, M); 200 μm (B–D, N–P); 20 μm (E–H, Q, R); 10 μm (I–L, S–V).
Splanchospora fulviconidia. A–L Sexual morph. M–Z Asexual morph. A Ascomata in face view. B Slimy mass of ascospores exuded though the ostiole of ascoma. C Ascoma in lateral section. D Ascoma in longitudinal section. E Ascomatal wall at side. F Pseudoparaphyses. G, H Asci. I–K Ascospores. L Germinating ascospore. M, O Conidiomata in face view (M on natural substrate, O on a rice straw in culture). N, P Slimy mass of conidia exuded though the ostiole of conidiomata (N on natural substrate, P on a rice straw in culture). Q Conidioma in longitudinal section. R Conidiomatal wall at side. S Paraphyses. T Conidiogenous cell and immature conidium (arrowheads indicate annellations). U, V, X–Z Conidia (U, V from specimens; X–Z from culture; X immature; Z senescent). W Germinating conidium. Specimens: RY 73 (A–E, G, H, J, L); RY 91 (F, K); RY 69 (I); RY 71 (M, N, Q, R, V, W); RY 112 (U). Strains: culture RY 92 (O, P, S, X); culture RY 71 (T, Y); culture RY 91 (Z). Scale bars: 1 mm (A, M, O); 200 μm (B–D, N, P, Q); 20 μm (E–H, R, S); 10 μm (I–L, T–Z).
Splanchospora microspora. A–L Sexual morph. M–Y Asexual morph. A Ascomata in face view. B Ostiole in face view. C Ascoma in lateral section. D Ascoma in longitudinal section. E Ascomatal wall at side. F Pseudoparaphyses. G, H Asci. I–K Ascospores. L Germinating ascospore. M, Q Conidiomata in face view (M on natural substrate, Q on a beech twig in culture). N, R Slimy mass of conidia exuded though the ostiole of conidiomata (N on natural substrate, R on a rice straw in culture). O Ostiole in face view. P Conidioma in lateral section. S Conidioma in longitudinal section. T Conidiomatal wall at side. U Paraphyses. V Conidiogenous cell and immature conidium. W–Y Conidia (W from specimen; X, Y from culture). Specimens: KT 2906 (A–L); KT 2905 (M–P, S, T, V, W). Strains: culture KT 2905 (Q, R, U, X); culture KT 2906 (Y). Scale bars: 1 mm (A, M, Q); 200 μm (B–D, N–P, R, S); 20 μm (E–H, T, U); 10 μm (I–L, V–Y).
Splanchospora oblongiconidia. A–L Sexual morph. M–Y Asexual morph. A Ascomata in face view. B Slimy mass of ascospores exuded though the ostiole of ascoma. C Ascoma in lateral section. D Ascoma in longitudinal section. E Ascomatal wall at side. F Pseudoparaphyses. G, H Asci. I–K Ascospores. L Germinating ascospore. M, O Conidiomata in face view (M on natural substrate, O on a rice straw in culture). N, P Slimy mass of conidia exuded though the ostiole of conidiomata (N on natural substrate, P on a beech twig in culture). Q Conidioma in longitudinal section. R Conidiomatal wall at side. S Paraphyses. T Conidiogenous cell (arrowheads indicate annellations). U Conidiogenous cell and conidium. V, X, Y Conidia (V from specimen; X, Y from culture). W Germinating conidium. Specimens: RY 83 (A–D, F–J, M, N, Q); AH 478 (E, K, L); KT 3610 (V, W). Strains: culture RY 83 (O, S–U, Y); culture AH 478 (P, R, X). Scale bars: 1 mm (A, M, O); 200 μm (B–D, N, P, Q); 20 μm (E–H, R, S); 10 μm (I–L, T–Y).
Splanchospora pseudomicrospora. A–M Sexual morph. N–X Asexual morph. A Ascomata in face view. B Slimy mass of ascospores exuded though the ostiole of ascoma. C Ascoma in lateral section. D Ascoma in longitudinal section. E Ascomatal wall at side. F Pseudoparaphyses. G–I Asci. J–L Ascospores. M Germinating ascospore. N, P Conidiomata in face view (N on natural substrate, P on a rice straw in culture). O, Q Slimy mass of conidia exuded though the ostiole of conidiomata (O on natural substrate, Q on a beech twig in culture). R Conidioma in longitudinal section. S Conidiomatal wall at side. T Paraphyses. U Conidiogenous cell and immature conidium (arrowheads indicate annellations). V–X Conidia (V from specimen; W, X from culture). Specimens: RY 50 (A–I, L); KT 3680 (J); KT 4246 (K, M); KT 4247 (N, O, R, S); KT 4324 (V). Strains: culture KT 4247 (P, Q, T); culture KT 4324 (U, X); culture RY 50 (W). Scale bars: 1 mm (A, N, P); 200 μm (B–D, O, Q, R); 20 μm (E–I, S, T); 10 μm (J–M, U–X).
Splanchospora sp. A, B Conidiogenous cells and conidia. C–E Conidia. Specimen: L181 (A–E). Scale bars: 10 μm (A–E).
Colony characters of Splanchospora species on PDA at 20 °C in the dark after 2 weeks. A–E Surface; F–J Reverse. Species: S. ampullacea (CBS 116578) (A, F); S. fulviconidia (culture RY 91 = MAFF 248153) (B, G); S. microspora (culture KT 2906 = MAFF 248150) (C, H); S. oblongiconidia (culture RY 83 = MAFF 248152) (D, I); S. pseudomicrospora (culture KT 4246 = MAFF 248151) (E, J). Scale bar: 1 cm.
Type.
AUSTRIA • Burgenland, Hornstein, Lindenallee, T. cordata Mill., 4 Nov 2017, H. Voglmayr & I. Greilhuber L179 (neotypeWU-MYC 0057753 here designated, MBT 10029611, isoneotypeHHUF 30704, ex-neotype CBS 154683 = culture L179).
Description.
Sexual morph. Ascomata pseudoperithecial, scattered, immersed, erumpent at the ostiolar neck, subglobose to depressed globose, 480–770 μm high, 530–800 μm diam, ostiolate. Ostiolar neck central, 80–120 μm long, 110–180 μm wide. Ascomatal wall (24–)35–45 μm thick at the sides, composed of 5–6 layers of polygonal, 10–21 × 5–8(–9) μm, brown cells. Pseudoparaphyses 2.5–4 μm wide, septate, branched and anastomosed. Asci bitunicate, clavate, 181–230(–245) × 30–39 μm (mean ± SD = 212.6 ± 13.3 × 34.4 ± 2.2 μm, n = 33), short-stalked (30–70 μm long, mean ± SD = 46.1 ± 11.1 μm, n = 33), 8-spored, rarely 4-, 6- or 7-spored due to meiotic failure or stunted spores, with uni- to bi-seriate ascospores. Ascospores clavate-ellipsoid, 30–45 × 13–18 μm (mean ± SD = 37.0 ± 3.4 × 15.6 ± 1.2 μm, n = 197), l/w = 1.9–3.1(–3.3) (mean ± SD = 2.4 ± 0.3, n = 197), 1-septate, with a submedian septum [(0.55–)0.58–0.67(–0.70), mean ± SD = 0.63 ± 0.03, n = 100], strongly constricted at the septum, brown to dark brown, smooth to faintly roughened, with an entire sheath.
Asexual morph. Conidiomata pycnidial, scattered, immersed, ampulliform, 310–560 μm high, 240–450 μm diam, ostiolate. Ostiolar neck central, 80–140 μm long, 100–220 μm wide. Conidiomatal wall 20–47 μm thick, composed of 4–6 layers of polygonal, 7.5–16 × 4.5–8 μm, brown cells. Paraphyses 2.5–3.5 μm wide, septate, branched. Conidiophores absent. Conidiogenous cells annellidic, cylindrical, (12–)15–23(–29) × 3–7.5(–11) μm (mean ± SD = 19.8 ± 5.1 × 5.8 ± 2.0 μm, n = 13). Conidia globose to subglobose, 14.5–22.5 × 14–17 μm (mean ± SD = 18.8 ± 1.7 × 15.1 ± 0.7 μm, n = 102), l/w = 1.1–1.4(–1.6) (mean ± SD = 1.2 ± 0.1, n = 102), aseptate, hyaline, smooth, with thick spore wall (1–1.5 μm).
Colonies on PDA (after 2 weeks) attaining a diam of 2.7–3.2 cm, white; reverse isabelline; no pigment produced.
Distribution.
Asia [Georgia (Nakhutsrishvili 1986), the Russian Far East (Vasilyeva 1998)]; Europe [Austria (this study), Belgium, Czech Republic, Denmark (Munk 1957), Finland, France, Germany, Hungary, Italy, Lithuania (Treigienė 1999), Luxembourg, Netherlands, Norway (Nordén et al. 2018), Poland (Mułenko et al. 2008), Portugal, Russia (Mel’nik 2011), Spain, Sweden (Eriksson 2014; this study), Switzerland (Tonolo 1956), UK (Pirozynski and Morgan-Jones 1968; Barr 1982; this study), Ukraine]; North America [Canada, USA (Barr 1982)]. Records of countries without references are based on specimens recorded in Cybertruffle (http://www.cybertruffle.org.uk/eng/), GBIF Secretariat (2023), or MyCoPortal (https://www.mycoportal.org/portal/index.php); true species identities of collections outside of Europe require additional studies.
Additional specimens examined.
AUSTRIA • Wien, 3. Bez., Botanical Garden of the University of Vienna (HBV), Tilia platyphyllos Scop., 4 Apr 2006, H. Voglmayr, L20 (WU-MYC 0057750 = HHUF 30703; CBS 122932 = culture L20); Wien, 21. Bez., Donauinsel near Nordbrücke, Tilia cf. cordata, 5 May 2013, W. Jaklitsch, L104 (WU-MYC 0057752, culture L104). UK • London, Richmond, Kew Gardens, Tilia sp., 11 Nov 2008, H. Voglmayr, L39 (WU-MYC 0057751, CBS 124477 = culture L39).
Additional strain examined.
SWEDEN • Uppland, Dalby parish, Ekbacken, Tilia cordata, 9 Mar 1991, K. & L. Holm, CBS 116578 = UPSC 3376.
Notes.
For further synonyms, please refer to Barr (1982). The location of the type specimen of Sphaeria ampullacea is unknown, and the original description of the species did not provide information on the ascospores (Persoon 1801). The only extant collection of Sphaeria ampullacea in the Persoon herbarium (L0112460) cannot represent the type, because this collection was evidently collected after its description (Persoon 1801), as brought forward in a detailed note by R. A. Shoemaker attached to this specimen (see https://bioportal.naturalis.nl/nl/multimedia/L0112460). To stabilize the species concept, we therefore here designate collection WU-MYC 0057753, for which detailed morphological data as well as sequences are available, as the neotype for Sphaeria ampullacea Persoon. Shoemaker and LeClair (1975) designated the neotype specimen of Massaria curreyi, which is a heterotypic synonym of Sphaeria ampullacea, and examined its ascospores. Although the ascospores from our specimens (30–45 × 13–18 μm) are broader than those from the neotype of Massaria curreyi (36–42 × 13–14.5 μm; Shoemaker and LeClair 1975), the values partially overlap. In our specimens, pycnidia with 1-celled conidia are closely associated with the species. Although the strains examined in this study were sterile in culture, we recognised the pycnidia on the natural specimens as being asexual morphs on the basis of their similarity to those of other Splanchospora spp.
Splanchospora
fulviconidia
Taxon classificationFungiPleosporalesNeohendersoniaceae
R. Yoshioka, Voglmayr & Kaz. Tanaka sp. nov.
CB97A67B-BB2D-5EB4-BCE9-526B4EFA716C
860946
Etymology.
From the Latin fulvi, meaning tawny, in reference to the tawny conidia.
Diagnosis.
Splanchospora fulviconidia can be easily distinguished from other species by its large-sized, yellowish-brown conidia.
Type.
JAPAN • Ibaraki: Tsukuba, Kannondai, Forest Research and Management Organization, Arboretum 2, dead twigs of Tilia japonica (Miq.) Simonk., 27 Mar 2024, R. Yoshioka, Y. Kudo & K. Tanaka, RY 91 (holotypeHHUF 30708, ex-type MAFF 248153 = culture RY 91).
Description.
Sexual morph. Ascomata pseudoperithecial, scattered, immersed, erumpent at the ostiolar neck, subglobose to depressed globose or ampulliform, 560–570 μm high, 500–600 μm diam, ostiolate. Ostiolar neck central, 90–120 μm long, 80–110 μm wide. Ascomatal wall 34–39 μm thick at the sides, composed of 4–5 layers of polygonal, 6–14.5(–17) × 4–7 μm, brown cells. Pseudoparaphyses 2.5–3.5 μm wide, septate, branched and anastomosed. Asci bitunicate, clavate, 162–212(–296) × 27.5–41(–44) μm (mean ± SD = 196.3 ± 23.0 × 34.1 ± 3.3 μm, n = 62), short-stalked (25–63 μm long, mean ± SD = 40.1 ± 11.5 μm, n = 11), 8-spored, rarely 2- or 6-spored due to meiotic failure or stunted spores, with bi- to tri-seriate ascospores. Ascospores clavate-ellipsoid to fusoid, 34.5–45 × 14–20 μm (mean ± SD = 39.4 ± 2.4 × 17.1 ± 1.2 μm, n = 128), l/w = 1.9–3.0 (mean ± SD = 2.3 ± 0.3, n = 128), 1-septate, with a submedian septum [0.54–0.62(–0.68), mean ± SD = 0.59 ± 0.02, n = 128), strongly constricted at the septum, pale brown to brown, smooth, with an entire sheath.
Asexual morph. Conidiomata pycnidial, scattered, immersed, subglobose to depressed globose or ampulliform, 460–490 μm high, 350–460 μm diam, ostiolate. Ostiolar neck central, 150–170 μm long, 100–110 μm wide. Conidiomatal wall 19–26 μm thick, composed of 4–5 layers of polygonal, 5–11(–12.5) × 2.5–5 μm, brown cells. Paraphyses 2.5–4 μm wide, septate, branched. Conidiophores absent. Conidiogenous cells annellidic, cylindrical to ampulliform, 11–17.5(–20) × 3–5 μm (mean ± SD = 15.6 ± 2.3 × 4.2 ± 0.6 μm, n = 11). Conidia ellipsoid to irregularly ellipsoid, 29.5–37(–38.5) × 15–19 μm (mean ± SD = 33.9 ± 1.9 × 17.0 ± 0.8 μm, n = 90), l/w = (1.7–)1.8–2.2(–2.3) (mean ± SD = 2.0 ± 0.1, n = 90), aseptate, pale to yellowish brown, becoming dark brown during senescence, smooth, with thick spore wall (2–3 μm).
Colonies on PDA (after 2 weeks) attaining a diam of 3.2–4 cm, white; reverse sienna; no pigment produced. In culture same coelomycetous asexual morph formed. Conidia 30.5–42 × 15–20 μm (mean ± SD = 36.2 ± 2.7 × 16.8 ± 1.0 μm, n = 150), l/w = (1.7–)1.8–2.5(–2.8) (mean ± SD = 2.2 ± 0.2, n = 150).
Distribution.
Japan.
Additional specimens examined.
JAPAN • Aomori: Aomori, Komagome, Fukazawa, Tsukimino Forest Park, dead twigs of T. japonica or T. maximowicziana Shiraz., 14 Oct 2023, R. Yoshioka, Y. Kudo & K. Tanaka, RY 69 (paratypeHHUF 30705, culture RY 69); ibid., RY 71 (paratypeHHUF 30706, culture RY 71); ibid., RY 73 (paratypeHHUF 30707, culture RY 73); ibid., dead twigs of T. japonica or T. maximowicziana, 9 Sep 2024, R. Yoshioka & Y. Kudo, RY 152 (paratypeHHUF 30711, culture RY 152); ibid., RY 153 (paratypeHHUF 30712, culture RY 153); ibid., RY 161 (paratypeHHUF 30713, culture RY 161); Ibaraki: Tsukuba, Kannondai, Forest Research and Management Organization, Arboretum 2, dead twigs of T. japonica, 27 Mar 2024, R. Yoshioka, Y. Kudo & K. Tanaka, RY 92 (paratypeHHUF 30709, culture RY 92); Tsukuba, Matsunosato, Forest Research and Management Organization, Arboretum 1, dead twigs of T. kiusiana, 27 Mar 2024, R. Yoshioka, Y. Kudo & K. Tanaka, RY 112 (paratypeHHUF 30710, culture RY 112).
Notes.
Splanchospora fulviconidia differs from all other Splanchospora species in having yellowish-brown conidia. Moreover, the conidia [29.5–37(–38.5) × 15–19 μm] are the largest amongst the six Splanchospora species examined. The position of the primary septum in the ascospores (mean 0.59) is more central than that of the other Splanchospora species (mean ≥0.63).
Splanchospora
microspora
Taxon classificationFungiPleosporalesNeohendersoniaceae
R. Yoshioka, Voglmayr & Kaz. Tanaka sp. nov.
F77A4A81-9641-5A26-923E-1E2079EA6F03
860947
Etymology.
From the Greek micro, meaning small, referring to the small ascospores and conidia of this species relative to those of other Splanchospora species in Japan.
Diagnosis. Splanchospora microspora can be distinguished from S. ampullacea in having a thicker conidial wall and being native to Japan.
Type. JAPAN • Aomori: Aomori, Komagome, Fukazawa, Tsukimino Forest Park, dead twigs of Tilia sp., 19 Sep 2011, K. Tanaka, KT 2906 (holotypeHHUF 30715, ex-type MAFF 248150 = culture KT 2906).
Description
Sexual morph. Ascomata pseudoperithecial, scattered, immersed, depressed globose, 320–570 μm high, 410–700 μm diam, ostiolate. Ostiolar neck central, 60–100 μm long, 80–200 μm wide. Ascomatal wall 23–30 μm thick at the sides, composed of 4–5 layers of polygonal, 6.5–16(–29) × 3–6.5(–8.5) μm, brown cells. Pseudoparaphyses 2–3 μm wide, septate, branched and anastomosed. Asci bitunicate, clavate, (118–)127–179 × 27.5–35 μm (mean ± SD = 155.4 ± 20.1 × 30.8 ± 2.4 μm, n = 14), short-stalked (23–50 μm long, mean ± SD = 37.5 ± 7.9 μm, n = 14), 8-spored, rarely 4-, or 6-spored due to meiotic failure, with uni- to tri-seriate ascospores. Ascospores clavate-ellipsoid, 25–36 × (11–)12–17.5 μm (mean ± SD = 30.0 ± 2.1 × 14.3 ± 1.3 μm, n = 137), l/w = (1.7–)1.8–2.5(–2.6) (mean ± SD = 2.1 ± 0.2, n = 137), 1-septate, with a submedian septum (0.59–0.71, mean ± SD = 0.65 ± 0.02, n = 137), strongly constricted at the septum, pale brown to brown, smooth, with an entire sheath.
Asexual morph. Conidiomata pycnidial, scattered, immersed, depressed globose, 280–500 μm high, 380–520 μm diam, ostiolate. Ostiolar neck central, 65–110 μm long, 90–160 μm wide. Conidiomatal wall 27–37 μm thick, composed of 4–6 layers of polygonal, 6–11.5 × 3–6(–7) μm, brown cells. Paraphyses 2–3 μm wide, septate, branched. Conidiophores absent. Conidiogenous cells annellidic, cylindrical to ampulliform, 6.5–19(–23) × 3–5(–6.5) μm (mean ± SD = 11.7 ± 3.8 × 4.7 ± 0.5 μm, n = 42). Conidia subglobose to ellipsoid, 19–25(–28) × 13–17 μm (mean ± SD = 22.3 ± 1.7 × 15.0 ± 0.9 μm, n = 80), l/w = 1.3–1.8(–1.9) (mean ± SD = 1.5 ± 0.1, n = 80), aseptate, hyaline, smooth, with thick spore wall (2.5–3 μm).
Colonies on PDA (after 2 weeks) attaining a diam of 4.6–4.8 cm, white; reverse buff; no pigment produced. In culture same coelomycetous asexual morph formed. Conidia 19–25.5 × 13–18 μm (mean ± SD = 23.1 ± 1.0 × 15.3 ± 0.9 μm, n = 112), l/w = (1.1–)1.3–1.7(–1.8) (mean ± SD = 1.5 ± 0.1, n = 112).
Distribution.
Japan.
Additional specimen examined.
JAPAN • Aomori: Aomori, Komagome, Fukazawa, Tsukimino Forest Park, dead twigs of Tilia sp., 19 Sep 2011, K. Tanaka, KT 2905 (paratypeHHUF 30714, culture KT 2905).
Notes.
Splanchospora microspora is phylogenetically close to S. ampullacea and Splanchospora sp. from Europe (Fig. 2, Suppl. material 1: fig. S1B). However, its conidial length [19–25(–28) μm, mean 22.3 μm] is somewhat longer than that of S. ampullacea (14.5–22.5 μm, mean 18.8 μm) and Splanchospora sp. (14–20.5 μm, mean 17.0 μm). Moreover, the conidial wall of S. microspora (2.5–3 μm) is thicker than that of S. ampullacea (1–1.5 μm) and Splanchospora sp. (0.9–1.1 μm).
Splanchospora
oblongiconidia
Taxon classificationFungiPleosporalesNeohendersoniaceae
R. Yoshioka, Voglmayr & Kaz. Tanaka sp. nov.
E4AE44F2-6CDB-5238-A9CD-113155A2E922
860948
Etymology.
Meaning oblong, referring to the longest conidia of Splanchospora species with hyaline ones.
Diagnosis.
Splanchospora oblongiconidia can be distinguished from S. microspora and S. pseudomicrospora by its longer conidia.
Type.
JAPAN • Aomori: Aomori, Katta, Heiwa Park, dead twigs of T. maximowicziana, 22 Oct 2022, Y. Kudo, RY 83 (holotypeHHUF 30718, ex-type MAFF 248152 = culture RY 83).
Description.
Sexual morph. Ascomata pseudoperithecial, scattered, immersed, erumpent at the ostiolar neck, subglobose to depressed globose, 310–560 μm high, 320–750 μm diam, ostiolate. Ostiolar neck central, 90–160 μm long, 175–200 μm wide. Ascomatal wall (19–)29–55(–60) μm thick at the sides, composed of 4–6 layers of polygonal, 7–19 × 3–10 μm, brown cells. Pseudoparaphyses 2.5–4 μm wide, septate, branched and anastomosed. Asci bitunicate, clavate, (125–)139–193 × 29.5–38(–42) μm (mean ± SD = 163.2 ± 16.9 × 34.3 ± 3.3 μm, n = 46), short-stalked (24–53 μm long, mean ± SD = 37.6 ± 7.5 μm, n = 35), 8-spored, rarely 2- to 7-spored due to meiotic failure or stunted spores, with uni- to tri-seriate ascospores. Ascospores clavate-ellipsoid, (28.5–)30–40.5 × (11.5–)14–18(–19.5) μm (mean ± SD = 34.7 ± 2.5 × 15.8 ± 1.3 μm, n = 111), l/w = 1.7–2.7(–3.1) (mean ± SD = 2.2 ± 0.3, n = 111), 1-septate, with a submedian septum [(0.57–)0.60–0.71, mean ± SD = 0.65 ± 0.03, n = 111], strongly constricted at the septum, pale brown to brown, smooth, with an entire sheath.
Asexual morph. Conidiomata pycnidial, scattered, immersed, erumpent at the ostiolar neck, subglobose, 470–490 μm high, 400–450 μm diam, ostiolate. Ostiolar neck central, 140–165 μm long, 100–125 μm wide. Conidiomatal wall 23–52 μm thick, composed of 4–5 layers of polygonal, 7.5–15(–17) × 3.5–8 μm, brown cells. Paraphyses 3–3.5 μm wide, septate, branched. Conidiophores absent. Conidiogenous cells annellidic, cylindrical to ampulliform, (14.5–)18.5–31 × 3–6 μm (mean ± SD = 23.1 ± 4.7 × 4.2 ± 0.9 μm, n = 10). Conidia ellipsoid to irregularly ellipsoid, 24–31.5 × 14–18 μm (mean ± SD = 26.6 ± 1.8 × 16.1 ± 0.9 μm, n = 100), l/w = 1.4–2.1 (mean ± SD = 1.7 ± 0.2, n = 100), aseptate, hyaline, smooth, with thick spore wall (2–3 μm).
Colonies on PDA (after 2 weeks) attaining a diam of 4–4.5 cm, olivaceous grey at central part, white at margin; reverse olivaceous grey at central part, buff at margin; no pigment produced. In culture same coelomycetous asexual morph formed. Conidia (24–)26–31(–35) × 14–18 μm (mean ± SD = 28.3 ± 1.4 × 15.5 ± 0.7 μm, n = 110), l/w = 1.5–2.1 (mean ± SD = 1.8 ± 0.1, n = 110).
Distribution.
Japan.
Additional specimens examined.
JAPAN • Hokkaido: Sapporo, Chuo-ku, Mt. Maruyama, dead twigs of Ostrya japonica, 17 Oct 2015, K. Tanaka, KT 3610 (paratypeHHUF 30717, culture KT 3610); Nagano: Ueda, Sugadaira Highland, University of Tsukuba, Mountain Science Center, Sugadaira Research Station, dead twigs of Carpinus cordata, 2 Jul 2016, A. Hashimoto, AH 478 (paratypeHHUF 30716, culture AH 478).
Notes.
The conidia of S. oblongiconidia (24–31.5 × 14–18 μm) are longer than those of S. microspora [19–25(–28) × 13–17 μm] and S. pseudomicrospora [20.5–25.5(–27) × 12.5–16 μm] and shorter those of than S. fulviconidia [29.5–37(–38.5) × 15–19 μm]. In addition to Tilia maximowicziana, S. oblongiconidia was found to be associated with Carpinus cordata and Ostrya japonica, indicating that Splanchospora spp. may have some latitude in host selection, although their main host is linden trees.
Splanchospora
pseudomicrospora
Taxon classificationFungiPleosporalesNeohendersoniaceae
R. Yoshioka, Voglmayr & Kaz. Tanaka sp. nov.
DF4E4780-B9B9-5052-B478-6205CE7B279E
860949
Etymology.
From the Greek pseudo, meaning spurious, in reference to morphological similarity to Splanchospora microspora.
Diagnosis.
It can be distinguished from S. microspora by its slightly elongated, slender conidia and variable-sized ascospores.
Type.
JAPAN • Aomori: Tsugaru, Kizukuritateoka, Bense Marsh, dead twigs of T. japonica, 28 Aug 2021, K. Tanaka, KT 4246 (holotypeHHUF 30721, ex-type MAFF 248151 = culture KT 4246).
Description.
Sexual morph. Ascomata pseudoperithecial, scattered, immersed, erumpent at the ostiolar neck, subglobose to depressed globose or ampulliform, 420–720 μm high, 410–560 μm diam, ostiolate. Ostiolar neck central, 170–200 μm long, 170–210 μm wide. Ascomatal wall (20–)30–39(–51) μm thick at the sides, composed of 4–5 layers of polygonal, 7–18(–22) × 3–7.5(–11) μm, brown cells. Pseudoparaphyses 2.5–4 μm wide, septate, branched and anastomosed. Asci bitunicate, clavate, 146–193(–212) × 27.5–36 μm (mean ± SD = 172.3 ± 16.5 × 31.5 ± 2.2 μm, n = 23), short-stalked (27–62 μm long, mean ± SD = 41.5 ± 9.7 μm, n = 17), 8-spored, rarely 2- to 7-spored due to meiotic failure or stunted spores, with uni- to bi-seriate ascospores. Ascospores clavate-ellipsoid, 27.5–37(–45.5) × 9.5–16(–20) μm (mean ± SD = 32.4 ± 3.2 × 13.7 ± 1.8 μm, n = 176), l/w = (1.8–)1.9–2.9(–3.3) (mean ± SD = 2.4 ± 0.3, n = 176), 1-septate, with a submedian septum (0.57–0.71, mean ± SD = 0.64 ± 0.02, n = 176), strongly constricted at the septum, pale brown to brown, smooth, with an entire sheath.
Asexual morph. Conidiomata pycnidial, scattered, immersed, erumpent at the ostiolar neck, ampulliform, 270–400 μm high, 500–550 μm diam, ostiolate. Ostiolar neck central, 210–350 μm long, 190–220 μm wide. Conidiomatal wall 23–30(–34) μm thick, composed of 4–5 layers of polygonal, 5.5–12(–15) × 2.5–6 μm, brown cells. Paraphyses 2.5–3.5 μm wide, septate, branched. Conidiophores absent. Conidiogenous cells annellidic, cylindrical to ampulliform, 12–22(–38) × (3–)5–8(–11) μm (mean ± SD = 18.2 ± 5.4 × 6.9 ± 1.9 μm, n = 30). Conidia ellipsoid to irregularly ellipsoid, 20.5–25.5(–27) × 12.5–16 μm (mean ± SD = 23.6 ± 1.5 × 14.5 ± 0.9 μm, n = 50), l/w = 1.3–1.9(–2.1) (mean ± SD = 1.6 ± 0.2, n = 50), aseptate, hyaline, smooth, with thick spore wall (2–3 μm).
Colonies on PDA (after 2 weeks) attaining a diam of 4.3–4.5 cm, white; reverse olivaceous grey, buff; no pigment produced. In culture same coelomycetous asexual morph formed. Conidia 22.5–29(–31) × 12.5–16 μm (mean ± SD = 25.4 ± 1.5 × 14.3 ± 0.8 μm, n = 230), l/w = (1.5–)1.6–2.0(–2.1) (mean ± SD = 1.8 ± 0.1, n = 230).
Distribution.
Japan.
Additional specimens examined.
JAPAN • Iwate: Hachimantai, Hirakasa, Iwate-san SA, dead twigs of T. maximowicziana or T. platyphyllos, 13 Aug 2016, KT 3680 (paratypeHHUF 30719, culture KT 3680); ibid., KT 3681 (paratypeHHUF 30720, culture KT 3681); ibid., dead twigs of T. maximowicziana, 2 Jan 2022, K. Tanaka, KT 4283 (paratypeHHUF 30723, culture KT 4283); Aomori: Tsugaru, Kizukuritateoka, Bense Marsh, dead twigs of T. japonica, 28 Aug 2021, K. Tanaka, KT 4247 (paratypeHHUF 30722, culture KT4247); Hirosaki, Shimoshirogane, Hirosaki Castle Botanical Garden, dead twigs of T. japonica, 5 Jun 2022, K. Tanaka, KT 4324 (paratypeHHUF 30724, culture KT 4324); Aomori, Komagome, Fukazawa, Tsukimino Forest Park, dead twigs of T. japonica or T. maximowicziana, 19 Sep 2024, R. Yoshioka & Y. Kudo, RY 154 (paratypeHHUF 30726, culture RY 154); ibid., RY 158 (paratypeHHUF 30727, culture RY 158); Hokkaido: Sapporo, Shiroishi-ku, Nango-dori Avenue, Bansei Park, dead twigs of T. japonica, 18 Jun 2023, R. Yoshioka, RY 50 (paratypeHHUF 30725, culture RY 50).
Notes.
Although S. pseudomicrospora is similar to S. microspora, the conidia of the former (mean 23.6 × 14.5 μm in the specimen and 25.4 × 14.3 μm in culture) are slightly longer and narrower than those of the latter (mean 22.3 × 15.0 μm in the specimen and 23.1 × 15.3 μm in culture) (Fig. 3B, C). These two species differ in various sequences: namely, at four sites with one gap in the ITS (99.1% homology = 448/452), at 27 sites with a single amino acid substitution in RPB2 (97.5% = 1,038/1,065), and at 24 sites with four gaps and a single amino acid substitution in TEF1 (98.0% = 1,206/1,230).
The ascospores size [27.5–37(–45.5) × 9.5–16(–20) μm (n = 176)] of S. pseudomicrospora is highly variable. This is probably due to occasional meiotic failure or stunted spores, resulting in enlargement of the remaining ascospores in the ascus. The size of ascospores from 8-spored asci are 27.5–37 × 11–15.5 μm (n = 54), whereas those from 4-spored asci are 38–40 × 17.5–20 μm (n = 4). This variability makes the identification of Splanchospora species on the basis of the ascospores alone more difficult.
Splanchospora
Taxon classificationFungiPleosporalesNeohendersoniaceae
sp.
ADC2BF09-AD45-5B61-9000-B7B205E14BD5
Description.
Sexual morph. Not observed.
Asexual morph. Conidiogenous cells annellidic, cylindrical, 13–24 × 3–4 μm (mean ± SD = 17.0 ± 4.4 × 3.6 ± 0.3 μm, n = 4). Conidia globose to subglobose, 14–20.5 × 12.5–16 μm (mean ± SD = 17.0 ± 1.2 × 13.7 ± 0.6 μm, n = 56), l/w = 1.0–1.4 (mean ± SD = 1.2 ± 0.1, n = 56), aseptate, hyaline, smooth, with a spore wall thickness of 0.9–1.1 μm.
Specimen examined.
AUSTRIA • Wien, 3. Bez., Botanical Garden of the University of Vienna (HBV), T. cordata, 31 Jan 2018, H. Voglmayr, L181 (WU-MYC 0057754, culture L181).
Notes.
Splanchospora sp. is most closely related to S. ampullacea (Fig. 2, Suppl. material 1: fig. S1A, B). Sequence differences exist between these two species: namely, at two sites in the ITS (99.6% homology = 450/452), eight sites with a single amino acid substitution in RPB2 (99.2% = 1,057/1,065), and 14 sites with a single amino acid substitution in TEF1 (98.9% = 1,216/1,230). The conidia of the Splanchospora sp. (mean 17.0 × 13.7 μm) are slightly smaller than those of S. ampullacea (mean 18.8 × 15.1 μm). However, the only specimen (L181) of this fungus lacks sexual morph and was collected in the same place as a specimen of S. ampullacea (L20). For now, we have postponed the description of this fungus as a new species, pending the availability of additional materials with sexual morphs to avoid confusion owing to premature decision.
Discussion
In this study, we have clarified the asexual morphs and phylogenetic position of Splanchospora for the first time. We provided a revised generic concept of Splanchospora on the basis of both its sexual and asexual characteristics and demonstrated the taxonomic validity of this genus in Neohendersoniaceae. Additionally, we identified four new species and discovered one undescribed species in this genus.
Splanchospora ampullacea was previously classified as Splanchnonema on the basis of the generic characteristics of having clavate, asymmetrically septate ascospores (Shoemaker and LeClair 1975). The genus Splanchnonema was established in 1829 with Splanchnonema pustulatum (current name: Splanchnonema foedans) as the type species and the asexual morph was revealed by Minter and Cannon (2013). In the asexual stage, these two genera share aseptate, thick-walled conidia and paraphyses (the latter in Splanchnonema were described as ‘paraphysis-like hyphae’ in Minter and Cannon 2013). They differ, however, in that the conidia of Splanchospora are globose to ellipsoid, while those of Splanchnonema are cylindrical to narrowly elongate. In the sexual stage, Splanchospora has 1-septate, thick-walled ascospores, whereas Splanchnonema has 2(rarely 3)-septate ascospores with an acute base (Zhang et al. 2012; Minter and Cannon 2013; Senn-Irlet et al. 2021).
Splanchnonema is known to be a polyphyletic group. To advance its taxonomic revision, information from the asexual morph is likely to be key for phylogenetic inference. For example, species whose asexual morphology differs from that of Splanchnonema sensu stricto have been excluded from this genus when considered together with molecular phylogenetic data: they are Splanchnonema platani (Ces.) M.E. Barr [current name: Macrodiplodiopsis desmazieri (Mont.) Petr.] with pycnidia producing mostly 3-septate, pigmented conidia (Crous et al. 2015, Macrodiplodiopsidaceae), Splanchnonema phorcioides (I. Miyake) P. Leroy, L. Gauthier & M.E. Barr, [current name: Pseudosplanchnonema phorcioides (I. Miyake) S. Konta, Camporesi & K.D. Hyde] with phoma-like asexual morph (Chethana et al. 2015, Massarinaceae), and Splanchnonema kalakadense Subram. & Sekar and Splanchnonema quercicola M.E. Barr (both are currently treated as Helminthosporium) with tretic, distoseptate conidia borne on erect conidiophores (Subramanian and Sekar 1987; Barr 1993; Voglmayr and Jaklitsch 2017, Massarinaceae). Splanchnonema pupula (Fr.) Kuntze, which produces acervuli with muriform conidia bearing lateral appendages (Tonolo 1956; Voglmayr and Jaklitsch 2008), should not be retained within Splanchnonema but should instead be placed in a separate genus, as previously proposed under the name Stigmatomassaria (nom. inval., Munk 1953). Further culture studies of other Splanchnonema species and the observation of their asexual morphs are expected to provide important clues to their phylogenetic placement. In addition, obtaining sequence data for the type species of Splanchnonema and clarifying its phylogenetic position remain essential tasks for the future.
In the genus Splanchospora, we identified the type species and one undescribed species from Europe, and described four new species from Japan. Although the ascospore sizes amongst these species overlapped except for the differentiation between S. fulviconidia and S. microspora (Fig. 3A), their conidial sizes (Fig. 3B, C), shapes, colours, wall thicknesses, and homology in DNA sequences were distinct. All six Splanchospora species were associated with dead twigs of linden trees. Our findings further support the host preference of Splanchospora for linden while also noting their occasional occurrences on Fagales trees, such as Carpinus betulus (Cybertruffle http://www.cybertruffle.org.uk/cgi-bin/robispec.pl?&colno=103169&glo=eng) and Quercus macrocarpa (The Kew Data Portal, https://records.data.kew.org/, Catalogue number K-M000458185) for S. ampullacea, and C. cordata and O. japonica for S. oblongiconidia (this study). Splanchospora species have multiple hosts and show overlap in their hosts. Our S. ampullacea specimens were associated with T. cordata and T. platyphyllos in Europe. The host, T. japonica, is shared by S. fulviconidia and S. pseudomicrospora in Japan. Interestingly, S. fulviconidia (RY 152) and S. pseudomicrospora (RY 154) were collected from the same linden individual at the same time. This indicates that although the two species co-exist in the same habitat, they are reproductively isolated.
Splanchospora ampullacea had been considered to be distributed throughout the mid-latitudes of the Northern Hemisphere and the only species in this genus. However, some specimens identified as S. ampullacea may be distinct species. The specimens in North America (Barr 1982) and the Russian Far East (Vasilyeva 1998) are not likely to be S. ampullacea. Based on specimens with identified host species, S. ampullacea in Europe is associated with T. cordata (Mel’nik 2011), T. platyphyllos (Nordén et al. 2018), Tilia × europaea K.Koch (as Tilia × vulgaris, The Kew Data Portal, Catalogue number K-M000084479), and Tilia tomentosa Moench (as T. argentea, Tonolo 1956), whereas the North American individuals are associated with T. americana and rarely T. cordata (MyCoPortal https://www.mycoportal.org/portal/index.php) and those from the Russian Far East are associated with Tilia amurensis Rupr. (Vasilyeva 1998). Considering the confusion caused by non-native linden trees, the fungal populations in these two regions may be considered distinct from those of the European species, reflecting differences in their original host species. As we identified four species of Splanchospora in Japan and a cryptic species of S. ampullacea in Europe, it is highly likely that multiple species also exist in North America and the Russian Far East.
The emphases on the ascospore morphology and host preference for species identification may be the reason why S. ampullacea has been the only species recognised for a long time. In this study, we found that the ascospore size was very similar amongst Splanchospora species (Fig. 3A). We also clarified that the preference for linden as the host is a common trait in this entire genus rather than a specific trait of S. ampullacea. Our results reveal that asexual characters, especially conidial features, combined with molecular data, provide valuable markers for species identification.
Conclusion
By providing new morphological, ecological, and molecular data, this study enhances our understanding of both the sexual and asexual morphs of Splanchospora and clarifies their interspecific relationships. On the basis of phylogenetic analyses and comparative morphological studies, we re-enforce the generic circumscription of Splanchospora and classify this genus in the family Neohendersoniaceae (Pleosporales, Dothideomycetes). Furthermore, we recognise six species within this genus. Members of Splanchospora exist on dead twigs of Tilia spp. and co-exist on the same host species within their native region. Conidial morphology and secondary DNA barcodes, such as TEF1 and RPB2, are useful for identifying species in this genus and will reveal unknown species diversity.
Supplementary Material
XML Treatment for Neohendersoniaceae
XML Treatment for Splanchospora
XML Treatment for Splanchospora ampullacea
XML Treatment for Splanchospora fulviconidia
XML Treatment for Splanchospora microspora
XML Treatment for Splanchospora oblongiconidia
XML Treatment for Splanchospora pseudomicrospora
XML Treatment for Splanchospora
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ahmed SA, van de Sande WWJ, Stevens DA et al. (2014) Revision of agents of black-grain eumycetoma in the order Pleosporales. Persoonia 33: 141–154. 10.3767/003158514 X 684744 PMC 431293025737597 · doi ↗ · pubmed ↗
- 2Akaike H (1974) A new look at the statistical model identification. IEEE Transactions on Automatic Control 19(6): 716–723. 10.1109/TAC.1974.1100705 · doi ↗
- 3Ariyawansa HA, Hyde KD, Jayasiri SC et al. (2015) Fungal diversity notes 111–252—taxonomic and phylogenetic contributions to fungal taxa. Fungal Diversity 75: 27–274. 10.1007/s 13225-015-0346-5 · doi ↗
- 4Ashrafi S, Wennrich JP, Becker Y et al. (2023) Polydomus karssenii gen. nov. sp. nov. is a dark septate endophyte with a bifunctional lifestyle parasitising eggs of plant parasitic cyst nematodes (Heterodera spp.). IMA Fungus 14: 6. 10.1186/s 43008-023-00113-w PMC 1006453836998098 · doi ↗ · pubmed ↗
- 5Barr ME (1982) On the Pleomassariaceae (Pleosporales) in North America. Mycotaxon 15: 349–383. 10.5962/p.417409 · doi ↗
- 6Barr ME (1993) Notes on the Pleomassariaceae. Mycotaxon 49: 129–142. 10.5962/p.416488 · doi ↗
- 7Cannon PF, Minter DW (2014) Splanchnonema ampullaceum. IMI Descriptions of Fungi and Bacteria 2000. 10.1079/DFB/20143338515 · doi ↗
- 8Chethana KWT, Liu M, Ariyawansa HA et al. (2015) Splanchnonema-like species in Pleosporales: introducing Pseudosplanchnonema gen. nov. in Massarinaceae. Phytotaxa 231(2): 133–144. 10.11646/phytotaxa.231.2.2 · doi ↗