Genetic and phenotypic characterization of the plasmid-encoded NDM-80 metallo-β-lactamase in Escherichia coli isolated from a pediatric patient
Jinlan Zhou, Qing Meng, Qimeng Fan, Weiwei Yang, Li Ding, Yan Guo, Fupin Hu, Guoping Lu, Gangfeng Yan

TL;DR
A new antibiotic-resistant gene variant, blaNDM–80, was found in E. coli from a child in China, showing resistance to most beta-lactam antibiotics.
Contribution
This is the first report of blaNDM–80, revealing its genetic structure and higher resistance compared to other NDM variants.
Findings
E. coli with blaNDM–80 showed resistance to most beta-lactam antibiotics except aztreonam, aztreonam-avibactam, and cefiderocol.
blaNDM–80 has unique mutations and a plasmid structure that may enhance its resistance and transmission potential.
NDM-80 hydrolyzes certain antibiotics more effectively than NDM-1, contributing to higher resistance.
Abstract
Carbapenem-resistant Enterobacterales (CRE) strains carrying blaNDM variants pose a significant threat to the health of infected patients worldwide. This study isolated a carbapenem-resistant Escherichia coli (E. coli) strain carrying blaNDM–80 from a patient in an intensive care unit in China. Antimicrobial susceptibility testing, whole-genome sequencing (WGS), plasmid transformation assay, cloning experiment, and steady-state kinetic determinations were performed to investigate antimicrobial susceptibility, the characteristics of the genetic environment, the mechanism of resistance gene transmission, resistance gene function, and antibiotic hydrolysis ability. The results indicated that E. coli carrying blaNDM–80 showed significant resistance to all β-lactam antibiotics except for aztreonam, aztreonam-avibactam, and cefiderocol. WGS analysis revealed that the strain belonged to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
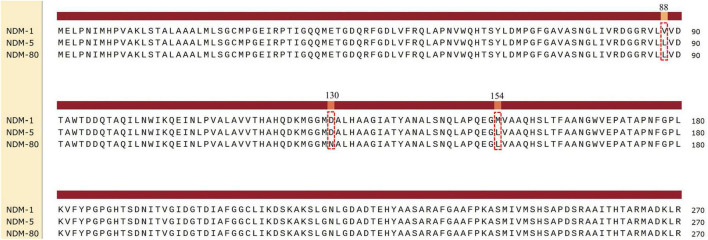 FIGURE 1
FIGURE 1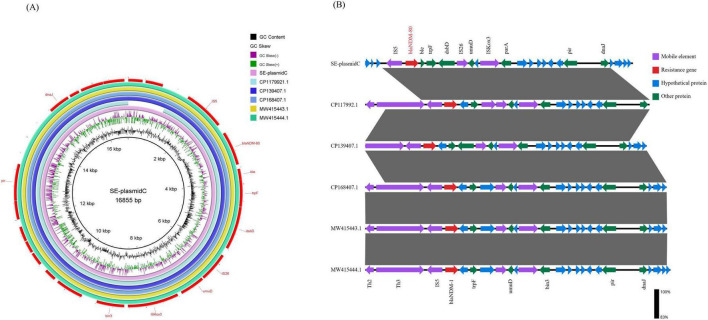 FIGURE 2
FIGURE 2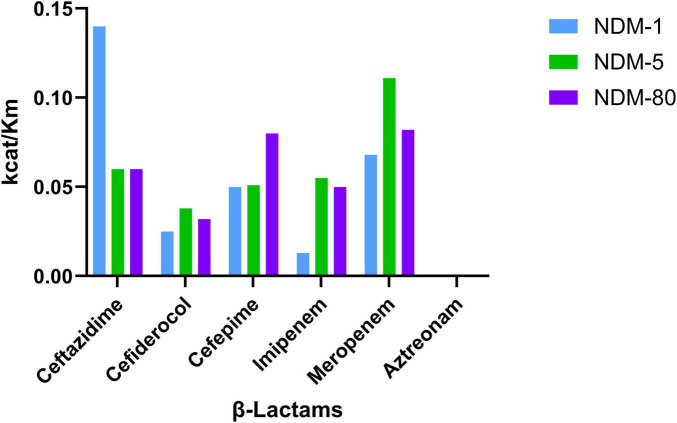 FIGURE 3
FIGURE 3| Isolates no. |
| MIC (mg/L) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IPM | MEM | MEV | CZT | CZA | AZA | ERV | AMK | FEP | CAZ | ||
|
| 8 | 32 | 32 | >128 | >64 | 0.06 | 0.25 | 2 | >32 | >32 | |
| – | 0.125 | <0.03 | <0.03 | 0.5 | 0.06 | <0.03 | 0.125 | 2 | 0.5 | 0.5 | |
|
| 8 | 16 | 16 | >128 | >64 | 0.06 | 0.125 | 2 | 32 | >32 | |
| – | 0.125 | <0.03 | <0.03 | 0.25 | 0.25 | <0.03 | 0.125 | <1 | <0.25 | <0.25 | |
|
| 1 | 0.125 | 0.125 | >128 | 32 | <0.03 | 0.125 | <1 | 1 | >32 | |
|
| 1 | 0.125 | 0.125 | >128 | >64 | <0.03 | 0.125 | <1 | 1 | >32 | |
|
| 0.5 | 0.06 | 0.125 | 128 | 32 | <0.03 | 0.125 | <1 | 0.5 | >32 | |
|
| 64 | 64 | 64 | >128 | >64 | 0.06 | 0.25 | 2 | >32 | >32 | |
|
| 64 | 64 | 64 | >128 | >64 | <0.03 | 0.125 | <1 | >32 | >32 | |
|
| 64 | 64 | 64 | >128 | >64 | <0.03 | <0.06 | <1 | >32 | >32 | |
|
| |||||||||||
|
|
|
|
|
|
|
|
|
|
| ||
| >32 | <1 | >8 | 4 | >128 | >256 | 0.25 | 0.25 | >16 | 1 | ||
| <0.25 | <1 | <0.06 | <0.06 | <1 | 4 | 0.125 | 0.25 | <0.25 | <0.03 | ||
| >32 | <1 | <0.06 | <0.06 | >128 | >256 | 0.125 | 0.25 | <0.25 | 0.125 | ||
| <0.25 | <1 | <0.06 | <0.06 | <1 | 4 | 0.125 | 0.25 | <0.25 | 0.06 | ||
| 16 | <1 | <0.06 | <0.06 | 8 | 16 | 0.125 | 0.25 | <0.25 | 1 | ||
| 16 | <1 | <0.06 | <0.06 | 8 | 32 | 0.125 | 0.25 | <0.25 | 1 | ||
| 16 | <1 | <0.06 | <0.06 | 8 | 16 | 0.125 | 0.25 | <0.25 | 0.5 | ||
| >32 | <1 | <0.06 | <0.06 | >128 | >256 | 0.125 | 0.25 | <0.25 | 1 | ||
| >32 | <1 | <0.06 | <0.06 | >128 | >256 | <0.06 | 0.25 | <0.25 | 1 | ||
| >32 | <1 | <0.06 | <0.06 | >128 | >256 | 0.125 | 0.25 | <0.25 | 0.5 | ||
| Sample name | Genome size (bp) | MLST | Serological typing | Plasmid Inc., type | Antibiotic resistance gene(s) |
|---|---|---|---|---|---|
| Chromosome | 5,057,113 bp | 155 | O4:H51 | NA |
|
| SE-plasmidA | 184,004 bp | NA | NA | IncHI2, RepA, IncHI2A | |
| SE-plasmidB | 138,252 bp | NA | NA | IncFIB/IncFIC(FII) | |
| SE-plasmidC | 17,067 bp | NA | NA | IncX3 |
|
| SE-plasmidD | 4,308 bp | NA | NA | Col156 | NA |
| β -Lactams | NDM-1 | NDM-5 | NDM-80 | ||||||
|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
|
|
|
|
| |
| Ceftazidime | 47.2432 | 6.509 | 0.14 | 177.8154 | 11.3705 | 0.06 | 324.05 | 20.7 | 0.06 |
| Cefiderocol | 148.414 | 3.7265 | 0.025 | 149.6593 | 5.6435 | 0.038 | 478.9576 | 15.485 | 0.032 |
| Cefepime | 150.4413 | 1.51 | 0.050 | 153.496 | 7.8005 | 0.051 | 396.1818 | 32.128 | 0.08 |
| Imipenem | 126.3622 | 1.6305 | 0.013 | 225.5201 | 14.2675 | 0.055 | 698.3814 | 34.9645 | 0.050 |
| Meropenem | 16.7962 | 1.1385 | 0.068 | 25.4624 | 2.84 | 0.111 | 72.0383 | 5.9245 | 0.082 |
| Aztreonam | ND | ND | ND | ND | ND | ND | ND | ND | ND |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Antibiotics Pharmacokinetics and Efficacy · Bacterial Identification and Susceptibility Testing
Introduction
1
Carbapenem-resistant Enterobacterales (CRE) have emerged and spread rapidly worldwide, posing a significant threat to human health (Yigit et al., 2001; Mackow and van Duin, 2024). The most common clinical isolates of CRE are Klebsiella pneumoniae and Escherichia coli (Zhang et al., 2018). In 2024, the World Health Organization listed CRE as a critical priority pathogen for the urgent development of new antibiotics (World Health Organization [WHO], 2024). Epidemiological research has shown that the overall annual incidence of CRE in the United States is 2.93 per 100,000 people, whereas in China it is 4 per 10,000 hospital discharges (Guh et al., 2015; Zhang et al., 2018). Furthermore, clinical studies indicate that, compared with infections caused by carbapenem-sensitive Enterobacterales, those caused by CRE are associated with a higher mortality rate (Stewardson et al., 2019).
The main mechanisms causing CRE resistance are porin mutations, efflux pump overexpression, and carbapenemase production. Of these, carbapenemase production is the most important CRE resistance mechanism, and it is divided into classes A, B, and D according to the Ambler classification system. Classes A and D consist of serine β-lactamases, while class B comprises metallo-β-lactamases (MBLs), such as New Delhi metallo-β-lactamase (Al-Marzooq et al., 2024; Dixon et al., 2022). K. pneumoniae isolates from adults primarily produce class A enzymes, predominantly KPC. KPC-2 is the most prevalent in China, whereas KPC-3 and KPC-2 predominate jointly in the Americas and Europe (Falagas et al., 2025). In pediatric patients, metallo-β-lactamases (including NDM-1 and NDM-5) are the primary resistance mechanism, followed by KPC and class D carbapenemases (with OXA-232 and OXA-48 being the most common) (Zhou et al., 2024). The primary resistance mechanism identified in E. coli isolates from both adult and pediatric patients was MBL production, with NDM-5 and NDM-1 being the most commonly detected (Xiong et al., 2023; Boutzoukas et al., 2023). MBLs can hydrolyze almost all β-lactam antibiotics except aztreonam, and new β-lactamase inhibitors are ineffective against them, posing a significant challenge for clinical treatment (Theuretzbacher et al., 2021; Feng et al., 2017). NDM is a typical representative of MBLs. Since NDM-1 was first isolated from K. pneumoniae in 2008 (Yong et al., 2009), more than 90 NDM variants have been identified worldwide, according to the NCBI database (National Center for Biotechnology Information [NCBI], 2025). These variants exhibit different levels of hydrolytic activity against β-lactam antibiotics. For instance, NDM-5 exhibits increased hydrolytic activity against carbapenem antibiotics compared to NDM-1 (Hornsey et al., 2011), whereas NDM-9 and NDM-35 demonstrate enhanced hydrolytic activity against cefiderocol (Poirel et al., 2022; Gaillot et al., 2023). Therefore, strengthening surveillance of NDM-carrying pathogens is essential to track their resistance profiles and guide clinical therapy.
This study describes the recently identified blaNDM variant (bla*NDM–*80) carried by E. coli and isolated from a fecal specimen taken from a 1-year-old patient in the pediatric intensive care unit in China. Through antimicrobial susceptibility testing, whole-genome sequencing, molecular cloning, and enzymatic kinetics assays, we explored its genetic characteristics and resistance mechanisms.
Materials and methods
2
Bacterial strain
2.1
A strain of E. coli carrying a new NDM variant (designated E. coli SE-eco-21) was isolated from a 1-year-old boy hospitalized with severe cough at the pediatric intensive care unit of the Children’s Hospital of Shenzhen. The strain was obtained from stool after 5 days hospitalization and no pathogen was isolated from the sputum. The patient was only successively treated with the ceftazidime and ceftriaxone during the hospitalization. Interestingly, on the seventh day of hospitalization, the follow-up stool specimen showed no detection of E. coli. Subsequently, the patient’s symptoms improved significantly, and he was discharged on the tenth day of hospitalization.
Antimicrobial susceptibility testing
2.2
The minimum inhibitory concentrations (MICs) of antibiotics against E. coli were determined using the broth microdilution method. These were interpreted according to the 2025 breakpoints for all agents tested by the Clinical and Laboratory Standards Institute (CLSI), except for aztreonam-avibactam, sitafloxacin, cefoperazone-sulbactam, eravacycline, and tigecycline (Clinical and Laboratory Standards Institute [CLSI], 2025). The MIC of aztreonam-avibactam was interpreted according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoints (≤4 mg/L susceptible; ≥4 mg/L resistant) (European Committee on Antimicrobial Susceptibility Testing [EUCAST], 2025). The sitafloxacin MIC was interpreted according to the epidemiological cutoff value (ECOFF) breakpoints (≤0.03 mg/L susceptible; ≥0.06 mg/L resistant) (Zhang et al., 2025). The cefoperazone-sulbactam MIC was interpreted according to the report by Barry and Jones (1988) (≤16 mg/L susceptible, 32 mg/L intermediate, ≥64 mg/L resistant). The MICs of eravacycline and tigecycline were interpreted according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST) breakpoints (≤0.5 mg/L susceptible and >0.5 mg/L resistant, respectively) (European Committee on Antimicrobial Susceptibility Testing [EUCAST], 2025). E. coli ATCC 25922 was used as a quality control for antimicrobial susceptibility testing.
Whole-genome sequencing and bioinformatic analysis
2.3
Genomic DNA of E. coli SE-eco-21 was extracted using a commercial kit (TIANGEN, Beijing, China), according to the manufacturer’s protocols. Bacterial genomic DNA was sequenced using the Illumina MiSeq platform (Illumina Inc.) with paired-end reads (2 × 150 bp) for short-read sequencing, and using the Oxford Nanopore MinION platform (Oxford Nanopore, Oxford, United Kingdom) for long-read sequencing. Sequencing reads were de novo assembled using SPAdes software (version 3.13.0) (Bankevich et al., 2012). Antimicrobial resistance genes and plasmid replicon analysis were executed through the Abricate software, referencing the CARD (Zankari et al., 2012) and PlasmidFinder database (Carattoli et al., 2014), respectively. Multilocus sequence typing (MLST) analysis was conducted via the PubMLST database.^1^ Serotyping using ECTyper (version 1.0) with default parameters (Bessonov et al., 2021). Genome annotation was performed using Prokka (Seemann, 2014). The genetic environment structure was analyzed by Easyfig tools (Sullivan et al., 2011).^2^ Comparative genome circle map was conducted by BLAST Ring Image Generator (BRIG) (Alikhan et al., 2011).
Horizontal transfer assays
2.4
A plasmid conjugation assay was performed to characterize the bla*NDM–*80-carrying plasmids. E. coli strains EC600 and J53 were used as recipient strains, while E. coli SE-eco-21 carrying bla*NDM–*80 served as the donor strain in the conjugation experiments. Transconjugants were selected on Luria-Bertani (LB) agar supplemented with rifampicin (50 mg/L) and ampicillin (50 mg/L) for E. coli EC600, and with sodium azide (50 mg/L) and ampicillin (50 mg/L) for E. coli J53. The presence of blaNDM was confirmed by PCR followed by sequencing. Plasmid DNA was extracted from E. coli SE-eco-21 and transferred into E. coli DH5α by electroporation. Transformants were selected on Luria–Bertani (LB) agar supplemented with ampicillin (50 mg/L). The presence of blaNDM was confirmed by PCR and PCR-based sequencing. Antimicrobial susceptibility testing of transformants was determined by the broth microdilution method.
Cloning experiment
2.5
Analysis of antibiotic resistance mediated by bla*NDM–*80 in comparison with bla*NDM–*1 and bla*NDM–*5 was investigated by amplifying either the entire open reading frame (primer: F1: 5′-CCATGATTACGAATTCATGGAATTGCCCAATATTATGCACCC-3′; R1: 5′-CGACTCTAGAGGATCCTCAGCGCAGCTTGTCGG-3′) or the complete gene with its native promoter (primer: F2: 5′-TGACCATGATTACGAATTCGGGACTTGTTCGCACCTTCC-3′; R2: 5′-CGACTCTAGAGGATCCTCAGCGCAGCTTGTCGG-3′). Purified PCR amplicons were cloned into PHSG398 vectors and transformed into E. coli DH5α. Transformants were selected on LB agar supplemented with ampicillin (50 mg/L). The sequence and phenotype mediated by blaNDM were verified by PCR-based sequencing and antimicrobial susceptibility test.
Expression and purification of NDM proteins
2.6
The sequences of NDM-80, NDM-1, and NDM-5 without the peptide signal region were amplified by PCR using primers EcoRI-NDM29-271: 5′-GCAAATGGGTCGCGGATCCGGTGAAAT CCGCCCGACG-3′ and BamHI-NDM29-271: 5′-GTCG ACGGAGCTCGAATTCTCAGCGCAGCTTGTCGG-3′. Purified PCR amplicons were cloned into Pet-28a vectors and transformed into E. coli BL21(DE3). NDM Protein was overexpressed in E. coli BL21(DE3) overnight at a temperature of 19 °C. After that, the protein was collected and purified by nickel affinity chromatography and imidazole elution. The concentration of NDM protein was determined by measuring the absorbance at 280 nm, and concentrated it to 1.5–2.0 mg/mL.
Steady-state kinetic determinations
2.7
Kinetic parameters were determined using purified NDM-80, NDM-1, and NDM-5 in 50 mM HEPES (pH 7.5) supplemented with 100 μM ZnSO4 to detect the hydrolysis of the β-lactams at 25 °C. The real-time absorbances for ceftazidime (257 nm), cefiderocol (259 nm), cefepime (254 nm), imipenem (297 nm), meropenem (298 nm), and aztreonam (318 nm) were detected with a UVProbe spectrophotometer (Kyoto, Japan). The Michaelis–Menten equation was used to calculate and analyze kinetic parameters.
Results
3
Antimicrobial susceptibility testing
3.1
The results of antimicrobial susceptibility testing revealed that E. coli SE-eco-21 exhibited resistance to imipenem, meropenem, meropenem-vaborbactam, ceftolozane-tazobactam, ceftazidime-avibactam, cefepime, ceftazidime, ceftriaxone, ciprofloxacin, cefoperazone-sulbactam, piperacillin-tazobactam, and trimethoprim-sulfamethoxazole. Conversely, the strain demonstrated susceptibility to aztreonam, aztreonam-avibactam, eravacycline, sitafloxacin, tigecycline, colistin and cefiderocol (Table 1).
Whole-genome sequencing assay and characterization of the blaNDM-carrying plasmid
3.2
Next-generation sequencing analysis revealed that E. coli SE-Eco-21 belongs to sequence type (ST) 155 and the O4:H51 serotype. One novel NDM-encoding gene, designated bla*NDM–*80 (Genebank accession: PV430024.1), was identified in E. coli SE-eco-21. The bla*NDM–*80 contains three-point mutations relative to bla*NDM–*1 (Genebank accession: NG_049326.1) at positions 262 (Gat p 388 (G→A), and 460 (A→C). These mutations generate the amino acid substitutions Val88Leu, Asp130Asn, and Met270Leu. The bla*NDM–*80 differs from bla*NDM–*5 (Genebank accession: NG_049337.1) by a single nucleotide at position 388 (G→A), resulting in an amino acid substitution Asp130Asn (Figure 1). Third-generation sequencing revealed that E. coli SE-eco-21 harbored a 5,057,113 bp chromosome and four plasmids ranging in size from 4,308 to 184,004 bp (Table 2). The bla*NDM–*80 was carried by SE-plasmidC, which belongs to the IncX3 plasmid type. The bla*NDM–*80 was the only antibiotic resistance gene present on SE-plasmidC, which was almost identical (100% query coverage and >99.9% nucleotide identity) to many other IncX3-type plasmids carrying blaNDM, such as plasmids GenBank numbers CP017992.1, CP0139407.1, CP0168407.1, MW415443.1, and MW415444.1 (Figure 2A). The complete genetic structure of bla*NDM–*80 was IS5-bla*NDM–*80-ble-trpF-dsbD-IS26-nmuD-ISKox3. Compared to other similar IncX3-type plasmids in the NCBI database, it was found to be absent of the Tn2 and Tn3 transposons (Figures 2A, B).
Alignment of NDM-1 (NG_049326.1), NDM-5 (NG_049337.1), and NDM-80 (PV430024.1) amino acid sequences.
(A) Alignments of plasmids. Comparison of the plasmids SE-plasmidC and other IncX3 plasmids using BRIG. (B) The genetic environment surrounding blaNDM in SE-plasmidC and other IncX3 plasmids.
Plasmid transformation assay
3.3
The results of the plasmid transformation assay (Table 1) show that, compared to recipient E. coli DH5α, the MICs of the transformant E. coli DH5α- SE-plasmidC (bla*NDM–*80 positive) to imipenem, meropenem, meropenem-vaborbactam, ceftolozane-tazobactam, ceftazidime-avibactam, cefepime, ceftazidime, ceftriaxone, cefoperazone-sulbactam, and piperacillin-tazobactam increased by between 64 and 1067-fold. This is consistent with the resistance phenotype observed in the original strains. Notably, no bla*NDM–*80-positive conjugants were detectable when either E. coli J53 or E. coli EC600 was used as the recipient strain. In this study, the plasmid carrying the bla*NDM–*80 gene appears to lack the ability to be transmitted by conjugation, probably due to the absence of the type IV secretion system (T4SS), the type IV coupling protein gene (T4CP), and the relaxase.
Functional analysis of blaNDM–80
3.4
To compare the antibiotic activity profiles of NDM-80 with those of NDM-5 and NDM-1, the relevant genes were cloned and transformed into E. coli DH5α. Two different types of blaNDM-positive transformants were generated (see Table 1). The first type contained only the complete blaNDM open reading frame and was designated E. coli DH5α-PHSG398-bla*NDM–*80, E. coli DH5α-PHSG398-bla*NDM–*5, and E. coli DH5α-PHSG398-bla*NDM–*1. The second type contained the complete blaNDM with its native promoter and was designated E. coli DH5α-PHSG398-promoter-bla*NDM–*80, E. coli DH5α-PHSG398-promoter-bla*NDM–*5, and E. coli DH5α-PHSG398-promoter-bla*NDM–*1. The results of the antimicrobial susceptibility test showed that E. coli DH5α harboring bla*NDM–*80, bla*NDM–*5, or bla*NDM–*1 exhibited reduced susceptibility to all tested β-lactam antibiotics, including carbapenems, cephalosporins, and β-lactam/β-lactamase inhibitors, when compared with E. coli DH5α-PHSG398 (Table 1). Notably, the basal expression of blaNDM open reading frame under the vector promoter (lacZ) affected the MICs toward imipenem, meropenem, meropenem-vaborbactam, and cefepime in DH5α strains. The E. coli DH5α-PHSG398-bla*NDM–*80 transformant showed similar β-lactam antibiotic resistance to the E. coli DH5α-PHSG398-bla*NDM–*5 transformant. Compared to the E. coli DH5α-PHSG398-bla*NDM–*1 transformant, the E. coli DH5α-PHSG398-bla*NDM–*80 transformant had a twofold increase in the MIC of imipenem, meropenem, cefepime, and cefiderocol. However, these values remain within the susceptible range in the absence of the native promoter. Interestingly, transformants expressing blaNDM with the native promoter exhibited significantly higher MICs for imipenem, meropenem, meropenem-vaborbactam, and cefepime, suggesting that the wild-type promoter plays a significant role in enhancing resistance to these antibiotics (see Table 1).
Enzyme activity analysis
3.5
To characterize NDM-80 and investigate the impact of the amino acid point mutation on its enzymatic activity, NDM-80, NDM-5, and NDM-1 were purified, and their kinetic parameters were determined. NDM-80 was found to hydrolyze all of the tested β-lactams except aztreonam (see Figure 3 and Table 3). Compared to NDM-1, NDM-80 exhibited a higher k_(cat)/K_m ratio for cefiderocol, cefepime, imipenem, and meropenem, but a lower k_(cat)/K_m ratio for ceftazidime. Kinetic analysis revealed that NDM-80 probably exhibits higher enzymatic activity toward cefiderocol, cefepime, imipenem, and meropenem, but lower activity toward ceftazidime than NDM-1. Compared to NDM-5, NDM-80 had a lower k_(cat)/K_m ratio for cefiderocol, imipenem, and meropenem, but a higher k_(cat)/K_m ratio for cefepime. This suggests that NDM-80 likely exhibits lower enzymatic activity for cefiderocol, imipenem, and meropenem, but higher enzymatic activity for cefepime than NDM-5.
Catalytic efficiency of NDM-1, NDM-5, and NDM-80 against β-lactam antibiotics.
Discussion
4
Carbapenem-resistant Enterobacterales are widespread in the pediatric population, particularly among children in pediatric intensive care units, where they cause infections and pose a serious global public health issue (Fu et al., 2023; Ruvinsky et al., 2022). The production of Class B β-lactamase NDM is the primary mechanism by which CRE isolated from pediatric patients exhibit carbapenem resistance. NDM exhibits significant hydrolytic activity against most β-lactam antibiotics, resulting in limited treatment options for infection control (Han et al., 2020). According to recommendations from the Infectious Diseases Society of America, the preferred treatment options for NDM and other MBL-producing Enterobacterales infections are ceftazidime-avibactam in combination with aztreonam or cefiderocol as monotherapy (Tamma et al., 2024). Aztreonam is resistant to hydrolysis by NDMs, and avibactam is generally effective at inhibiting the activity of other co-produced β-lactamases, such as KPCs, ESBLs, and AmpCs. When used in combination, enhanced antibacterial efficacy can be achieved (Tamma et al., 2024). Cefiderocol, which functions as an iron-chelating cephalosporin, is stable against metalloenzymes and is a significant therapeutic alternative; however, it is not currently available for clinical use in China (Sato and Yamawaki, 2019; Aoki et al., 2018). This study reports a novel blaNDM variant, designated blaNDM–80, identified in carbapenem-resistant E. coli for the first time. Antimicrobial susceptibility testing showed that the clinical strain harboring the blaNDM–80 variant exhibited high-level resistance to all β-lactam antimicrobial agents except for aztreonam, aztreonam-avibactam, and cefiderocol. Furthermore, accumulating evidence indicates that NDM is the only metallo-β-lactamase capable of mediating cefiderocol resistance, primarily via blaNDM overexpression, alterations to transferrin, and the emergence of novel mutants (e.g., blaNDM–9 and blaNDM–35) (Simner et al., 2022; Nurjadi et al., 2022; Poirel et al., 2022). In our study, molecular cloning results showed that the cefiderocol MIC for the blaNDM–80-positive transformant (1 mg/L) was double that for the blaNDM–1-positive transformant (0.5 mg/L), yet both values remain well within the susceptible range. This single dilution increase, which may fall within the inherent variability of the testing method, suggests that the NDM-80 variant probably does not confer a clinically meaningful reduction in cefiderocol susceptibility compared to NDM-1. However, it is crucial to strengthen surveillance of pathogens harboring blaNDM variants in order to prevent the emergence of high-level cefiderocol-resistant strains and to inform clinical antimicrobial therapy.
The blaNDM resistance genes are typically located on conjugative plasmids, with IncFII and IncX3 plasmids being the most prevalent. This enables them to be disseminated widely among pathogenic bacteria. To date, the NCBI database has documented more than 90 blaNDM variants, 49 of which were initially isolated from E. coli. Internationally, blaNDM–1 is the most prevalent variant among E. coli isolates, followed by blaNDM–5, blaNDM–9, and blaNDM–7 (Wu et al., 2019). In contrast, blaNDM–5 is most prevalent in CRE strains isolated in China, followed by blaNDM–1, with other variants rarely detected (Li et al., 2024; Han et al., 2020). Compared with blaNDM–1, the blaNDM–80 variant identified in E. coli in this study harbors three amino acid mutations (including V88L, D130N, and M154L). By contrast, only a single amino acid mutation (D130N) is present compared to blaNDM–5. The blaNDM–80 is located on an IncX3 plasmid. Previous studies have demonstrated that the IncX3 plasmid can transfer blaNDM between different enterobacterial species (Shao et al., 2020; Moussa et al., 2024). Interestingly, no blaNDM–80-positive conjugants were detectable when E. coli J53 or E. coli EC600 were used as receptors. Sequence alignment analysis of this plasmid with other IncX3 plasmids carrying blaNDM resistance genes revealed that, compared with other plasmids (Figure 2B), the upstream region of blaNDM–80 lacks the Tn2 and Tn3 transposons. Accumulating evidence has demonstrated that the Tn3 family of transposons plays a crucial role in mediating the horizontal transfer of resistance genes (Tang et al., 2024). Furthermore, the plasmid carrying blaNDM–80 lacks the T4SS, T4CP, and relaxase. Therefore, we hypothesize that these common factors have rendered the IncX3 plasmid in this study non-conjugative. This may restrict the dissemination of blaNDM–80 among Enterobacterales.
Different blaNDM variants exhibit distinct levels of drug resistance. For example, blaNDM–5 and blaNDM–7 enhance resistance to meropenem, imipenem, and ertapenem more than blaNDM–1 does (Hornsey et al., 2011; Göttig et al., 2013), while blaNDM–9 and blaNDM–35 increase resistance to cefiderocol more than blaNDM–1 does (Gaillot et al., 2023; Poirel et al., 2022). The blaNDM–80 variant isolated in this study exhibits a similar antimicrobial resistance profile to blaNDM–5. Studies on crystal structures have shown that the active site of blaNDM is located at the bottom of a shallow groove formed by loops L3 and L10 (Tada et al., 2013). In the present study, the other two mutation sites (V88L and D130N), except for the M154L mutation, are not located within this active site. This may account for the similar antimicrobial resistance profiles between blaNDM–80 and blaNDM–5. Additionally, electroporation and molecular cloning studies revealed that transformants harboring plasmid vectors with the wild-type promoter upstream of blaNDM exhibited significant resistance to imipenem, meropenem, meropenem-vaborbactam, and cefepime. Conversely, transformants lacking this wild-type promoter exhibited only a slight increase in MICs for these antimicrobials, which did not reach the threshold for drug resistance. These findings further confirm that the wild-type promoter upstream of blaNDM is a crucial factor in blaNDM-mediated resistance to these agents, consistent with previous research reports (Göttig et al., 2013).
There are several limitations to our study. First, the conjugation experiments were conducted exclusively under controlled in vitro conditions and lack validation in clinical settings. The vitro environment differs substantially from the complex physiological milieu in vivo. Second, our study lacks therapeutic efficacy testing; antibiotics that are sensitive in vitro may not be effective in vivo. In conclusion, the present study identified a novel blaNDM variant, designated blaNDM–80, in E. coli isolated from a 1-year-old patient. Investigating blaNDM–80 expands our understanding of blaNDM variants, which may arise from the spontaneous adaptive evolution of pathogens under selective antimicrobial pressure, particularly carbapenems. Therefore, establishing a drug resistance surveillance network for pathogens producing NDM enzyme variants is imperative in the future. This will clarify the global incidence, resistance mechanisms, transmission characteristics, and risk factors for infection with these drug-resistant strains, thereby providing a scientific basis for curbing the further dissemination of such resistant pathogens.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alikhan N. F. Petty N. K. Ben Zakour N. L. Beatson S. A. (2011). BLAST ring image generator (BRIG): Simple prokaryote genome comparisons. BMC Genomics 12:402. 10.1186/1471-2164-12-402 21824423 PMC 3163573 · doi ↗ · pubmed ↗
- 2Al-Marzooq F. Ghazawi A. Allam M. Collyns T. Saleem A. (2024). Novel variant of New Delhi metallo-beta-lactamase bla NDM–60 discovered in a clinical strain of Escherichia coli from the United Arab Emirates: An emerging challenge in antimicrobial resistance. Antibiotics 13:1158. 10.3390/antibiotics 13121158 39766548 PMC 11672588 · doi ↗ · pubmed ↗
- 3Aoki T. Yoshizawa H. Yamawaki K. Yokoo K. Sato J. Hisakawa S. (2018). Cefiderocol (S-649266), A new siderophore cephalosporin exhibiting potent activities against Pseudomonas aeruginosa and other gram-negative pathogens including multi-drug resistant bacteria: Structure activity relationship. Eur. J. Med. Chem. 155 847–868. 10.1016/j.ejmech.2018.06.014 29960205 · doi ↗ · pubmed ↗
- 4Bankevich A. Nurk S. Antipov D. Gurevich A. A. Dvorkin M. Kulikov A. S. (2012). SP Ades: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19 455–477. 10.1089/cmb.2012.0021 22506599 PMC 3342519 · doi ↗ · pubmed ↗
- 5Barry A. L. Jones R. N. (1988). Criteria for disk susceptibility tests and quality control guidelines for the cefoperazone-sulbactam combination. J. Clin. Microbiol. 26 13–17. 10.1128/jcm.26.1.13-17.1988 3343304 PMC 266166 · doi ↗ · pubmed ↗
- 6Bessonov K. Laing C. Robertson J. Yong I. Ziebell K. Gannon V. P. J. (2021). EC Typer: In silico Escherichia coli serotype and species prediction from raw and assembled whole-genome sequence data. Microb. Genom. 7:000728. 10.1099/mgen.0.000728 34860150 PMC 8767331 · doi ↗ · pubmed ↗
- 7Boutzoukas A. E. Komarow L. Chen L. Hanson B. Kanj S. S. Liu Z. (2023). International Epidemiology of carbapenemase-producing Escherichia coli. Clin. Infect. Dis. 77 499–509. 10.1093/cid/ciad 288 37154071 PMC 10444003 · doi ↗ · pubmed ↗
- 8Carattoli A. Zankari E. García-Fernández A. Voldby Larsen M. Lund O. Villa L. (2014). In silico detection and typing of plasmids using Plasmid Finder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 58 3895–3903. 10.1128/aac.02412-14 24777092 PMC 4068535 · doi ↗ · pubmed ↗