Bactericidal and antibiofilm activity of lactic acid bacteria-derived cell free extracts against dairy-associated spoilage and pathogenic bacteria
Andressa Falqueto, Rafaela da Silva Rodrigues, Luana Virgínia Souza, Antônio Fernandes de Carvalho, Cinzia Caggia, Luiz Augusto Nero, Solimar Gonçalves Machado, Cinzia Lucia Randazzo

TL;DR
This study shows that extracts from lactic acid bacteria can effectively kill and prevent biofilms of spoilage and harmful bacteria in dairy products.
Contribution
The study identifies specific lactic acid bacteria strains and their extracts with strong antimicrobial and antibiofilm properties for use in dairy systems.
Findings
Non-neutralized cell-free supernatants showed strong bactericidal activity against spoilage and pathogenic bacteria.
Lactiplantibacillus plantarum Q4C3 and other strains were identified as the most effective biocontrol agents.
SEM analysis revealed membrane disruption in treated cells, confirming the efficacy of the extracts.
Abstract
This study aimed to evaluate the antimicrobial and antibiofilm potential of 15 LAB strains using five types of LAB-derived preparations: cell-free supernatants (CFS), sonicated-inactivated cells (IC), their combination (ICS), and their neutralized variants (CFS N, ICS N) to identify the most effective strain-extract combinations for potential application as natural biocontrol agents in dairy systems. Antimicrobial activity was assessed through agar diffusion assays, growth and biofilm inhibition, and determination of minimum inhibitory (MIC) and bactericidal (MBC) concentrations. The extracts were further characterized by pH and organic acid profiles using high-performance liquid chromatography (HPLC), and their mechanisms of action were investigated through cellular leakage assays, time–kill kinetics, and scanning electron microscopy (SEM). Several LAB-derived extracts exhibited strong…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
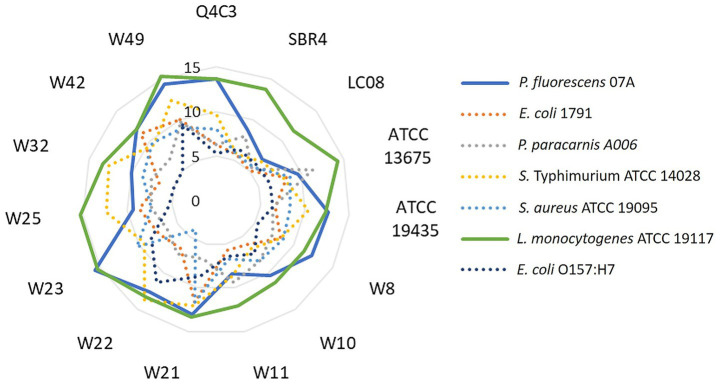 Figure 1
Figure 1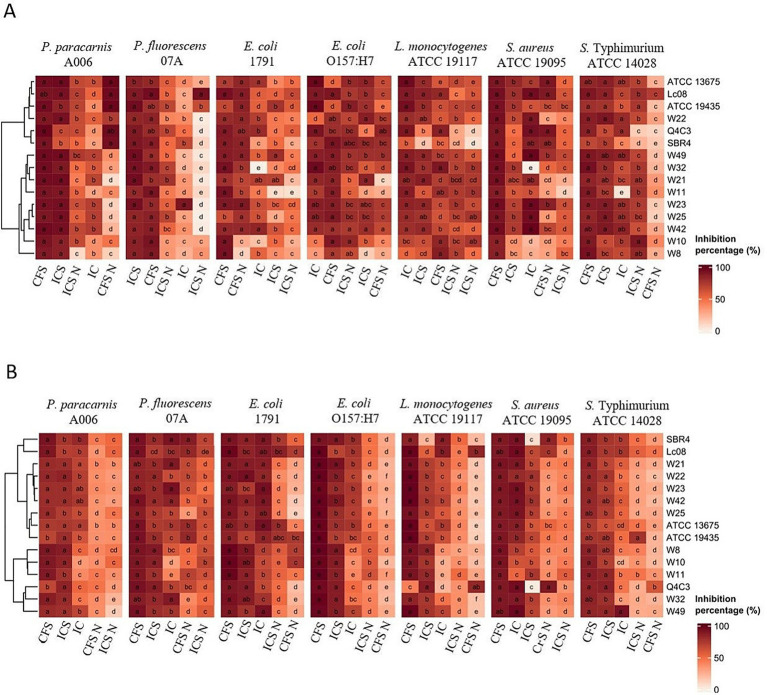 Figure 2
Figure 2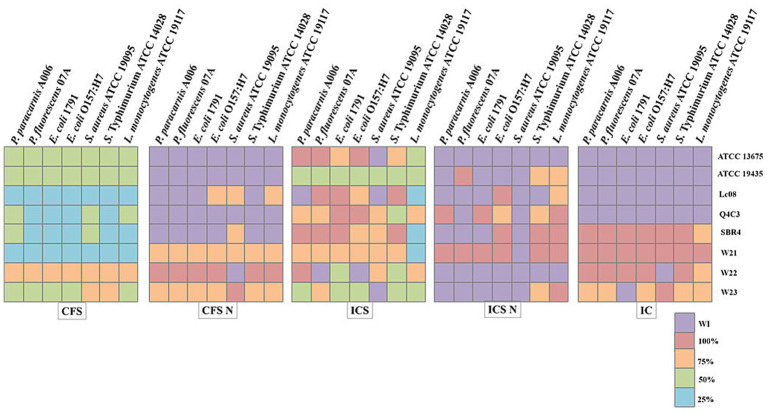 Figure 3
Figure 3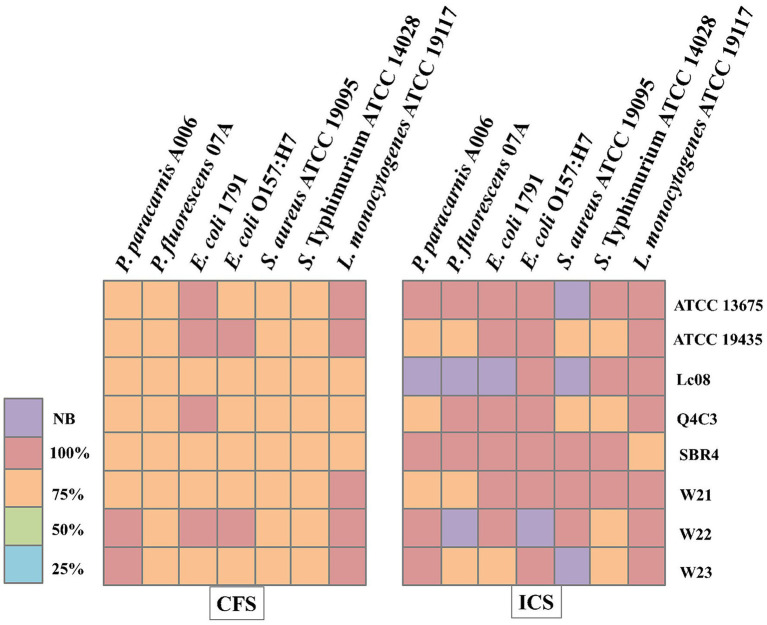 Figure 4
Figure 4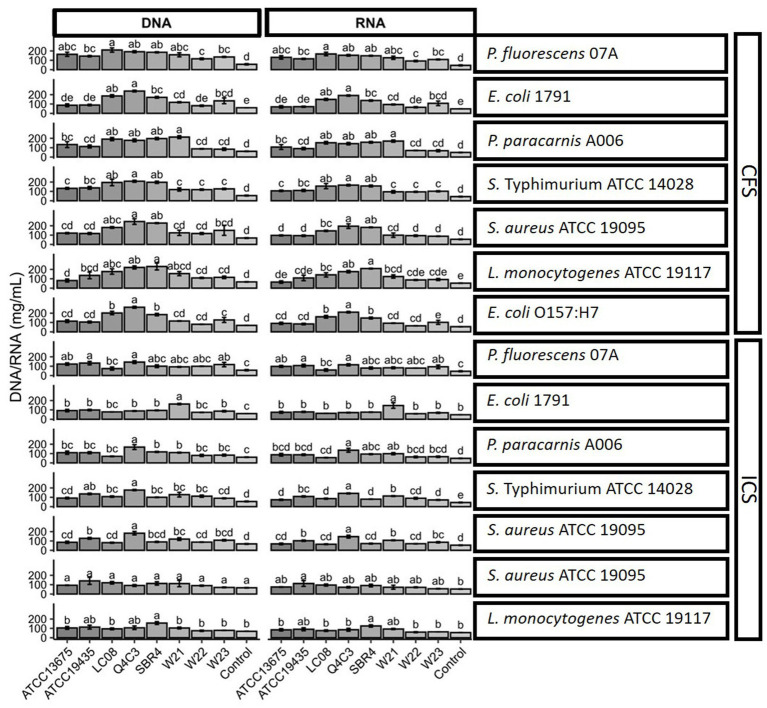 Figure 5
Figure 5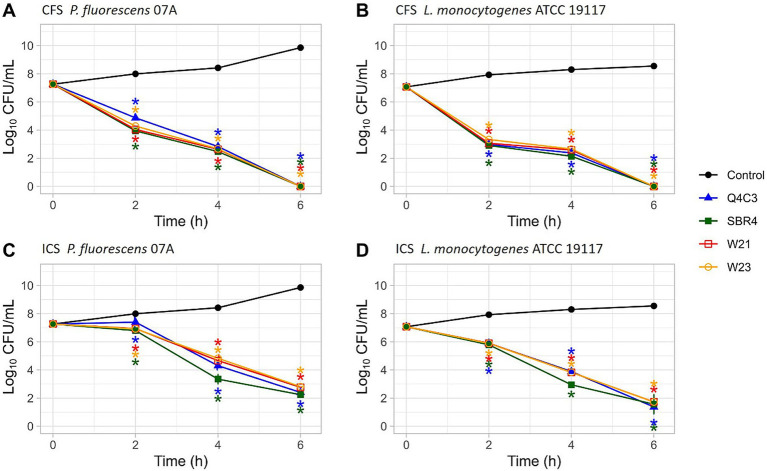 Figure 6
Figure 6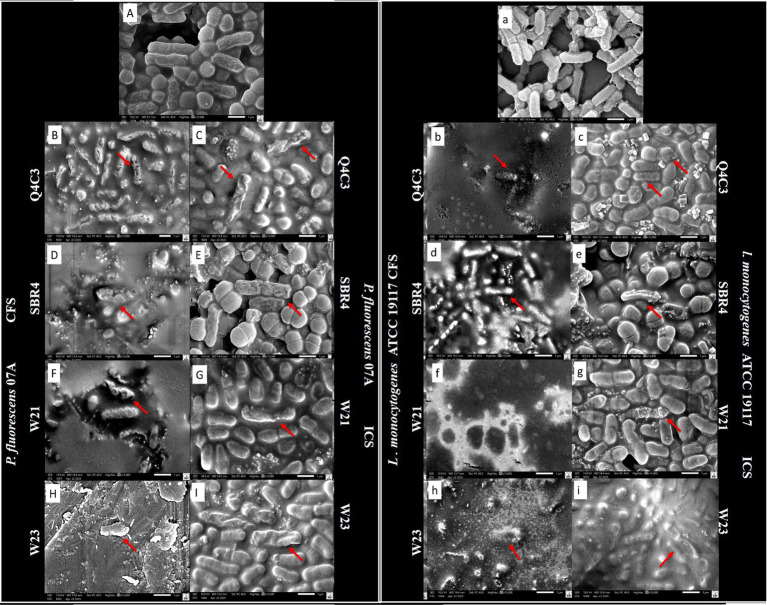 Figure 7
Figure 7| Microorganisms | Source | Growth conditions | Abbreviations | Assays performed | References |
|---|---|---|---|---|---|
| LAB strains | |||||
| Artisanal cheese/Ilha do Marajó – Pará, Brazil | Cultured in MRSa broth or agar at 30 °C (±2 °C) for 24–48 h | Antagonistic/antibiofilm/MIC/MBC/cell-leakage/Time–kill/SEM |
| ||
| Silage | Antagonistic/antibiofilm/MIC/MBC/cell-leakage/Time–kill/SEM |
| |||
| Raw Milk | Antagonistic/antibiofilm/MIC/MBC/cell-leakage/Time–kill |
| |||
| Pasture/Campos das vertentes – Minas Gerais, Brazil | Antagonistic/antibiofilm/MIC/MBC/cell-leakage/Time–kill/SEM |
| |||
| Raw milk /Ilha do Marajó – Pará, Brazil | Antagonistic and antibiofilm activity | ||||
| Artisanal cheese/Sul do Pará - Pará, Brazil | Antagonistic and antibiofilm activity | ||||
| Artisanal cheese/Ilha do Marajó - Pará, Brazil | Antagonistic and antibiofilm activity | ||||
| Soil/Campos das vertentes – Minas Gerais, Brazil | Antagonistic and antibiofilm activity | ||||
| Antagonistic and antibiofilm activity | |||||
| Silage/Campos das vertentes – Minas Gerais, Brazil | Antagonistic and antibiofilm activity | ||||
| Pasture/Campos das vertentes - Minas Gerais, Brazil | Antagonistic/antibiofilm/MIC/MBC/cell-leakage/Time–kill/SEM | ||||
| Antagonistic/antibiofilm/MIC/MBC/cell-leakage | |||||
| Antagonistic and antibiofilm activity | |||||
| Culture collection | Antagonistic/antibiofilm/MICC/MBC/cell-leakage | ATCC* | |||
| Raw milk, culture collection | Antagonistic/antibiofilm/MIC/MBC/cell-leakage | ATCC* | |||
| Pathogen strains | |||||
| Pooled heart and liver tissue of four-week-old chickens | Cultured in BHIb broth or agar at 37 °C (±2 °C) for 24 h | Antagonistic/antibiofilm/MIC/MBC/cell-leakage | ATCC* | ||
| Sheep | Antagonistic/antibiofilm/MIC/MBC/cell-leakage/Time–kill/SEM | ATCC* | |||
| Patient’s leg abscess | Antagonistic/antibiofilm/MIC/MBC/cell-leakage | ATCC* | |||
| Culture collection | Antagonistic/antibiofilm/MIC/MBC/cell-leakage | InovaLeite** | |||
| Spoilage strains | |||||
| Fresh Cheese/MG | Cultured in BHIb broth or agar at 25 °C (±2 °C) for 24 h | Antagonistic/antibiofilm/MIC/MBC/cell-leakage |
| ||
| Raw Milk | Antagonistic/antibiofilm/MIC/MBC/cell-leakage/Time–kill/SEM |
| |||
| Raw Milk | Cultured in BHIb broth or agar at 37 °C (±2 °C) for 24 h | Antagonistic/antibiofilm/MIC/MBC/cell-leakage | INSPOA*** | ||
| LAB | CFS | CFS N | ICS | ICS N | IC |
|---|---|---|---|---|---|
| 4.46 ± 0.11 | 7.04 ± 0.2 | 4.52 ± 0.01 | 6.96 ± 0.016 | 6.37 ± 0.11 | |
| 4.18 ± 0.06 | 7.18 ± 0.01 | 4.35 ± 0.02 | 6.89 ± 0.07 | 6.21 ± 0.15 | |
| 4.11 ± 0.30 | 6,98 ± 0.04 | 4.61 ± 0.04 | 6.94 ± 0.07 | 6.19 ± 0.04 | |
| 4.36 ± 0.41 | 7.06 ± 0.03 | 4.32 ± 0.04 | 6.89 ± 0.13 | 6.22 ± 0.06 | |
| 4.03 ± 0.13 | 7.02 ± 0.01 | 4.23 ± 0.01 | 6.65 ± 0.24 | 6.17 ± 0.07 | |
| 3.84 ± 0.06 | 7.12 ± 0.01 | 3.87 ± 0.04 | 6.85 ± 0.08 | 6.02 ± 0.08 | |
| 4.08 ± 0.01 | 7.06 ± 0.01 | 4.19 ± 0.01 | 7.02 ± 0.04 | 6.21 ± 0.14 | |
| 4.33 ± 0.17 | 6.96 ± 0.04 | 4.32 ± 0.04 | 7.02 ± 0.01 | 6.26 ± 0.16 | |
| 4.13 ± 0.10 | 7.17 ± 0.04 | 4.10 ± 0.02 | 6.92 ± 0.11 | 6.18 ± 0.08 | |
| 4.33 ± 0.03 | 7.18 ± 0.01 | 4.22 ± 0.02 | 7.15 ± 0.15 | 6.16 ± 0.04 | |
| 4.45 ± 0.10 | 7.18 ± 0.04 | 4.27 ± 0.06 | 7.06 ± 0.11 | 6.30 ± 0.10 | |
| 4.40 ± 0.10 | 7.13 ± 0.05 | 4.23 ± 0.05 | 6.94 ± 0.08 | 6.24 ± 0.14 | |
| 4.39 ± 0.06 | 7.15 ± 0.02 | 4.25 ± 0.08 | 7.12 ± 0.06 | 6.28 ± 0.14 | |
| 4.43 ± 0.16 | 6.98 ± 0.08 | 4.53 ± 0.03 | 7.15 ± 0.06 | 6.23 ± 0.15 | |
| 4.37 ± 0.04 | 6.97 ± 0.07 | 4.51 ± 0.08 | 7.01 ± 0.08 | 6.21 ± 0.12 |
| LAB | Latic | Citric | Succinic | Formic | Acetic | Propionic | Isobutiric | Butiric | Isovaleric | Valeric |
|---|---|---|---|---|---|---|---|---|---|---|
| CFS | ||||||||||
| Q4C3 | 189.657 | 0.000 | 4.124 | 94.381 | 90.179 | 13.167 | 0.511 | 28.253 | 0.000 | 0.071 |
| SBR4 | 256.962 | 10.365 | 2.252 | 101.279 | 72.974 | 0.270 | 0.533 | 2.871 | 0.083 | 0.038 |
| LC08 | 279.431 | 12.502 | 3.077 | 79.968 | 88.306 | 1.734 | 0.944 | 0.205 | 0.707 | 0.160 |
| ATCC 13675 | 169.543 | 0.000 | 2.823 | 0. 000 | 137.138 | 3.358 | 1.089 | 0.504 | 0.000 | 0.000 |
| ATCC 19435 | 191.744 | 11.328 | 3.947 | 68.389 | 78.583 | 13.247 | 0.883 | 26.950 | 0.000 | 0.110 |
| W21 | 164.604 | 11.099 | 3.883 | 60.599 | 79.309 | 14.017 | 0.897 | 33.268 | 0.000 | 0.135 |
| W22 | 183.114 | 3.212 | 6.550 | 83.348 | 94.540 | 15.415 | 1.149 | 34.518 | 0.000 | 0.140 |
| W23 | 164.000 | 11.162 | 4.050 | 71.696 | 79.494 | 14.668 | 1.378 | 34.466 | 0.000 | 0.000 |
| ICS | ||||||||||
| Q4C3 | 167.664 | 0.000 | 3.086 | 73.892 | 92.033 | 12.350 | 0.665 | 32.878 | 0.000 | 0.100 |
| SBR4 | 187.829 | 10.247 | 0.938 | 77.992 | 69.889 | 0.211 | 0.483 | 3.124 | 0.000 | 0.119 |
| LC08 | 181.668 | 11.212 | 1.130 | 70.564 | 77.135 | 0.879 | 0.657 | 0.204 | 0.381 | 0.130 |
| ATCC 13675 | 158.351 | 0.000 | 3.297 | 0.000 | 90.769 | 20.496 | 1.283 | 0.464 | 0.000 | 0.135 |
| ATCC 19435 | 188.513 | 11.299 | 4.197 | 67.659 | 79.593 | 15.140 | 1.284 | 24.378 | 0.000 | 0.114 |
| W21 | 158.572 | 10.614 | 3.892 | 61.267 | 77.673 | 14.458 | 1.229 | 31.670 | 0.000 | 0.047 |
| W22 | 154.039 | 10.148 | 3.409 | 60.374 | 74.436 | 12.518 | 0.844 | 33.325 | 0.000 | 0.099 |
| W23 | 157.988 | 10.578 | 3.906 | 68.385 | 77.738 | 0.151 | 1.308 | 31.509 | 0.000 | 0.139 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Bacterial biofilms and quorum sensing · Milk Quality and Mastitis in Dairy Cows
Introduction
1
Microbiological contamination remains one of the major challenges in cheese production, where both pathogenic and spoilage microorganisms can compromise quality, reduce shelf life, and pose health risks to consumers (Fusco et al., 2020). Pathogens, such as Salmonella spp., Listeria monocytogenes, Staphylococcus aureus, and pathogenic Escherichia coli strains continue to be frequently reported in dairy products (Mantovam et al., 2025), with contamination often linked to failures during production, handling, or storage including inadequate hygiene during milking and herd management, insufficient cleaning and sanitation of equipment that promotes biofilm formation, improper temperature control and breaks in the cold chain, post-pasteurization contamination, and poor environmental management (Jayarao and Henning, 2001; Oliver et al., 2005; Kousta et al., 2010; Martin et al., 2023). Spoilage organisms, particulary Pseudomonas spp. and coliforms, also contribute to product deterioration through pigment formation, enzymatic degradation, and gas production, leading to undesirable sensory and structural changes (Martin et al., 2016; Kumar et al., 2019).
These challenges underscore the need for effective and natural strategies to control both pathogenic and spoilage organisms in dairy matrices. Lactic acid bacteria (LAB) play a central role in food fermentation and are well known for producing antimicrobial metabolites, including organic acids and bacteriocins, that inhibit undesirable microorganisms (Snyder et al., 2024). Recently, interest has expanded toward the use of LAB-derived extracts, which contain bioactive compounds obtained from inactivated cells and culture supernatants. Unlike probiotics or bio-preservative cultures that rely on the presence of viable microorganisms, LAB-derived extracts offer antimicrobial functionality through metabolites alone, making them attractive for clean-label food applications (Raman et al., 2022; Banicod et al., 2025; Choi et al., 2024).
In this context, preparations obtained from LAB are frequently discussed in the literature within the framework of postbiotics. However, according to the International Scientific Association for Probiotics and Prebiotics (ISAPP), postbiotics require demonstrated health benefit to the host (Salminen et al., 2021). When no host-related effects are addressed to the LAB evaluated strains, the preparations are not classified as postbiotics and are instead framed as LAB-derived antimicrobial extracts intended for food biocontrol applications. These preparations include cell-free supernatants, culture media enriched with extracellular metabolites, and intracellular components released after cell inactivation, representing a diverse set of bioactive microbial products (Karabacak Aydin et al., 2026).
Although combined physical, chemical, and biological treatments often enhance microbial inactivation (Ashrafudoulla et al., 2024), the synergistic action of multiple metabolites within a single LAB-derived extract may also potentiate antimicrobial effects (Soltani et al., 2022; Teneva and Denev, 2023). LAB-derived compounds have demonstrated promising activity against biofilm formation and microbial contamination in food systems, aligning with the growing demand for natural and minimally processed products (Yildirim et al., 2018; Moradi et al., 2020).
Comparative evaluations of multiple LAB strains and their derived extracts, particularly in the context of foodborne pathogens and dairy-related spoilers, are still limited. Furthermore, the relative contribution of organic acids, cell-associated components, and other metabolites to the antimicrobial activity of different extract preparations is not yet fully understood. Evaluating extracts containing only supernatants, only inactivated cells, or combinations of both, either native or neutralized, can help clarify the mechanisms underlying microbial inhibition and support their rational application in dairy systems.
Accordingly, this study aimed to comparatively screen 15 LAB strains for antimicrobial and antibiofilm activity against dairy-associated spoilage and pathogenic microorganisms. Five types of LAB-derived preparations (cell-free supernatants, inactivated cells, their combination, and their neutralized variants) were systematically evaluated to determine the contribution of organic acids and cell-associated components to microbial inhibition. This approach enabled the identification and selection of the most effective strain–extract combinations with potential application as natural biocontrol agents in dairy systems. Although antimicrobial activity of LAB-derived cell-free supernatants has been widely reported, most studies focus on a limited number of strains and extract types. The novelty of the present work lies in the systematic, strain-level comparison of 15 LAB strains using five distinct extract preparations, including neutralized variants, combined with an integrated functional and mechanistic evaluation. By linking antimicrobial and antibiofilm performance to organic acid profiles, membrane damage, and killing kinetics, this study provides a rational framework for selecting LAB-derived extracts with enhanced efficacy and industrial relevance, rather than reporting isolated antimicrobial effects.
Materials and methods
2
Microorganisms and growth conditions
2.1
Fifteen strains of LAB and seven strains of pathogenic and spoilage microorganisms (hereafter referred to as contaminants) were used (Table 1). The contaminants strains were maintained at −80 °C (±2 °C) in Brain Heart Infusion (BHI) broth (Kasvi, Curitiba, Brasil) while the LAB were maintained at the same temperature in Man, Rogosa and Sharpe (MRS) broth (Kasvi, Curitiba, Brasil). The culture medium was supplemented with 20% (v/v) glycerol.
Before each experiment, the stock cultures stored at −80 °C (±2 °C) were refreshed by inoculating 1% (v/v) into fresh medium (Table 1). Incubation was performed under the optimal growth conditions for each microorganism under static conditions (Table 1). After growth, cell suspensions were diluted in 0.85% (w/v) saline solution to an optical density (OD) at 600 nm of 0.200 to 0.280 corresponding to approximately 1.5 × 10^8^ CFU/mL. OD was measured using a UV-5100 UV–Vis spectrophotometer (Global Trade Technology, China). The bacterial population in the standardized cell suspensions was confirmed by plate counting on solid media using the microdrop technique (Morton, 2001), followed by incubation under the conditions provided for each microorganism (Table 1).
Preparation of LAB-derived extracts
2.2
LAB-derived extracts evaluated in this study comprised cell-free supernatants (CFS), inactivated cells (IC), their combination (ICS), and their neutralized variants (CFS N, ICS N) prepared as described below.
Cell-free supernatants (CFS) were obtained from the 15 standardized LAB suspensions (1.5 × 10^8^ CFU/mL) by centrifugation at 10,000 g for 10 min at 4 °C. The supernatants were filtered through 0.22 μm pore-size membranes to obtain the CFS which was stored frozen (−20 °C) until use (İncili et al., 2021). For inactive cells plus cell-free supernatants (ICS), standardized LAB suspensions were sonicated for 10 min in an ice bath using a probe sonicator (Ultrasonic Probe System QR850, Ecosonics, Brazil) operating at 40 kHz and 100% amplitude. Similarly, to obtain the inactive cells (IC), the standardized LAB suspensions were centrifuged at 10,000 g at 4 °C for 15 min. The recovered pellets were washed three times, resuspended in sterile distilled water and sonicated under the same conditions used for ICS (Sharafi et al., 2023). The neutralized cell free supernatant (CFS N) and the neutralized inactive cells plus cell-free supernatant (ICS N) were prepared in the same way, with 1 M NaOH added to adjust the pH to approximately 7.0. Cell death was confirmed for all extracts by plating them under optimal conditions (Table 1), as well as for the negative growth control in all subsequent analyses.
Once obtained, the extracts had their pH measured. The measurements were conducted according to Fusieger et al. (2020a), using a calibrated digital pH meter (Hanna Instruments, São Paulo, SP, Brazil) and the extracts were subsequently stored at −20 °C.
Antibacterial activity
2.3
Agar diffusion method
2.3.1
The LAB antimicrobial activity was screened using a modified colony overlay method as described by Jeong et al. (2024). Briefly, a loopful of the revived LAB cultures was streaked onto MRS agar plates which was left half-open to dry for 1 h under aseptic conditions. The plates were overlaid with 10 mL of previously homogenized BHI agar (cooled to approximately 45 °C) containing each pathogenic or spoilage strain at a final population of 10^7^ CFU/mL. After incubation for 24–48 h under the optimal conditions of each bacteria (Table 1), the presence of an inhibition zone (a clear halo ≥10 mm in diameter from the streak edge), was considered indicative of positive antagonistic activity.
Antagonistic and antibiofilm activity
2.3.2
To the antagonistic activity, the extracts were tested against pathogens and spoilage organisms using a microplate assay. Aliquots of 90 μL of BHI broth (1×) were mixed with an equal volume of each LAB-derived extract in 96-well microtiter plates. Then, each well was inoculated with 20 μL of contaminant cell suspensions standardized to 10^6^ CFU/mL. The plates were incubated at 37 or 25 °C, depending on the inoculated microorganism (Table 1), for 24 h in a microplate spectrophotometer (Multiskan™ GO, Thermo Fisher, Scientific Inc., Finland). The OD was measured at 600 nm every 60 min for 24 h (Arena et al., 2016).
Using the same incubation conditions employed for the antagonistic activity, the antibiofilm activity of LAB-derived extracts was evaluated as described by Kim et al. (2022), with minor modifications. Briefly, contaminants were co-incubated with the LAB-derived extracts in 96-well microtiter plates under the conditions described above, allowing biofilm formation in the presence of the extracts for 24 h. After incubation, the culture was carefully removed, and the wells were gently washed three times with phosphate-buffered saline (PBS; pH 7.4; Sigma-Aldrich) to remove non-adhered cells. The adhered biofilm was stained with 200 μL of 0.1% (w/v) crystal violet (Sigma-Aldrich) at 25 °C (±2 °C) for 20 min. Excess stain was removed by washing the wells three times with PBS, followed by drying at 37 °C for 20 min. Subsequently, 200 μL of 95% (v/v) ethanol (Sigma-Aldrich) was added to solubilize the bound dye for 15 min at room temperature. Biofilm biomass was quantified by measuring absorbance at 590 nm (OD590) using a microplate spectrophotometer (Multiskan™ GO, Thermo Fisher Scientific Inc., Finland).
Positive controls consisted of wells containing the contaminants cell suspensions (10^6^ CFU/mL) and BHI broth without LAB-derived extracts, whereas negative controls contained BHI broth supplemented with the corresponding LAB-derived extract.
Absorbance values were baseline-corrected by subtracting the negative controls, and biofilm inhibition (%) was calculated relative to the positive control according to Equation 1. The OD590 values of the positive controls (ODc) were compared with those of the treated cultures (ODt), and biofilm inhibition (%) was calculated according to Equation 1.
The results obtained from the antagonistic activity and antibiofilm assays were normalized using Z-scores, calculated according to Equation 2 (Du et al., 2022), where X represents the observed value, μ the dataset mean, and σ the standard deviation. The Z-score indicates how many standard deviations a value deviates from the mean, facilitating data interpretation. Z-score values were considered to rank the LAB-derived extracts, prioritizing those that showed Z-score < 1 (less than one standard deviation above or below the population mean).
All experiments were carried out in triplicate with two independent biological replicates. The LAB-derived extracts from the eighth strains showing the strongest antimicrobial and antibiofilm activity were selected for further analysis.
Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC)
2.4
The MIC and MBC of LAB-derived extracts were determined based on the microplate dilution technique detailed by Sadekuzzaman et al. (2018) with modifications. The LAB-derived extracts were diluted in BHI to obtain concentrations: 100 (no dilution), 75, 50, 25 and 12.5% (v/v) and distributed in the 96-well microplate (180 μL/well). Aliquots of 20 μL of contaminant cell suspension, adjusted to 10^5^ CFU/mL were added to each well. Plates were incubated at the optimal growth temperature for each evaluated strain (Table 1), and OD measurements at 600 nm were recorded after 24 h using a microplate spectrophotometer (MultiscanGo, ThermoFisher, Finland). Controls included fresh BHI broth (blank), BHI broth inoculated with contaminants without LAB-derived extracts (positive control), and BHI broth with LAB-derived extracts without contaminants (negative control). All controls were incubated under the same conditions as the test samples. The MIC was defined as the lowest extract concentration that completely inhibited bacterial growth (ΔD ≈ 0). To determine MBC, the content of each well was plated on BHI agar using microdrop technique (Morton, 2001). The Petri dishes were incubated as described in Table 1. The MBC was defined as the lowest LAB-derived extract concentration that prevented colony formation. The experiments were carried out in triplicate with two independent biological replicates. For the subsequent characterization of organic acid profiles and the further assays, extracts that demonstrated bactericidal activity were selected.
Characterization of the organic acid profile
2.5
Organic acid analysis was performed using a high-performance liquid chromatography system (HPLC; Shimadzu, São Paulo, Brazil) equipped with a refractive index detector (RID-20A), following the methodology described by Zhang et al. (2024), with minor modifications. Initially, the selected LAB-derived extracts were centrifuged at 10,000×g for 15 min at 4 °C and subsequently filtered through a 0.22 μm pore-size membrane filter. A 10 μL aliquot of each sample was injected into an Aminex® HPX-87H column (300 mm × 7.8 mm), coupled with a guard column (Bio-Rad Laboratories, Rio de Janeiro, Brazil). The mobile phase consisted of 0.005 mol/L sulfuric acid (H₂SO_4_) in ultrapure water, delivered at a constant flow rate of 0.7 mL/min. The column was maintained at 45 °C, and the total run time for each analysis was 30 min. Chromatographic data were acquired and processed using LabSolutions software (Shimadzu Corporation, 2013). Standards of lactic, citric, succinic, formic, acetic, propionic, isobutyric, butyric, isovaleric, and valeric acids were used to identify the chromatogram peaks, compare the retention times, and calculate their concentration in the samples. One sample of each extract was analyzed in duplicate.
Cell leakage assay
2.6
Cell leakage assessment was performed following Sulistyani et al. (2023) and Wang and Zeng (2022), with adaptations. Ten milliliters of bacterial suspensions (10^8^ CFU/mL) of P. paracarnis A006, P. fluorescens 07A, E. coli 1791, E. coli O157: H7, L. monocytogenes ATCC 19117, S. aureus ATCC 19095, and S. typhimurium ATCC 14028 were centrifuged for 20 min at 4,500 rpm. The pellet was collected and washed three times with PBS (pH 7.4), then resuspended in LAB-derived extracts that demonstrated bactericidal activity against at least one contaminant at 1 × MBC. The mixture was incubated as described in Table 1 for 24 h. Subsequently, the samples were centrifuged for 20 min at 4,500 rpm, and the absorbance of the supernatant was measured at 260 and 280 nm to detect nucleic acids (DNA and RNA) using a Shimadzu UV-1800 UV–Vis Spectrophotometer (Shimadzu, Japan). DNA and RNA concentrations were calculated according to Equations 3 and 4, respectively. The LAB-derived extracts from four strains exhibiting the highest potential for cellular damage, leading to nucleic acid leakage, were selected for the time-kill assay and scanning electron microscopy (SEM) imaging. The experiments were carried out in triplicate with two independent biological replicates.
Time-kill assay
2.7
The time-kill assay was conducted following the method described for Sulistyani et al. (2023). The contaminants P. fluorescens 07A and L. monocytogenes ATCC 19117 were selected as target microorganisms. BHI broth containing both contaminants at 10^5^ CFU/mL was mixed with the LAB-derived extract at a concentration of 1×MBC, and then incubated as mentioned in Table 1 for 0, 2, 4 and 6 h. An aliquot of 20 μL of each sample was collected and inoculated with a pour plate method using BHI agar and then incubated at 37 °C for 18–24 h. The colonies were counted and then time-kill curves were constructed by plotting log_10_ CFU/mL against time (h). The experiments were carried out in triplicate with two independent biological replicates.
SEM analysis
2.8
The morphological changes in planktonic cells of P. fluorescens 07A and L. monocytogenes ATCC 19117 treated with CFS and ICS were evaluated by SEM, as previously described by Jiang et al. (2022). Initially, 1 mL of L. monocytogenes ATCC 19117 and P. fluorescens 07A cell suspension (10^7^ CFU/mL) was centrifuged at 6,000 rpm for 5 min at 4 °C, followed by three washing steps with PBS. The recovered cells were then treated with the LAB-derived extracts at 1 × MBC and incubated following the conditions described in Table 1 for 4 h. After treatment, the cell suspension was centrifuged again, washed three times with PBS and fixed on brass stubs with 2.5% glutaraldehyde at 4 °C for 8 h. The samples were dried in a desiccator at room temperature for 7 days (no dehydration step was performed), sputter-coated with gold (Quorum—Q150TES Plus, East Sussex, England), and examined using a scanning electron microscope (JSM-IT700HR, JEOL, Tokyo, Japan). Untreated cells were used as controls.
Statistical analysis
2.9
Statistical analyses and data visualization were performed using R software version 4.3.3 and RStudio 2023.03.0 + 386 (RStudio Team, 2023; R Core Team, 2024). The Shapiro–Wilk test and Bartlett’s test were used to assess the normality of residuals and the homogeneity of variances, respectively. For the antagonism assays, an analysis of variance (ANOVA) was performed for each of the seven target microorganisms within each LAB studied, considering the six evaluated treatments. Tukey’s test was applied as a post hoc procedure. For the time-kill assay, statistical analysis was conducted using the Kruskal–Wallis test followed by Conover’s test, whereas cell leakage data were analyzed using ANOVA and Tukey’s test. Statistical significance was set at p < 0.05.
Results
3
Antagonistic effects and prevention of biofilm formation
3.1
Antibacterial activity of fifteen LAB strains against seven food-related spoilage and pathogenic microorganisms was initially assessed using agar diffusion assays. L. monocytogenes ATCC 19117 and P. fluorescens showed the largest inhibition zones (Figure 1), with inhibition patterns varying among strains.
Radar chart illustrating the inhibition halo diameters (mm) generated by Lpb. plantarum Q4C3, Lc. lactis subsp. lactis biovar diacetylactis SBR4, Lc. lactis subsp. lactis Lc08, W. viridescens W23, W. confusa W8, W. paramesenteroides W11, W. paramesenteroides W10, W. cibaria W42, W. cibaria W32, W. cibaria W49, W. cibaria W21, W. cibaria W22, W. cibaria W25, Lc. lactis subsp. lactis biovar diacetylactis ATCC 13675, Lc. lactis subsp. lactis ATCC 19435 against the contaminants: Salmonella Typhimurium ATCC 14028, L. monocytogenes ATCC 19117, S. aureus ATCC 19095, E. coli O157: H7, P. paracarnis A006, P. fluorescens 07A, E. coli 1791, after 24 h of incubation in the agar diffusion assay. Each value represents the mean of triplicate.
Following the initial screening by agar diffusion assays, the antimicrobial potential of LAB-derived extracts was further assessed through growth inhibition and antibiofilm assays. Table 2 presents the pH values of the LAB-derived extracts. The pH ranged from 3.84 to 4.46 for CFS, 6.96 to 7.18 for CFS N, 3.87 to 4.61 for ICS, 6.65 to 7.15 for ICS N, and 6.02 to 6.37 for IC.
After 24 h of incubation, the LAB-derived extracts from Lc. lactis Lc. lactissubsp. Lactis biovar diacetylactis ATCC 13675, Lc. lactis Lc. lactissubsp. Lactis Lc08, Lc. lactis Lc. lactissubsp. Lactis ATCC 19435, W. cibaria W22, and Lpb. plantarum Q4C3 exhibited the highest inhibition of microbial growth (Figure 2A). For biofilm inhibition (Figure 2B), the most effective extracts were those derived from Lc. lactis subsp. lactis biovar diacetylactis SBR4, Lc. lactis subsp. lactis Lc08, W. cibaria W21, W. cibaria W22, and W. viridescens W23.
Heatmap of growth inhibition (%) (A: OD600) and biofilm inhibition (%) (B: OD590) of P. paracarnis A006, P. fluorescens 07A, E. coli 1791, E. coli O157: H7, L. monocytogenes ATCC 19117, S. aureus ATCC 19095, and S. typhimurium ATCC 14028 in response to six treatments: CFS, Cell-free supernatants; ICS, inactive cells plus cell-free supernatants; CFS N, neutralized cell free supernatant; ICS N, neutralized inactive cells plus cell-free supernatant, and IC, inactive cells, prepared from Lpb. plantarum Q4C3, Lc. lactis subsp. lactis biovar diacetylactis SBR4, Lc. lactis subsp. lactis Lc08, W. viridescens W23, W. confusa W8, W. paramesenteroides W11, W. paramesenteroides W10, W. cibaria W42, W. cibaria W32, W. cibaria W49, W. cibaria W21, W. cibaria W22, W. cibaria W25, Lc. lactis subsp. lactis biovar diacetylactis ATCC 13675, Lc. lactis subsp. lactis ATCC 19435. The colors of the heatmaps agree with the percent inhibition. Letters indicate statistically significant differences between treatments (columns) in each lactic acid bacteria, based on Tukey’s test (p < 0.05). Tree densities represent hierarchical clustering applying the Euclidean distance matrix calculation and the Ward D2 method.
Z-score analysis (Supplementary material) identified Lpb. plantarum Q4C3, Lc. lactis subsp. lactis biovar diacetylactis ATCC 13675, Lc. lactis subsp. lactis ATCC 19435, Lc. lactis subsp. lactis Lc08, and W. cibaria W21 as the strains with the most consistent inhibitory performance across assays. This standardized comparison also indicated that CFS and ICS were the most effective extract types overall.
Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC)
3.2
Figure 3 shows the MIC values of the LAB-derived extracts against representative foodborne pathogens and spoilage microorganisms. CFS and ICS exhibited the lowest MIC values (25–100%), depending on the strain and target species. Lower MIC values (25–50%) were most frequently observed for extracts from Lpb. plantarum Q4C3, Lc. lactis subsp. lactis Lc08, and W. cibaria W21. In contrast, neutralized extracts (CFS N and ICS N) displayed reduced or no inhibitory activity (WI) for most target microorganisms. Only extracts from W. cibaria W21, W. cibaria W22, and W. viridescens W23 retained partial inhibitory capacity after neutralization. Among the evaluated extracts, only CFS and ICS displayed bactericidal activity (Figure 4). These treatments were therefore selected for subsequent cell leakage, time–kill, and SEM analyses.
Minimum inhibitory concentration (MIC) values of LAB extracts in percentage determined against foodborne pathogens and spoilage microorganisms. CFS, Cell-free supernatants; ICS, inactive cells plus cell-free supernatants; CFS N, neutralized cell free supernatant; ICS N, neutralized inactive cells plus cell-free supernatant; IC, inactive cells and WI, without inhibition. Lactococcus lactis subsp. lactis biovar diacetylactis ATCC 13675, Lactococcus lactis subsp. lactis ATCC 19435, Lactplantibacillus plantarum Q4C3, Lactococcus lactis subsp. lactis biovar diacetylactis SBR4, Lactococcus lactis subsp. lactis Lc08, Weissela cibaria W21, Weissela cibaria W22, Weissella viridescens W23.
Minimum bactericidal concentration (MBC) values determined against foodborne pathogens and spoilage microorganisms. CFS, Cell-free supernatants; ICS, inactive cells plus cell-free supernatants; CFS N, neutralized cell free supernatant; ICS N, neutralized inactive cells plus cell-free supernatant; IC, inactive cells, and NB, Non-bactericida Lactococcus lactis subsp. lactis biovar diacetylactis ATCC 13675, Lactococcus lactis subsp. lactis ATCC 19435, Lactplantibacillus plantarum Q4C3, Lactococcus lactis subsp. lactis biovar diacetylactis SBR4, Lactococcus lactis subsp. lactis Lc08, Weissela cibaria W21, Weissela cibaria W22, Weissella viridescens W23.
Cell leakage
3.3
Cell leakage analysis revealed that CFS treatments resulted in higher extracellular nucleic acid levels than ICS treatments across all evaluated strains (Figure 5). Among the extracts, those from Lc. lactis subsp. lactis biovar diacetylactis SBR4, Lpb. plantarum Q4C3, W. cibaria W21, and W. viridescens W23 produced the highest leakage values.
Bar chart illustrating cellular leakage in foodborne contaminants, quantified through DNA and RNA release after treatment with extracts of Lc. lactis subsp. lactis biovar diacetylactis ATCC 13675, Lc. lactis subsp. lactis ATCC 19435, Lc. lactis subsp. lactis Lc08, Lpb. plantarum Q4C3, Lc. lactis subsp. lactis biovar diacetylactis SBR4, W. cibaria W21, W. cibaria W22, and W. viridescens W23. The letters indicate statistically significant differences between each lactic acid bacteria, based on Tukey’s test (p < 0.05).
Time-kill assay
3.4
For time–kill assessment, P. fluorescens 07A and L. monocytogenes were selected as representative Gram-negative and Gram-positive species. Extracts from the four LAB strains that produced the highest leakage values were included in this assay.
CFS treatments resulted in reductions of more than 3 log CFU/mL within 4 h, while ICS tretments produced reductions of approximately 2 log CFU/mL over the same time (Figure 6). For CFS, no viable cells were detected after 6 h. For ICS, viable P. fluorescens 07A cells were detected at 4 h, followed by a marked reduction at 6 h. Therefore, 4 h was selected as the contact time for ICS and CFS treatments for next assays.
Time–kill curves of P. fluorescens 07A (A: CFS, C: ICS) and L. monocytogenes ATCC19117 (B: CFS, D: ICS) treated with four LAB-derived extracts (Lpb. plantarum Q4C3, Lc. lactis subsp. lactis biovar diacetylactis SBR4, Lc. lactis subsp. lactis Lc08, W. cibaria W21, W. viridescens W23) compared with the untreated control. Panels A and B correspond to cell-free supernatants (CFS), while panels C and D correspond to inactive cells plus cell-free supernatant (ICS). Bacterial counts (log₁₀ CFU/mL) were determined at 0, 2, 4, and 6 h of incubation. The asterisks indicate statistically significant differences between each lactic acid bacteria, based on Tukey’s test (p < 0.05).
Organic acid quantification assay
3.5
The organic acid profile varied considerably among the eight tested LAB-derived extracts (Table 3). Detected organic acids included lactic, formic, acetic, citric, succinic, propionic, isobutyric, butyric, isovaleric, and valeric acids. Lc. lactis subsp. lactis biovar diacetylactis ATCC 13675 did not produce formic acid and showed the highest acetic acid levels. This strain and Lpb. plantarum Q4C3 did not produce citric acid. Isovaleric acid was detected only in Lc. lactis subsp. lactis biovar diacetylactis SBR4 and Lc. lactis subsp. lactis Lc08. Acid concentrations were lower in ICS compared with CFS for all strains.
SEM
3.6
SEM images revealed structural alterations in bacterial cells following exposure to LAB-derived extracts (Figure 7). For P. fluorescens 07A, CFS-treated cells showed more extensive deformation than ICS-treated cells. Similar patterns were observed for L. monocytogenes. Control cells of both species maintained intact morphology, whereas treated cells exhibited varied degrees of surface irregularity, including depressions and distortions. Morphological changes were consistently more pronounced in cells treated with CFS compared with ICS.
SEM images of Pseudomonas fluorescens 07A cells (A: Control, B, C, D, E, F, G, H, I) and Listeria monocytogenes ATCC 19117 cells (a: Control, b, c, d, e, f, g, h, i) treated with LAB-derived extract at 1×MBC from Lpb. plantarum Q4C3 (B/b: CFS; C/c: ICS), Lc. lactis subsp. lactis biovar diacetylactis SBR4 (D/d: CFS; E/e: ICS), W. cibaria W21 (F/f: CFS; G/g: ICS) and W. viridescens 23 derived extracts (H/h: CFS; I/i: ICS) for 4 h. The red arrows indicate irregular and collapsed surfaces, fissures, and deformations, which are indicative of membrane damage and possible cell lysis.
Discussion
4
In this study, LAB-derived extracts exhibited marked antimicrobial and antibiofilm activities against foodborne pathogens and spoilage microorganisms relevant to dairy systems. However, the magnitude and spectrum of inhibition were clearly strain dependent. In agar diffusion assay, L. monocytogenes ATCC 19117 and P. fluorescens were among the most susceptible targets, but inhibition patterns varied considerably across the 15 LAB strains (Figure 1), indicating that antagonistic effects are strain specific rather than uniform at the species level. The inhibition observed in agar assay reflects the direct interactions with the solid medium, where metabolite diffusion and localized competition play a key role, and may differ from liquid co-culture systems, in which nutrient availability, mixing, and metabolite dispersion influence the extent of antagonism (Selegato and Castro-Gamboa, 2023).
A consistent finding across assays was the superior performance of non-neutralized extracts (CFS and ICS) compared with their neutralized counterparts (CFS-N and ICS-N), both in terms of MIC values and antibiofilm activity. This pattern supports the central role of organic acids in the antimicrobial action of LAB-derived extracts. Partial or total loss of activity, following pH neutralization, as observed for several strains, reinforces this interpretation and agrees with reports in which non-neutralized supernatants from Lpb. plantarum exhibited strong antibacterial activity that was abolished upon neutralization (Grigore-Gurgu et al., 2024). In this study, Lpb. plantarum Q4C3 stood out not only for its antibacterial potential but also corroborates previous work in which this strain ranked among the most effective LAB against filamentous fungi such as Aspergillus niger IOC 207 and Penicillium chrysogenum IOC 132 (Souza et al., 2023), highlighting its versatility as a biocontrol agent.
The organic acid profiles obtained for selected strains help to explain these observer results in this studye observations. Lactic, acetic and propionic acids were detected in the highest concentrations, with additional contributions from formic, citric, succinic, butyric and valeric acids. The transition from CFS to ICS was accompanied by a general decrease in organic acid concentrations and a subtle increase in pH, suggesting partial neutralization of acids due to the release of intracellular fluid following cell lysis. Weak acids display greater antimicrobial activity at low pH, and acetic and propionic acids are typically more potent inhibitors than lactic acid because of their higher pK_a_ values, which favor a higher proportion of undissociated molecules in acidic environments (Lahtinen et al., 2011; Wang et al., 2014; Ray and Sandine, 2019). In mixed acid systems, lactic acid contributes predominantly to pH reduction, whereas acetic and propionic acids act as the main antimicrobial components, often exhibiting synergistic effects (Zavišić et al., 2024). The composition and concentration of these acids in CFS and ICS therefore strongly influence the observed inhibitory profiles.
The biological relevance of the organic acid profiles obtained by HPLC can be further interpreted by comparison with inhibitory concentrations reported in food-relevant conditions. Wemmenhove et al. (2016) determined the MIC of undissociated lactic, acetic, citric, and propionic acids against L. monocytogenes at cheese-relevant pH conditions (5.2–5.6), reporting average MIC of approximately 5 mM for lactic acid, 19 mM for acetic acid, and 11 mM for propionic acid. These values overlap with the organic acid concentrations detected in the most active LAB-derived extracts evaluated in the present study, supporting the biological plausibility of the observed antimicrobial effects in dairy matrices. In addition, salt content and other intrinsic factors in cheese matrices may further reduce the effective MICs of organic acids, thereby enhancing their inhibitory potential.
However, in complex food environments such as the moisture phase of cheeses, dilution effects and matrix buffering can decrease the proportion of undissociated acids available to exert antimicrobial activity. Factors including pH, water activity, ionic strength, and the presence of competitive microflora can modulate pathogen inhibition and should be considered when extrapolating in vitro efficacy to real food systems (Engstrom et al., 2020; Falih et al., 2024). Together, these considerations indicate that while organic acid concentrations measured in LAB-derived extracts are within inhibitory ranges reported for dairy-relevant conditions, their effective antimicrobial performance in foods will depend on matrix-specific interactions.
Although pH-neutralization experiments provide valuable insight into acid-mediated antimicrobial mechanisms, they do not exclude the contribution of pH-sensitive bacteriocins or other metabolites. Several antimicrobial peptides produced by LAB are known to be more active and stable under acidic conditions, while exhibiting reduced activity or structural destabilization at neutral or alkaline pH due to changes in ionization state and partial peptide denaturation (Liang et al., 2025). A similar pH-dependent loss of activity has been reported for nisin, which shows significantly reduced inhibitory effects as pH increases (Zhang et al., 2016). Therefore, the reduction in antimicrobial efficacy observed after neutralization may reflect not only the loss of organic acid activity but also diminished functionality of bacteriocins and other acid-dependent metabolites.
Consistent with this interpretation, residual antimicrobial activity detected in some neutralized extracts indicates the contribution of additional acid-independent metabolites. In the present study, Lc. lactis subsp. lactis Lc08 retained antimicrobial activity after neutralization, in agreement with previous reports describing the production of antagonistic compounds that remain stable over a wide pH range and are only inactivated after autoclaving (Perin et al., 2013), in this study retained antimicrobial activity after neutralization, consistent with the presence of acid-independent compounds. Similarly, Lc. lactis subsp. lactis bv. diacetylactis SBR4, characterized as a nisin Z producer with high solubility and diffusibility at neutral pH (Fusieger et al., 2020b), showed consistent behavior with its known characteristics. Together, these results support a model in which organic acids provide a basal antimicrobial effect that can be further enhanced by bacteriocins, in line with previous evidence that the co-application of acids and bacteriocins results in synergistic inhibition of pathogens and spoilers (Yu et al., 2023).
Weissella strains also emerged as promising candidates for biocontrol in dairy environments. In agreement with previous studies showing that W. cibaria W21 and related strains inhibit a range of foodborne (Dey et al., 2019; Teixeira et al., 2021), this results demonstrate that W. cibaria W21, W. cibaria W22 and W. viridescens W23 combine relevant antimicrobial and antibiofilm activities. The fact that some Weissella-derived extracts retained partial activity after neutralization suggests the production of additional bioactive metabolites beyond organic acids, reinforcing the potential of this genus as a source of bacteriocins and extracts preparations.
Although Weissella strains have demonstrated relevant antimicrobial and antibiofilm potential, their application as live cultures should be approached with caution. This genus lacks Qualified Presumption of Safety (QPS) status according to the EFSA and GRAS recognition by the FDA, and safety has not been established for all strains (FDA, 2024; EFSA, 2026). This regulatory limitation highlights the need for strain-specific safety assessments prior to industrial use. In this context, the application of cell-free extracts derived from Weissella represents a safer alternative, allowing the exploitation of bioactive metabolites such as organic acids and bacteriocins, without the risks associated with administering live microrganisms (Fusco et al., 2023). The use of Weissella derived cell-free extracts or LAB-derived extracts strain-level antagonistic evaluation and supports the development of natural, safe, and industrially applicable biocontrol strategies.
Beyond safety considerations, an additional layer of complexity in biocontrol strategies is that antimicrobial efficacy against planktonic cells does not necessarily translate into effective biofilm inhibition. This discrepancy indicates that biofilm inhibition cannot be ascribed solely to reductions in viable cell numbers. As highlighted by Sharma et al. (2023), surface adhesion represents only the initial step of biofilm development; and compounds that directly interfere with exopolysaccharide synthesis, motility, or quorum sensing can markedly alter biofilm architecture. In this context, LAB-derived metabolites, such as hydrogen peroxide, oxygenated compounds, exopolysaccharides, bacteriocins, and fatty acids with surfactant properties, have been identified as key antibiofilm agents (Moradi et al., 2019; Rather et al., 2021). In this study, LAB-derived extracts exhibited strain-dependent antimicrobial and antibiofilm activities against both spoilage and pathogenic microorganisms, highlighting the diversity of antagonistic mechanisms. Z-score-based ranking and growth and biofilm inhibition data (Figure 2) confirmed that only a subset of LAB strains combines consistently high performance across both phenotypes, underscoring the need for strain-level selection when designing biocontrol strategies.
Within this context, the physical composition of LAB-derived preparations, particularly the presence of cellular debris in ICS, emerges as a critical factor influencing antimicrobial and antibiofilm performance. The presence of cellular debris in some treatments was associated with a reduction in antimicrobial potential, particularly for ICS. One plausible explanation is that cell remnants, extracellular polymeric substances, proteins, lipids, and nucleic acids may form a physical barrier around target cells, partially shielding them from diffusible antimicrobial metabolites (Zhao et al., 2017). Similar mechanisms have been proposed to explain decreased antimicrobial activity in non-filtered preparations or in systems with high organic load (Ostad et al., 2009). Conversely, other studies suggest that inactivated LAB cells can reduce pathogen adhesion by forming a physical layer on the surface, thereby limiting colonization (Singh et al., 2017; Zielińska et al., 2021). These apparently conflicting observations highlight that the role of cellular debris is context dependent and influenced by factors such as matrix composition, microbial species and the relative contribution of diffusible versus cell-associated components. In our experimental conditions, filtration and removal of cellular remnants improved antimicrobial performance, which is consistent with the higher activity observed for CFS compared with ICS. In pratical applications, the behavior of LAB-derived extracts in real food systems is influenced by the surrounding matrix. Organic matter can modulate acid efficacy, with some components reducing and others enhancing antimicrobial effects, depending on the type of acid and the food environment (Guan and Liu, 2020; Lund et al., 2020).
In addition to matrix-related effects, intrinsic structural differences between target microorganisms also influence the efficacy of LAB-derived extracts. A notable outcome was the ability of CFS to inhibit both Gram-positive and Gram-negative bacteria. While bacteriocins are generally more active against Gram-positive organisms because of the protective outer membrane in Gram-negative bacteria (Darbandi et al., 2022), the combination of organic acids and other metabolites present in CFS may compromise this barrier by increasing membrane permeability. Previous studies have shown that organic acids and CFS from LAB can induce leakage of K^+^ ions, nucleic acids and proteins, reflecting direct damage to the cytoplasmic membrane (Bajpai et al., 2016; Sun et al., 2024; Wang et al., 2023). MIC and time–kill data, together with leakage assays, support a mechanism in which acids and associated metabolites facilitate access of bioactive compounds to the cell membrane, broadening the inhibitory spectrum to include Gram-negative spoilers such as Pseudomonas spp.
Cell leakage and SEM analyses provided further insight into the mode of action of LAB-derived extracts. CFS treatments consistently resulted in higher extracellular nucleic acid levels than ICS, and SEM images revealed pronounced morphological alterations in P. fluorescens and L. monocytogenes cells exposed to CFS, including surface collapse, wrinkling and structural distortion. These findings are in line with previous studies reporting membrane damage, cell swelling, vacuole formation and leakage of intracellular components in bacteria treated with LAB CFS (Abou Elez et al., 2023; Wang et al., 2023; Sun et al., 2024). Taken together, these observations support the notion that LAB-derived metabolites interact with membrane phospholipids and proteins, alter membrane fluidity, promote pore formation and ultimately lead to leakage and cell death (Anumudu et al., 2024; Che et al., 2024; Rahman et al., 2025; Banicod et al., 2025).
The membrane damage and leakage effects observed by SEM and nucleic acid release assays are further reflected in the rapid bactericidal kinetics revealed by the time–kill experiments. The time-kill assays highlight not only the bactericidal capacity of the extracts but also the speed of their action. For L. monocytogenes, CFS caused rapid reductions in viable counts, whereas P. fluorescens required slightly longer exposure for comparable effects. ICS required longer contact times than CFS in both cases. When compared with previous work in which LAB CFS required longer exposures or higher multiples of the MIC to achieve modest log reductions (Kho et al., 2024; Rotta et al., 2025; Liang et al., 2025), the extracts evaluated here displayed faster and more pronounced bactericidal activity. This enhanced efficacy may reflect the combined effects of a favorable organic acid profile and additional diffusible metabolites, as suggested by both the MIC/MBC results and the leakage and SEM data.
Conclusion
5
This present work demonstrated that LAB-derived extracts, particularly CFS from Lpb. plantarum Q4C3, Lc. lactis subsp. lactis biovar diacetylactis SBR4, W. cibaria W21, and W. viridescens W23, combine rapid bactericidal activity, antibiofilm effects, and a broad inhibitory spectrum against dairy-related contaminants. By systematically comparing multiple LAB strains and extract types, this study identified the most effective strain and extract combinations for potential industrial application. These findings support the use of these extracts as natural, fast-acting biocontrol agents to improve the microbial safety and quality of dairy products, while also providing a framework for future studies in real food matrices and pilot-scale applications.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abou Elez R. M. M. Elsohaby I. al-Mohammadi A. R. Seliem M. Tahoun A. B. M. B. Abousaty A. I. . (2023). Antibacterial and anti-biofilm activities of probiotic Lactobacillus plantarum against Listeria monocytogenes isolated from milk, chicken and pregnant women. Front. Microbiol. 14:1201201. doi: 10.3389/fmicb.2023.1201201, 37538844 PMC 10394229 · doi ↗ · pubmed ↗
- 2Anumudu C. K. Miri T. Onyeaka H. (2024). Multifunctional applications of lactic acid bacteria: enhancing the safety, quality, and nutritional value of fermented foods and beverages. Foods 13:3714. doi: 10.3390/foods 1323371439682785 PMC 11640447 · doi ↗ · pubmed ↗
- 3Arena M. P. Silvain A. Normanno G. Grieco F. Drider D. Spano G. . (2016). Use of Lactobacillus plantarum strains as a bio-control strategy against food-borne pathogenic microorganisms. Front. Microbiol. 7:464. doi: 10.3389/fmicb.2016.00464, 27148172 PMC 4829616 · doi ↗ · pubmed ↗
- 4Ashrafudoulla M. Park J. Toushik S. H. Shaila S. Ha A. J.-w. Rahman M. A. . (2024). Synergistic mechanism of UV-C and postbiotic of Leuconostoc mesenteroides (J.27) combination to eradicate Salmonella Thompson biofilm in the poultry industry. Food Control 164:110607. doi: 10.1016/j.foodcont.2024.110607 · doi ↗
- 5Bajpai V. K. Han J. H. Rather I. A. Park C. Lim J. Paek W. K. . (2016). Characterization and antibacterial potential of lactic acid bacterium Pediococcus pentosaceus 4I 1 isolated from freshwater fish Zacco koreanus. Front. Microbiol. 7:2037. doi: 10.3389/fmicb.2016.02037, 28066360 PMC 5167689 · doi ↗ · pubmed ↗
- 6Banicod R. J. S. Tabassum N. Javaid A. Kim Y. M. Khan F. (2025). Lactic acid bacteria–derived secondary metabolites: emerging natural alternatives for food preservation. Probiotics Antimicrob. Proteins, 1–38. doi: 10.1007/s 12602-025-10672-640751876 · doi ↗ · pubmed ↗
- 7Che J. Shi J. Fang C. Zeng X. Wu Z. Du Q. . (2024). Elimination of pathogen biofilms via postbiotics from lactic acid bacteria: a promising method in food and biomedicine. Microorganisms 12:704. doi: 10.3390/microorganisms 12040704, 38674648 PMC 11051744 · doi ↗ · pubmed ↗
- 8Choi D. Bedale W. Chetty S. Yu J. H. (2024). Comprehensive review of clean-label antimicrobials used in dairy products. Compr. Rev. Food Sci. Food Saf. 23:e 13263. doi: 10.1111/1541-4337.13263, 38284580 · doi ↗ · pubmed ↗