Life table of Myrmeleon fuscus Yang (Neuroptera: Myrmeleontidae) fed on different natural diets
Tingting Zhang, Xiaokang Li, Meng Liu, Lisheng Zhang, Mengqing Wang, Yuyan Li, Zhongjian Shen, Li Zheng, Jianjun Mao

TL;DR
This study examines how different diets affect the life cycle of Myrmeleon fuscus antlion larvae and adults, finding that certain prey species improve survival and reproduction.
Contribution
The study identifies optimal prey species for M. fuscus larvae and adults, aiding in their potential use for biological control.
Findings
M. fuscus larvae fed on vetch aphids had the shortest larval stage and highest survival rates.
Adult M. fuscus showed better reproductive performance when fed on Tenebrio molitor compared to other prey.
Feeding on T. molitor resulted in the highest body weight for M. fuscus at the third instar.
Abstract
Antlion larvae are sit-and-wait predators that capture arthropods in conical sand traps. The antlion Myrmeleon fuscus Yang (Neuroptera: Myrmeleontidae) is a potential biocontrol agent for the red imported fire ant Solenopsis invicta Buren (Hymenoptera: Formicidae). In this study, we evaluated the life table of M. fuscus when fed on 4 different prey species. The larval stage was shortest when the vetch aphid Megoura crassicauda Mordvilko (Hemiptera: Aphididae) was provided as prey for M. fuscus larvae. Furthermore, the larval survival rate, pupation rate, pupal weight, and emergence rate of M. fuscus fed on M. crassicauda were the highest, suggesting that aphids are the most palatable prey for M. fuscus larvae. M. fuscus adults did not lay eggs when fed with S. invicta or M. crassicauda. They showed a longer oviposition period, higher fecundity, greater intrinsic rate of increase (r),…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
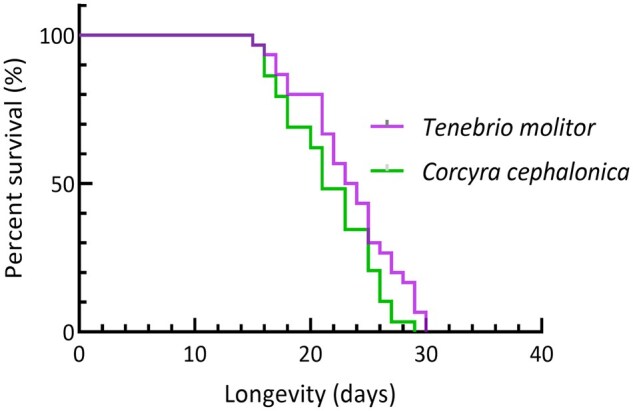 Fig. 1
Fig. 1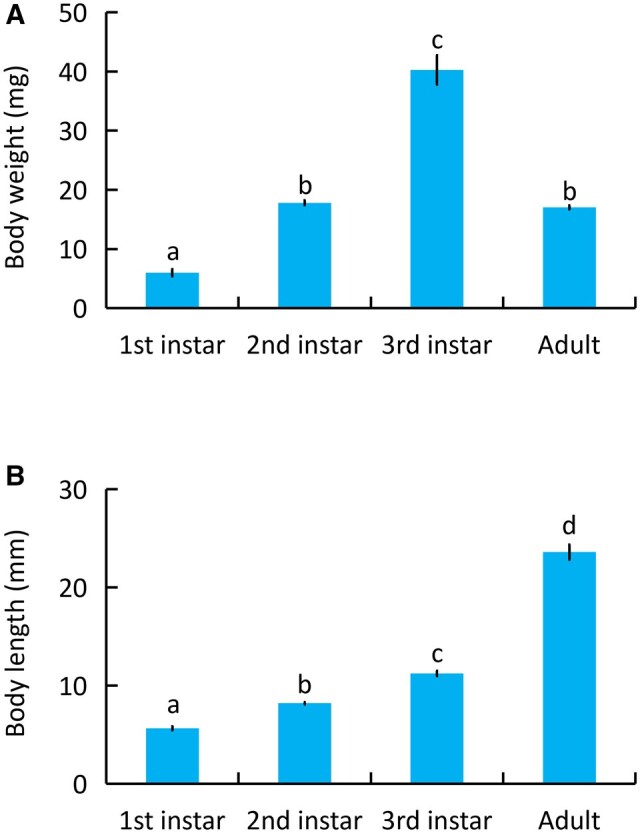 Fig. 2
Fig. 2|
| Mass of prey daily offered to | |||||||
|---|---|---|---|---|---|---|---|---|
|
|
|
|
| |||||
| Type | Mass (mg) | Stage | Mass (mg) | Stage | Mass (mg) | Stage | Mass (mg) | |
|
| Small workers | 1.08 | 1st instar | 1.80 | 1st instar | 1.61 | Adult | 2.68 |
|
| Small workers | 2.15 | 1st instar | 3.59 | 1st instar | 3.25 | Adult | 5.48 |
|
| Small workers | 2.71 | 1st instar | 3.57 | 1st instar | 3.23 | Adult | 8.14 |
|
| Small workers | 3.78 | 4-5th instar | 51.50 | 5th instar | 25.40 | Adult | 10.75 |
| Prey | Developmental period (d) | |||||
|---|---|---|---|---|---|---|
| 1st instar | 2nd instar | 3rd instar | Larval stage | Pupal stage | Adult lifespan | |
|
| 13.48 ± 0.34a | 14.67 ± 0.33a | 18.41 ± 0.41a | 46.56 ± 0.64a | 26.92 ± 0.53a | 4.22 ± 0.19b |
|
| 9.27 ± 0.32b | 12.46 ± 0.22b | 16.00 ± 0.26bc | 37.73 ± 0.47c | 25.90 ± 0.53a | 23.33 ± 0.79a |
|
| 9.40 ± 0.28b | 13.83 ± 0.25a | 16.27 ± 0.34b | 39.50 ± 0.45b | 26.06 ± 0.42a | 21.73 ± 0.72a |
|
| 7.66 ± 0.19c | 10.20 ± 0.24c | 14.73 ± 0.26c | 32.60 ± 0.42d | 26.43 ± 0.40a | 3.83 ± 0.18b |
| Prey | Initial population | Survival rates at different stages (%) | |||
|---|---|---|---|---|---|
| 1st instar | 2nd instar | 3rd instar | Larval stage | ||
|
| 30 | 96.67 ± 3.33a | 93.33 ± 3.33a | 100.00 ± 0.00a | 90.00 ± 5.77a |
|
| 30 | 100.00 ± 0.00a | 100.00 ± 0.00a | 100.00 ± 0.00a | 100.00 ± 0.00a |
|
| 30 | 100.00 ± 0.00a | 100.00 ± 0.00a | 100.00 ± 0.00a | 100.00 ± 0.00a |
|
| 30 | 100.00 ± 0.00a | 100.00 ± 0.00a | 100.00 ± 0.00a | 100.00 ± 0.00a |
| Prey | Initial population | Pupation rate (%) | Pupal weight (mg) | Diameter (mm) | Emergence rate (%) |
|---|---|---|---|---|---|
|
| 30 | 93.33 ± 3.33a | 320.43 ± 6.50a | 8.91 ± 0.09a | 96.67 ± 3.33a |
|
| 30 | 100a | 327.95 ± 6.55a | 9.16 ± 0.06a | 100a |
|
| 30 | 100a | 321.54 ± 5.78a | 9.02 ± 0.06a | 100a |
|
| 30 | 100a | 335.08 ± 5.40a | 9.11 ± 0.06a | 100a |
| Prey | Preoviposition period (d) | Oviposition period (d) | Fecundity (ind.) | F1 hatchability (%) |
|---|---|---|---|---|
|
| — | — | — | — |
|
| 8.23 ± 0.23a | 17.00 ± 0.75a | 14.38 ± 0.73a | 87.00 ± 0.50a |
|
| 8.54 ± 0.27a | 15.00 ± 0.58b | 10.39 ± 0.59b | 83.00 ± 0.37a |
|
| — | — | — | — |
| Prey | Intrinsic rate of increase ( | Net reproductive rate ( | Finite rate of increase ( | Mean generation time ( |
|---|---|---|---|---|
|
| — | 0 | — | — |
|
| 0.021 ± 0.003a | 6.233 ± 1.363a | 1.020 ± 0.0033a | 88.851 ± 0.911a |
|
| 0.017 ± 0.003a | 4.500 ± 0.967a | 1.017 ± 0.003a | 89.278 ± 1.237a |
|
| — | 0 | — | — |
- —National Key R&D Program of China10.13039/501100012166
- —MNR Key Laboratory of Eco-Environmental Science and Technology
- —National Key R&D Program of China10.13039/501100012166
- —Science and Technology Plan Project of Quanzhou City
- —Education and Research Foundation of the Ministry of Education of Fujian Province for Young Teachers
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect and Arachnid Ecology and Behavior · Insect-Plant Interactions and Control · Fossil Insects in Amber
Introduction
The red-imported fire ant, Solenopsis invicta Buren, belongs to the order Hymenoptera and family Formicidae. S. invicta is one of the most devastating insect pests worldwide. It poses serious threats to agriculture, public safety, human health, and ecological security (Zeng et al. 2005). In 2018, S. invicta spread to more than 390 counties, 15 provinces, and regions in southern China. Currently, S. invicta is rapidly expanding (Wang et al. 2020, Li et al. 2023).
Management of S. invicta depends mainly on the use of chemical baits, which effectively suppress its populations. However, the toxicity of chemical pesticides to nontarget organisms and their persistence as environmental residues are indisputable (Li et al. 2018). Biological control is preferred for the long-term management of S. invicta. Natural enemy insects can either directly kill S. invicta or indirectly compete with native ants to suppress its populations. Among these natural enemies, the phorid fly Pseudacteon has demonstrated potential for controlling S. invicta (Lawrence and Lloyd 1997, Chen and Fadamiro 2018, Li et al. 2018). However, the parasitism rate of phorid flies is very low, making the eradication of S. invicta difficult (Calcaterra et al. 2008). To date, the predatory natural enemies of S. invicta have rarely been reported.
One of the potential natural enemies of ants are antlions (Neuroptera: Myrmeleontidae), which are predators widely distributed in tropical regions. Myrmeleontidae is a species-rich family within the order Neuroptera. To date, approximately 2,000 extant species belonging to more than 200 genera have been identified worldwide (Devetak et al. 2010, Lu et al. 2019). According to the “Catalog of the World Antlions,” the genus Myrmeleon comprises 176 described species (Stange 2004). Ten of these species have been reported from mainland China, including Myrmeleon fuscus Yang (Neuroptera: Myrmeleontidae), which is mainly distributed in the central and southern regions (Bao et al. 2009). Antlion larvae are sit-and-wait predators that capture prey using ambush techniques. They constructed funnel-shaped traps in sandy soil and remained buried while waiting for the prey to pass through the soil surface (Abot et al. 2022). They detect the presence of prey at a distance of several centimeters by sensing vibrations generated through prey movement (Devetak et al. 2007, Mencinger-Vracko and Devetak 2008). The behavior and survival of antlions are influenced by both biotic and abiotic environmental factors. Antlions were found to be equally abundant in microhabitats with varying levels of disturbance. However, their density was greater in protected microhabits than in exposed microhabits (Lima 2020). The investment made by antlions in building traps represents a plastic phenotypic response that varies with the surrounding environment. In protected areas, antlions invest more in trap size, which consequently enhances their capture success (Abot et al. 2022). Larval antlions in large traps captured both large and small prey, whereas those in small traps captured only small prey. The trap size and capture success are proportional to the size of antlion larvae (Klokočovnik and Devetak 2013). After a month without food, larval antlions move on average once every 10 days, construct progressively smaller traps, and travel longer distances before establishing a new trap (Heinrich and Heinrich 1984).
Larval antlions are generalist arthropod predators. A large proportion of their diet consists of ants (Topoff 1977). Other arthropods include ticks, beetles, isopods, flies, caterpillars, wasps, lepidopteran larvae, spiders, and mites (Topoff 1977, Crowley and Linton 1999). Analysis of gut contents revealed that many adult Neuroptera are omnivorous or phytophagous. Adult Myrmeleontidae occasionally consume nonanimal foods (New 1991). Larval antlions pass through 3 instars before pupation. They have a remarkable ability to withstand hunger and can survive for up to 3 months without eating. Mature antlion larvae spin silk cocoons before entering the pupal stage, which they remain in for approximately 1 month. Adult antlions are nocturnal predators that are frequently attracted to light (Heinrich and Heinrich 1984, New 1991). They lay eggs in the sand and usually live for about 1 month (Arnett and Gotelli 2001). The life cycle of antlions lasts for 6 months to 2 years, depending on factors such as the availability of food sources, photoperiod, temperature, and metabolic rate (Fisher 1989, Arnett and Gotelli 1999, 2001, Missirian et al. 2006). To date, most studies on antlions have focused on the behavior of larvae in predation and intra- and interspecific competition (Crowley and Linton 1999, Lima 2020, Abot et al. 2022), and some have addressed their energy requirements, fat storage, and tolerance to starvation (Griffiths 1982, 1985, 1991, Lucas 1985, Van Zyl et al. 1997). However, the biology, nutritional requirements, predation spectra, and life tables of adult antlions have rarely been documented.
Recently, the strong predatory ability of M. fuscus against S. invicta has been demonstrated. The third-instar M. fuscus larvae had a theoretical maximum predation amount of 47.17 small S. invicta workers per day. In the field mesocosm experiment, foraging S. invicta workers were reduced by 24.43% on the 30th day after release of the third-instar M. fuscus larvae (Zhang et al. 2026). These studies demonstrates that M. fuscus is a potential biological control agent of S. invicta (Zhang et al. 2026). However, to date, the biological characteristics of this antlion species remain completely unknown, and substitute prey suitable for mass rearing has not yet been identified. Are the prey that do not naturally occur in the antlions’ habitat palatable to them? To answer this question, in the present study, we compared the effects of several substitute prey species on the life history and reproduction of M. fuscus with the aim of identifying suitable prey for the mass production of this natural predator.
Materials and Methods
Collection and Rearing of Antlions
M. fuscus larvae collected from Shakeng Village, Guangchang County, Fuzhou City, Jiangxi Province, China (26°45′N, 116°18′E) were individually housed in plastic cups (8 cm diameter and 15 cm height). The bottom of the cups was covered with a 5-cm-thick layer of sand with a grain diameter of 0.425 mm. First-instar larvae were fed Tenebrio molitor Linnaeus (Coleoptera: Tenebrionidae) once daily; the second- and third-instar larvae were fed twice daily. The pupae were transferred to transparent plastic feeding boxes (33 cm long, 21.7 cm wide, 15.5 cm high) without lids. The bottoms of the boxes were covered with sand similar to that used in cups. A Petri dish containing a cotton ball saturated with water was placed inside each feeding box. Dry branches were provided as habitat for M. fuscus adults, which were also supplied with T. molitor. The masses of prey supplied to M. fuscus are shown in Table 1. The feeding box was placed in a cage (45 × 45 × 50 cm^3^). M. fuscus culture and prey were maintained at 28 °C, 60% relative humidity under a 14 h:10 h light:dark photoperiod.
Developmental Duration and Survival of M. fuscus Larvae
Freshly laid M. fuscus eggs were collected and buried approximately 2 cm deep in sand. An initial population of 30 newly hatched M. fuscus larvae was used for each prey treatment. The M. fuscus larvae were transferred to plastic cups using a fine brush and reared individually to avoid cannibalism. The bottom of each cup was covered with sand, as described above. The M. fuscus larvae were provided with ample prey, including small S. invicta workers, T. molitor larvae, rice moth Corcyra cephalonica Stainton (Lepidoptera: Pyralidae) larvae, and vetch aphid Megoura crassicauda Mordvilko (Hemiptera: Aphididae) adults. The masses of the supplied prey are shown in Table 1. The first-instar M. fuscus larvae were fed once daily and the second- and third-instar M. fuscus larvae were fed twice daily. The prey carcasses were cleaned, and metamorphosis and death were monitored daily. The first-instar stage was defined as the period from hatching to the first ecdysis, the second-instar stage as the period from the first to the second ecdysis, and the third-instar stage as the period from the second ecdysis to pupation.
Pupation and Emergence
After pupation, cocoon size and mass were measured. Pupal size was recorded as the diameter of the cocoons. The pupation rate was calculated as the ratio of successfully pupated larvae to the initial number of larvae. Cocoons were poured into a sieve, and the surrounding sand was removed by gentle shaking prior to weighing on a precision scale (XP205; Mettler Toledo, Switzerland) with an accuracy of 0.0001 g. After weighing, the cocoons were transferred to feeding boxes without lids. Cotton balls saturated with water and dry branches were provided. The feeding boxes were then placed in mesh cages. Adult emergence was monitored daily.
Reproduction and Adult Life Span
Male and female M. fuscus were paired in plastic boxes (25.2 × 17.4 × 9.3 cm^3^) with mesh lids. The 4 prey species were provided twice a day. The heads of T. molitor and C. cephalonica larvae were removed to expose their body fluids as their big size and hard exoskeleton makes it difficult for the antlion adults to prey on them. Inside the boxes, the 2 prey items were held by hand, attracting the antlions to fly over and feed on them. Feeding lasted for 2 min. S. invicta small workers and M. crassicauda adults were dropped to the bottom of the box. The number of eggs (F1 generation, offspring of male and female pairs) was counted daily. The lifetime fecundity (number of eggs oviposited by each female throughout the adult stage), preoviposition period (between emergence and the first day of oviposition), and oviposition period (between the first and last days of oviposition) were recorded. To measure the hatching rate, 10 F_1_ eggs were randomly collected and neonates were counted. The hatching rate was evaluated across 10 replicates. Adult lifespan is expressed as the period from emergence to death.
Body Length and Weight of M. fuscus Fed on T. molitor
The body length and weight of M. fuscus fed on T. molitor were measured in separate experiments. To estimate the size of M. fuscus larvae and adults, body length (from the tip of the head to the end of the abdomen) was measured using calipers. Larvae and adults were weighed on a precision scale (XP205, Mettler Toledo, Switzerland). The sizes and masses of the first-, second-, and third-instar M. fuscus larvae were measured 8, 10, and 15 days after hatching. M. fuscus adults were measured on day 2 after eclosion. Ten individuals were examined for each parameter at each developmental stage.
Data Analysis
The demographic characteristics of M. fuscus that fed on S. invicta workers, T. molitor larvae, C. cephalonica larvae, and adult M. crassicauda were analyzed using the TWOSEX-MSChart program (Chi 2022) based on the age-stage, two-sex life table theory (Chi et al. 2020). Age-specific survival rate (l_x_), age-specific fecundity (m_x_), net maternity (l_x_m_x_), intrinsic rate of increase (r), net reproductive rate (R0), finite rate of increase (λ) and mean generation time (T) were calculated as previously described (Chi and Liu 1985). The differences in these life parameters among different prey treatments were analyzed by paired bootstrap test (B = 1,000) based on 95% CIs of the difference. These are expressed as follows:
Homogeneity of variance was confirmed before testing the means. Differences in the duration of the larval stage among different prey treatments, and body weight and body length among different developmental stages were determined using 1-way analysis of variance (ANOVA Least Significant Difference). Differences in first-, second-, and third-instar stages, pupal stage, adult lifespan, survival rate, pupation rate, pupal weight, pupal diameter, and emergence rate among the 4 prey treatments were determined using the nonparametric Kruskal–Wallis test. Differences in the oviposition period and lifetime fecundity between the T. molitor and C. cephalonica groups were determined using the Student t test. Differences in the preoviposition period and egg hatching rate between the T. molitor and C. cephalonica groups were analyzed using the nonparametric Mann–Whitney test.
Results
Duration of Larval and Pupal Stages and Adult Lifespan
The M. fuscus first-instar stage (H = 69.872, *df *= 3, *P *< 0.01), second-instar stage (H = 69.421, *df *= 3, *P *< 0.01), and third-instar stage (H = 40.051, *df *= 3, *P *< 0.01) were significantly affected by the prey. The larval stage of M. fuscus that fed on S. invicta workers was significantly longer than that of M. fuscus that fed on T. molitor larvae, C. cephalonica, and M. crassicauda (F3,113=132.133, *P *< 0.001). The larval stage of M. fuscus feeding on M. crassicauda adults was the shortest, suggesting a rapid development rate. No significant differences were observed in the duration of pupal stage among the four prey treatments, suggesting that pupal development was not significantly affected by prey (H = 3.578, *df *= 3, *P *= 0.311). M. fuscus adults did not consume T. molitor and C. cephalonica larvae. They died of starvation at early adult stage. The M. fuscus adult lifespan was significantly longer when reared on these 2 prey species than on S. invicta workers and M. crassicauda adults (H = 89.253, *df *= 3, *P *< 0.01) (Table 2, Fig. 1).
Life expectancy of Myrmeleon fuscus adults fed on different prey.
Survival Rate
The survival rates of M. fuscus at different larval stages were all above 90% when fed on S. invicta workers and reached 100% when fed on T. molitor larvae, C. cephalonica larvae, and adult M. crassicauda. The survival rates at the first-instar stage (H = 3.000, *df *= 3, *P *= 0.392), second-instar stage (H = 6.600, *df *= 3, *P *= 0.086), third-instar stage (H = 0.000, *df *= 3, *P *= 1.000), and throughout the larval stage (H = 6.545, *df *= 3, *P *= 0.088) did not differ significantly among the four prey treatments (Table 3).
Pupation Parameters
There were no significant differences in pupal mass among the 4 prey treatments (H = 4.211, *df *= 3, *P *= 0.240). The M. fuscus pupation rate was not significantly affected by the prey (H = 6.600, *df *= 3, *P *= 0.086). The differences in pupal diameters among the 4 prey treatments were not significant (H = 5.050, *df *= 3, *P *= 0.168). The emergence rate was not significantly affected by the prey (H = 3.000, *df *= 3, *P *= 0.392) (Table 4).
Reproduction
Female adults supplied with S. invicta workers and M. crassicauda adults did not oviposit, as they neither attacked nor consumed these 2 prey species. The preoviposition period (U = 70,000, Z = −0.789, *P *= 0.479) and egg hatchability (U = 40.500, Z = −0.744, *P *= 0.481) did not differ significantly between the T. molitor and C. cephalonica treatments; however, the oviposition period (*P *= 0.045) and lifetime fecundity (P< 0.001) did (Table 5).
Population Parameters
The net reproductive rate (R0) was zero, and the intrinsic rate of increase (r), finite rate of increase (λ) and mean generation time (T) were not provided by the TWOSEX-MSChart program in the S. invicta and M. crassicauda treatments, as M. fuscus supplied with these 2 prey did not lay eggs. The 4 population parameters did not differ significantly between T. molitor and C. cephalonica treatments (Table 6).
Size of M. fuscus Fed on T. molitor
The body weight of M. fuscus fed on T. molitor increased rapidly at the larval stage and declined at the adult stage. However, the body length increased continuously with increasing age and peaked at the adult stage. The average body weights of the first instar, second instar, third instar, and adults were 5.98 ± 0.59, 17.81 ± 0.38, 40.26 ± 2.50, and 17.01 ± 0.32 mg, respectively (F3,36=121.12, *P<*0.001) (Fig. 2A). The mean body lengths of the first instar, second instar, third instar, and adults were 5.64 ± 0.19, 8.21 ± 0.10, 11.24 ± 0.26, and 23.60 ± 0.76 mm, respectively (F3,36=457.67, *P<*0.001) (Fig. 2B).
Body weight and length of M. fuscus fed on T. molitor. A) Body weight. B) Body length. Bars represent means ± SE of 10 replicates. Different letters above the bars indicate significant difference (n = 10, 1-way ANOVA followed by LSD, P < 0.05).
Discussion
Augmentative biological control relies on mass production of high-quality natural enemies. An ample and cost-effective diet is fundamental for mass production of generalist predators (Sahayaraj 2001, Riddick 2009). The diets of generalist predators can be classified into 2 major categories based on their sources: natural prey and artificial diets, each with its distinct advantages and disadvantages. Currently, biocontrol producers usually raise natural enemies on their natural prey or host, which, in turn, maintained on their host plants. Unlike natural prey, an artificial diet does not involve maintenance of the 3 trophic levels (Riddick 2009). However, developing a palatable artificial diet that meets the nutritional requirements of predators and supports the continuous production of offspring is a challenging task (Riddick 2009). Furthermore, to date, natural prey suitable for the mass rearing of antlion species, especially antlion adults, has not been exploited. Therefore, in the present study, we investigated the life table of a Myrmeleontidae species, when fed on 4 prey species from different orders, with the aim of identifying palatable natural prey suitable for its mass production.
Although antlion larvae feed on ant species such as the red imported fire ant and leaf-cutting ants (Atta spp.) (Missirian et al. 2006, Zhang et al. 2026), these prey are not suitable for the mass rearing of antlions. This is because indoor rearing and maintenance of ant colonies are difficult and sufficient supply of ants are not available in the market. Therefore, we selected 3 nonant insects, M. crassicauda, C. cephalonica, and T. molitor as prey of M. fuscus, using S. invicta as a reference. We argue that whether antlions attack the 3 nonant species in the wild is irrelevant, as the goal of the present study was to screen for the best prey for indoor mass production of M. fuscus. The vetch aphid, M. crassicauda can be raised on a large scale using broad bean seedlings as hosts. C. cephalonica and T. molitor are in ample supply in the market at low cost. Therefore, if these prey prove palatable, the mass production of M. fuscus could be achieved cost-effectively. As expected, M. fuscus showed the longest larval stage when fed on S. invicta (Table 2). This indicates that the nonant prey species are more palatable to M. fuscus larvae than S. invicta, because nutritious diet can shorten larval developmental period (Omkar and Srivastava 2003, Arshad et al. 2021). Most important of all, M. fuscus did not consume S. invicta and died of starvation, implying that S. invicta is not palatable to M. fuscus adults. The possible reason is that antlion adults have different digestive physiology from larvae and cannot metabolize the venom proteinaceous and alkaloidal toxins contained in ants (Touchard et al. 2016). It is worth mentioning that the manual feeding method for the M. fuscus adults may cause deviations in their behavior, as we found that they did not consume T. molitor and C. cephalonica larvae placed at the bottom of the box. We cannot completely rule out the possibility that the M. fuscus adults were attracted by the manual feeding action, rather than by the exposed body fluid of the prey.
We found that M. fuscus larvae and adults have different prey preferences by comparing the effects of prey on the demographic parameters of M. fuscus larvae and adults. Among the 3 nonant prey species that do not naturally occur in the habitat of M. fuscus, M. crassicauda proved to be most palatable to M. fuscus larvae. This conclusion is supported by the short larval stage and high larval survival rate, pupation rate, pupal weight, and emergence rate of M. fuscus larvae fed with M. crassicauda. However, among the 3 nonant prey species, T. molitor was the most palatable to M. fuscus. This is because the adult lifespan and oviposition period were the longest, and fecundity, intrinsic rate of increase (r), net reproductive rate (R0), and finite rate of increase (λ) were the highest when T. molitor was provided to M. fuscus adults. R0 is an important parameter that is closely related to population growth and fecundity (Sayyed et al. 2008). The maximum R0 value of M. fuscus adults fed on T. molitor suggests that this prey promoted rapid development and consequently led to a high reproductive rate and a long oviposition period for the predator (Amer et al. 2018, Azher et al. 2019).
The pupation parameters of M. fuscus were not significantly affected by prey. This is consistent with the results of another antlion species, Myrmeleon brasiliensis Navás (Neuroptera, Myrmeleontidae). When M. brasiliensis larvae were fed different prey, including leaf-cutting ants (Atta spp.), fruit fly larvae (Anastrepha spp. and Ceratitis capitata), and a mixed diet (Atta spp. plus fruit flies), neither the pupal period nor the pupal size was significantly influenced by the prey type (Missirian et al. 2006). Similar results indicated that natural prey with different palatability had no discernible effects on the pupation of the two antlion species.
M. fuscus adults died soon after being supplied with S. invicta workers and M. crassicauda adults because they did not consume the prey. This outcome may not occur under natural conditions, where a variety of arthropods in the natural habitat are available as potential prey for M. fuscus adults. M. fuscus adults fed on C. cephalonica and T. molitor showed normal reproductive performance, demonstrating that these 2 prey species successfully sustained the reproduction of M. fuscus adults. However, the lifetime fecundity (14.38) of M. fuscus adults that fed on T. molitor was relatively low, suggesting that this prey was not palatable enough for M. fuscus adults. To address this limitation, further studies are required to identify prey species that can better sustain adult reproduction in M. fuscus. Another solution is to provide multiple prey species that can improve food quality and increase the reproductive output of polyphagous predators (Topoff 1977). For example, M. brasillensis antlions showed the shortest developmental duration when fed a mixed diet (leaf-cutting ants plus fruit fly larvae), which was related to a more comprehensive composition and better nutritional quality of the diet. Finally, supplementation with plant components, such as pollen, may contribute to the reproduction of antlions (Missirian et al. 2006).
This study indicated that the vetch aphid M. crassicauda was the most suitable for mass rearing of M. fuscus larvae, followed by T. molitor. Although T. molitor is not as palatable as M. crassicauda for M. fuscus larvae, it is cost-effective and has an abundant supply in the market. T. molitor was the most palatable species for M. fuscus adults. Considering the palatability and availability of the prey tested, we conclude that T. molitor is the most suitable species for mass production of M. fuscus throughout its life stage. Based on this conclusion, we measured the body length and weight of M. fuscus fed on T. molitor. The M. fuscus demographic parameters obtained will contribute to the screening of more palatable prey and development of artificial diets.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abot AR , Arguelho EG, do Nascimento Lima T. 2022. Foraging behavior plasticity in antlion larvae Myrmeleon brasiliensis (Neuroptera, Myrmeleontidae). Int. J. Trop. Insect Sci. 42:591–595. 10.1007/s 42690-021-00577-6 · doi ↗
- 2Amer MES , Fu Y, Niu L. 2018. Biological aspects of Orius similis Zheng reared on two preys at three constant temperatures. J. Agric. Sci. Technol. 8:350–363. 10.17265/2161-6256/2018.06.002 · doi ↗
- 3Arnett AE , Gotelli NJ. 1999. Bergmann’s rule in the ant lion Myrmeleon immaculatus De Geer (Neuroptera: Myrmeleontidae): geographic variation in body size and heterozygosity. J. Biogeogr. 26:275–283. 10.1046/j.1365-2699.1999.00271.x. · doi ↗
- 4Arnett AE , Gotelli NJ. 2001. Pit-building decisions of larval antlions: effects of larval age, temperature, food, and population source. J. Insect. Behav. 14:89–97. 10.1023/A : 1007853730317
- 5Arshad M , Ullah MI, Khan RR, et al 2021. Demographic parameters of the reduviid predator, Rhynocoris marginatus (Reduviidae: Hemiptera) fed on two lepidopterous insect pests. Bio Control 66:227–235. 10.1007/s 10526-020-10060-9 · doi ↗
- 6Azher F , Khan MM, Bilal M, et al 2019. The development of Antilochus coquebertii Fabr. (Heteroptera: Pyrrhocoridae) on different artificial diets. J. Kansas Entomol. Soc. 91:192–208. 10.2317/0022-8567-91.3.192 · doi ↗
- 7Bao R , Wang XL, Liu JZ. 2009. A review of the species of Myrmeleon (Neuroptera: Myrmeleontidae) from mainland China, with the description of a new species. Entomol. News 120:18–24. 10.3157/021.120.0108 · doi ↗
- 8Calcaterra LA , Delgado A, Tsutsui ND. 2008. Activity patterns and parasitism rates of fire ant-decapitating flies (Diptera: Phoridae: Pseudacteon spp.) in their native Argentina. Ann. Entomol. Soc. Am. 101:539–550. 10.1603/0013-8746(2008)101[539:APAPRO]2.0.CO;2 · doi ↗