Morpho-molecular approach reveals three novel endophytic fungi in Polyschema (Pleosporales, Latoruaceae) associated with roots of baobab trees in Yunnan, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
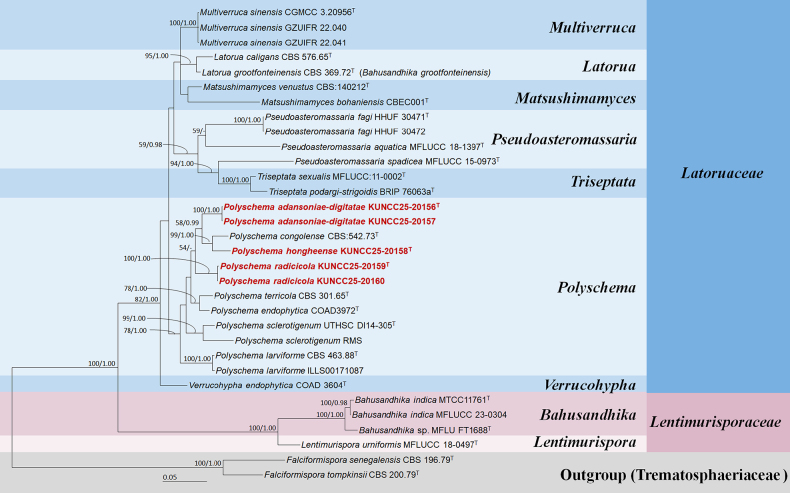 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4| Species names | Strain numbers | GenBank accession numbers | |||
|---|---|---|---|---|---|
|
|
|
| |||
|
| – |
| – | – | |
|
|
|
| – | – | |
|
|
|
|
| – | |
| MFLU FT1688 |
|
|
| – | |
|
|
|
|
|
| |
|
|
|
|
|
| |
|
|
|
| – | – | |
|
| – |
|
| – | |
|
| CBEC001T |
|
| – | – |
|
|
|
| – | – | |
|
|
|
| – | – | |
|
|
|
| – | – | |
|
|
|
| – | – | |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| – |
| |
|
| COAD3972T |
|
| – | – |
|
|
|
|
|
|
|
|
| – |
| – | – | |
|
| ILLS00171087 |
| – | – | – |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| – | – | |
|
|
|
| – | – | – |
|
|
|
|
|
| |
|
|
|
|
| – | |
|
|
|
|
| – | |
|
|
|
| – | – | |
|
|
|
| – | – | |
|
|
|
| – | – | |
|
|
|
|
| – | |
|
|
|
| – | – | |
| Group | Morphological characteristics | Sequence data | References | ||||
|---|---|---|---|---|---|---|---|
| Conidiophores | Conidiogenous cells | Conidia | Chlamydospores | ||||
| Group I |
| Macronematous or micronematous, hyaline, aseptate, or reduced to conidiogenous cells | 4–6.9 × 2.8–5.6 µm, globose or clavate, mono- or polytretic, discrete, smooth-walled | 18.5–27.6 × 9.7–14.3 µm, (1–)2–3-septate, ellipsoidal to obovoid, or oblong, brown to dark brown, versicolored, echinulate | 11.2–18.7 × 9.1–12.5 µm, terminal or intercalary, appear in short chains, globose to subglobose or ovoid, dark brown, 0–1-septate, thick, and smooth-walled | This study | |
|
| Micronematous and finely echinulate | 4–6 μm diam., spherical to subspherical, mono- or polytretic, discrete, sometimes in short chains | 13.5–49.5 × 6–11.5 um, 3–9-septate, commonly with 4–6 septa, clavate, obclavate, cylindrical to sigmoid, pale brown, finely echinulate, mostly formed singly, sometimes in short chains | Absent | Unavailable |
| |
|
| Mostly reduced to conidiogenous cells | 4.5–9.5 × 3–6 μm, spherical or enlarged at the apices, mono- or polytretic, discrete, smooth-walled, or slightly echinulate | 16–26 × 8–11 μm, 2–3-septate, commonly with 2 septa, clavate, ellipsoidal, brown, finely echinulate | Absent |
| ||
|
| 2.6–4.6 × 3.3–6.6 µm, short and subglobose, sometimes elongate, and curved, macronematous, or micronematous, or reduced to conidiogenous cells, septate, smooth to finely echinulate | 4.6–6.9 × 4–8.9 µm, globose or clavate, mono- or polytretic, discrete, smooth-walled, or slightly echinulate | 12.8–36.6 × 7.4–13.6 µm, 1–4-septate, commonly with 1–2 septa, fusoid-ellipsoidal, pale brown to dark brown, verruculose or echinulate | Absent | This study | ||
|
| Absent | 6–8 μm diam., spherical or subspherical, monotretic, discrete, verruculose or echinulate | 15–30 × 10–14 μm, 2–4-septate, ellipsoidal, or pyriform, dark golden brown, echinulate or verruculose | Absent | Unavailable |
| |
|
| Semi-micromematous and verruculose | 5–9 μm diam., spherical to subspherical, mono- or polytretic, discrete, solitary | 30–80 × 16–20 μm, 4–12-septate, commonly with 7–10 septa, clavate, obclavate, or ellipsoidal, sometimes sigmoid, dark reddish brown, verrucose | Absent | |||
|
| Absent | 6–9 μm diam., spherical or subspherical, monotretic, discrete, sometimes in short chains and flattened dorsiventrally, echinulate | 17–37 × 6–9 μm, 2–7-septate, cylindrical, pale to mid golden brown, verruculose or echinulate | Absent | Unavailable |
| |
|
| Absent | 6–10 × 5–6.5 μm, subglobose or clavate, mono- or polytretic, discrete, verruculose | 20–36 × 9–13 µm, 2–4-septate, commonly with 3 septa, cylindrical to ellipsoidal, pale brown at both ends, dark brown at the middle, verruculose | Absent | Unavailable |
| |
|
| 7.2–10.3 × 3.4–6.1 µm, unbranched, clavate or short cylindrical, micronematous, or reduced to conidiogenous cells | 4.4–7.2 × 4.3–7.3 µm, globose, clavate or short cylindrical, monotretic, discrete, smooth, or slightly verrucose | 20.4–40.9 × 7.5–13.1 µm, 2–5-septate, cylindrical, pale brown, verruculose or echinulate | Absent | This study | ||
|
| 6.6–59.4 × 3.3–4.9 μm, unbranched or frequently bifurcated at base, 1–4-septate, smooth, or rarely echinulate | 6.6–13.2 × 3.3–4.9 μm, globose, mono- or polytretic, discrete, smooth, or finely echinulate | 5–20 × 3–8 µm, 1–5-septate, commonly with 3 septa, cylindrical, pale brown, echinulate | Absent | Unavailable |
| |
| Group I |
| Up to 45 µm long, 2.5–3 µm wide, unbranched, subcylindrical, smooth or verruculose | 5–7.5 × 5–7 µm, spherical to subspherical, monotretic, rarely polytretic, discrete, sometimes in short chains, verruculose to tuberculate | 36–65(–75) × 7–8 µm, 5–16-septate, cylindrical, or sigmoid, brown to dark brown, verruculose to tuberculate, with | Absent |
| |
|
| Absent | 4–8 μm diam., spherical to subspherical, mostly monotretic, discrete, smooth or verruculose | 15–35 × 13–18 μm, 0–3-septate transversally, and sometimes 1 longitudinal or oblique septum, subspherical, pyriform or clavate, dark brown, verruculose | 8–15 μm diam. |
| ||
|
| 6–9(–16) μm long, clavate or doliiform, 0–1-septate, slightly verruculose | 6.5–8.5 × 6–8 μm, globose or widely obovate, mostly polytretic, discrete, verruculose | (15–)26–73 × 7–10 μm, (2–)3–11-septate, cylindrical, brown, finely verruculose | Absent | Unavailable |
| |
|
| Semi-micronematous and verruculose | 4.5–5.5 × 3–4.5 μm, subglobose (or 9.5 × 4 μm, clavate), mono- or polytretic, discrete, sometimes in short chains | 9.5–28.5 × 6–11.5 μm, 0–5-septate, commonly with 2–3 septa, clavate, straight or curved, rarely sigmoid, greyish brown to dark brown, finely echinulate, mostly solitary or in pairs, sometimes in short chains of 2–3 | 12.5–17.5 × 10.5–14.5 μm, numerous, solitary in chains or in dense clusters, globose to oval, occasionally with transverse and longitudinal septa, verrucose or smooth | Unavailable | ||
|
| Micronematous | Intercalary, incorporated in repent and aerial hyphae, laterally forming one or more conidia directly | (13–)15–22 × 6–9 μm, (2–)3-septate, cylindrical to elliptical, dark brown at the middle, subhyaline at both ends, smooth to verruculose | Absent | Unavailable |
| |
| Group II |
| 25–50 × 3–4 µm, mostly moniliform, semi-macronematous, mononematous, smooth-walled | 5–8 × 3–3.5 µm, globose, doliiform to ellipsoid, monotretic, determinate, terminal, sometimes sympodially proliferation become intercalary, smooth-walled | 25–29 × 8–10 µm, 4–5-septate, mostly fusiform to navicular, rarely clavate, dark brown to black at the middle, pale brown at both ends, smooth-walled, with a schizolytic conidial secession | Absent | Unavailable |
|
|
| Absent | 3–5 µm diam., spherical to subspherical, monotretic, discrete, smooth-walled | 10–12 × 5–6 µm, 1-septate, cylindrical to clavate, reddish brown, smooth-walled, with germ pore present | Absent | Unavailable |
| |
|
| Micronematous, mononematous, integrated, inconspicuous | Smooth, 4–7 μm in diam., spherical or subspherical, monotretic, smooth-walled | 30–46 × 12–16 μm, 3–5-septate, cylindrical to clavate, pale to rather dark reddish brown with a paler basal cell, smooth-walled | Absent | Unavailable | ||
|
| Micronematic or semi-macronematic | 4–8.5 × 4.6 μm, subspherical or ampulliform, monotretic, smooth-walled | 11–20 × 4.8–8 μm, 1–3-septate, cylindrical to ellipsoidal or clavate, brown to dark brown, smooth-walled | Absent | Unavailable |
| |
|
| Up to 45 μm tall, 3–5 μm wide, micronematous, mononematous | 6–8.5 × 2–7 μm, hemispherical, subspherical or cylindrical, monotretic, discrete, smooth-walled | Aseptate, subglobose to ellipsoidal, dark brown, smooth-walled | Absent |
| ||
|
| Micronematous, mononematous | 2.8–4.2 μm diam., spherical or subspherical, monotretic, rarely polytretic, discrete, or integrated, determinate, sometimes in short chains of 2–3, smooth-walled or verrucose | 14–21 μm long, 4.2–7 μm at base, and 7–11.2 μm wide at apex, always 1-septate, obovoid or pyriform, pale to mid dark brown, smooth-walled | Formed in culture | Unavailable |
| |
| Group II |
| 20–38 × 5–7 µm, moniliform, smooth-walled | 8–10 × 7–10 µm, globose or clavate, monotretic, determinate, smooth-walled | 41–45 × 19–22 µm, 2–3(–5)-septate, clavate to ovoid, brown, dark brown to black at the septa, smooth-walled | Absent | Unavailable |
|
|
| 11–20 × 4–5 µm, moniliform, smooth-walled | 5–6 × 3–5 μm, globose or clavate, monotretic, determinate, smooth-walled | 30–48 × 6–7 μm, (4–)5-septate, obclavate, brown at the middle and pale brown at the ends, smooth-walled, seceding schizolytically | Absent | Unavailable |
| |
|
| Micronematous, mononematous | 3–4.5 μm diam., spherical, monotretic, determinate, smooth-walled | 21–31 × 7–11 μm, usually 3–5-septate transversally, and generally one vertical septum towards the lower end, mid brown to dark or blackish brown, dark brown to black at the septa, smooth-walled | Absent | Unavailable |
| |
| Genera | Species | Life mode | Habitat | Isolation source | Geographic contribution | References |
|---|---|---|---|---|---|---|
|
|
| Saprobic | Terrestrial | Soil | Brazil |
|
|
| Saprobic | Terrestrial | Brown sandy soil | South Africa |
| |
|
|
| Saprobic | Terrestrial | Soil | India |
|
|
| Saprobic | Terrestrial | Decaying leaf | Cuba |
| |
|
|
| Saprobic | Terrestrial | Soil | China (Zhejiang) |
|
|
|
| Endophytic | Terrestrial | Baobab roots | China (Yunnan) | In this study |
|
| Saprobic | Terrestrial | Twig of unidentified plant | Venezuela |
| |
|
| Saprobic | Terrestrial | Twig of | USA (Illinois) |
| |
|
| Saprobic | Terrestrial | Soil | India |
| |
|
| Saprobic | Terrestrial | Fallen wood of | Mexico and USA (California) | ||
|
| Saprobic | Terrestrial | Tropical forest soil | Democratic Republic of the Congo |
| |
|
| Saprobic | Terrestrial | Dead leaves and petioles of | Cuba |
| |
|
| Endophytic | Terrestrial | Healthy roots of coffee plant | Brazil |
| |
|
| Endophytic | Terrestrial | Baobab roots | China (Yunnan) | In this study | |
|
| Saprobic | Terrestrial | Grassland soil | India |
| |
|
| Saprobic | Terrestrial | The cut surface of old firewood | USA (New York) |
| |
|
| Saprobic | Terrestrial | The decorticated wood | India |
| |
|
| Saprobic | Terrestrial | Mangrove debris | Mexico |
| |
|
| Saprobic | Terrestrial | Fallen, decaying leaves of an unidentified plant in rainforest | Cuba |
| |
|
| Saprobic | Terrestrial | Old culms of | USA (New Jersey) |
| |
|
| Saprobic | Terrestrial | Forest soil | Australia |
| |
|
| Endophytic | Terrestrial | Baobab roots | China (Yunnan) | In this study | |
|
| Saprobic | Terrestrial | Red-colored soil of | India |
| |
|
| Pathogenic | Terrestrial | Human clinic tissue | USA (Texas) |
| |
|
| Saprobic | Terrestrial | Soil | Brazil |
| |
|
| Saprobic | Terrestrial | Leaves of | China (Taiwan) |
| |
|
| Saprobic | Terrestrial | Soil | India |
| |
|
| Saprobic | Terrestrial | Forest soil | Japan |
| |
|
| Saprobic | Terrestrial | Dead unidentified twigs | India |
| |
|
|
| Saprobic | Freshwater | Decaying wood submerged in freshwater | Thailand |
|
|
| Pathogenic | Terrestrial | Twigs of | Japan |
| |
|
| Saprobic | Freshwater | Submerged wood in freshwater | Thailand |
| |
|
|
| Saprobic | Terrestrial | Dried branches of an unidentified plant | Thailand |
|
|
| Saprobic | Terrestrial | Decayed wood of | Australia |
| |
|
|
| Endophytic | Terrestrial | Macaw palm roots | Brazil |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Microbial Natural Products and Biosynthesis
Introduction
Latoruaceae is a family within Pleosporales, Dothideomycetes, that was introduced by Crous et al. (2015a) to accommodate the genera Latorua and Polyschema, with Latorua designated as the type genus and Latorua caligans as the type species. The teleomorph of Latoruaceae is characterized by immersed, compressed, globose ascomata with a papillate ostiole, 8-spored, bitunicate, fissitunicate, cylindrical to clavate asci with a clear ocular chamber, and 1–3-septate, fusiform, brown ascospores (Ariyawansa et al. 2015). The teleomorph is mostly presented as dark brown, verruculose hyphae (Crous et al. 2024) and is characterized by erect, moniliform, brown conidiophores or reduced conidiogenous cells and smooth to verruculose, brown, polyblastic conidiogenous cells or reduced inconspicuous loci on hyphae, bearing smooth or verrucose, septate, fusoid-ellipsoidal conidia in chains or solitary, sometimes becoming cupulate, with secondary conidia and cells or septa darker pigmented than the rest of the conidium (Crous et al. 2015a). Besides Latorua, six other genera, including Matsushimamyces (Sharma et al. 2015), Multiverruca (Wang et al. 2023), Polyschema (Crous et al. 2015a), Pseudoasteromassaria (Ariyawansa et al. 2015), Triseptata (Boonmee et al. 2020), and Verrucohypha (Crous et al. 2024), have been accommodated within the family. Genera within Latoruaceae are mainly distributed in North America (Ellis 1976; Shearer 1982), South America (Crous et al. 2015a; Crous et al. 2024), Africa (Crous et al. 2015a), East Asia (Ariyawansa et al. 2015; Wang et al. 2023), South Asia (Ellis 1976; Boonmee et al. 2020), and Australia (Matsushima 1989; Tan and Bishop-Hurley 2024). They are predominantly saprobic, associated with plant leaves (Ariyawansa et al. 2015), roots (Crous et al. 2024; Crous et al. 2025), soil (Wang et al. 2023), and decaying wood (Mota et al. 2008). Furthermore, some species in Verrucohypha and Polyschema are reported to be endophytic and associated with roots (Crous et al. 2024; Crous et al. 2025), while Polyschema sclerotigenum and Pseudoasteromassaria fagi were reported as a human pathogenic fungi (Crous et al. 2015b) and a plant pathogenic fungus, respectively (Ariyawansa et al. 2015).
Several genera within Latoruaceae are considered monotypic. The genus Latorua became monotypic following the recombination of L. grootfonteinensis into Bahusandhika (Crane and Miller 2016). Pseudoasteromassaria was incorporated into Latoruaceae by Ariyawansa et al. (2015) to accommodate the type species, Ps. fagi, associated with the twigs of Fagus crenata (Fagaceae) in Japan, based on morphological observations and combined SSU and LSU phylogenetic analyses. Subsequently, two freshwater species from Thailand, Ps. aquatica (Dong et al. 2020) and Ps. spadicea (Tibpromma et al. 2017), were accommodated in this genus. Matsushimamyces was established to accommodate keratinophilic fungi from soil in central India. Matsushimamyces bohaniensis (the type species) and M. venustus (≡ Polyschema venustum) were isolated from decaying leaves of an unidentified tree in Cuba (Sharma et al. 2015). Multiverruca was introduced by Wang et al. (2023) as a monotypic genus to accommodate a thermotolerant fungus isolated from soil in Zhejiang, China. Based on a combined LSU, ITS, and SSU phylogenetic analysis, Triseptata was introduced to accommodate a single species, T. sexualis, isolated from dead branches of an unidentified plant in Thailand (Boonmee et al. 2020). Later, Tan and Bishop-Hurley (2024) included T. podargi-strigoidis [as “podargusstrigoides”], isolated from decayed wood of Acacia in Queensland, Australia. Verrucohypha was assigned to Latoruaceae based on an ITS phylogenetic analysis, forming a clade closely related to Polyschema, and is the most recently described genus in this family (Crous et al. 2024). The genus was introduced as monotypic to accommodate the dark septate endophyte, V. endophytica (type species), isolated from roots of Acrocomia aculeata (Arecaceae) in Brazil (Crous et al. 2024).
Polyschema is a species-rich genus in Latoruaceae compared to other genera in this family. The genus was introduced by Upadhyay (1966) with P. terricola as its type species. The teleomorph of species within Polyschema remains undiscovered. However, the hyphomycetous asexual morphological characteristics were described as micronematous or semi-macronematous, mononematous conidiophores, mono- or polytretic, smooth or echinulate, determinate, discrete or integrated conidiogenous cells, bearing ellipsoidal, cylindrical, clavate, or obclavate, septate, pigmented, smooth to verrucose or echinulate conidia (Ellis 1976; Hongsanan et al. 2020). Species of Polyschema lack comprehensive modern taxonomic treatment. Based primarily on morphological features, species within the Polyschema were divided into two distinct groups on the basis of their conidial surfaces by Rao and Reddy (1984), and with further refinement by Mota et al. (2008). In accordance with Rao and Reddy (1984) and Mota et al. (2008), the first group (group I) comprises taxa with verrucose or echinulate conidia, including P. chambalense (Joshi et al. 1983), P. congolense (Reisinger and Kiffer 1974), P. indicum, P. larviforme, P. olivaceum (Ellis 1976), P. queenslandicum (Matsushima 1989), P. sagari (Pandeya and Saksena 1978), P. terricola (type species; Ellis 1976), P. variabile (Tiwari et al. 1977), and P. yakuense (Matsushima 1975). The second group (group II) comprises taxa with smooth-walled conidia, including P. bicellulare (Shearer 1982), P. clavulatum (Ellis 1976), P. cubense (Mercado-Sierra and Mena-Portales 1992), P. lignicola (Rao and Reddy 1984), P. nigroseptatum (Mota et al. 2008), and P. obclaviforme (Ruiz et al. 2000). Polyschema venustum was reassigned to the genus Matsushimamyces according to a combined morphological and ITS-LSU phylogenetic analysis (Sharma et al. 2015).
Previously, the phylogenetic position and relationships of Polyschema within Pleosporales remained unclear. Most species placed in this genus were based on morphological characteristics and were introduced in the 19^th^ century. There are 22 species accepted as members of Polyschema (Index Fungorum 2025; accessed 04 November 2025). Furthermore, 18 species of Polyschema lack molecular data to clarify their phylogenetic placement. Shenoy et al. (2010) revealed the preliminary phylogenetic affinity of Polyschema in Pleosporales based on phylogenetic analyses of LSU sequence data, which placed the genus as a monophyletic lineage related to Lentithecium and Leptosphaeria in Pleosporales. Presently, molecular data are available for only five species of Polyschema, viz. P. congolense, P. larviforme, P. sclerotigenum, P. terricola (Hongsanan et al. 2020), and P. endophytica (Crous et al. 2025). Additionally, most available phylogenetic markers in the above five taxa used to identify species within Polyschema include only ITS and LSU data.
Species within Polyschema typically appear saprobic in various habitats, including forest soil (Crous et al. 2015b), grassland soil, decaying wood (Ellis 1976), leaves (Ruiz et al. 2000), and old firewood (Ellis 1976). In rare cases, they have been found to be pathogenic in human clinical tissue (Crous et al. 2015b) or endophytic in healthy roots of coffee trees (Crous et al. 2025). These species are mainly distributed across North America (Ellis 1976; Shearer 1982; Mota et al. 2008; Crous et al. 2015b), South America (Ellis 1976; Mercado-Sierra and Mena-Portales 1992; Ruiz et al. 2000; Castañeda Ruíz et al. 2009; Crous et al. 2025), East Asia (Matsushima 1975, 1980), and South Asia (Ellis 1976; Tiwari et al. 1977; Pandeya and Saksena 1978; Joshi et al. 1983). As individual cases, P. queenslandicum has been reported exclusively from Australia (Matsushima 1989), and P. congolense from Africa (Ellis 1976).
While conducting fieldwork to collect endophytic fungi associated with higher plants in Yunnan Province, China, five Polyschema strains were isolated from healthy roots of the baobab tree (Adansonia digitata). Through morphological comparisons coupled with multi-locus phylogenetic analyses based on a combined dataset of ITS, LSU, SSU, and rpb2, three novel species in Polyschema (Latoruaceae) from Yunnan are introduced and illustrated in the present study. Additionally, the ecological information and genera of the family Latoruaceae are updated.
Materials and methods
Sample collection, fungal isolation, examination, and preservation
Fresh, healthy roots of baobab trees from Honghe County (altitude 443.2–510.3 m), Honghe Prefecture, Yunnan Province, China, were collected on 25 June 2024. Samples were enclosed in sealed plastic Ziploc bags, stored in cooling boxes, and transported to the laboratory within 24 hours, accompanied by labels containing collection details. Endophytes were isolated from root samples following the protocols outlined by Mattoo and Nonzom (2022), Suwannarach et al. (2023), and Luo et al. (2024). Each sample was cut into 1 × 1 cm^2^ pieces after being thoroughly cleaned with running tap water. Root fragments were surface sterilized using 75% ethanol for 4 min, 5% sodium hypochlorite (NaOCl) for 3 min, and 80% ethanol for 30 s, followed by three rinses with sterilized water and drying with sterile paper tissues. Root pieces were then transferred onto potato dextrose agar (PDA) plates with 0.03% rose Bengal and incubated at 25–30 °C in the dark for 2–5 days. Mycelia emerging from the cut end of root pieces were picked using sterilized needles, purified, and subcultured onto new PDA plates when individual hyphal tips grew out from the root pieces. After purification, the cultures were incubated at 25–30 °C for 1–2 months under dark conditions for sporulation and morphological examination (Mattoo and Nonzom 2022). Culture characteristics, growth, and sporulation on the medium were observed and recorded. Pure fungal cultures are preserved on PDA slants for short-term storage and in 20% glycerol at −20 °C for long-term preservation. Dry culture materials were deposited in the herbarium of the Cryptogams, Kunming Institute of Botany, Academia Sinica (KUN–HKAS). Living cultures were conserved at the Kunming Institute of Botany Culture Collection (KUNCC). New fungal taxa were registered in the MycoBank database (MycoBank 2025).
Microscopic examination
Micromorphological features were examined and captured using a Nikon ECLIPSE Ni-U compound microscope equipped with a Nikon DS-Ri2 camera. All morphological characteristics (e.g., conidiophore, conidiogenous cell, conidia, and chlamydospores) were measured using the Tarosoft® Image Framework version 0.9.7. Photographic plates were edited and combined in Adobe Photoshop CS6 (Adobe Systems Inc., United States) to present the size and morphology of the above-mentioned micromorphological features.
DNA extraction, PCR amplification, and sequencing
Fungal genomic DNA was extracted from fresh mycelia grown on PDA for 5 days at 25 °C using the Biospin Fungus Genomic DNA Extraction Kit (BioFlux®, Hangzhou, China) according to the manufacturer’s instructions. The internal transcribed spacer (ITS: ITS1-5.8S-ITS2), 28S large subunit rDNA (LSU), 18S small subunit rDNA (SSU), and RNA polymerase II second largest subunit (rpb2) loci were amplified using the primer pairs ITS5/ITS4 (White et al. 1990), LR0R/LR5 (Vilgalys and Hester 1990), NS1/NS4 (White et al. 1990), and fRPB2-5F/fRPB2-7cR (Liu et al. 1999), respectively. The PCR reaction mixture was performed in a total volume of 25 µL, containing 2 µL of DNA template, 1 µL of each forward and reverse primer (10 µM), 12.5 µL of 2 × Power Taq PCR Master Mix (mixture of EasyTaq™ DNA Polymerase, dNTPs, and optimized buffer; Beijing Bio Teke Corporation, China), and 8.5 µL of double-distilled water (ddH_2_O). The thermal cycling conditions for PCR amplification of ITS, LSU, SSU, and rpb2 followed those described by Xu et al. (2022). PCR products were sent to TsingKe Biological Technology (Beijing) Co., Ltd., China, for purification and sequencing. All newly generated consensus sequences were deposited in the GenBank database, and the corresponding accession numbers are presented in Table 1.
Sequence alignment and phylogenetic analyses
Sequence data for all loci were subjected to a BLAST search to identify sequences closely related to the five new fungal strains (KUNCC25-20156, KUNCC25-20157, KUNCC25-20158, KUNCC25-20159, and KUNCC25-20160) in the NCBI database (https://blast.ncbi.nlm.nih.gov/Blast.cgi; accessed 25 September 2025). Reference sequences from relevant publications and BLAST results of closely related species were downloaded from GenBank to supplement the datasets (Table 1). To determine the accurate phylogenetic placement of the five new strains, a concatenated ITS–LSU–SSU–rpb2 phylogenetic tree was analyzed using maximum likelihood (ML) and Bayesian inference (BI) methods. Initially, individual DNA sequence matrices were aligned using the online platform MAFFT v. 7.511 (Katoh et al. 2019). The resulting alignments were then trimmed and refined as necessary using BioEdit v. 6.0.7 (Hall 1999). Missing nucleotide bases at the start and end of the consensus sequences were trimmed. ML analysis was performed using the CIPRES Science Gateway v. 3.3 platform (Miller et al. 2010), employing the RAxML-HPC v.8 on XSEDE (version 8.2.12) software. Analyses utilized default parameters except for the implementation of the GAMMA nucleotide substitution model and the execution of 1,000 rapid bootstrap replicates (MLBS). Individual locus datasets were first analyzed by ML to assess the congruence of tree topologies, after which the concatenated ITS–LSU–SSU–rpb2 sequence matrix was analyzed.
For BI analysis, the optimal nucleotide substitution model was independently determined for each locus using MrModeltest 2.3 (Nylander 2008). Based on the Akaike Information Criterion (AIC), the GTR+I+G model was selected as the best-fitting model for ITS, LSU, SSU, and rpb2 loci. BI analyses were conducted via the online portal CIPRES Science Gateway v. 3.3 (Miller et al. 2010) using MrBayes v. 3.2.7 (Ronquist and Huelsenbeck 2003), with Markov chain Monte Carlo (MCMC) sampling to estimate posterior probabilities (PP). Specifically, six concurrent Markov chains were run for 2,000,000 generations, with trees sampled every 100^th^ generation. A burn-in of 25% was applied, and the analysis was automatically terminated once the mean standard deviation of split frequencies fell below 0.01, following the criteria outlined by Maharachchikumbura et al. (2015).
The tree topologies generated in this study were visualized using FigTree v. 1.4.0 (Rambaut 2016). The phylogram was edited and formatted using Microsoft Office PowerPoint 2016 (Microsoft Inc., Redmond, WA, USA) and converted to a TIFF file with Adobe Photoshop CS6 (Adobe Systems Inc., San Jose, CA, USA). The final phylogram was submitted to Figshare (doi: 10.6084/m9.figshare.30716888; https://www.figshare.com; accessed 26 November 2025).
Results
Phylogenetic analyses
The concatenated ITS–LSU–SSU–rpb2 sequence dataset comprises 26 strains of representative species within Latoruaceae and four strains of representative species within Lentimurisporaceae, with Falciformispora senegalensis (CBS 196.79) and F. tompkinsii (CBS 200.79) as the outgroup taxa. The dataset consists of 5,368 total characters, including gaps (ITS: 1–1089 bp, LSU: 1090–2851 bp, SSU: 2852–4260 bp, rpb2: 4261–5368 bp). The final optimized likelihood score for the ML analysis was −19,187.116587. All free model parameters were estimated by the RAxML GAMMA model of rate heterogeneity, with 1,096 distinct alignment patterns and 68.51% undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.248430, C = 0.233171, G = 0.273844, T = 0.244555, with substitution rates AC = 1.489123, AG = 2.533300, AT = 1.245085, CG = 0.740529, CT = 6.110617, GT = 1.000000. The gamma distribution shape parameter alpha = 0.160265, and the tree length = 1.397439. The final average standard deviation of split frequencies at the end of the total MCMC generations was calculated as 0.008629 in the BI analysis.
Tree topologies generated based on ML and BI analyses were similar in the present study. The phylogenetic tree, constructed through analyses of the combined ITS, LSU, SSU, and rpb2 sequence dataset, demonstrated the relationships among taxa within Latoruaceae. Therefore, the phylogenetic tree obtained from the BI analysis was selected and is presented in Fig. 1. These findings are consistent with phylogenetic trees from previous studies (Crous et al. 2024; Crous et al. 2025). The present phylogenetic analyses revealed that seven genera, viz. Latorua, Matsushimamyces, Multiverruca, Polyschema, Pseudoasteromassaria, Triseptata, and Verrucohypha, were placed in Latoruaceae. In the phylogenetic tree, two new strains, KUNCC25-20156 and KUNCC25-20157, formed a robust subclade and have a close relationship with P. congolense (CBS 542.73) and P. hongheense (KUNCC 25-20158) (58% MLBS and 0.99 BYPP; Fig. 1). The new strain KUNCC25-20158 formed a separate branch and is sister to P. congolense (CBS 542.73) with strong statistical support (99% MLBS and 1.00 BYPP; Fig. 1). In addition, the other two new strains (KUNCC25-20159 and KUNCC25-20160) formed a robust subclade basal to P. adansoniae-digitatae, P. congolense, and P. hongheense with 54% MLBS support in the ML analysis but low support in the BI analysis (Fig. 1). Notably, Pseudoasteromassaria spadicea (MFLUCC 15-0972) did not cluster with Ps. aquatica (MFLUCC 18-1397) and Ps. fagi (HHUF 30471, HHUF 30472) in Pseudoasteromassaria, concurring with previous studies (Crous et al. 2024; Crous et al. 2025). The species formed a distinct branch clustering with Triseptata sexualis (MFLUCC 11-0002) and T. podargi-strigoidis (BRIP 76063a) with high statistical support (94% MLBS and 1.00 BYPP; Fig. 1) in the present study. Furthermore, Bahusandhika grootfonteinensis (≡ Latorua grootfonteinensis) formed a strongly supported subclade with Latorua caligans (95% MLBS and 1.00 BYPP; Fig. 1) in Latorua, concurring with previous studies (Crous et al. 2015a; Wang et al. 2023; Crous et al. 2024; Crous et al. 2025). The taxonomic revision of Pseudoasteromassaria spadicea and Bahusandhika grootfonteinensis is formally needed to be reclassified pending further study.
Bayesian inference tree generated by MrBayes v. 3.2.7 on XSEDE in the CIPRES Science Gateway based on a concatenated ITS, LSU, SSU, and rpb2 sequence dataset. The tree is rooted to Falciformispora senegalensis (CBS 196.79) and F. tompkinsii (CBS 200.79). Maximum likelihood bootstrap values (MLBS) greater than 50% and Bayesian posterior probabilities (BYPP) greater than 0.95 are written above the nodes as MLBS/BYPP. Ex-type strains are labeled with the superscript “T,” and newly generated sequences are in red bold.
Taxonomy
Polyschema
adansoniae-digitatae
Taxon classificationFungiPleosporalesLatoruaceae
F.Q. Sun, Kumla, Phookamsak & Suwannar. sp. nov.
20DF5763-FDFF-5BCE-9491-A56F310B1B6E
MB861189
Etymology.
The species epithet “adansoniae-digitatae” refers to the host, Adansonia digitata, from which the fungus was isolated.
Polyschema adansoniae-digitatae (KUN-HKAS 150832, holotype). A, B. Colony on PDA from above and reverse; C. Hyphae; D, E. Conidiogenous cells with conidia; F, G. Conidia growing directly on hyphae; H, I. Chlamydospores; J–P. Conidia. Scale bars: 10 mm (A, B); 50 µm (C); 30 µm (H, I, K); 20 µm (D–G); 15 µm (J); 10 µm (L–P).
Type.
China • Yunnan Province: Honghe Prefecture, Honghe County, isolated from healthy root of baobab tree, 23°15'18"N, 102°14'12"E, elevation 459.8 m, 25 June 2024, F.Q. Sun, BR11-5, dried herbarium culture KUN-HKAS 150832 (Holotype, preserved in a metabolically inactive state), ex-type living culture KUNCC25-20156.
Description.
Dark-brown, filamentous, septate, endophyte isolated from healthy root of baobab tree (Adansonia digitata). Mycelia 1.7–2.2 µm wide, immersed to superficial, composed of branched, septate, initially hyaline to pale brown, smooth hyphae, later becoming brown to dark brown, finely verruculose or warty-surfaced hyphae. Conidiophores semi-macronematous or micronematous, hyaline, aseptate, or reduced to conidiogenous cells. Conidiogenous cells 4.0–6.9 × 2.8–5.6 µm (x̄ = 5.6 × 4.2 μm, n = 30), mono- or polytretic, solitary, discrete, globose or clavate, lateral, rarely terminal, pale brown, smooth-walled, with inconspicuous 1–2 conidiogenous loci. Conidia 18.5–27.6 × 9.7–14.3 µm (x̄ = 23.4 × 12.0 μm, n = 50), solitary to aggregated, acrogenous or catenate, in short branched chains, sometimes born directly on hyphae, ellipsoidal to obovoid, or oblong, pale brown, smooth-walled when young, becoming brown to dark brown at maturity, versicolored, paler brown at apical and basal cells, darker brown in the middle cells, (1–)2–3-septate, slightly constricted at septa, with thick and dark-brown, pigmented, echinulate or spikey warty surface; secondary conidia arising via apical cells. Chlamydospores 11.2–18.7 × 9.1–12.5 µm (x̄ = 15.6 × 11.0 μm, n = 50), terminal or intercalary, appear in short chains, varied in shape, usually globose to subglobose, or ovoid, dark brown, 0–1-septate, thick, smooth-walled.
Culture characteristics.
Colony grows slowly on PDA, reaching 12 mm in diameter after seven days at room temperature (25–30 °C) under dark conditions, floccose, raised, greyish green, white at margin when immature, becoming greyish green to almost blackish olive when mature, producing yellowish-brown pigment.
Additional material examined.
China • Yunnan Province: Honghe Prefecture, Honghe County, isolated from root of baobab tree, 23°15'18"N, 102°14'12"E, elevation 459.8 m, 25 June 2024, F.Q. Sun, BR11-5-1, ex-paratype living cultureKUNCC 25-20157.
Notes.
The present phylogeny demonstrated that the new strains KUNCC 25-20156 (ex-holotype) and KUNCC 25-20157 (ex-paratype) are phylogenetically situated within the genus Polyschema (Fig. 1). The newly identified species, P. adansoniae-digitatae, has a close relationship with the clade including P. congolense and P. hongheense. Base pair comparisons in the ITS, LSU, and rpb2 loci between P. adansoniae-digitatae KUNCC25-20156 (holotype) and P. congolenseCBS 542.73 (holotype) showed 10.1% nucleotide differences (58/575 bp, including 27 gaps) in ITS, 2.3% nucleotide differences (19/840 bp, including four gaps) in LSU, and 6.8% nucleotide differences (52/761 bp, no gaps) in rpb2. Due to the unavailability of the SSU sequence for P. congolense, comparative analyses for this locus could not be conducted. Polyschema adansoniae-digitatae KUNCC25-20156 and P. hongheenseKUNCC 25-20158 showed nucleotide differences of 9.6% (52/544 bp, including 22 gaps) in ITS, 2.3% (19/834 bp, including four gaps) in LSU, 2.8% (38/1344 bp, including 18 gaps) in SSU, and 6.7% (70/1042 bp, including one gap) in rpb2, respectively.
Accompanied by phylogenetic evidence, morphological characteristics provide clear distinctions among P. adansoniae-digitatae, P. congolense, and P. hongheense, particularly in terms of conidial features. The conidia of P. adansoniae-digitatae are 2–3-septate, generally larger, and exhibit more variable shapes, ranging from ellipsoidal to obovoid or oblong, whereas those of P. hongheense are 1–2-septate (sometimes reaching up to four septa), more uniformly sized, and generally narrower (Table 2). Furthermore, conidiogenous cells of P. adansoniae-digitatae are significantly smaller than those of P. congolense and P. hongheense (Table 2).
Polyschema
hongheense
Taxon classificationFungiPleosporalesLatoruaceae
F.Q. Sun, Kumla, Phookamsak & Suwannar. sp. nov.
F3CCFE63-F074-57B3-9B17-4E6FC4F3B35D
MB861247
Etymology.
The species epithet “hongheense” refers to the locality, Honghe County, Yunnan, China, from where the fungus was collected.
Polyschema hongheense (KUN-HKAS 150833, holotype). A, B. Culture from above and reverse; C, D. Hyphae; E–H. Conidiogenous cells; I. Conidia borne parallel to the axis of the conidiogenous cell; J. Conidia borne obliquely to the axis of the conidiogenous cell; K. Conidia borne on the axis of the conidiogenous cell but obliquely oriented; L. Conidia growing directly on hyphae; M, N. Conidiophores; O. Conidia in mass; P–S. Conidia. Scale bars: 10 mm (A, B); 20 µm (C, D, M–O); 10 µm (I–L, P–S); 5 µm (E–H).
Type.
China • Yunnan Province: Honghe Prefecture, Honghe County, isolated from healthy root of baobab tree, 23°15'18"N, 102°14'12"E, elevation 459.8 m, 25 June 2024, F.Q. Sun, BR11-10, dried herbarium culture KUN-HKAS 150833 (Holotype, preserved in a metabolically inactive state), ex-holotype living culture KUNCC25-20158.
Description.
Dark brown, filamentous, septate, endophyte isolated from healthy root of baobab tree (Adansonia digitata). Mycelia 2.2–4.3 µm wide, superficial, composed of branched, narrow, septate, hyaline to light brown hyphae, smooth-walled when young, coarsely roughened, minutely echinulate or verruculose when mature. Conidiophores 2.6–4.6 × 3.3–6.6 µm (x̄ = 3.7 × 5.0 μm, n = 50), macronematous or micronematous, solitary, erect or flexuous, hyaline to pale brown, unbranched, usually short and subglobose, sometimes elongated and curved, septate, 2.4–2.5 × 11.2–38.5 µm, or reduced to conidiogenous cells. Conidiogenous cells 4.6–6.9 × 4.0–8.9 µm (x̄ = 5.8 × 6.5 μm, n = 30), mono- or polytretic, solitary, globose or clavate, hyaline, smooth-walled. Conidia solitary, borne parallelly or obliquely to main axis of the conidiogenous cells, sometimes borne on the axis of conidiogenous cells, but obliquely oriented, sometimes arising directly from hyphae, fusoid-ellipsoidal, slightly constricted at septa, hyaline when young, brown when mature, paler at end cells, dark brown to black at the centre, verrucose to echinulate at surface, (1–4)-septate, usually (1–2)-septate; 1-septate measuring 12.8–19.8 × 7.4–12.7 µm (x̄ = 16.5 × 10.2 μm, n = 50), 2-septate 16.5–22.4 × 9.5–13.6 µm (x̄ = 19.6 × 11.6 μm, n = 50), 3-septate 20.0–32.7 × 8.0–11.8 µm (x̄ = 26.5 × 10.0 μm, n = 50), 4-septate 34.5–36.6 × 10.1–12.6 µm (x̄ = 35.6 × 11.4 μm, n = 50).
Culture characteristics.
Colony grows slowly on PDA, reaching 14 mm in seven days at room temperature (25–30 °C) under dark conditions, colonies irregular, fluffy asteroid, raised, brown, white at the margin when immature, becoming completely brown to olive brown when mature.
Notes.
Polyschema hongheense (KUNCC25-20158) formed a distinct lineage sister to P. congolense, supported by significant statistical values (99% MLBS and 1.00 BYPP; Fig. 1), which indicates a close phylogenetic relationship. Base pair comparisons in the ITS, LSU, and rpb2 loci between P. hongheense KUNCC25-20158 (holotype) and P. congolenseCBS 542.73 (holotype) showed 3.3% nucleotide differences (17/521 bp, including five gaps) in ITS, 0.7% nucleotide differences (6/845 bp, no gaps) in LSU, and 5.9% nucleotide differences (45/760 bp, including one gap) in rpb2. Due to the absence of SSU sequences for P. congolense, comparisons at this locus were not available.
Morphologically, P. hongheense can be distinguished from P. congolense by its wider mycelium (2.2–4.3 µm wide vs. 1.5–2.5 μm wide) and conidia that are larger (12.8–36.6 × 7.4–13.6 µm vs. 16–26 × 8–11 μm) and reach up to four septa, whereas P. congolense has 2–3-septate, commonly with two septa (Table 2).
Polyschema
radicicola
Taxon classificationFungiPleosporalesLatoruaceae
F.Q. Sun, Kumla, Phookamsak & Suwannar. sp. nov.
942E2874-E09E-5BD3-90A2-297CD53CEC5B
MB861248
Etymology.
The species epithet “radicicola” refers to the root from which the species was isolated.
Polyschema radicicola (KUN-HKAS 150834, holotype). A, B. Culture from above and reverse; C, D. Hyphae; E–H. Conidiogenous cells; I. Conidia growing terminally; J, K. Conidiophores; L–N. Conidia in mass; O–R. Conidia. Scale bars: 10 mm (A, B); 50 µm (M, N); 20 µm (C); 15 µm (L); 10 µm (D, G–K, O–R); 5 µm (E, F).
Type.
China • Yunnan Province: Honghe Prefecture, Honghe County, isolated from healthy root of baobab tree, 23°25'21"N, 102°14'19"E, elevation 503.7 m, 25 June 2024, F.Q. Sun, BR16-1, dried herbarium culture KUN-HKAS 150834 (Holotype, preserved in a metabolically inactive state), ex-type living culture KUNCC25-20159.
Description.
Dark brown, filamentous, septate endophyte (DSE) isolated from healthy root of baobab tree (Adansonia digitata). Mycelium 1.7–4.0 µm wide, superficial, composed of branched, narrow, septate, hyaline to light brown hyphae, flexuous hyphae, smooth-walled when young, straight or curved, coarsely roughened when mature, minutely echinulate or verruculose. Conidiophores 7.2–10.3 × 3.4–6.1 µm (x̄ = 8.8 × 4.8 μm, n = 50), micronematous, solitary, erect, hyaline to pale brown, unbranched, clavate, or short cylindrical, or reduced to conidiogenous cells. Conidiogenous cells 4.4–7.2 × 4.3–7.3 µm (x̄ = 5.8 × 5.9 μm, n = 30), monotretic, solitary, terminal or lateral, globose, clavate or short cylindrical, hyaline, smooth-walled. Conidia solitary, mostly pleurogenous, at times acrogenous, usually borne along the axis of conidiogenous cells, but sometimes obliquely oriented, cylindrical, slightly constricted at septa, pale brown, end cells paler, dark brown to black at the septa, verrucose to echinulate surfaced, (2–5)-septate, 20.4–40.9 × 7.5–13.1 µm (x̄ = 31.0 × 10.5 μm, n = 50).
Culture characteristics.
Colony grows slowly on PDA, reaching 15 mm in seven days at room temperature (25–30 °C) under dark conditions, colonies irregular, felted, slightly heaped and folded, pale brown when immature, becoming completely brown when mature, producing orange pigment.
Additional materials examined.
China • Yunnan Province: Honghe Prefecture, Honghe County, isolated from healthy root of baobab tree, 23°25'21"N, 102°14'19"E, elevation 503.7 m, 25 June 2024, F.Q. Sun, BR16-5, ex-paratype living cultureKUNCC 25-20160.
Notes.
Polyschema radicicola is basal to a subclade containing P. adansoniae-digitatae (KUNCC25-20156 and KUNCC25-20157), P. congolense (CBS 542.73), and P. hongheense (KUNCC 25-20158). These species form a subclade and have a close relationship with P. endophytica (COAD3972) and P. terricola (CBS 301.65). Based on nucleotide comparisons, P. radicicola KUNCC25-20159 (holotype) differs from P. congolenseCBS 542.73 (holotype) in the ITS sequence by 34/548 bp (6.2%, including 14 gaps), in the LSU sequence by 14/846 bp (1.7%, including one gap), and in the rpb2 sequence by 82/768 bp (10.7%, including three gaps). Polyschema radicicola KUNCC25-20159 differs from P. hongheense KUNCC25-20158 by 5.8% nucleotides in ITS (30/518 bp, including nine gaps), 1.4% nucleotides in LSU (12/844 bp, including one gap), 2.3% nucleotides in SSU (31/1336 bp, including nine gaps), and 8.9% nucleotides in rpb2 (94/1060 bp, including four gaps). Polyschema radicicola KUNCC25-20159 differs from P. endophytica COAD3972 (holotype) by 9.4% nucleotides in ITS (52/556 bp, including 28 gaps) and 2.0% nucleotides in LSU (17/849 bp, including four gaps), and differs from P. terricolaCBS 301.65 (holotype) by 11.4% nucleotides in ITS (57/501 bp, including 33 gaps), 2.3% nucleotides in LSU (20/852 bp, including seven gaps), and 10.0% nucleotides in rpb2 (77/769 bp, including three gaps).
Morphologically, Polyschema radicicola is distinguishable by its colony producing orange pigment (Table 2). Furthermore, P. radicicola exhibits mycelial hyphae that are occasionally narrow, flexuous, and smooth-walled, yet remain capable of generating conidiogenous cells and conidia. Conidia of P. radicicola differ from those of P. terricola (Ellis 1976) by being longer and narrower (20.4–40.9 × 7.5–13.1 µm vs. 15–35 × 13–18 μm), cylindrical, distinctly echinulate at the surface, and transversely 2–5-septate, lacking vertical septa, whereas P. terricola has transversely 0–3-septate conidia with sometimes one longitudinal or oblique septum (Ellis 1976). Polyschema congolense (Reisinger and Kiffer 1974) can be distinguished from P. radicicola by having smaller (16–26 × 8–11 μm) and 2–3-septate conidia, commonly with two septa. Polyschema hongheense differs by having shorter (12.8–36.6 × 7.4–13.6 µm), paler brown, 1–4-septate conidia, commonly with 1–2 septa and occasionally formed terminally. Polyschema endophytica (Crous et al. 2025) differs by having aseptate, globose, subglobose to ellipsoidal, smooth-walled conidia.
Discussion
Species of Polyschema were divided into two groups by Rao and Reddy (1984) based on their conidial surfaces and were further divided by conidial septa, shapes, colors, and sizes. This grouping was later updated by Mota et al. (2008). Group I has conidia with ornamented walls, whereas Group II has smooth-walled conidia. Based on their conidial surfaces, the three new Polyschema species, P. adansoniae-digitatae, P. hongheense, and P. radicicola, obtained in this study were classified into Group I (ornamented-walled conidia). A morphological comparison between our new species and other Polyschema species is provided in Table 2. However, morphological classification in Polyschema remains limited by ambiguous descriptions; therefore, integrating morphological data with molecular phylogenetic analysis provides a more robust and reliable framework for identifying and confirming new species. By adding our newly introduced species, there are a total of 24 species accommodated in Polyschema (Tables 2, 3). However, only eight species are confirmed with molecular evidence (Table 2). Furthermore, most sequences in these eight species belong to ITS and LSU; therefore, previous phylogenies including Polyschema (Crous et al. 2024; Crous et al. 2025) relied mainly on ribosomal DNA regions (ITS and LSU). The present study employs a combined analysis of ITS, LSU, SSU, and rpb2 regions, which is mostly consistent with earlier approaches but provides clearer phylogenetic relationships among closely related species with stronger statistical support. The phylogenetic reconstruction reveals a progressive, stepwise pattern of separation within species of Polyschema. Through nucleotide comparisons in the present study while introducing P. adansoniae-digitatae, P. hongheense, and P. radicicola, these comparisons indicate minor differences in LSU and SSU loci, while there are significant base pair differences in ITS and rpb2. Therefore, it suggests that ITS and rpb2 sequences are efficient for resolving the ambiguous placement of Polyschema species in the future.
Members of the family Latoruaceae play various ecological roles, including endophytic, pathogenic, and saprobic life modes (Table 3). Most Polyschema species have been documented as saprobes on various hosts, including soil, decaying wood, and leaves worldwide (Ariyawansa et al. 2015; Crous et al. 2015b; Wang et al. 2023; Table 3). More recent findings, including P. sclerotigenum from human clinical material (Crous et al. 2015b) and P. endophytica (Crous et al. 2025) as a root endophyte of coffee, suggest that the genus is not limited to a saprobic lifestyle. The three root endophytes introduced in this study, representing the first records of Polyschema from mainland China, further underscore its ecological plasticity. Furthermore, this is the first report of members of Polyschema living in baobab trees. Other genera within the Latoruaceae are mostly isolated from soil and decaying or decayed wood. The identification of Pseudoasteromassaria aquatica (Dong et al. 2020) and Ps. spadicea (Tibpromma et al. 2017) expanded the known habitats of this family to freshwater environments. Pseudoasteromassaria fagi (Ariyawansa et al. 2015) was reported as pathogenic on decaying wood submerged in freshwater in Thailand, exhibiting a lifestyle similar to Polyschema sclerotigenum. Verrucohypha endophytica (Crous et al. 2024) was discovered as an endophyte, a lifestyle also observed in Polyschema endophytica (Crous et al. 2025) and in the three new taxa introduced in this study. These observations point to a family characterized by ecological plasticity, morphological variability, and an expanding distribution range. Continued sampling from varied habitats will be crucial to uncover hidden knowledge of Polyschema, species diversity in Latoruaceae, and their extended ecological significance.
There is a controversial point in the present phylogeny. Bahusandhika grootfonteinensis is clustered with Latorua and remained distinct from the Bahusandhika group within Lentimurisporaceae, concurring with previous studies (Crous et al. 2015a; Wang et al. 2023; Crous et al. 2024; Crous et al. 2025). Latorua grootfonteinensis was transferred to Bahusandhika by Crane and Miller (2016) due to its similarities to Bahusandhika in conidiogenous cells, conidial development, overall conidial morphology, and lack of molecular data support. However, the present phylogeny and the phylogenies in Wang et al. (2023) and Crous et al. (2024; Crous et al. 2025) do not support this synonym. Therefore, B. grootfonteinensis should be reinstated in Latorua as L. grootfonteinensis. Similarly, Pseudoasteromassaria spadicea (MFLUCC 15-0972) clustered within Triseptata and remained distant from Pseudoasteromassaria. This species should be reclassified within Triseptata based on phylogenetic analyses. However, further work is needed to confirm the taxonomic placement of these two species, particularly by using more taxa sampling and morphological determination.
Supplementary Material
XML Treatment for Polyschema adansoniae-digitatae
XML Treatment for Polyschema hongheense
XML Treatment for Polyschema radicicola
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ariyawansa HA, Hyde KD, Jayasiri SC, Buyck B, Chethana KT, Dai DQ, Dai YC, Daranagama DA, Jayawardena RS, Lücking R, Ghobad-Nejhad M, Niskanen T, Thambugala KM, Voigt K, Zhao RL, Li GJ, Doilom M, Boonmee S, Yang ZL, Cai Q, Cui YY, Bahkali AH, Chen J, Cui BK, Chen JJ, Dayarathne MC, Dissanayake AJ, Ekanayaka AH, Hashimoto A, Hongsanan S, Jones EBG, Larsson E, Li WJ, Li QR, Liu JK, Luo ZL, Maharachchikumbura SSN, Mapook A, Mc Kenzie EHC, Norphanphoun C, Konta S, Pang KL, Perera RH, Phookamsak R, Ph · doi ↗
- 2Boonmee S, Calabon MS, Phookamsak R, Elgorban AM, Hyde KD (2020) Triseptata sexualis gen. et sp. nov. in Latoruaceae (Pleosporales). Phytotaxa 447: 252–264. 10.11646/phytotaxa.447.4.3 · doi ↗
- 3Castañeda Ruíz RF, Iturriaga T, Minter DW, Abarca GH, Stadler M, Saikawa M, Fernández R (2009) Two new anamorphic fungi and some microfungi recorded from ‘El Ávila,’ Venezuela. Mycotaxon 107: 225–237. 10.5248/107.225 · doi ↗
- 4Crane JL, Miller AN (2016) Studies in genera similar to Torula: Bahusaganda, Bahusandhika, Pseudotorula, and Simmonsiella gen. nov. IMA Fungus 7: 29–45. 10.5598/imafungus.2016.07.01.03PMC 494168627433439 · doi ↗ · pubmed ↗
- 5Crous PW, Carris LM, Giraldo A, Groenewald JZ, Hawksworth DL, Hernández-Restrepo M, Jaklitsch WM, Lebrun MH, Schumacher RK, Stielow JB, van der Linde EJ, Vilcāne J, Voglmayr H, Wood AR (2015 a) The genera of fungi—fixing the application of the type species of generic names—G 2: Allantophomopsis, Latorua, Macrodiplodiopsis, Macrohilum, Milospium, Protostegia, Pyricularia, Robillarda, Rotula, Septoriella, Torula, and Wojnowicia. IMA Fungus 6: 163–198. 10.5598/imafungus.2015.06.01.11PMC 4500082262034 · doi ↗ · pubmed ↗
- 6Crous PW, Sutton DA, Acharya K, Barber PA, Boekhout T, Dimitrov RA, Dueñas M, Dutta AK, Gené J, Gouliamova DE, Groenewald M, Lombard L, Morozova OV, Sarkar J, Smith MT, Stchigel AM, Wiederhold NP, Alexandrova AV, Antelmi I, Armengol J, Barnes I, Cano-Lira JF, Castañeda Ruiz RF, Contu M, Courtecuisse PR, da Silveira AL, Decock CA, de Goes A, Edathodu J, Ercole E, Firmino AC, Fourie A, Fournier J, Furtado EL, Geering AD, Gershenzon J, Giraldo A, Gramaje D, Hammerbacher A, He XL, Haryadi D, Khemmuk · doi ↗ · pubmed ↗
- 7Crous PW, Wingfield MJ, Jurjević Ž, Balashov S, Osieck ER, Marin-Felix Y, Luangsa-ard JJ, Mejí LC, Cappelli A, Parra LA, Lucchini G, Chen J, Moreno G, Faraoni M, Zhao RL, Weholt Ø, Borovička J, Jansen GM, Shivas RG, Tan YP, Akulov A, Alfenas AC, Alfenas RF, Altes A, Avchar R, Barreto RW, Catcheside DEA, Chi TY, Esteve-Raventos F, Fryar SC, Hanh LTM, Larsbrink J, Oberlies NH, Olsson L, Pancorbo F, Raja HA, Thanh V, Thuy NT, Ajithkumar K, Akram W, Alvarado P, Angeletti B, Arumugam E, Atashi K, Hal · doi ↗ · pubmed ↗
- 8Crous PW, Catcheside DEA, Catcheside PS, Alfenas AC, Alfenas RF, Barreto RW, Lebel T, Balashov S, Broadbridge J, Jurjević Ž, De la Peña-Lastra S, Hoffmann R, Mateos A, Riebesehl J, Shivas RG, Soliz Santander FF, Tan YP, Altés A, Bandini D, Carriconde F, Cazabonne J, Czachura P, Gryta H, Eyssartier G, Larsson E, Pereira OL, Rigueiro-Rodríguez A, Wingfield MJ, Ahmad W, Bibi S, Denman S, Esteve-Raventós F, Hussain S, Illescas T, Luangsa-Ard JJ, Möller L, Mombert A, Noisripoom W, Olariaga I, Pancorb · doi ↗ · pubmed ↗