Comparison of virulence factors genes between exoU and exoS Pseudomonas aeruginosa in microbial keratitis
Tanzina Akter, Fiona Stapleton, Jake Ireland, Mark Willcox

TL;DR
This study compares virulence genes in two types of Pseudomonas aeruginosa linked to eye infections, finding that exoU strains have higher levels of certain harmful genes.
Contribution
The study identifies pldA and flaG as potentially important virulence factors specific to exoU Pseudomonas aeruginosa in microbial keratitis.
Findings
exoU strains showed significantly higher prevalence of pldA and flaG genes compared to exoS strains.
PLD activity was correlated with the presence of the pldA gene.
Findings were consistent across isolates from India and Australia, though differences were more pronounced in Indian isolates.
Abstract
This study aimed to investigate differences in virulence factors between exoU and exoS lineages of Pseudomonas aeruginosa from microbial keratitis (MK). Initially, the whole genome sequence (WGS) data of 20 exoU and 19 exoS P. aeruginosa keratitis isolates from India and Australia retrieved from NCBI genome database was assessed for the presence of different virulence genes using the Virulence Factors Database (VFDB). To confirm the VFDB results, a separate set of keratitis isolates consisting of 148 P. aeruginosa (36 from India and 112 from Australia) were screened by PCR for the presence of exoU, exoS, and the virulence genes found to be significantly different in the VFDB analysis. Flagellar length was measured by transmission electron microscopy (TEM) and phospholipase D (PLD) activity was determined by Amplex Red Phospholipase D Assay Kit. From 327 virulence-associated genes, the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
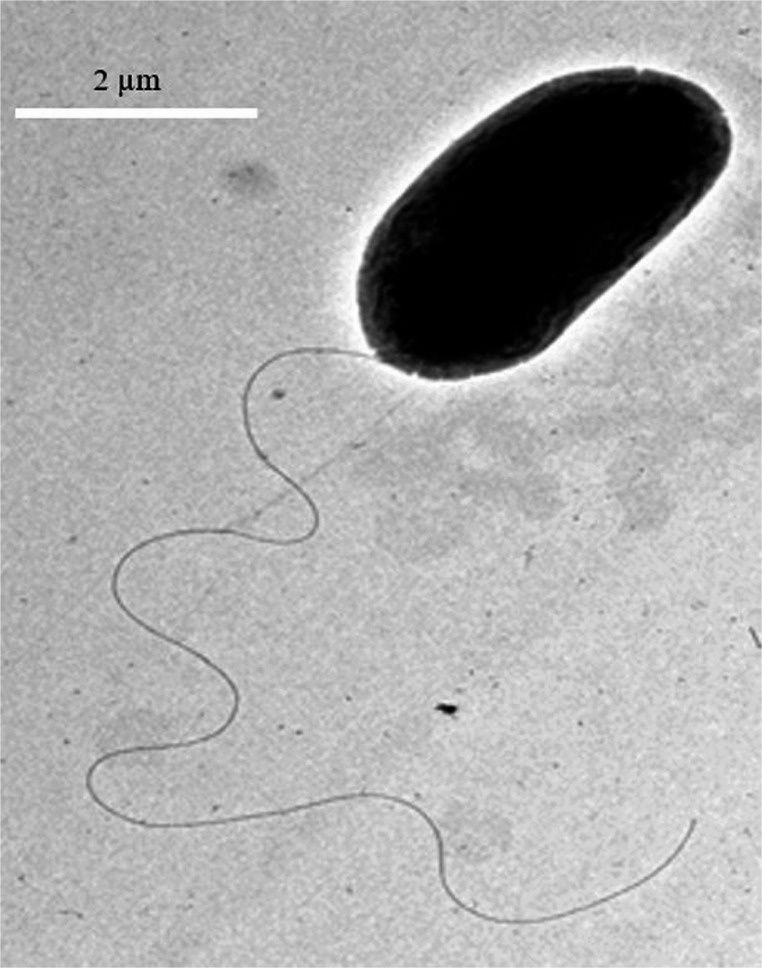 Figure 1
Figure 1- —University of New South Wales
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial biofilms and quorum sensing · Ocular Infections and Treatments · Legionella and Acanthamoeba research
Introduction
Pseudomonas aeruginosa is an opportunistic pathogen that can cause microbial keratitis (MK), a severe sight-threatening infection of the cornea, and P. aeruginosa is one of the most commonly isolated bacteria from these infections [1]. P. aeruginosa possesses an extensive array of both cell-associated and secreted virulence factors which can be chemical or proteinaceous and can contribute to its complex pathogenicity in MK [2, 3]. Among its cell-associated structures, type IV pili and a single unsheathed polar flagellum play crucial roles in adhesion to host surfaces [4–6]. Type IV pili also facilitates twitching motility, while swimming motility is achieved through the coordinated activity of the flagellum and type IV pili [2, 7, 8].
To deliver its proteinaceous secreted virulence factors, P. aeruginosa utilizes six distinct secretion systems, designated as the Type 1 secretion system (T1SS) to Type 6 secretion system (T6SS) [9, 10]. Among these, the Type III secretion system (T3SS) is particularly associated with high virulence leading to severe infections. This sophisticated system enables the bacterium to inject effector proteins directly into the cytoplasm of eukaryotic host cells, where they disrupt the cell and interfere with essential cellular signalling pathways [11]. To date, four effector proteins secreted by the T3SS have been identified: ExoU, ExoS, ExoT, and ExoY [12]. ExoU can rapidly kill host cells by hydrolyzing host membrane lipids through its phospholipase activity [13, 14]. ExoS is a bifunctional effector, containing an N-terminal GTPase-activating protein (GAP) domain and a C-terminal ADP-ribosyltransferase domain [15–18]. ExoS disrupts the host’s actin cytoskeleton, encouraging endocytosis of the bacteria, interferes with vesicular trafficking, induces the formation of membrane blebs, all of which are thought to eventually lead to cell death with features of apoptosis or necrosis [15–18]. ExoS and ExoT are homologous and both are bifunctional, though they differ in their cellular targets and pathogenic roles [19, 20]. Additionally, ExoY functions as an adenylate cyclase, increasing intracellular cAMP levels and further impairing host cell processes [12]. Based on their T3SS effector profiles, P. aeruginosa strains can be broadly classified into two groups: exoU containing strains are called cytotoxic strains as they kill mammalian cells rapidly while exoS possessing strains are termed as invasive strains causing a slower cell death. It is worth noting that the distribution of these effector genes is not uniform, and in many cases, exoS and exoU genes are mutually exclusive [21]. This distribution pattern may be attributed to their enhanced fit in distinct ecological niches, as these genes likely provide specific advantages in different environments. ExoU strains have been associated with more severe clinical outcomes than exoS strains in ocular infections [22–25], which might be attributed to differential expression of additional virulence factors during infection.
Specific virulence factors of P. aeruginosa from different infections including those from keratitis have been explored [3, 26–30]. However, differences in virulence factors between exoU and exoS strains of P. aeruginosa have not been previously investigated in keratitis isolates. Therefore, this study examined the differences in virulence factor profiles of these two lineages in strains isolated from MK.
Materials and methods
P. aeruginosa isolates
A total of 187 P. aeruginosa isolates from MK comprising 132 samples from Australia and 55 from India (collected between 1993 and 2022) were included in this study. Each isolate was obtained from a separate patient with a unilateral infection and with no personal or identifying information was collected. The bacterial strains were stored at −80 °C in the microbial culture collection at the School of Optometry and Vision Science, UNSW Sydney, Australia, and later revived on nutrient agar (Oxoid Ltd., Basingstoke, Hampshire, UK). Species identification was verified using the VITEK-2 system for Gram-negative bacteria (BioMérieux, Baulkham Hills, NSW, Australia).
Study of differences of virulence factors genes between exoU and exoS isolates
The WGS data of 20 exoU (13 from India and 7 from Australia) and 19 exoS (6 from India and 13 from Australia) P. aeruginosa keratitis isolates were retrieved from NCBI genome database (Bio project numbers: PRJNA590804 and PRJNA431326). These 39 strains were chosen as cluster analysis from the core and pan genome indicated that these exoU and exoS strains generally segregate into two distinct phylogenetic clusters, normally exoS tends to Group 1 and exoU tends to Group 2, indicating broadly representative of the genomic divergence between these two lineages [27, 31]. In addition, these isolates were sequenced using the same technology, with comparable genome coverage and identical assembly methods, minimizing potential methodological variability that could affect gene detection [27, 31]. These WGS data were assessed for the presence of different virulence genes in these two lineages by the Virulence Factors Database (VFDB) [32]. P. aeruginosa PAO1 and P. aeruginosa PA14 were used as reference strains for analysis of exoS and exoU strains respectively. VFDB uses VFanalyzer for construction of orthologous groups within the query genome and pre-analyzed reference genomes from VFDB. This step reduces false positives caused by paralogous genes. After that, it performs iterative and comprehensive sequence similarity searches across the hierarchical VFDB datasets, enabling detection of strain-specific virulence factors. Lastly, a context-based refinement is applied for virulence factors encoded by gene clusters, improving both specificity and sensitivity. Genes found to differ significantly between exoU and exoS strains in VFDB analysis, were further confirmed by BLAST searches using reference strains gene sequences. To confirm the VFDB results, the remainder (148 keratitis isolates) were screened for the presence of exoU and exoS and virulence genes that were found to be significantly different using PCR following DNA extraction in genes identified in the VFDB analysis.
PCR for the prevalence of virulence factors genes
DNA was extracted from fresh bacteria grown overnight on Tryptone Soy Broth (TSB; Oxoid Ltd., Basingstoke, UK) at 37 °C using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions and guidelines. After extraction, the DNA concentration was determined in µg/ml at an optical density reading of A260/A280. The DNA was stored at −20 °C for PCR. The prevalence of exoU in 29% and exoS in 71% genes of isolates had been reported previously [33]. Strains that were double-positive or double-negative for exoU and exoS genes were excluded from the present study. Amplification of pilA,* pldA*,* algP*, and flaG genes was performed using gene specific primers. The primer sequences, PCR cycling condition, amplicon size (bp), and composition of PCR reaction mixture are presented in Supplementary Tables 1 and 2. Following PCR, 1.0% agarose gel containing Sybr Safe (Thermo Fisher Scientific, USA) was used to confirm the size of the amplified DNA fragments of the genes comparing to the known DNA ladder fragments. To minimize false-positive and false-negative PCR results, primers for exoU, exoS, pldA, and pilA were selected from previous studies where they had been successfully applied and shown to be highly conserved [34–36]. FlaG and algP primer pairs were designed in conserved regions, and reproducible PCR results were obtained across multiple isolates. Prior to amplifying the genes from the 148 study samples, primers specificity was evaluated using 39 bacterial isolates with available WGS data. The PCR results were positive for all WGS-positive samples and negative for all WGS-negative samples, demonstrating complete concordance between the two methods.
Determination of pldA activity
The pldA gene encodes for a PLD enzyme which breakdowns phosphatidylcholine (contained in lecithin) to produce choline and phosphatidic acid. P. aeruginosa isolates possessing pldA gene (5 isolates) and negative for pldA gene (5 isolates) were randomly selected from the 187 strains to evaluate the activity of PldA in both exoU and exoS groups. Briefly, the bacterial strains were cultured overnight in Luria-Bertani medium supplemented with phosphatidylcholine until OD reached to 0.4. Cells were harvested by centrifugation at 2,000 × g for 10 min at 4 °C to collect supernatant for the measurement of PLD activity using the Amplex Red Phospholipase D Assay Kit (Invitrogen), following the manufacturer’s instructions. In this assay, firstly, PLD hydrolyzes phosphatidylcholine (lecithin) to produce choline and phosphatidic acid. The choline oxidase subsequently oxidizes choline, generating betaine and hydrogen peroxide (H₂O₂). In the presence of horseradish peroxidase (HRP), the H₂O₂ reacts with the Amplex Red reagent (10-acetyl-3,7-dihydrophenoxazine) in a 1:1 stoichiometric ratio to produce resorufin, a highly fluorescent product which has excitation and emission maxima at approximately 530 nm and 590 nm, respectively. For each reaction, 100 µL of the bacterial supernatant was added to a 100 µL mixture containing 50 µM Amplex Red reagent, 1 U/mL horseradish peroxidase, 0.1 U/mL choline oxidase, 0.25 mM lecithin in a Coster 96 well microplate reader and then incubated for 30 min at 37 °C, and fluorescence was measured using excitation at 530 ± 12.5 nm and fluorescence detection at 590 ± 17.5 nm. A standard curve was prepared using H₂O₂ (Supplementary Table 3), and the PLD activity of the test samples was expressed as the equivalent concentration of H₂O₂ (µM) generated during the reaction.
Determination of effects of the presence of the flaG gene on flagellar length
For determination of effects of the flaG gene on flagellar length, a total of 10 randomly selected P. aeruginosa strains which included two exoU strains possessing the flaG gene (PA2 and PA23), three exoU strains without the flaG gene (PA4, PA46, PA169), two exoS strains with the flaG gene (PA11 and PA125), and three exoS strains without the flaG gene (PA3, PA129 and PA140) were chosen and flagellar length was measured by transmission electron microscopy (TEM) following a standard negative staining protocol [37]. Briefly, P. aeruginosa isolates were cultured overnight in tryptic soy broth at 37 °C with shaking at 120 rpm. The following day, bacterial cells were gently resuspended by flicking the culture tube to avoid mechanical disruption of the flagella. Carbon-coated TEM grids were glow-discharged for 20 s to improve sample adherence. A 10 µL aliquot of the live bacterial suspension was carefully applied to each grid and allowed to settle for 5 min at room temperature. Excess liquid was gently removed with filter paper, and grids were inverted onto droplets of 4% paraformaldehyde (PFA) for 5 min to fix the cells. Following fixation, excess PFA was removed by wicking, and grids were sequentially inverted onto two droplets of 2% uranyl acetate for negative staining - the first for 1 min and the second for 5–10 s. The grids were then washed by inverting onto three consecutive droplets of double-distilled, filtered water for 5–10 s each, without drying between washes. After air-drying at room temperature, flagella were visualized using TEM (Thermo Fisher Scientific Talos L120C LaB₆, USA) operating at 120 kV and images were taken with a digital camera (Thermo Fisher Scientific Ceta-S CMOS, USA). Flagellar lengths were measured using ImageJ software (National Institutes of Health, USA) [38].
Statistical analysis
To compare differences in the frequency of different virulence genes and flagellar length between exoU and exoS groups, Fisher’s exact test and unpaired t-tests were performed respectively using GraphPad prism, 2025, v8.0.2 (263). During analysis, a p-value of less than 0.05 was considered statistically significant.
Results
Difference between virulence factors genes in exoU and exoS P. aeruginosa
VFDB compared 17 virulence factors classes encompassing 327 virulence-associated genes in 20 exoU and 19 exoS P. aeruginosa keratitis isolates (Supplementary Table 4). This analysis identified significant differences in four virulence factor genes (pilA,* flaG*,* pldA*, and algP) belonging to three classes: adherence, anti-phagocytosis, and enzyme. PilA was detected in 0% of exoU isolates compared to 47% of exoS isolates (p < 0.01), flaG in 100% vs. 74% (p < 0.05), algP in 90% vs. 58% (p < 0.05), and pldA in 85% vs. 16% (p < 0.01) respectively.
Based on the PCR and VFDB results from 187 keratitis strains, the prevalence of pilA (10.9% vs. 12.9%, p > 0.05), pldA (83.6% vs. 31.8%, p < 0.01), flaG (80% vs. 55.3%, p < 0.01) and algP (90.9% vs. 81.1%, p > 0.05) differed between the exoU and exoS groups (Table 1 and Supplementary Table 5). The frequency of pldA, flaG and algP genes were higher in the exoU group and pilA was more common in the exoS group. However, differences between the two groups were statistically significant only for pldA and flaG genes (Table 1).Table 1. Differences of pilA,* pldA*,* flaG* and algP genes between exoU and exoS P. aeruginosaGenesexoU (%)exoS (%)p valueTotal isolates (N = 187)pilA10.912.9> 0.05pldA83.631.8< 0.01flaG8055.3< 0.01algP90.981.1> 0.05Indian isolates (n = 19) from whole genome sequenced datapilA050< 0.05pldA92.333.3< 0.05flaG10083.3> 0.05algP84.683.3> 0.05All Indian isolates (n = 55)pilA9.514.7> 0.05pldA77.335.3< 0.01flaG85.764.7> 0.05algP85.779.4> 0.05Australian isolates (n = 20) from whole genome sequenced datapilA046.20.05pldA71.47.7< 0.01flaG10069.3> 0.05algP10046.2< 0.05All Australian isolates (n = 132)pilA11.812.2> 0.05pldA85.330.6< 0.01flaG73.552.1< 0.05algP94.181.6> 0.05Bold indicates value that are statistically significant
Analysis of 19 Indian whole-genome sequenced P. aeruginosa (13 exoU and 6 exoS) using the VFDB revealed differences in the distribution of certain virulence-associated genes (Table 1 and Supplementary Table 6). The pilA gene was detected in 50% of exoS strains but absent in exoU strains (p < 0.05). Conversely, the pldA was more common among exoU (92.3%) compared to exoS (33.3%) (p < 0.05). 100% exoU strains had the flagellar gene flaG while 83.3% of exoS possessed this gene, though this difference was not statistically significant (p > 0.05). Similar results were observed for algP gene (84.6% in exoU vs. 83.3% exoS, p > 0.05) (Table 1).
Combined VFDB results for the 19 strains (13 exoU and 6 exoS) and a separate set of PCR results from 36 strains (8 exoU and 28 exoS) from India showed that the pldA gene was present at a significantly higher frequency in exoU (77.3%) compared to exoS isolates (35.3%) (p < 0.01) (Table 1 and Supplementary Table 5). No statistically significant differences were observed in the prevalence of other three genes (pilA, flaG, and algP). PilA was detected in 9.5% of exoU and 14.7% of exoS (p > 0.05), flaG in 85.7% vs. 64.7% (p > 0.05), and algP in 85.7% vs. 79.4% respectively (p > 0.05) (Table 1).
VFDB analysis of 7 exoU and 13 exoS P. aeruginosa isolates from Australia revealed differences in two virulence factors genes - pldA and algP (Table 1 and Supplementary Table 7). The pldA was significantly more frequent in exoU (71.4%) compared to exoS (7.7%) (p < 0.01). Similarly, algP was significantly more common in exoU (100%) than in exoS (46.2%) (p < 0.05). 46.2% of exoS carried pilA but no exoU strain was positive for this gene with the difference significant at the 10% level (p = 0.05). Although all exoU strains carried the flaG gene, its prevalence among exoS strains was 69.3%, and this difference was not statistically significant (p > 0.05) (Table 1).
Combined results from VFDB and PCR data of 132 P. aeruginosa isolates from Australia, comprising 34 exoU and 98 exoS strains, highlighted significant associations between pldA and flaG with exoU isolates in the Australian isolates, while other virulence genes had a similar distribution across both the exoU and exoS groups (Table 1 and Supplementary Table 5). The pldA gene was more common in exoU (85.3%) compared to exoS (30.6%) (p < 0.01). FlaG gene was also more common in exoU (73.5%) than in exoS (52.1%) (p < 0.05). No statistically significant differences were observed in the frequency of pilA and algP genes between the two groups. PilA was present in 11.8% of exoU and 12.2% of exoS isolates (p > 0.05), while algP was detected in 94.1% and 81.6% of isolates, respectively (p > 0.05) (Table 1).
Measurement of pldA activity
To determine the PLD activity, the association between the presence of the pldA and H₂O₂ production was assessed in a panel of 5 exoU and 5 exoS P. aeruginosa strains with and without pldA gene (Table 2). All strains carrying the pldA gene generated detectable levels of H₂O₂, ranging from 1.02 to 3.19 µM. The highest concentration of H₂O₂ was produced by exoU strain PA4 (3.19 µM), while exoS strains possessing the pldA produced comparatively lower H₂O₂ than the exoU strains. In contrast, strains lacking the pldA gene showed negligible or negative H₂O₂ values, confirming the role of pldA in phospholipase-mediated H₂O₂ generation. These findings indicated a strong correlation between pldA possession and oxidative activity, regardless of the presence of the exoU or exoS genes.Table 2PLD activity of pldA positive and negative exoU and exoS P. aeruginosa keratitis isolates. Positive values indicate detectable H₂O₂ production (µM), while negative values represent background-subtracted readings below detection limitsT3SSStrains IDpldAH₂O₂ conc (µM)exoUPA4+3.19 ± 0.16exoUPA23+2.49 ± 0.19exoSPA112+1.76 ± 0.32exoSPA114+1.02 ± 0.21exoUPA46-−12.28 ± 1.53exoUPA49-−10.41 ± 0.25exoUPA154-−8.54 ± 0.77exoSPA3-−11.72 ± 0.75exoSPA11-−11.98 ± 1.58exoSPA125-−0.06 ± 1.91 + indicates the presence of pldA gene and – indicates the absence of pldA gene, Data are presented as mean ± SD from three independent experiments (n = 3)
Determination of flagellar length of strains
TEM was used to measure flagellar lengths in a total of 10 randomly selected P. aeruginosa strains from both exoU and exoS groups (Table 3; Fig. 1). Among the exoU strains, those carrying the flaG gene (PA2 and PA23) had flagellar lengths of 7.9 ± 0.66 μm and 5.2 ± 0.58 μm, respectively. In contrast, exoU strains lacking the flaG gene (PA4, PA46, PA169) had flagellar lengths ranging from 4.9 ± 0.57 μm to 5.3 ± 0.59 μm. Like exoU strains, in exoS strains, PA11 and PA125 possessing the flaG gene showed flagellar lengths 5.1 ± 0.66 μm and 5.4 ± 0.59 μm correspondingly, while flaG-negative exoS strains (PA3, PA129, PA140) had lengths ranging from 2.6 ± 0.61 μm to 5.4 ± 0.56 μm. Most strikingly, the exoS strain PA140 (without flaG) had the shortest flagella measuring 2.6 ± 0.61 μm in length. Overall, while there was some variation between strains, no consistent increase in flagellar length was observed with the absence of the flaG gene. Statistical analysis using an unpaired t-test confirmed that the differences in flagellar lengths between flaG-positive and flaG-negative groups were not statistically significant (p > 0.05).Table 3. Flagellar lengths of flaG positive and flaG negative exoU and exoS P. aeruginosa keratitis isolatesStrains IDT3SSflaGFlagellar Length (µm)PA2exoU+7.9 ± 0.66PA23exoU+5.2 ± 0.58PA4exoU-4.9 ± 0.57PA46exoU-5.1 ± 0.50PA169exoU-5.3 ± 0.59PA11exoS+5.4 ± 0.59PA125exoS+5.1 ± 0.66PA3exoS-4.7 ± 0.55PA129exoS-5.4 ± 0.56PA140exoS-2.6 ± 0.61 + indicates the presence of* flaG* gene and – indicates the absence of* flaG* gene, Flagellar lengths are expressed as mean ± SD from 20 measurements (n = 20)Fig. 1. Transmission Electron Microscopy (TEM) of P. aeruginosa strain PA2 (exoU + and flaG+, Bar 2 μm)
Discussion
ExoU strains of P. aeruginosa are associated with more severe keratitis infection than the exoS [22, 25]. This may be due to the product of exoU which is a highly effective cytotoxin. However, there is the possibility of other virulence factors being involved. Previous studies showed that exoU strains were more resistant to the commonly prescribed antibiotics used for MK treatment because of using different resistance mechanisms, for instance possessing a greater number of mutations along with novel mutations in target genes, efflux pumps and their regulators and having more acquired resistance genes underlying resistance to antibiotics [33, 39]. However, there has been no previous comprehensive study to compare possession of many virulence factors genes between exoU and exoS P. aeruginosa in MK. This study compared the prevalence of 327 virulence factors genes from 17 different classes using VFDB and confirmed these findings using PCR to demonstrate distinct genetic virulence profiles between exoU and exoS lineages, highlighting specific virulence factors potentially contributing to differential disease outcomes in keratitis infections. Overall, exoU containing isolates more commonly possessed pldA and flaG compared to exoS strains.
The gene pldA encodes a PLD enzyme which is secreted by the haemolysin coregulated protein secretion island II- type VI secretion system (H2-T6SS) [40]. This is a novel class of T6SS effectors and the first example of a trans-kingdom virulence effector which not only helps the P. aeruginosa in internalization into non-phagocytic host cells by targeting the host PI3K (phosphoinositide 3-kinase)/Akt pathway [41, 42] but also can kill competitor bacteria [43, 44] which may aid in its colonisation. The pldA gene exerts a bactericidal effect through the breakdown of phospho-diester bond of the bacterial cell membrane phospholipid [43]. Bacteria acquire this gene horizontally as the flanking regions of pldA are highly conserved, proposing a specific insertion site [45], due to that not all P. aeruginosa possess pldA. The more common presence of pldA in exoU is consistent with a previous study that examined clinical isolates associated with infections such as septicaemia, pulmonary infections, urinary tract infections, and chronic infections in cystic fibrosis patients, along with environmental strains where the pldA was present in hypervirulent and multi-drug resistant exoU isolates responsible for severe acute infections [26].
Given the role of pldA in enhancing cytotoxicity and promoting competitive fitness through T6SS-mediated effector delivery, its higher prevalence in exoU strains suggests a possible synergistic role in exacerbating keratitis severity. The concurrent presence of both exoU, a potent type III secretion system effector, and pldA, a T6SS-associated virulence factor, may enhance the capacity of these strains in the lysis of epithelial cells, immune evasion, and bacterial competition within the ocular environment. Future functional studies, including targeted knockouts of pldA in representative exoU keratitis strains, are warranted to elucidate its specific contribution to the pathogenesis of keratitis in animal models. Such investigations would also provide insight about the interplay between T3SS and T6SS effectors in ocular infections and potentially identify new targets for adjunctive therapeutic interventions.
The flaG gene was more common in exoU, though its prevalence varied between regions. In the Australian collection, the flaG gene was more common in the exoU group, while in the Indian cohort, although the gene was more common overall, it did not reach statistical significance in the comparison between the groups. The flaG gene is considered to have role in bacterial filament length control by maintaining the normal filament length through a mechanism that is still not known [46]. Deletion of flaG from P. fluorescens resulted in longer flagella without changing the motility and colonization properties of the strain [47]. To explore the functional consequence of flaG carriage, flagellar lengths were assessed from both the exoU and exoS strains with and without flaG gene by TEM. In contrast to previous literature [47], there was no significant difference in flagellar lengths between flaG-positive and flaG-negative strains within either exoU or exoS groups. While some individual variations were observed, including notably shorter flagella in a flaG-negative exoS strain PA140, the lack of consistent correlation implies that other regulatory factors may compensate for the absence of the flaG gene in flagellar biogenesis or that its role in ocular isolates may differ from strains studied in other infection sites and in other species of Pseudomonas. Further studies with a larger sample size and complementary assays such as flagellar gene expression analysis or study of the isogenic mutant are warranted to clarify this relationship.
Type IV pili plays a vital role in P. aeruginosa motility by enabling twitching movement through the extension and retraction of surface appendages which allows the bacteria to migrate across smooth surfaces and contributes to bacterial colonisation [48]. In P. aeruginosa, Type IV pili are assembled from pilin, a 15-kDa protein encoded by the pilA gene, which is essential for the formation of long pili filaments involved in bacterial virulence [48–50]. Before infection or corneal epithelial cell killing, both exoU and exoS P. aeruginosa strains use twitching motility to travel along the basolateral membranes of epithelial cells [51]. Through this process, Type IV pili can bind to asialo-GM1 receptors on the surface of epithelial cells leading to either bacterial internalisation or cytotoxicity [52]. Previous studies have shown that the pilA mutant can still exhibit twitching motility, invade corneal epithelial cells, and accumulate intracellularly in greater numbers than wild-type P. aeruginosa [53]. In the present study, the pilA gene was predominantly detected in exoS isolates, as confirmed by both whole-genome sequencing and PCR analysis of Australian and Indian strains. This result indicated an association between pilA and exoS genes. Previous studies reported that Type IV pili are necessary for the injection of exoS into epithelial cells [54]. The observed association between exoS strains and adhesion-related mechanisms suggests that exoS strains may rely more on pilus-mediated attachment and internalisation, potentially compensating for the absence of exoU-mediated rapid cytotoxicity.
Conclusion
ExoU keratitis strains possessed a distinct virulence gene repertoire compared to exoS strains, particularly enriched for pldA and, in certain settings, flaG, which may contribute to a more severe disease outcome. Conversely, exoS strains more commonly had the adhesion-associated factor pilA. Clinical data were not available to explore association between disease severity and the presence of the virulence factors. Incorporating detailed clinical data in future studies could further help correlate virulence gene profiles with disease severity. In addition, future studies should focus on generating targeted gene knockouts of pldA, flaG, and pilA in representative exoU and exoS keratitis strains and see their effects both in vitro and in vivo models to directly assess their contribution in pathogenesis of keratitis. These functional studies are essential to validate the genomic associations identified in this study which could ultimately identify novel targets for anti-virulence therapies in MK.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1 (DOCX 50.0 KB)
Supplementary Material 2 (DOCX 25.9 KB)
Supplementary Material 3 (XLSX 16.3 KB)
Supplementary Material 4 (XLSL 120 KB)
Supplementary Material 5 (XLSL 19.3 KB)
Supplementary Material 6 (XLSL 61.1 KB)
Supplementary Material 7 (XLSL 62.6 KB)