Activation of 5-HT1A autoreceptors causes actin depolymerization in Drosophila serotonergic neurons
Ava Kinser, Jessie K. Rhodes, Douglas H. Roossien

TL;DR
This study explores how serotonin receptors affect axon growth in fruit fly neurons, finding that they cause actin structures to break down.
Contribution
The study reveals a novel cellular mechanism linking 5-HT1A autoreceptor activation to actin depolymerization in serotonergic neurons.
Findings
Activation of 5-HT1A increases G/F-actin ratios in Drosophila serotonergic neurons.
5-HT1A negatively regulates axon outgrowth through actin depolymerization.
Abstract
Serotonergic neurons extend long, highly branched axons throughout the brain during development and are responsible for the modulation of many behaviors. Because proper behavioral output is dependent on precise outgrowth and targeting of serotonergic axons, it is important to understand how serotonergic axon outgrowth is regulated during development. Our previous pharmacological experiments suggest that autoreceptor 5-HT1A negatively regulates axon outgrowth and branching of Drosophila serotonergic neurons in vitro , though the cellular mechanisms are unknown. Here we show that pharmacological activation of 5-HT1A leads to increases in G/F-actin ratios, suggesting 5-HT1A negatively regulates serotonergic axon outgrowth through actin depolymerization.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
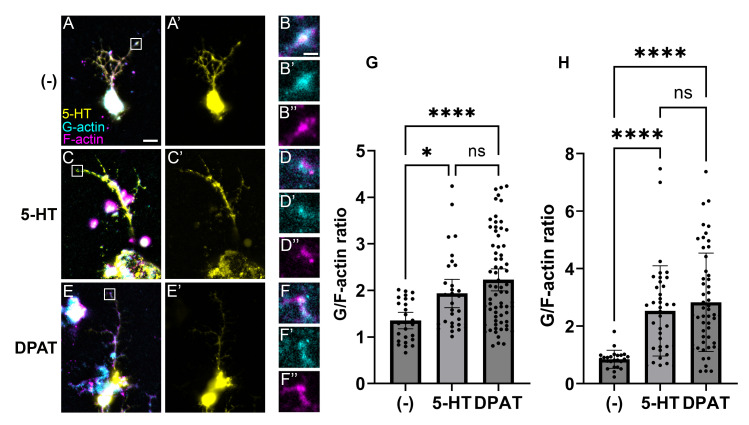 Figure 1
Figure 1- —National Science Foundation (United States)https://ror.org/021nxhr62
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurobiology and Insect Physiology Research · Developmental Biology and Gene Regulation · Neurotransmitter Receptor Influence on Behavior
Description
In addition to its canonical role as a neurotransmitter, serotonin (5-hydroxytryptamine or 5-HT) negatively autoregulates the outgrowth of serotonergic axons (Budnik et al., 1989; Daubert & Condron, 2010; Diefenbach et al., 1995; Migliarini et al., 2013). Previous experiments in primary Drosophila serotonergic neurons showed this occurs through 5-HT1A receptor signaling (Long et al., 2023). Growth cones at the distal tip of actively growing axons contain radially arranged bundles of F-actin, which aid in adhesion, navigation, and elongation of the axon (Miller & Suter, 2018). In non-serotonergic snail growth cones, addition of 5-HT causes F-actin depolymerization (Torreano et al., 2005). We therefore hypothesized that activation of 5-HT1A negatively regulates axon outgrowth in serotonergic neurons by inducing actin depolymerization. Here we tested this hypothesis using a previously established fluorescence method of estimating the ratios of G-actin and F-actin (Torreano et al., 2005) after pharmacological treatment with 5-HT or the 5-HT1A agonist 8-OH-DPAT (DPAT).
Primary serotonergic neurons were harvested from Stage 11 *Drosophila * embryos, the point at which serotonergic neurons are committed to their cell fate and are undergoing active periods of axon outgrowth (Chen & Condron, 2008; Vallés & White, 1988). Neurons were cultured for 24 hours then treated 2 hours with either negative control media, 5-HT, or DPAT. The neurons were then stained with α5-HT, SiR-Actin (F-actin stain), and DNAseI (G-actin stain) to identify serotonergic neurons and estimate F- and G- actin levels by creating ROIs as the most distal 5 μm ends of axons in each channel. The samples were imaged on a Zeiss Apotome, which is capable of both standard epifluorescent imaging and optical sectioning. Using standard epifluorescence images, we found the average G/F actin ratio in controls was 1.35 (n = 25 ROIs measured from 11 neurons), whereas treatment with 5-HT and DPAT increased the average ratios to 2.01 (n = 31 ROIs measured from 22 neurons) and 2.23 (n = 70 ROIs from 51 neurons), respectively. In addition, we created maximum projections from the Z-stacks to eliminate any potential bias due to epifluorescent focusing. We found similar results, with a mean control G/F-actin ratio of 0.834, whereas 5-HT and DPAT treatment increased ratios to 2.56 and 2.823, respectively.
In these experiments, higher ratios correlate with either an increase in G-actin and/or a decrease in F-actin. The interpretation of these higher ratios, therefore, is that 5-HT and DPAT induced actin depolymerization. Previous studies have shown that 5-HT1A activation in serotonergic neurons can modulate ERK1/2 via several mechanisms, including b-arrestin scaffolding (Della Rocca et al., 1999), heterodimerization with FGFRs (Borroto-Escuela et al., 2013, 2015), and alternative G-protein subunit coupling (Albert & Vahid-Ansari, 2019; Kushwaha & Albert, 2005; Rojas & Fiedler, 2016). ERK1/2 signaling has been shown to regulate actin dynamics through phosphorylation of RhoA (Kim et al., 2004; Kumar et al., 2005), highlighting its potential role in 5-HT1A mediated F-actin depolymerization. The increase in G/F-actin ratio observed upon addition of 5-HT or DPAT supports a model in which 5-HT1A signaling induces F-actin depolymerization, possibly through downstream ERK1/2 activation and subsequent modulation of actin regulator proteins such as Rho GTPases. Consistent with our previous work, we found the distribution of our negative control data to be normal, whereas treatment of 5-HT and DPAT causes a shift to a non-normal distribution. We speculate here that this occurs because only a subset of serotonergic neurons expresses 5-HT1A in this system, and therefore only a subpopulation of axons responds. However, this should be tested in future experiments using 5-HT1A expression markers to delineate these subpopulations.
Methods
Primary neuronal culture of serotonergic neurons was performed as described previously (Long et al., 2023). In brief, stage 11 Drosophila embryos were collected and dechorionated in 50% bleach before cells were aspirated and transferred to Schnieder’s Drosophila medium (Fisher 21-720-024) supplemented with 20% FBS (Fisher 16-140-063), 0.02% P/S, and 0.2 mg/mL insulin (Sigma 10515-5ML). The cells were chemically digested with collagenase followed by physical trituration before plating on 0.01% poly-L-lysine treated coverslips. Serotonin hydrochloride (Sigma H9523) was suspended at 10mM in sterile PBS prior to suspending in media at 100 μM final concentration. The 5-HT1A agonist 8-hydroxy-DPAT hydrobromide (Tocris Bioscience 0529) was suspended in DMSO prior to diluting to final concentration of 10 μM in 1 mL of culture media. This resulted in a final percentage of 0.01 DMSO in culture media. Drugs were added to cultures 2 hours prior to fixing. Cells were fixed with 4% paraformaldehyde for 15 min at room temperature, washed three times with PBS for 3 min, then incubated with rabbit anti-serotonin (ImmunoStar 20080) at 1:500 dilution in 1× PBS with 0.1% Triton X-100 for 45 min at room temperature. After washing for 5 min in 1× PBS with 0.1% Triton X-100 three times, samples were incubated in donkey anti-rabbit IgG AMCA (Jackson ImmunoResearch 711-155-152) at 1:500 dilution, 0.4 mg/mL SiR-Actin, (Cytoskeleton CYSC001), and 0.4 mg/mL DNAse I-Alexa488 (Invitrogen D12371) in 1× PBS for 30 min at room temperature. Neurons were washed three times in 1× PBS for 10 min at room temperature before being briefly rinsed in Milli-Q water. Samples were mounted in Vectashield Antifade Mounting Medium (Vector Labs H-1000).
Neurons were imaged on a Zeiss Apotome Axio Imager.M2 with a 63x NA 1.4 objective, X-cite 120LED Lamp, and Zeiss 807 mono camera. AMCA was excited at 353 nm with emission collected at 465 nm. SiR-actin was excited at 650 nm with emission collected at 673nm. DNAse I was excited at 488 nm with emission collected at 509 nm. Serotonergic neurons were identified using fluorescent AMCA channel. In FIJI, the background was subtracted from a composite image of all three channels, with the best focal plane for the actin channels selected for analysis. For each neuron, the distal 5 μm tip of each axon branch was selected as an ROI and used to measure the mean pixel intensity for each actin channel in the selected area. The G/F actin ratio at each distal axon tip was determined by dividing the mean pixel intensity of G-actin by the mean pixel intensity of F-actin (Torreano et al., 2005). In addition, maximum projections of the optical sectioning stacks were created and analyzed as above. All statistical analyses were conducted in GraphPad Prism. Normality was tested using the Shapiro–Wilk test. An analysis of variance was run on the G/F-actin ratios with Kruskal-Walis post hoc comparisons.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Albert PR Vahid-Ansari F 20181025 The 5-HT 1A receptor: Signaling to behavior.Biochimie 1610300-9084344510.1016/j.biochi.2018.10.01531079617 · doi ↗ · pubmed ↗
- 2Borroto-Escuela DO Corrales F Narvaez M Oflijan J Agnati LF Palkovits M Fuxe K 20131021 Dynamic modulation of FGFR 1-5-HT 1A heteroreceptor complexes. Agonist treatment enhances participation of FGFR 1 and 5-HT 1A homodimers and recruitment of β-arrestin 2.Biochem Biophys Res Commun 44120006-291X 38739210.1016/j.bbrc.2013.10.06724157794 · doi ↗ · pubmed ↗
- 3Borroto-Escuela DO Pérez-Alea M Narvaez M Tarakanov AO MudóG Jiménez-Beristain A Agnati LF Ciruela F Belluardo N Fuxe K 201556 Enhancement of the FGFR 1 signaling in the FGFR 1-5-HT 1A heteroreceptor complex in midbrain raphe 5-HT neuron systems. Relevance for neuroplasticity and depression.Biochem Biophys Res Commun 46330006-291X 18018610.1016/j.bbrc.2015.04.13325957476 · doi ↗ · pubmed ↗
- 4Budnik V Wu CF White K 198981 Altered branching of serotonin-containing neurons in Drosophila mutants unable to synthesize serotonin and dopamine.J Neurosci 980270-64742866287710.1523/JNEUROSCI.09-08-02866.19892570137 PMC 6569695 · doi ↗ · pubmed ↗
- 5Chen J Condron BG 200849 Branch architecture of the fly larval abdominal serotonergic neurons.Dev Biol 32010012-1606303810.1016/j.ydbio.2008.03.03818561908 PMC 2610461 · doi ↗ · pubmed ↗
- 6Daubert EA Condron BG 2010618 Serotonin: a regulator of neuronal morphology and circuitry.Trends Neurosci 3390166-223642443410.1016/j.tins.2010.05.00520561690 PMC 2929308 · doi ↗ · pubmed ↗
- 7Della Rocca GJ Mukhin YV Garnovskaya MN Daaka Y Clark GJ Luttrell LM Lefkowitz RJ Raymond JR 1999219 Serotonin 5-HT 1A receptor-mediated Erk activation requires calcium/calmodulin-dependent receptor endocytosis.J Biol Chem 27480021-92584749475310.1074/jbc.274.8.47499988712 · doi ↗ · pubmed ↗
- 8Diefenbach TJ Sloley BD Goldberg JI 199511 Neurite branch development of an identified serotonergic neuron from embryonic Helisoma: evidence for autoregulation by serotonin.Dev Biol 16710012-160628229310.1006/dbio.1995.10237851649 · doi ↗ · pubmed ↗