Phages and quorum sensing: findings to consider in phage therapy
Laura Fernández-Garcia, Lucia Blasco, Inés Bleriot, Lucía Arman, Clara Ibarguren-Quiles, Antonio Barrio-Pujante, Manuel González de Aledo, Rodolfo García-Contreras, Rafael Cantón, Thomas K. Wood, María Tomas

TL;DR
This paper reviews how bacteria use quorum sensing to defend against phages and how manipulating this system could improve phage therapy.
Contribution
The paper highlights the dual role of quorum sensing in both defending against and promoting phage infection, and explores novel treatment approaches using QS inhibitors.
Findings
Quorum sensing can both protect bacteria from phages and sometimes promote phage infection.
QS inhibitors like SsoPox-W2631 and penicillinic acid may enhance phage therapy.
Modifying quorum sensing can influence bacterial defense mechanisms against phages.
Abstract
This review aims to provide an overview of current knowledge on the involvement of QS in phage infection. The role of QS in bacterial defence against phages is emphasized, without overlooking the fact that QS can sometimes also promote phage infection. We also review the implications of QS in phage therapy and current perspectives. For the bibliographic review, PubMed and Google Scholar were used to search for publications on “quorum-sensing” and “phage infection”. The relationships between bacteria and phages are extremely complicated and involve several mechanisms. Quorum sensing (QS) is a communication system involved in controlling bacterial fitness, both at population and individual levels. Phages (viruses that infect bacteria) play a major role in the natural regulation of bacterial populations. In order to protect themselves, bacteria have developed several defence mechanisms…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
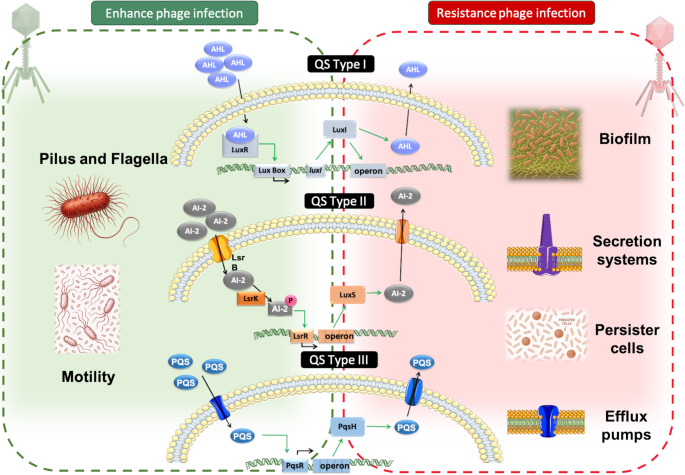 Figure 1
Figure 1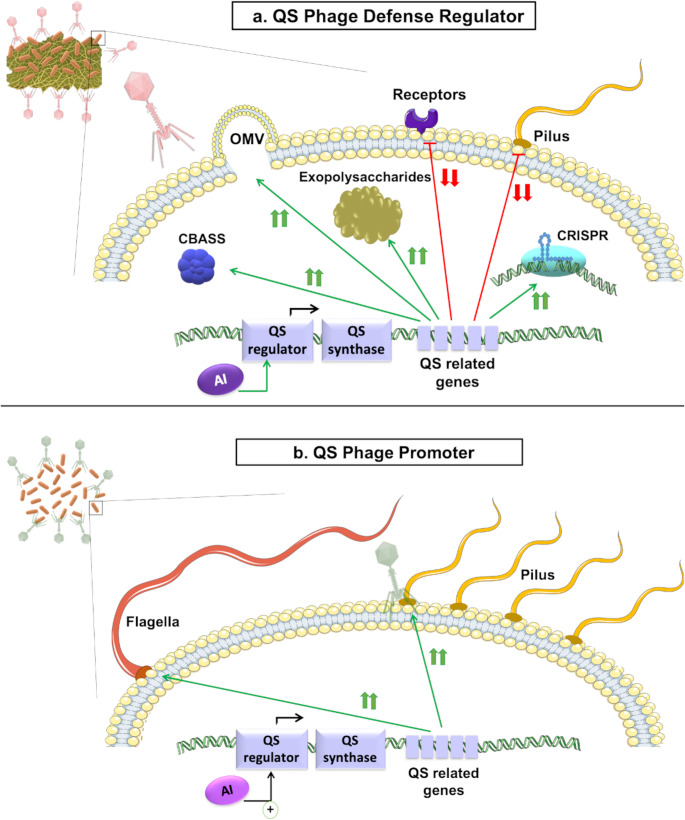 Figure 2
Figure 2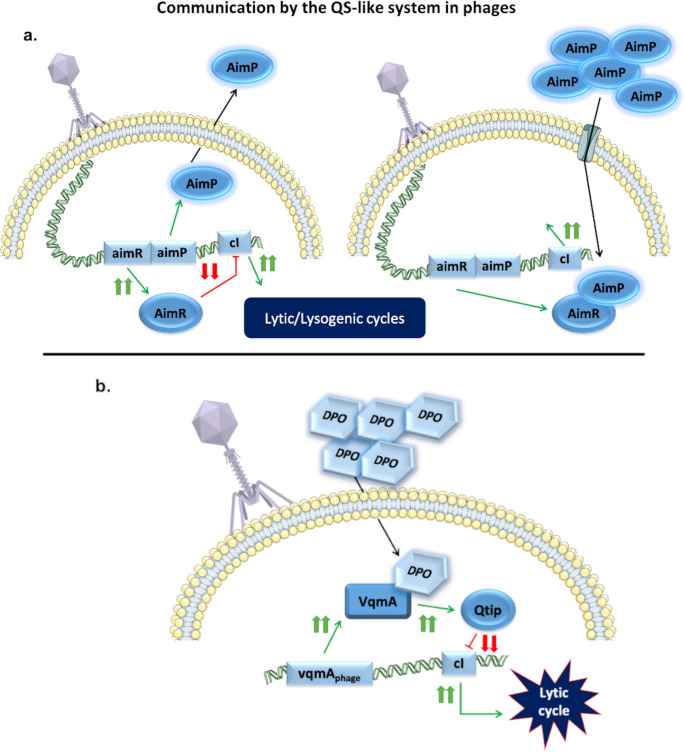 Figure 3
Figure 3- —Universidade da Coruña
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Vibrio bacteria research studies · Bacterial Genetics and Biotechnology
Introduction
Phages, viruses that infect bacteria, are considered a major natural regulator of bacterial populations. In order to manage this, they have two types of life cycles: the lysogenic cycle, in which the phage genome becomes integrated into the bacterial genome; and the lytic cycle, in which the phage enters the host, uses the bacterial machinery to replicate itself and then lyses the host to release its progeny [1]. Due to the constant interaction between phages and their bacterial hosts, both are permanently updating their defence/counter-defence strategies [2].
The use of phage therapy has recently been proposed as a promising alternative to antibiotic therapy. The increase in multiresistant bacteria for which no viable antibiotic treatments are available has become increasingly common in the last few decades and is now a worldwide health problem [3]. Clinical trials are being developed and legal regulations implemented to enable the use of phage therapy in hospitalized patients in several countries [4]. One of the requirements of phage therapy is the use of strictly lytic phages, as the nucleic acid of this type of phage will not become integrated in the bacterial genome, thus preventing horizontal gene transfer of virulence and resistant genes [5, 6]. Additionally, phage therapy is considered a safe and very specific treatment. Phages have a narrow spectrum of action, being able to infect only those bacterial targets that carry specific receptors [7]. This specificity enables directed treatment against the pathogen, evading commensal bacteria [4, 6].
Phage multiplication is directly related to the number of bacteria available. Thus, the greater the number of bacteria, the greater the number of potential hosts and the greater the number of new phages produced, which in turn increases phage pressure, driving bacterial resistance [8, 9]. Phage progeny rates have been shown to increase in logarithmic phase. Phages spread readily under conditions of high cell density (HCD) [10], although the bacterial quorum sensing (QS) communication system is also activated under such conditions [11]. Nevertheless, it is worth mentioning that HCD conditions might contribute to bacterial starvation conditions, significantly reducing bacterial activity and therefore, substantially reducing phage susceptibility [12, 13]. Bacteria have developed several phage-resistant mechanisms to protect themselves, especially during slow infection cycles, during which the bacteria in the population promote the activation of QS and related defence mechanisms. These strategies can be classified according to their mechanism of action into the following: prevention of phage adsorption, prevention of phage assembly, degradation of phage nucleic acid, and increase in the number of persister cells [14]. However, phages have developed anti-defence mechanisms that allow them to overcome the mentioned bacterial defences, keeping both bacteria and phages in a constant arms race [15].
The QS communication system is based on secreted signals (auto-inducers, AIs) that enable bacteria to determine population density and activate metabolic routes that are too energetically expensive to be activated individually (i.e., biofilm) [16]. QS is based on the accumulation of molecules, so that when there are few bacteria in the population, the number of AIs in the environment will not be enough to activate QS. However, as the population increases, accumulation of AIs in the environment can activate QS and related mechanisms [17]. The QS system is a global regulator that has been related to several bacterial mechanisms, ranging from bioluminescence to resistance and including motility, biofilm formation, and virulence [18]. The regulation is context-dependent, i.e., during the first stage of biofilm formation (attachment), QS promotes motility and also production of fimbriae; during the following steps (microcolony formation and maturation), the QS system represses these mechanisms and promotes polysaccharide excretion to form the biofilm matrix [19]. All QS systems have four main characteristics: (i) production of diffusible molecules, AIs; (ii) production of cytoplasmatic or inter-membrane receptors specific for the AIs; (iii) auto-stimulated production of AIs; and (iv) regulation of several bacterial mechanisms [20]. In addition to positive autoregulation, QS can also undergo negative autoregulation in order to balance the cost/benefit of resources, reducing the expression of QS-related genes when the bacterial population is not large enough to develop cooperative responses [21]. For this type of communication, bacteria use several types of AIs, including autoinducing peptides (AIPs), acyl-homoserine lactones (AHL) (previously known as AI-1), CAI-1 (S−3-hydroxytridecan-4-one), quinolones, autoinducer-2 (AI-2), and leaderless communication peptides [20, 22, 23]. The signals vary depending on the bacterial species, although many bacteria can detect signals from other species, such as Escherichia coli, which detects -but does not produce- AHL [24]. Some molecules related to cell communication, such as indole, are inter-kingdom, as well as inter-species, being detected and produced by bacteria, plants, and animals [23].
The AI-2 based system is found in both Gram-positive and Gram-negative bacteria. Thus, AI-2 molecules are recognized by their membrane detectors (LuxP, LsrB, Cah-R, YeaJ) [25–27], entering the cytoplasm through transport channels and phosphorylating LsrK kinase, activating the Lsr operon, producing more AI-2, and regulating bacterial mechanisms [18]. Gram-negative bacteria have another two systems - one based on homoserine lactones (HSLs) and the other based on quinolones. In Pseudomonas aeruginosa, HSLs diffuse through the membrane and activate a LuxR-like transcriptional regulator (LasR or RhlR), which activates the Lux-like operon, including LuxI-like synthase (LasI or RhlI), to produce more signals. Conversely, the Pseudomonas quinolone signal (PQS) enters the cell through transporters to bind to the PqsR regulator, activating the Pqs operon [28, 29]. In Gram-positive bacteria, such as Staphylococcus aureus, the AIPs are recognized by a membrane histidine kinase, AgrC, which activates the accessory gene regulator (agr) cluster, leading to overexpression of exoproteins and downregulation of cell-wall-associated proteins [28]. Activating QS operons generates a cascade of activation affecting multiple important systems, which regulate several bacterial mechanisms involved in defence (biofilm and persister cell formation, efflux pump and secretion system activation), metabolism (bioluminescence, polysaccharide synthesis), and virulence (motility, toxin production), among others (Fig. 1) [30, 31]. In the last few decades, QS has been shown to regulate bacterial phage-defence mechanisms, including the following: (i) expression of surface proteins, (ii) biofilm formation, both of which are involved in adsorption inhibition; (iii) Clustered Regularly Interspaced Short Palindromic Repeats and associated proteins (CRISPR-CAS), involved in degrading phage nucleic acid; and (iv) toxin-antitoxin systems, involved in persister cell formation [17].Fig. 1. Graphical representation of the three main QS mechanisms and how the bacterial metabolism is affected. The green background highlights the mechanisms by which QS, depending on the situation, enhances phage infection, while the red background highlights those involved in phage resistance
This mini-review aims to discuss the effects of QS in phage infection and how phages can control the host QS to their own benefit. In addition, some very promising molecules for use in phage therapy (based on recent findings) are mentioned. We consider here the complex interaction between QS and phages, and the possibility of exploiting this interaction to enhance phage therapy.
Basic knowledge
Phages use specific receptors to infect bacteria. These receptors are surface structural proteins that are involved in several bacterial properties or mechanisms, including membrane structure, molecular transportation, and bacterial motility [32]. As already mentioned, QS is a global regulator of cell metabolism, affecting several mechanisms. The global regulation of bacterial metabolism affects, among other things, the genes involved in the synthesis of some of these structural proteins [33]. It is precisely through this regulation that the activation of QS in bacteria can be indirectly involved in phage infection. During the early stages of HCD, QS promotes bacterial motility (upregulating flagellar and pili genes, used for many phages as receptors). Under HCD (High Cell Density) conditions, QS can cause an increase in the secretion of bacterial extracellular matrix, reducing motility to promote the formation of biofilms, which thus hide phage receptors and prevent access to the host [34].
QS as phage defence regulator
During phage infection, several stress responses are triggered in bacteria. One of the main molecules that bacteria activate under stress conditions is the alarmone signal or (p)ppGpp [35]. Stringent response (ppGpp) is known to activate QS [36]. It is possible that QS may be activated in response to phage infection through the stress response and (p)ppGpp.
One of the first defence mechanisms involves the modification or elimination of the membrane receptors, directly preventing phage adsorption. The reduction in receptors is closely related to QS. Although E. coli cannot produce AHLs, it can detect them through the SdiA regulator, which acts as a LuxR-type transcriptional regulator, regulating cell metabolism in response to external signals [24, 37, 38]. Through SdiA, E. coli strains have been observed to reduce susceptibility to phages such as λ and χ when exogenous AHLs are present in the environment. It has been demonstrated that in the presence of AHLs, E. coli reduced the amount of LamB protein, a maltose outer membrane channel used by phages as a receptor, and its depletion therefore reduces phage adsorption [39]. A similar increase in phage susceptibility was later observed when sdiA was deleted from Klebsiella pneumoniae strains. Although the authors did not observe any increase in phage resistance when C6-homoserine lactone (C6-HSL) was added to the cultures, proving that SdiA has a role independent from that of C6-HSL [40]. Moreover, Vibrio cholerae showed a reduction in the O1 antigen (highly variable surface polysaccharide region of Gram-negative bacteria [41]), used by many Vibrio phages as a receptor, when the high concentration of the auto-inducer is consistent with HCD conditions [42]. Likewise, other Vibrio spp. have shown similar responses to QS. Under HCD conditions or in the presence of synthetic AHLs, expression of the OmpK receptor decreased in V. anguillarum, and cell aggregation was reduced [43]. Li et al. (2025) discovered that under conditions of low cell density, QS regulates the expression of BcsE receptor in Vibrio alginolyticus [44].
The bacterial strategies used to hide phage receptor proteins also include the production of outer-membrane vesicles (OMVs) and the formation of capsules and biofilms (Fig. 2a). OMV production is a QS-dependent bacterial decoy that deflects phage attack, as the OMVs are produced when the phage is adsorbed to the bacterial surface, and phages then infect the OMVs instead of the actual bacteria [8, 45]. Biofilm production is strictly dependent on cell density and, therefore, on QS. Biofilms are bacterial communities in which cells are joined to each other and to surfaces by a matrix of extracellular polymeric substances, including polysaccharides, lipids, nucleic acids, and proteins [46, 47]. The polysaccharide matrix protects bacteria against adverse environments and several antimicrobial agents [48]. The increased production of biofilm creates a barrier that reduces phage access to their bacterial receptors [49]. Furthermore, Enterococcus faecalis QS controls the production of gelatinase GelE, as a bacterial defence mechanism. The GelE is a secreted metalloprotease, encoded in the Agr-like operon (Fsr cluster), which is directly activated by bacterial AIs, considered essential for induction of biofilm formation in Enterococcus sp., as well as being related to virulence [50, 51]. Sheriff et al. (2024) observed significant downexpression of GelE during phage infection, hypothesizing that phages reduce biofilm formation by repressing gelE, which produces downregulation of lrgA (murein hydrolase regulator) and lrgB (antiholin), both involved in protection from extracellular lysis of phages [52]. The previously mentioned SdiA regulator has also been related to biofilm production in the presence of external AHLs, both in E. coli and K. pneumoniae, reducing phage accessibility [39, 40]. Moreover, phage BUCT640 of P. aeruginosa uses the type Psl exopolysaccharides (which are highly dependent on QS) as receptors. The authors demonstrated that the susceptibility of P. aeruginosa QS mutants to the phage is significantly increased, showing that QS increases phage resistance by regulating biofilm production through Psl maturation [53].Fig. 2(a) QS as a regulator of phage defence. Graphical representation of the mechanism activated by QS to defend bacteria from a phage infection under conditions of high cell density. Green arrows denote activation, while red lines represent inhibition. (b)QS as a promoter of phage infection. Graphical representation of the mechanism activated by QS that promotes phage infection during early stages of HCD. Green arrows indicate activation
Bacterial QS has been related to membrane receptors and biofilm production, and also to internal defence systems such as CBASS (Cyclic oligonucleotide-based antiphage signalling system) and the CRISPR-Cas. The CBASS anti-phage defence system is an abortive infection-type system, which is formed by a cyclase, an effector protein, and, in most cases, accessory genes involved in cyclase regulation [54]. Activation of the cyclase produced cycled-nucleotides as second messengers, triggering effector proteins, causing reduction of cell energy, degradation of DNA, or disruption of the bacterial membrane, inhibiting phage propagation [55]. Hoque et al. (2016) observed QS-dependent overexpression of haemagglutinin protease (HAP), which reduced phage susceptibility by significantly decreasing phage stability, presumably by degrading phage particles. Additionally, the HapR regulator has been shown to upregulate the CBASS promoter in V. cholerae [54]. The authors demonstrated that under HCD conditions, QS enhances HapR, which activates the CBASS system, increasing anti-phage defence [56].
CRISPR-Cas systems are considered to represent an adaptive immune system in bacteria [57, 58]. These systems consist of repeat arrays separated by small foreign DNA known as spacers. These spacers are acquired by Cas proteins, which cut short fragments of foreign DNA, and are incorporated into the CRISPR array [59, 60]. CRISPR-Cas systems are classified into 6 types, divided into two classes depending on whether they use multi-subunit Cas proteins or a multi-domain Cas as the effector [61]. CRISPR-Cas systems react to previously encountered foreign DNA and cleave it, preventing phage replication [60]. Patterson et al. (2016) reported that under HCD conditions, QS promotes CRISPR-Cas immunity by enhancing spacer acquisition [62]. The ecological significance of QS interaction with CRISPR-Cas was explored by Goldberg et al. (2014), who demonstrated that CRISPR-Cas type III-A plays a role in controlling the induction of lysogenic phages in a transcription-dependent manner, avoiding self-targeting [63]. Although involving plasmids, Høyland-Kroghsbo et al. (2017) demonstrated the link between CRISPR-Cas activity and QS in P. aeruginosa. These authors observed a loss of the plasmid both in HCD conditions and when the media was supplemented with AIs, suggesting that CRISPR-Cas regulation by QS balances the cost/benefit ratio of CRISPR-Cas activation during bacterial growth [64]. According to Vale et al. (2005), activation of CRISPR-Cas systems is highly expensive for bacteria, creating a delicate energy balance when phages are involved [65]. Thus, in conditions where there are a large number of phages per bacteria, resistance by elimination of surface receptors is less costly; also, when there are very few phages per bacteria, activation of CRISPR-Cas could lead to autoimmunity [65, 66]. Similar studies have shown QS regulation of CRISPR-Cas activity, and in several of these, no relationship was observed between AI-2 presence and CRISPR-Cas mediated phage-defence in E. coli [67]. It has been suggested that QS adaptive immunity is a widespread mechanism across bacterial species under HCD conditions [62].
Bacteria can defend themselves by slowing their metabolism and entering a state of dormancy that prevents phage multiplication. In this respect, it has been shown that in the presence of AI-2, E. coli reduces cell metabolism through LsrB and not only downregulating T4P receptor [67]. Additionally, a QS-dependent phage-resistance evolution was observed in P. aeruginosa. The authors found that quinolones activate the alkyl-quinolone QS system and thus increase phage resistance. They suggested that external quinolones may be involved in phage resistance evolution either through activation of phage resistance mechanisms or by decreasing the cost of resistance in cells lacking a complete QS and thus altering the balance between cooperation and “cheating” in the population [68].
QS as a promoter of phage infection
Despite the above findings, QS has also been found to promote phage infection under certain conditions. The increase in phage infection is usually related to the fact that QS promotes the expression of some of the genes used by phages as receptors (Fig. 2b). The effect of QS as a global regulator in bacteria, together with the fact that phages use structural proteins, can cause this contradiction. Therefore, inhibition of QS would lead to a subsequent reduction in the number of phage receptors on the cell surface, thus decreasing the infectivity. Analysis of the behaviour of phage-resistant mutants of V. alginolyticus in response to phages Athena1 and VaphiSt2 revealed that several genes related to QS synthesis and detection were downregulated relative to the sensitive wild-type strain, showing that spontaneous mutants inhibit QS as a phage defence mechanism. These researchers found that the two-component sensor systems for AHL and AI-2 were inhibited in both of the mutants analysed, while in one of them (Athena1-resistant), deletion also occurred in the QS two-component CAI-1 histidine kinase sensing system and affected the cqsS gene. The researchers also found that many virulence factors were depleted in both of the phage-resistant mutants (Athena 1 and VaphiSt2 resistant-mutants) [69].
In contrast to previous findings in P. aeruginosa, it has been observed that inhibition of QS by Baicalein, a flavonoid compound extracted from plants [70], generates a reduction in pilus-dependent phage infection as well as an increase in CRISPR-Cas immunity [71]. Baicalein has been demonstrated to increase lysis of QS-receptor TraR in E. coli [72] and reduce expression of QS-genes in P. aeruginosa [73]. The authors noted that in the presence of this QS inhibitor, the amount of phage DMS3vir receptors decreased, also reducing the elimination of phages due to CRISPR-Cas activity caused by a reduction in phage adsorption rates. However, they found that in the presence of Baicalein and a high titre of phages, more cells were resistant due to CRISPR immunity than in the control. The authors also found that Baicalein promotes sensitive bacteria, through regulation of T4P-dependent genes when cocultured with receptor-lacking strains [71]. Likewise, Pseudomonas aeruginosa responds to environmental indole by reducing the availability of phage receptor T4P (type IV pilus). However, it was not established whether the deficit in the receptor was due to downregulation of the gene expression or inhibition of proper pili assembly [74]. Xuan et al. (2022) also observed an increase in phage adsorption in P. aeruginosa due to QS upregulation of GalU, a protein essential for lipopolysaccharide synthesis, and therefore in phage receptors. These researchers demonstrated the effect of lactose inhibitor (lacI) in upregulating galU and thus boosting phage adsorption [75]. Recently, Cao et al. (2024) found that LasR in P. aeruginosa also promotes phage infection by increasing the synthesis of T4P receptor [53]. Moreover, the SdiA receptor can promote phage infection in Cronobacter sakazakii by reducing surface hydrophobicity and extracellular matrix, which reduces biofilm formation and increases cell motility, thereby increasing accessibility to the receptors needed by the phage [76]. It has also been shown that in the presence of AI-2, V. cholerae activates VqmR, which represses biofilm formation, in turn, increasing access of phages to their host [77].
Communication by a QS-like system in phages
Improved understanding of the phage-bacteria relationship has led to the discovery of ways in which phages monitor and even use bacterial QS to their advantage. The arbitrium system is known as a phage QS-like communication system (Fig. 3a) [78]. This system, found in 72% of phages in the SPbeta group, allows the phages to enter the lytic or lysogenic cycle depending on the amount of recent infections in the population. Briefly, phages express aimR and aimP as early genes during infection; AimR activates aimX, which is a lysogeny inhibitor, while AimP is processed into a mature signalling peptide that is secreted from the host and incorporated in the rest of the cells in the population by the OPP transporter. In conditions of high infectivity, high levels of AimP will be secreted by the bacteria, thus blocking AimX activity, and the lysogenic cycle will therefore take place [78, 79]. However, the Arbitrum system is not the only way in which phages control their lytic-lysogenic cycles according to QS. Vibriophages are known to harbour a homolog of VqmA DPO-binding QS receptor (VqmAPhage). This receptor detects DPO and produces transcription of the antirepressor Qtip, which inhibits the cIVP882 repressor, activating the lytic cycle of the prophage (Fig. 3b) [80]. Thus, Vibriophages that encode VqmAPhage can regulate their life cycles according to when the probability of infection success is higher. In addition, these phages have also been found to control the host QS by using VqmAPhage to activate VqmR expression and therefore VqmA production [80]. Another well-known example of how phages control bacterial QS is the ability of some Clostridium difficile prophages to code for Agr genes. Phages have been described to harbour all these genes, except the Agr receptor (AgrA). Hargreaves et al. (2014) suggested that under suitable conditions, prophages could produce an HCD response without the number of bacteria usually required [81]. Recently, a LuxR-type receptor named Apop was discovered in an Aeromonas popoffii phage, although the mechanism of action remains unclear [82].Fig. 3. Communication by the QS-like system in phages.(a) Graphical representation of the arbitrium system. The image on the left represents the induction of the lytic cycle at low phage density. The image on the right shows the induction of the lysogenic cycle under high phage density conditions. (b) VqmA_phage_ system. Representation of how phage codifies VqmA and controls the choice between the lytic and lysogenic cycle on the basis of bacterial cell density. Green arrows indicate activation and synthesis, while red lines denote repression
Prophages are known to harbour genes that can regulate the host metabolism when integrated in the bacterial genome, and also protect the host from being infected by other phages [83]. Shah et al. (2021) investigated the ability of the DMS3 prophage from P. aeruginosa to inhibit the host QS. This prophage produces an anti-quorum-sensing protein (Aqs1), which binds the LasR regulator as well as the pili synthesis gene PilB at the early stages of infection, thus inducing superinfection exclusion by reducing expression of the phage receptor. On the other hand, inhibition of LasR by Aqs1 was proposed as a way of avoiding QS-related anti-phage defence mechanisms [39, 62, 84]. Similarly, the PfsE protein in filamentous P. aeruginosa prophages was shown to inhibit PQS as well as the pili synthesis gene PilC. PfsE binds PqsA, blocking QS signalling at the same time as binding PilC, reducing phage receptor synthesis. This double function, as in the previous case, allows the phage to protect the lysogen and therefore itself by preventing superinfection [85]. Moreover, the bci gene of P. aeruginosa filamentous prophages was demonstrated to respond to external AHLs and also regulate the host QS. Ambroa et al. (2020) observed that bci not only favours phage infectivity but also regulates expression of the QS regulator genes lasR,* rhlR*,* qscR*, and pqsR. In addition, the presence of this gene was shown to significantly increase the bacterial virulence, increasing pyocyanin levels, biofilm formation, and therefore reducing motility. The bci gene was thus suggested to be an important ally to the maintenance of P. aeruginosa in the lungs in cystic fibrosis patients [86]. Another example of phages modulating the host QS has been described in the P. aeruginosa lytic phage LUZ19, which codes for a QS targeting protein (Qst), which, in turn, interacts with PqsD, directly disrupting the PQS biosynthesis pathway and causing inhibition of the host QS. Moreover, Qst has been shown to interact with acetyl-CoA and thiamine metabolism, inhibiting cell division and reprogramming the host metabolism to promote phage replication. The authors hypothesized that Qst regulates bacterial energy by reducing the cost of both QS mechanisms, affecting phage-defence systems and acetyl-CoA pathways, to redirect energy towards phage propagation [87, 88]. Furthermore, Leblanc et al. (2009) discovered an acyl hydrolase in an Iodobacter phage that degrades AHLs and controls the host QS [89].
Phages can control and monitor bacterial QS to their own benefit to avoid bacterial defence mechanisms, redirect bacterial energy, and communicate with each other. However, prophages can also monitor the host QS to decide when to switch from lysogenic to lytic cycle. This activity has been observed in several E. coli phages, such as λ phage, which induce the lytic cycle when the host is present in an environment with a high concentration of AHLs [90]; or more recently, the prophage T1, which transcriptional regulator Pir monitors the levels of AI-2 and cAMP in the host to induce the lytic cycle and upregulate holin production, which is needed for degradation of the cell wall in the last states of the lytic cycle. However, this mechanism is predicted to be widely distributed among phages, as Pir homologues have been found in several phage species [91]. Nevertheless, Mauritzen et al. (2023) demonstrated that some V. anguillarum prophages inhibited the phage lytic cycle during HCD conditions [92].
Remarkably, it has been suggested that phages create selective pressure towards functional QS systems in bacteria [93, 94]. Lopez et al. (2018) observed the evolution of an Acinetobacter baumannii hospital collection in 10 years, from 2000 to 2010. In the 2010 collection, the acquisition of two temperate phages and a complete QS/QQ system was observed in the majority of the population [93]. In addition, Saucedo-Mora et al. (2017) found several P. aeruginosa phages that infect preferably functional QS strains rather than their ΔlasR/ΔrhlR mutants [95]. On the other hand, Davies et al. (2016) showed that some phages (such as P. aeruginosa φ4) prefer to infect QS-deficient bacteria involved in chronic infections, positively selected them in the population [96].
The same QS signal can interact with phage infection by reducing both bacterial metabolism and phage receptors. However, the same signal in a different bacterium may cause the opposite effect by increasing the effectiveness of the infection. These contradictory effects can be explained by the fact that phages use important host elements, which are regulated by QS. These findings must be considered when QS molecules are used with phage therapy, without overlooking the previously mentioned ability of some phages to regulate and monitor bacterial QS to their own benefit.
Community dynamics
The involvement of QS in phage defence is a complicated subject and involves a delicate equilibrium between QS-mediated gene expression and the activation of antiviral defence mechanisms. The QS is regulated in an HCD-dependent manner, and this system can act by favouring or avoiding phage infection. However, HCD conditions usually occur in bacterial populations composed of different strains and in many environments, of various species. Phage infection in heterogeneous populations and the consequent activation of defence mechanisms stimulate changes in the population, affecting the delicate balance. These heterogeneous populations usually contain QS-defective or “cheating” strains, which either lack QS receptors or QS effectors [97, 98]. Ahator et al. (2022) analysed the dynamics of QS and CRISPR-Cas defective strains in a mixed population of P. aeruginosa infected with phages. These authors observed a reduction in phage numbers when bacteria were grown in cocultures rather than monocultures, implying compensation of the different mechanisms that are lacking (QS or CRISPR) in the strains through cooperation between different types of bacteria cooperation between diverse types of bacteria. They also suggested that a mixed population may promote the emergence of resistance in bacteria [99]. These findings were corroborated by Li et al. (2025), who observed a reduction in phage abundance in cocultures of P. aeruginosa with and without effective QS. These researchers noted that the application of cell-free supernatant, rich in C4-HSL, to the cultures increased the susceptibility to the phage. The researchers suggested that QS is involved in reducing the metabolic activity of the cells, promoting their entry into a persister state, which would reduce the creation of QS-deficient mutants [100, 101]. They also hypothesized that the phage could recognize and synchronize with these persister cells while waiting for metabolic reactivation to resume propagation [101].
On the other hand, several authors examined how the population changes when multiple bacterial species are present in the population, in an attempt to better understand the changes that occur in the body when phage therapy is applied. To this end, Mumford et al. (2017) analysed the effects of Pseudomonas phage PT7 in a mixed population of P. aeruginosa/P. aeruginosa QS-deficient, Staphylococcus aureus, and Stenotrophomonas maltophilia. They found that the presence of phages can reduce the total number of bacteria, even though the P. aeruginosa QS-mutant was significantly more resistant to the phage than the wild-type. Nevertheless, they also observed an increase in the other pathogens when P. aeruginosa is targeted. In addition, the authors observed a reduction in phage-resistant appearance in the mixed communities, suggesting that the high cost of adaptation, with the need for competition between pathogens, could favour the effectiveness of phage therapy [102]. Moreover, the dynamics of a population consisting of P. aeruginosa, S. aureus,* A. baumannii*, and Burkholderia cenocepacia in the presence of Pseudomonas DMS3vir phage was analysed. Under these conditions, it was observed that P. aeruginosa dominates the population in the absence of phage, but it is substituted by A. baumannii in the presence of phage. The authors also noted that although P. aeruginosa CRISPR-mutant is fitter in the absence of phages, it is almost eradicated from the population when phage is involved. In addition, these authors proposed that targeted phage treatment of a dominant species in a mixed population helps to maintain the population diversity and that the other species in the community will block reinfection [103].
Phage therapy and qs, implications and perspectives
This section presents a case report showing the effect of phage infection on bacterial QS and highlighting the most promising molecules proposed for enhancing phage therapy.
Case report
Blasco et al. (2023) presented a case report of a patient with a multidrug-resistant P. aeruginosa infection in a prosthetic vascular graft and who was treated with a three-phage cocktail. After several months of infection and multiple antibiotic treatments, the phage therapy was administered in a regimen of one intravenous injection per day for a week, in combination with ceftazidime-avibactam. After administration of the phage therapy, the strain showed a reduction in resistance to both β-lactamics and quinolone antibiotics (previously resistant). The re-sensitized strain was sequenced and compared with the pre-treatment strain; several mutations in virulence-related (secretion systems, oxidative stress), antibiotic-resistant-related (ion transport, efflux pumps), and phage-resistant genes (surface receptors, QS) were observed. This report shows the evolutionary pathway of bacteria towards phage resistance by increasing biofilm production, and several mutations in surface receptors, toxin-antitoxin systems, metabolic genes, and prophages, directly reducing bacterial virulence and increasing antibiotic sensitivity. Importantly, mutations appeared in the PvdQ gene, an acylase that hydrolyses AHL, inhibiting QS [104]. The mutation in PvdQ increased the activation of QS and, allegedly, its regulated phage-defence mechanisms [105].
Molecules in use
Due to the involvement of QS in the activation of bacterial defence mechanisms, the use of anti-QS agents as treatment has been deeply studied, focusing on five main strategies: (i) inactivation of QS-receptors, (ii) inhibition, (iii) degradation, (iv) blockage of QS signals, and (v) combination of these agents and antibiotics [106, 107]. In light of the importance of QS also in bacterial phage defence, several molecules have been studied to enhance phage therapy by affecting QS. Some of the most promising ones are mentioned below (Table 1).Table 1. Molecules in use and promising genes for enhancing phage treatmentNameOrigenTargetTarget speciesObserved reactionReferencesMycotoxinPenicillic acid Aspergillus flavus LasR/RhlR P. aeruginosa QS reduction and derived resistance mechanisms [108, 109]LactonaseSsoPox-W163I Saccharolobus solfataricus AHLs/CRISPR genes P. aeruginosa Reduction of CRISPR activity and Pyocyanin [110–112]FlavonoidCinnamaldehyde (CAD) Cinnamomum spp.AI-2 K. pneumoniae QS reduction and derived resistance mechanisms [113–116]Lactonase geneAiiA Bacillus spp.AHLs/PilB P. aeruginosa/ E. coli Biofilm reduction and β-galactosidase and QS [117]Anti-QS geneAqs1DMS3 phageLasR P. aeruginosa QS reduction and derived resistance mechanisms [84]QS-targeting geneQstLuz19 phagePqsD/Acetyl-CoA P. aeruginosa Reduction of PQS synthesis [87]Phage geneGp70.1PaP3 phageRpoS P. aeruginosa Reduction of cell metabolism and stress response [118]
One of the compounds that was first found to have promising synergy with phage therapy was penicillic acid. This compound was found to increase the susceptibility of P. aeruginosa to phages by targeting LasR and RhlR regulators and thus inhibiting QS [108] and, therefore, also phage-resistant mechanisms regulated by QS. The authors also found that the RNA polymerase of the phage was affected by the presence of penicillic acid [109]. Similarly, the QS-inhibition activity of SsoPox-W163I lactonase (an enzyme that degrades the lactone ring of HSLs) [110] was analysed to determine its potential application in phage therapy. SsoPox-W263I significantly decreases secreted factors, like pyocyanin, in Pseudomonas spp., as well as biofilm production in P. aeruginosa and Chromobacterium violaceum, and violacein in the latter. The presence of SsoPox-W263I also produced downexpression of CRISPR-associated genes in both P. aeruginosa and C. violaceum [111]. The combination of SsoPox-W263I with phages was found to be a promising strategy, as this enzyme reduces virulence and also the appearance of phage-resistant mutants [112]. Cinnamaldehyde (CAD) is another QS inhibitor with potential in phage therapy as it interacts with AI-2 of different species of bacteria [113–116]. The presence of CAD was shown to increase phage susceptibility in K. pneumoniae clinical strains, presumably by avoiding the reduction of membrane proteins and the activation of phage-defence systems [116].
Some authors have used genetically modified phages to inhibit QS. The modified phage carries an enzyme able to disrupt biofilm formation by degrading AHLs (AiiA) [117]. It was demonstrated that the modified phage was able to reduce β-galactosidase activity in the bacterial target. These results open up a new pathway in phage therapy, as AiiA can disrupt the QS of several bacterial species [117]. Moreover, several candidate genes could be engineered in phages, such as the previously mentioned Aqs1 [84] and Qst [87], and also the Gp70.1 from P. aeruginosa phage PaP3 [118]. Gp70.1 has been shown to inhibit growth in P. aeruginosa and E. coli by interacting with the RpoS. The authors demonstrated that the presence of Gp70.1 substantially reduced motility, amino acid and sugar metabolism, stress response, and virulence, highlighting its potential as an antibacterial agent targeting the regulation of sigma factor [118]. However, the identification of new potentially therapeutic genes is complicated since most phage gene functions are unknown, and there is no bioinformatic reference [118].
The combination of anti-QS agents and phages has shown promising in vitro results, becoming an encouraging new path to enhance phage therapy and combat multiresistant bacteria. However, the specific mechanisms by which the above compounds work are still unknown, and diverse efficiencies were observed between species or even strains. Moreover, AiiA and Gp70.1 would require phage engineering, increasing the difficulty of their use. Nevertheless, deeper studies would fill these gaps in our knowledge, taking these methods a step closer to their application.
Conclusion
In this mini-review, multiple interactions between QS and phage infection are considered. The importance of QS in phage defence is highlighted, without disregarding the paradoxical effects on the expression of some phage receptors, and how phages can control or monitor bacterial QS to their benefit. Finally, several molecules with high synergic potential for use with phage therapy are proposed. In addition, a promising approach would be the use of a triple combination of anti-QS agents, phages, and antibiotics, reducing bacterial resistance. Although the interaction between bacterial QS and phages is complex, modifying bacterial QS as a method of enhancing phage therapy is a promising strategy. However, for effective use of QS in phage therapy, it is important to identify which phage to use, mainly in relation to its bacterial receptors.
The reference list from the paper itself. Each links out to its DOI / PubMed record.