Resistance Training Reshapes the Gut Microbiome in a Longitudinal 8-Week Intervention in Sedentary Adults
Daniel Straub, Till Englert, Antonia Beller, Josua Stadelmaier, Mark Stahl, Joachim Kilian, Jens Borzym, Carola Rotermund, Tanja Akbuğa-Schön, Sabrina Krakau, Stefan Czemmel, Sabine Weiler, Marc Pettenkofer, Jörg Pettenkofer, Ulli Maser, Sascha Dammeier, Andreas M. Nieß

TL;DR
Resistance training can change the gut microbiome, especially in people who gain more strength, leading to healthier bacteria linked to anti-inflammation and better gut function.
Contribution
This study shows resistance training induces time-dependent gut microbiome changes, particularly in individuals with significant strength gains.
Findings
Resistance training caused significant microbial shifts in participants with ≥33% strength gains.
Faecalibacterium and Roseburia hominis, linked to health benefits, were enriched in high responders.
Microbial changes correlated with strength improvement, suggesting a link between gut health and physical fitness.
Abstract
The gut microbiome plays a critical role in metabolism, immunity, and aging. While endurance training has been shown to beneficially modulate the microbiome, the effects of resistance training remain less clear, with some studies reporting minimal changes. This project aims to investigate whether structured resistance training elicits significant changes in gut microbiome composition and diversity in sedentary, healthy adults. 150 participants (85 female, 63 male), between 24 and 61 years of age, completed an 8-week supervised resistance training program between May 2022 and July 2023 in the cities of Tübingen and Rottenburg, Germany. Session-level training data, including weights and repetitions, were recorded alongside metrics like load and compliance. Fecal samples were collected throughout the study period at designated timepoints for 16S rRNA gene amplicon sequencing to assess…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
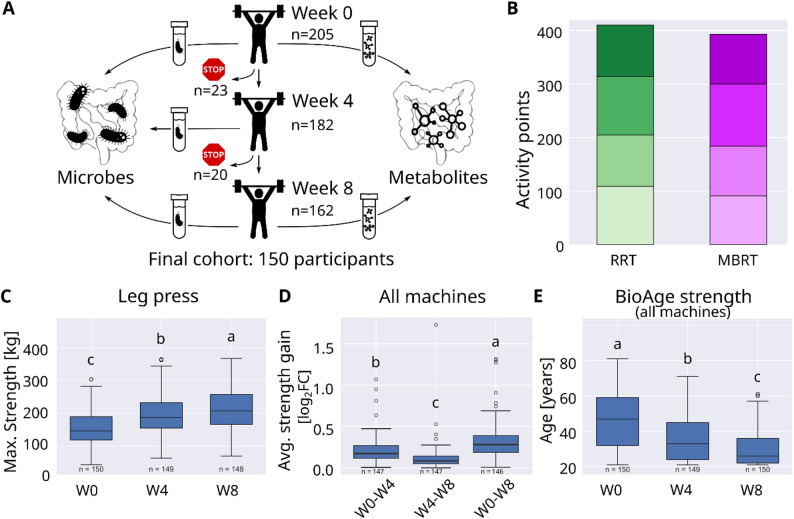 Figure 1
Figure 1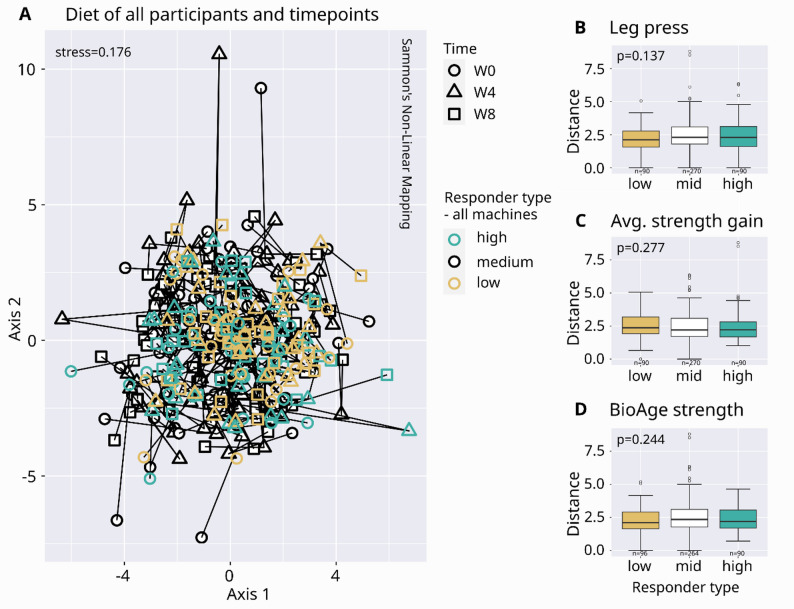 Figure 2
Figure 2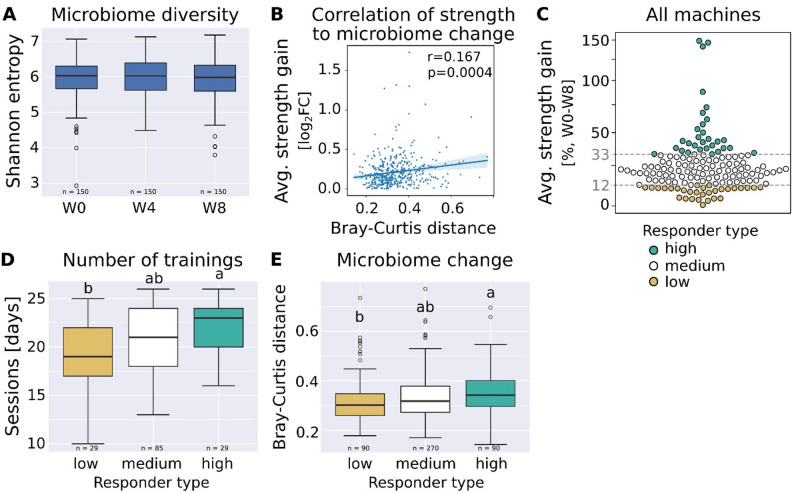 Figure 3
Figure 3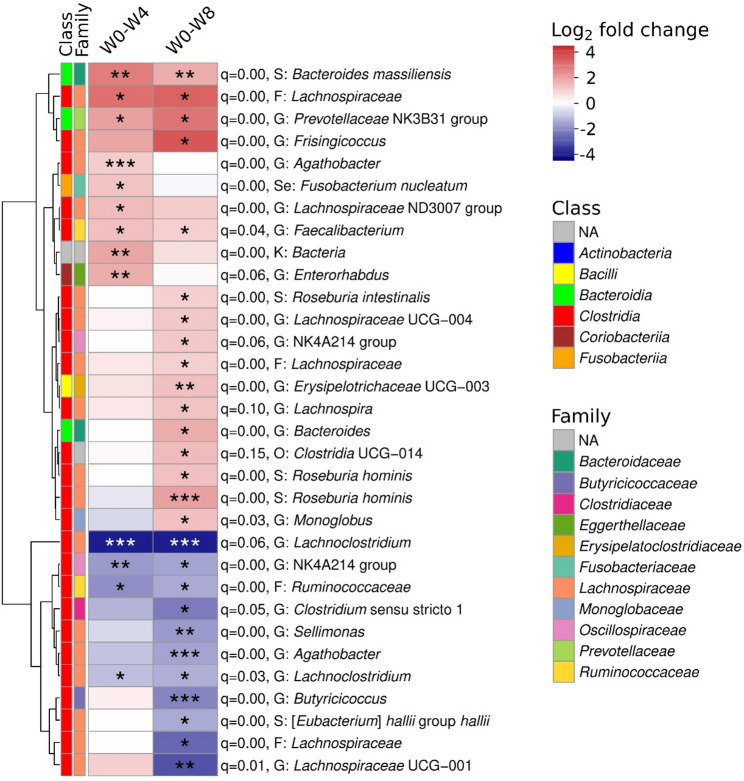 Figure 4
Figure 4- —Eberhard Karls Universität Tübingen (1020)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Muscle metabolism and nutrition · Metabolomics and Mass Spectrometry Studies
Introduction
The human gut microbiome, comprising trillions of microorganisms, plays a central role in digestion, metabolism, immune function, and overall health [1]. While diet and lifestyle are well-established modulators, growing evidence points to physical activity as a significant influence on gut microbial diversity and composition. Exercise affects key physiological processes such as inflammation and energy metabolism, which are closely linked to the gut microbiome and broader outcomes like physical fitness and aging [2].
The microbiome is increasingly recognized as a marker not only of overall health but also of aging [3, 4]. Age-related declines in microbial diversity and beneficial taxa are associated with inflammation, immune dysfunction, and metabolic deterioration [4]. In contrast, a microbiome enriched in bacteria such as Faecalibacterium, Akkermansia, and Roseburia, known for producing short-chain fatty acids (SCFAs) like butyrate, is linked to improved gut integrity, reduced inflammation, and healthier aging [5]. Endurance training has been consistently shown to promote such a microbial profile, increasing alpha diversity, SCFA production, and gut barrier function [2].
In contrast, the effects of resistance training on the gut microbiome are far less understood. Resistance exercise induces distinct physiological changes, including muscle hypertrophy, increased protein turnover, glycemic control [6] and hormonal adaptations [7]. However, current research offers conflicting findings: while some studies report microbiome shifts correlated with strength performance, others find only minimal or no changes [8, 9].
Given the inconsistent findings to date, larger and more detailed studies are needed to determine whether resistance training alone can induce measurable changes in the gut microbiome—particularly changes resembling the health-associated patterns seen with endurance training. To address this gap, we investigated microbiome shifts in a large cohort of previously inactive, healthy adults undergoing a structured resistance training program. Uniquely, we captured detailed session-level data (weights lifted, repetitions performed) to objectively assess training load and adherence. By linking these metrics with longitudinal microbiome and fitness data, this study offers the most comprehensive analysis to date of how resistance training may influence gut microbial diversity and taxa associated with metabolic and immune health.
Materials and Methods
Training Protocol and Exclusion Criteria
The intervention took place from May 2022 to July 2023 at two “Fitness- und Gesundheitsclub Mapet” locations in Tübingen and Rottenburg, Germany. Sedentary individuals (≥ 1 year) were screened and enrolled after completing a demographic questionnaire. Participants completed a baseline fitness assessment comprising anthropometric measurements (body weight, body mass index, and body fat percentage assessed via bioelectrical impedance analysis), cardiorespiratory evaluation (VO₂ max testing), and cardiovascular variables (resting blood pressure and heart rate). Subsequently, participants engaged in an 8-week supervised resistance training intervention (2–3 sessions per week). Follow-up fitness assessments were conducted at weeks 4 and 8. Participants completed pre-test questionnaires covering dietary and fluid intake, medication use and other relevant factors, and were instructed to maintain their usual dietary and fluid intake patterns throughout the study period (Suppl. File: Datenblatt Screening 2).
Training was conducted on seven EGYM Smart Strength machines (Seated row, Lat Pull, Chest press, Back trainer, Abdominal trainer, Leg curl, and Leg press) using one of two resistance training programs: a resistance training program aimed at improving general fitness, or muscle-building (Table 1). The equipment enabled standardized training through automated control of range of motion, movement speed, and digitally programmed resistance profiles, including eccentric overload phases and adaptive load adjustments based on user performance. These algorithm-driven protocols provided progressive, individualized stimuli not feasible with conventional gym equipment. Training machines not only guided each session in real time but also automatically recorded detailed exercise data (machine used, program, weight, repetitions, max strength, date) for every workout.
Table 1. Training protocol with EGYM resistance exercise machines and training algorithmsWarm-up10 minCardiovascular exercise machinesResistance training25 min2 sets of 7 EGYM Smart Strength machines with resistance training algorithmsMachines: Leg press, Leg curl, Back trainer, Abdominal trainer, Chest press, Seated Row, Lat PullResistance training algorithm for each training phase (a 2 weeks)GroupRegular resistance trainingMuscle building resistance training Phase 1Regular20 rep. at 45% of max. strengthNegative15 rep. at 38% concentric / 58% eccentric of max. strength Phase 2Negative15 rep. at 38% concentric / 58% eccentric of max. strengthAdaptive10 rep. starting at 68% of max. strength with automatic decrease Phase 3Regular20 rep. at 45% of max. strengthIsokinetic2 × 8 rep. starting at 73% concentric / 105% eccentric of max. strength with automatic decrease Phase 4Negative15 rep. at 38% concentric / 58% eccentric of max. strengthAdaptive10 rep. starting at 68% of max. strength with automatic decreaserep. = repetition
Of the 205 enrolled participants, 43 did not complete the full 8-week training protocol and were excluded from analysis. Six additional participants were excluded from the analysis due to data quality concerns (antibiotic use or improper stool sample storage). Another six participants were excluded as their samples were used for internal validation of sequencing and metabolomics workflows.
Strength and Training Metrics
Average strength gains were assessed by calculating the average log₂ fold change in maximum strength across all exercise machines between two time points.
BioAge Strength is an EGYM metric that estimates the biological age of muscular fitness based on a non-linear model of the typical relationship between age and strength. It uses a large dataset from approximately one million probands including chronological age, sex, and strength relative to body weight to determine whether an individual’s strength is above (younger BioAge) or below average (older BioAge) for their chronological age. The final score is calculated as the average across all exercise machines and can be improved by increasing strength or reducing body weight—but not below the minimum of 21 years.
Activity Points, another EGYM metric, were used to estimate the metabolic equivalent (MET) expended per session. This measure integrates training intensity, excess post-exercise oxygen consumption (EPOC), the mechanical work of each set, and an individualized estimate of the participant’s resting metabolic rate, providing a comprehensive representation of training load.
Training compliance was quantified as the percentage of targeted repetitions actually performed across all sessions, offering an objective measure of adherence to the training protocol.
Stool Sample Processing
Study participants were handed out self-sampling kits and instructed to sample stool 1–3 days before each fitness test. Stool was sampled at the start of the study (week 0), during the study (week 4) and at the end of the study (week 8) with OMNIgene^®^-GUT Kits OM-200 (DNA Genotek Inc., Ottawa, Canada) for 16S rRNA gene amplicon sequencing and at the start of the study (week 0) and at the end of the study (week 8) additionally with OMNImet^®^-GUT Kits ME-200 DNA Genotek Inc., Ottawa, Canada) for targeted metabolomics. Samples were stored at 4 °C before transport at ambient temperature and long-term storage at − 80 °C.
DNA was extracted using the DNeasy 96 PowerSoil Pro Kit and 16S rRNA V4 regions were amplified using primers 515 F/806R. Sequencing was performed on the Illumina MiSeq platform (2 × 250 bp, 69k reads/sample on average). Data processing, including quality control, reconstruction of sequences, taxonomic annotation, and diversity analysis, employed the nf-core/ampliseq pipeline (v2.7.0) [10] as detailed in the supplement. Essentially, 9,031 amplicon sequencing variants (ASVs) with more than 25 million counts were obtained across all samples. ASVs were taxonomically annotated with DADA2 v1.28.0 [11] and SILVA v138.1 [12] and diversity indices were determined with QIIME2 v2023.7.0 [13] after rarefaction.
For metabolomics, the targeted panel consisted of 34 compounds including short-chain fatty acids (SCFA), amino acids, bile acid derivates, choline metabolites, indole derivates, phenolic derivates, polyamines, and vitamins (see the supplment for details). Samples were prepared, analyzed and quantified via internal standards and dry weight normalization via LC-MS with Waters Acquity-SynaptG2 and GC-MS with Shimadzu TQ8040 as detailed in the supplement.
Statistical Analysis
Friedman rank sum test with unreplicated blocked data [14] and, in case of significance, was followed by Conover post hoc [15], to assess changes over time in fitness and survey data, because normal distribution was consistently rejected (Shapiro-Wilk test [16], p ≤ 0.05). Longitudinal trends between training groups were analyzed with non-parametric linear data analysis (nparLD v2.2 [17]) with spanova design (2 independent groups with 3 repeated measures) in R v4.3.3 [18]. Means of training days, fitness, and survey data were compared between training types with Kruskal-Wallis rank sum test followed by pairwise comparisons using Wilcoxon rank sum test with continuity correction and between reponder types using Dunn’s test of multiple comparisons using rank sums (R package “dunn.test” v1.3.6 [19]), respectively. Binary fitness and survey data data was compared with Pearson’s Chi-squared test in R. Kolmogorov-Smirnov test [20] was applied to distributions and Pearson correlation was applied to beta-diversity indices and strength improvements in R. Multiple testing correction was done with the Benjamini-Hochberg method [21]. Significance threshold was set at p ≤ 0.05 if not otherwise indicated.
Diet pattern changes within and between participants (food: fruit and vegetables, fish, meat, eggs, diary, grains, sweets, salty snacks; drinks: water and tea, coffee, juice and lemonade, alcohol in ml) were visualized with Sammon’s Non-Linear Mapping [22] based on euclidean distances of root-mean-square of centered values (R package MASS v7.3-60 [23]).
Differential microbial abundance was assessed with ANCOM-BC2 [24] using Dunnett’s test [25] with Holm’s family-wise error rate correction [26] based on phyloseq [27] objects from nf-core/ampliseq, modeling time point as a categorical variable. Extended information is available in the supplement.
HR and LR were predicted with Random Forest (scikit-learn v1.5.2 [28] in python v3.12.3 [29]) based on baseline fitness (strength and health data) and surveyed data (dietary intake, demographics, activity), or microbial abundances, or metabolite measurements. Key predictors were determined based on gini feature importance [30]. 1000 permutations determined one-sided p = 0.05 for the ROC-AUC score.
Results
Resistance Training Rapidly Enhances Muscle Strength
Study participants were predominately recruited through email-advertisement within the University of Tübingen, Germany, (40%) and press releases (32%) and no reimbursement was offered. The study participants were 24 to 61 years of age (42 years on average), of normal weight (BMI 16.9 to 34.2, average 24.5), sedentary (< 1 h of regular physical activity per week in the last 6 months), and healthy. Of the 205 participants initially enrolled, 150 were included in the final analysis (Fig. 1A). Table 2 summarizes the demographic and baseline characteristics of the final study cohort, more details in Suppl. Table 1, that was retrieved by a questionnaire (Suppl. File: Datenblatt Screening 2).
Table 2. Demographic data of participantsTraining groupAll participantsn = 150Regular resistance trainingn = 73Muscle-building resistance trainingn = 77p-valueGender (male)42.6%43.7%41.6%0.927^1^Age (years)41.7 ± 11.643.1 ± 11.440.5 ± 11.70.144^2^BMI (kg/m²)24.5 ± 3.524.7 ± 3.724.4 ± 3.40.683^2^Non-smoker76.7%78.1%75.3%0.827^1^Any allergy32.0%27.4%36.4%0.317^1^No special diet82.7%89.2%77.0%0.094^1^Born in Germany82.4%87.3%77.9%0.199^1^Values as percent or mean ± SD^1^: Pearson’s Chi-squared test^2^: Kruskal-Wallis rank sum test
Participants were randomly assigned to one of two training regimens: regular resistance training (higher repetitions, lower weights) or muscle-building resistance training (higher weights, fewer repetitions) (Table 2). The study initially aimed to compare fitness and microbiome responses between the two approaches. Despite differences in training style, both programs were similar in total weight moved and activity points (metabolic equivalent minutes) (Fig. 1B). Further, no significant differences in dietary intake (Dunn’s test or Pearson’s Chi-squared test, adjusted p > 0.35, Suppl. Table 1) or fitness improvements were observed between the groups (nparLD, adjusted p > 0.5). As a result, participants were pooled for all subsequent analyses to maximize statistical power.
Strength gains were evaluated using three metrics: (1) maximum strength leg press (kg), (2) average strength gain (average log₂ fold change across all machines), and (3) BioAge Strength (years). All measures showed substantial improvement within the first four weeks, followed by a smaller increase in the second half of the program. On average, leg press strength rose from 154 ± 54 kg to 217 ± 65 kg (Friedman test, adjusted p=3 × 10^− 45^, Fig. 1C), average strength increased by 24 ± 16% (Kruskal-Wallis rank sum test, p < 2.2 × 10^− 16^, Fig. 1D), and BioAge Strength decreased from 45.8 ± 14.7 to 29.6 ± 9.6 years (Friedman test, adjusted p=2 × 10^− 49^, Fig. 1E).
Fig. 1. Study participants, training and main performance metrics.** A** Study overview with number of participants enrolled in the study (n = 205 started, 150 in final cohort), with three fitness tests (week 0, week 4, week 8), three gut microbiome samples (left) and two gut metabolome samples (right) per participant. B Comparison of activity points (metabolic equivalents minutes) of regular resistance training (RRT) and muscle-building resistance training (MBRT).** C** Maximum strength of leg press over time.** D** Average strength gain over seven exercises.** E** Calculated BioAge Strength over time. Test tube symbols: self-sampling kit for microbiome or metabolites, stop symbols: study drop outs, log_2_FC: log_2_ fold change, W0: week 0, W4: week 4, W8: week 8. Letters denote p < 0.05
Beyond strength, participants also experienced statistically significant, though small, reductions in diastolic blood pressure (from 108 ± 17 to 105 ± 17 mmHg, Friedman test, adjusted p = 0.001) and body fat percentage (from 29.72 ± 8.01% to 29.34 ± 8.30%, Friedman test, adjusted p = 0.008). However, BMI, resting heart rate, and systolic blood pressure remained unchanged (Suppl. Figure 1). Importantly, the study participants appeared to follow the instruction to keep a stable diet and daily activity routine, because no significant nutritial or activity changes were found in the self-reported values over time over all participants (Firedman test, adjusted p > 0.15, Suppl. Table 2).
Changes in the Microbial Community is associated with Strength Gain
Resistance training did not significantly alter gut microbial alpha diversity, neither across all participants (Friedman test, p > 0.5, Fig. 3A and Suppl. Figure 2) nor within groups stratified by strength metrics. Similarly, no notable changes were observed in beta diversity across these groups, and stool metabolomic profiles remained unchanged (Suppl. Figure 3). Longitudinal within-subject analyses demonstrated that shifts in microbial community composition were modestly, yet significantly, associated with improvements in strength performance—most notably with average strength gains (Pearson correlation coefficient r = 0.167, p = 0.0004, Fig. 3B).
Based on this correlation, participants were stratified into high-responder (HR, top 20%) and low-responders (LR, bottom 20%) across three strength metrics (Fig. 3C), yielding three partially overlapping subsets (Suppl. Figure 4). Among these, the average strength gain subset showed the least demographic bias, while the BioAge Strength subset showed the most (Suppl. Figure 5). Noteably, all three HR subsets had with 43% to 50% regular resistance training a nearly balanced distribution of training types, excluding bias by training modality (Pearson’s Chi-squared test, adjusted p > 0.5, Suppl. Table 3). Neither overall dietary patterns differed significantly between HR and LR groups in any subset (Kruskal-Wallis rank sum test, p > 0.1, Fig. 2) nor surveyed dietary information in the average strength gain subset (Dunn’s test or Pearson’s Chi-squared test, adjusted p > 0.03; Suppl. Table 1), and no significant change in nutritional data or activity was found over time (Friedman test, adjusted p > 0.15, Suppl. Table 2).
Fig. 2. Diet differences within and between participants.** A** Ordination of summarized diet (food: fruit & vegetables, fish, meat, eggs, diary, grains, sweets, salty snacks; drinks: water & tea, coffee, juice & lemonade, alcohol in ml) based on euclidean distances of root-mean-square of centered values. Lines connect timepoints per participant. Stress of < 0.2 indicates a fair fit: usable representation, but higher values approach poor interpretation.** B**–D Diet distances within participants (W0-W4, W4-W8, W0-W8) stratified by responder type. p: Kruskal-Wallis rank sum test. W0: week 0, W4: week 4, W8: week 8
The number of training days differed significantly between HR and LR only in the average strength gain subset (21.9 vs. 19.1 sessions; Wilcoxon rank sum test, p = 0.007, Fig. 3D), while training compliance was similar across all groups. At baseline five BioAge measures (core, strength, legs, upper body, total) were 18% to 60% higher in HR than LR and six strength measures (leg curls, abdominal trainer, back trainer, leg press, lateral pulldown, rowing) were 19% to 31% lower in HR than LR (Dunn’s test, adjusted p < 0.02, Suppl. Table 1). A Random Forest model using baseline fitness- and metadata also identified strength measures as the strongest predictors of training response; microbial and metabolomic data were not predictive (Suppl. Figure 6).
Finally, within-individual beta diversity distances were significantly higher in HR than LR in the average strength gain subset (Fig. 3E), but not in the other two strength-based groupings.
Fig. 3. Microbiome change and strength gain.** A** Microbial diversity over the course of the training (Friedman test, p = 0.5).** B** Pearson correlation between changes of the microbial community and changes in fitness (average strength gain) over 8 weeks.** C** Subsetting participants in low, medium and high responders based on change in average strength: Low responders had less than 12.2% strength gain, high responders more than 33%.** D** Number of training days per participant stratified by responder type (average strength).** E** Microbial community changes (Bray-Curtis distance) within participants stratified by responder type (average strength). log_2_FC: log_2_ fold change, W0: week 0, W4: week 4, W8: week 8. Letters denote p < 0.05
Differential Microbiome Changes in High-Responder
As the correlation of longitudinal within-subject shifts in microbial community composition with strength performance was most pronounced in the average strength gain group, this subset was selected for further differential abundance analysis (DAA), focusing on HR. To ensure robustness, a permutation-based approach was used: 100 random subsets of non-high responders (not classified as high responders by any metric) were also subjected to DAA and 99 had a lower number of significant differences compared to HR (p = 0.01, Suppl. Figure 7), additionally, microbial changes in HR with a q < 0.05 were considered unlikely to be due to chance (Fig. 4).
DAA revealed distinct microbial shifts in HRs following resistance training. After four weeks, nine Amplicon Sequence Variants (ASVs) were significantly enriched and four were depleted compared to control subsets. By week eight, these differences became more pronounced, with sixteen ASV increased and eleven decreased in abundance. Notably, a large proportion of differentially abundant sequences belonged to the Lachnospiraceae family. However, no consistent phylogenetic pattern emerged, as both increases and decreases occurred across diverse microbial lineages.
Several ASVs were consistently and significantly enriched at both time points, including ASVs that were associated to Faecalibacterium, Bacteroides massiliensis, Lachnospiraceae, and the Prevotellaceae family. Conversely, consistently and significantly depleted ASVs included Lachnoclostridium, along with sequences from the Oscillospiraceae (NK4A214 group) and Ruminococcaceae families. Notably, an ASV classified as Roseburia hominis showed the most significant increase, while Agathobacter and Butyricicoccus were the most significantly depleted, changes that reached significance only at week eight (Fig. 4).
Fig. 4. Heatmap of significantly different Amplicon Sequence Variants (ASVs) with taxonomies within high responders (average strength gain) over time. The q-value indicates Benjamini-Hochberg corrected p-values that ASVs were found by chance (permutation test with 100 random subsets of non-high responder participants). For each ASV, class and family are indicated as colours, and the lowest reported informative taxonomic level is indicated as K: Kingdom, O: order, F: family, G: Genus, S: Species, Se: Species with exact sequence match. W0: week 0, W4: week 4, W8: week 8. ANCOM-BC2 (formula: asv ~ timepoint + (1|participant), dunnet test) with p ≤ 0.05 and log_2_ fold change ≤-1 or log_2_ fold change ≥ 1. *: p ≤ 0.05, **: p ≤ 0.001, ***: p ≤ 0.00001
Discussion
This study provides evidence that structured resistance training can induce significant and reproducible shifts in gut microbiome composition, particularly in individuals exhibiting pronounced adaptations in muscle strength. By utilizing a large cohort of 150 previously inactive but otherwise healthy adults and integrating digitally controlled resistance training protocols, individual strength trajectories were objectively linked to microbial dynamics. A key methodological strength was the use of digitally controlled resistance training equipment, which enabled individualized and progressively adjusted training stimuli. These standardized protocols likely enhanced the precision and consistency of training across participants, potentially contributing to the observed association between strength development across all exercises and microbial changes. Participants with lower baseline strength demonstrated the greatest relative improvements, consistent with the principle of diminishing returns in resistance training [31, 32]. These findings support the potential of resistance training, particularly when delivered through adaptive modalities, as a modulator of host–microbe interactions.
The absence of significant changes in alpha diversity following resistance training contrasts with findings from endurance-based exercise interventions, where increases in microbial richness and evenness are commonly reported [2, 33–35]. This suggests that alpha diversity may be less responsive to resistance training stimuli [9]. However, shifts in beta diversity among participants with greater strength gains indicate that compositional remodeling does occur and may depend on surpassing a physiological threshold, consistent with a dose–response relationship [36].
The enrichment of taxa such as Faecalibacterium and Roseburia hominis, both producers of SCFAs with anti-inflammatory properties, suggests that resistance training may foster a gut microbial profile conducive to metabolic health and immune regulation [5]. SCFAs have been implicated in maintaining gut barrier integrity, regulating glucose homeostasis [6], and exerting systemic anti-inflammatory effects [2]. These findings are consistent with those reported by Cullen et al. [8], who observed increased abundance of Roseburia and a non-significant trend toward higher levels of Faecalibacterium prausnitzii following a 6-week resistance training program in young adults with overweight and obesity. Their results further support the potential of resistance training to promote SCFA-producing taxa, with possible implications for improved insulin sensitivity [6] and cardiometabolic health [8]. Notably, similar microbial adaptations, including increases in SCFA-producing genera such as Roseburia, have been documented in response to endurance training, highlighting that despite differing exercise modalities, both resistance and endurance training may converge on shared microbiome-mediated pathways that enhance host metabolic and immune function [2, 37].
The modulation of Lachnospiraceae, a diverse bacterial family with both beneficial and potentially pathogenic members, illustrates their complex, context-dependent role in microbiome adaptation to physical activity. While some murine studies link certain strains to reduced endurance, others report performance-enhancing effects from taxa like Coprococcus eutactus [e.g. 38–40]. Importantly, all observed shifts in microbial composition—including the enrichment of Faecalibacterium and Roseburia hominis, as well as changes within the Lachnospiraceae family—occurred in the absence of dietary modification, indicating that resistance training alone may act as an independent driver of gut ecosystem remodeling.
Several mechanisms have been proposed to explain how these microbial shifts may influence muscular adaptations to resistance training. SCFAs—particularly butyrate, produced via bacterial fermentation of dietary fiber—have demonstrated beneficial effects on muscle metabolism and function [41]. The gut microbiota also play a key role in amino acid biosynthesis and metabolism, which are essential for muscle protein synthesis [42, 43], as well as in regulating energy homeostasis, which can impact muscle anabolism and recovery [44]. These interconnected pathways suggest that the gut microbiome may contribute to the physiological adaptations induced by resistance training, although further mechanistic studies are warranted to clarify these relationships.
Despite these compositional changes in the microbiome, no significant alterations were detected in the stool metabolomic profile, an observation that contrasts with previous reports demonstrating a correlation between gut microbiome composition and metabolomic output [45]. This discrepancy may reflect subtle or localized functional changes, limitations of the targeted metabolomics approach, or functional redundancy within the microbiome. It is also possible that transient metabolic responses were missed due to the exclusive use of long-term sampling; short-term assessments may reveal dynamic shifts not captured in this study.
It is also important to consider that stool metabolomics predominantly reflect luminal microbial activity and may not fully capture host–microbe metabolic interactions occurring at the mucosal interface or systemically. Mucosa-associated or circulating metabolites may provide a more sensitive measure of exercise-induced functional adaptation, e.g. Liu et al. found increased serum SCFA in exercise responders [46]. Resistance training may influence host metabolism through pathways not reflected in fecal metabolites, including muscle-derived signaling molecules, systemic inflammation, or shifts in energy substrate utilization. Taken together, these findings emphasize the need for future investigations to integrate multi-omics approaches, such as plasma metabolomics and metatranscriptomics, to more comprehensively characterize the functional impact of exercise-induced microbiome alterations.
From a translational perspective, these findings carry important implications for public health and clinical practice. Resistance training is already recommended for musculoskeletal health and metabolic disease prevention; our results suggest it may also serve as a non-pharmacological strategy to enhance gut health. This opens avenues for integrating resistance training into holistic health promotion programs, particularly for populations at risk of chronic inflammation, metabolic syndrome, or age-related decline in gut function.
Despite its strengths, this study has limitations. The lack of a non-exercising control group precludes definitive causal inference. Implementing a true inactive control group would have required ensuring that control participants remained sedentary for eight weeks and that was not feasible. We therefore adopted a within-subject longitudinal design, using each participant’s baseline as their own control. This approach is commonly used in exercise–microbiome research and provides strong sensitivity for detecting individual microbial and physiological changes (e.g. [47, 48]). Importantly, it allowed us to allocate resources toward a sufficiently large cohort and dense microbiome sampling, thereby increasing the statistical power and robustness of our findings. The generalizability of findings to clinical populations or older adults remains to be established. Additionally, while taxonomic shifts were identified, complementary functional analyses may be needed to fully elucidate the metabolic pathways involved and their relevance to host physiology. Furthermore, although diet logs were collected, the reliance on self-reported data posed a significant limitation to the accuracy of our dietary analysis. For example, the dietary fiber intake could not be estimated reliably but influences SCFA production and gut microbial composition [49]. Finally, biological variability, including factors such as menstrual cycle phase - which was not controlled for despite its known influence on the gut microbiome - may also have contributed to inter-individual differences and obscured subtle effects.
Conclusions
This study shows that resistance training can beneficially shape the gut microbiome, especially in individuals with marked strength gains. The enrichment of health-promoting taxa like Faecalibacterium and Roseburia hominis suggests potential anti-inflammatory and metabolic effects. However, these microbial shifts were not mirrored in the stool metabolome, indicating that functional outcomes may be subtle, localized, or require longer adaptation. Together with previous findings on endurance training, our results highlight physical activity—regardless of modality—as a promising, non-pharmacological approach to support metabolic and immune health. The consistent rise in SCFA-producing microbes across exercise types points to a shared mechanism, underscoring the value of multi-omics approaches in unraveling the complex interplay between exercise, the microbiome, and host physiology.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1.
Supplementary Material 2.
Supplementary Material 3.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ragonnaud E, Biragyn A. Gut microbiota as the key controllers of healthy aging of elderly people. Immun Ageing. 2021;18. 10.1186/s 12979-020-00213-w.10.1186/s 12979-020-00213-w PMC 778437833397404 · doi ↗ · pubmed ↗
- 2Wagner A, Kapounková K, Struhár I. The relationship between the gut microbiome and resistance training: a rapid review. BMC Sports Sci Med Rehabil. 2024;16. 10.1186/s 13102-023-00791-4.10.1186/s 13102-023-00791-4PMC 1076321138166998 · doi ↗ · pubmed ↗
- 3Straub D, Blackwell N, Langarica-Fuentes A, Peltzer A, Nahnsen S, Kleindienst S. Interpretations of Environmental Microbial Community Studies Are Biased by the Selected 16S r RNA (Gene) Amplicon Sequencing Pipeline. Front Microbiol. 2020;11. 10.3389/fmicb.2020.550420.10.3389/fmicb.2020.550420 PMC 764511633193131 · doi ↗ · pubmed ↗
- 4Noguchi K, Gel YR, Brunner E, Konietschke F. npar LD: An R software package for the nonparametric analysis of longitudinal data in factorial experiments. J Stat Softw. 2012;50. 10.18637/jss.v 050.i 12.
- 5Liu Z, Liu H-Y, Zhou H, Zhan Q, Lai W, Zeng Q, Ren H, Xu D. Moderate-intensity exercise affects gut microbiome composition and influences cardiac function in myocardial infarction mice. Front Microbiol. 2017;8. 10.3389/fmicb.2017.01687.10.3389/fmicb.2017.01687 PMC 558514328919891 · doi ↗ · pubmed ↗
- 6Cullen JMA, Shahzad S, Dhillon J. A systematic review on the effects of exercise on gut microbial diversity, taxonomic composition, and microbial metabolites: identifying research gaps and future directions. Front Physiol. 2023;14. 10.3389/fphys.2023.1292673.10.3389/fphys.2023.1292673 PMC 1077026038187136 · doi ↗ · pubmed ↗
- 7Anhê FF, Zlitni S, Barra NG, Foley KP, Nilsson MI, Nederveen JP, Koch LG, Britton SL, Tarnopolsky MA, Schertzer JD. Life-long exercise training and inherited aerobic endurance capacity produce converging gut microbiome signatures in rodents. Physiol Rep. 2022;10. 10.14814/phy 2.15215.10.14814/phy 2.15215 PMC 889774235246957 · doi ↗ · pubmed ↗
- 8Fernández J, Fernández-Sanjurjo M, Iglesias-Gutiérrez E, Martínez-Camblor P, Villar CJ, Tomás-Zapico C, Fernández-García B, Lombó F. Resistance and Endurance Exercise Training Induce Differential Changes in Gut Microbiota Composition in Murine Models. Front Physiol. 2021;12. 10.3389/fphys.2021.748854.10.3389/fphys.2021.748854 PMC 873999735002754 · doi ↗ · pubmed ↗