Morphological variation in the Rhododendron pseudochrysanthum Hayata (Ericales, Ericaceae) species complex from Taiwan
Ya-Rong Zheng, Yen-Hsueh Tseng, Hsy-Yu Tzeng

TL;DR
This paper identifies a distinct subspecies of Rhododendron pseudochrysanthum in Taiwan based on unique leaf, flower, and seed traits.
Contribution
The study provides new evidence for taxonomic differentiation within the R. pseudochrysanthum species complex using morphological and statistical analysis.
Findings
R. pseudochrysanthum ssp. morii var. taitunense is distinct in leaf, flower bud, pollen, and seed morphology.
The taxon has a restricted distribution in low-elevation northern Taiwan.
Statistical analysis confirms morphological differentiation from other members of the species complex.
Abstract
Our study examines the morphological and statistical differentiation within the Rhododendron pseudochrysanthum species complex through comparative analyses of macro- and micro-morphological characters. Using significance testing and cluster analysis, our results demonstrate that R. pseudochrysanthum Hayata ssp. morii (Hayata) Yamazaki var. taitunense Yamazaki is distinct from other members of the complex, namely R. morii Hayata, R. pseudochrysanthum Hayata, and R. hyperythrum Hayata. This taxon is characterized by glabrous mature leaves with revolute margins, larger flower buds with an elongated conical shape, larger pollen and seed sizes, and distinct pollen and seed morphology. Furthermore, R. pseudochrysanthum ssp. morii var. taitunense exhibits a restricted and localized distribution, occurring exclusively in low-elevation mountainous areas of Northern Taiwan. Overall, our results…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1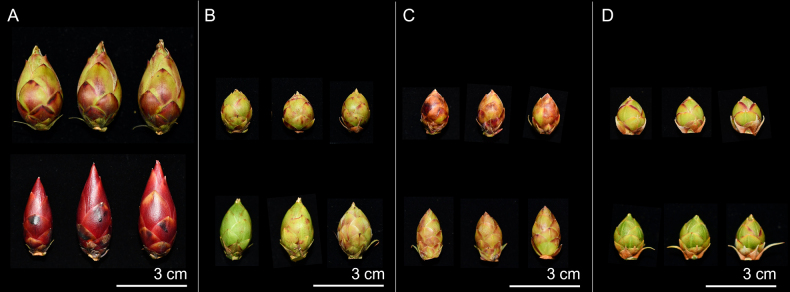 Figure 2
Figure 2 Figure 3
Figure 3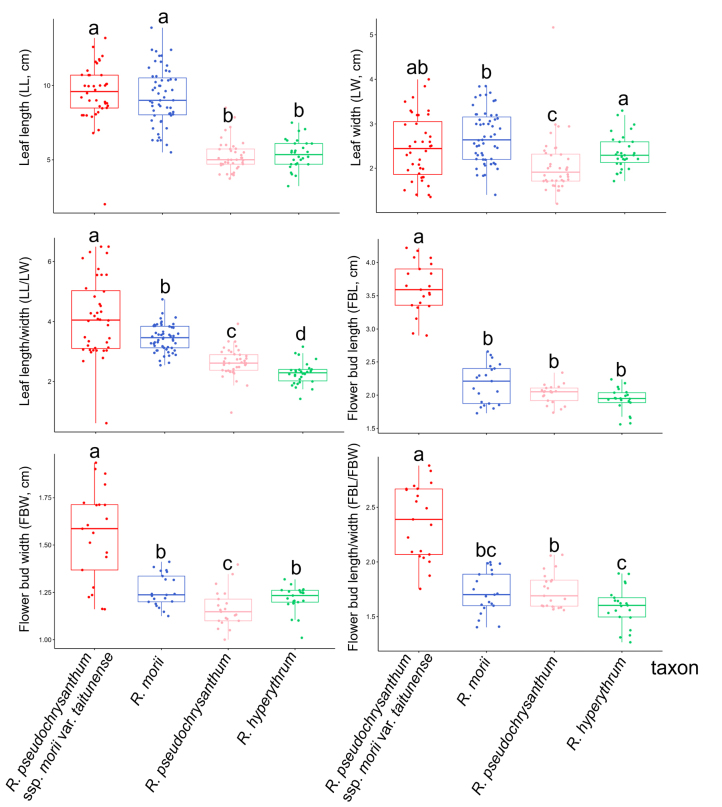 Figure 4
Figure 4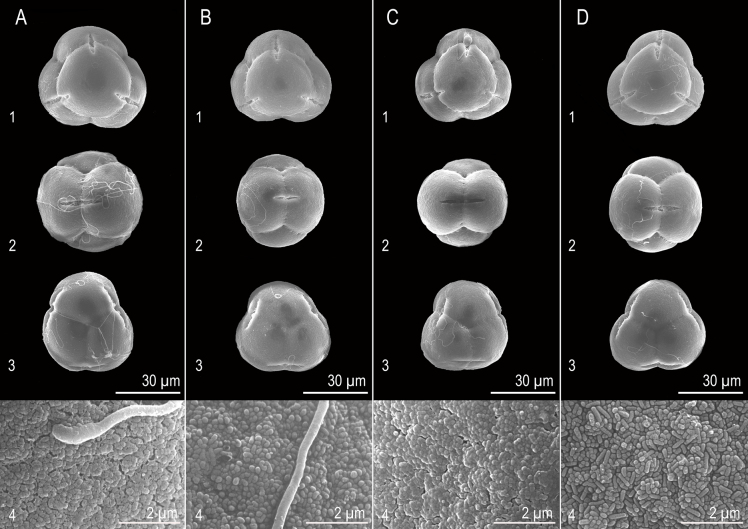 Figure 5
Figure 5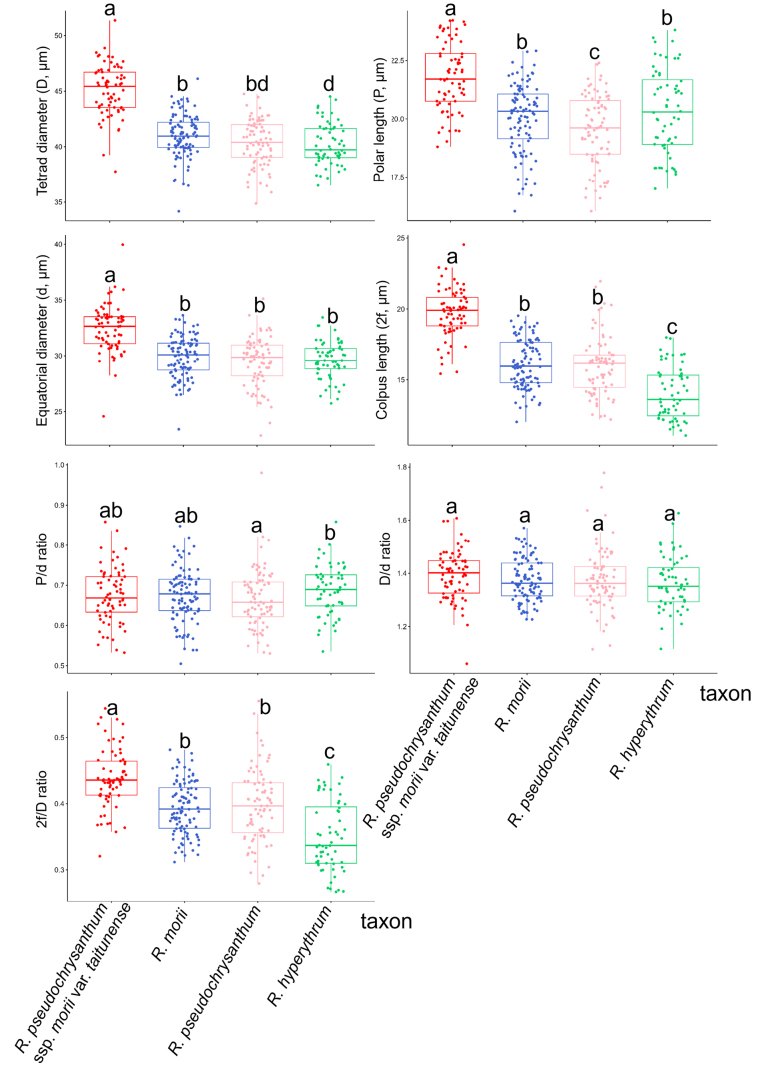 Figure 6
Figure 6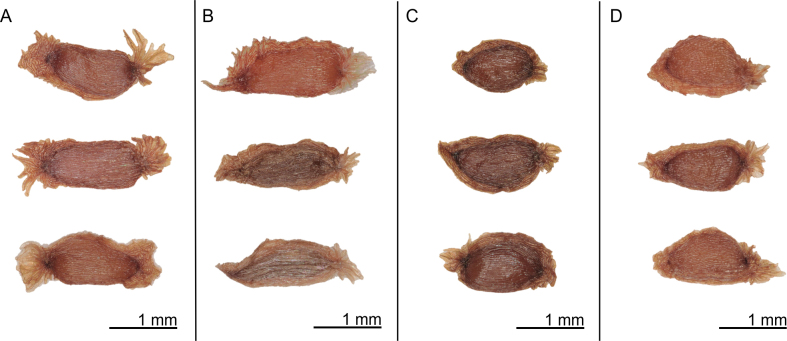 Figure 7
Figure 7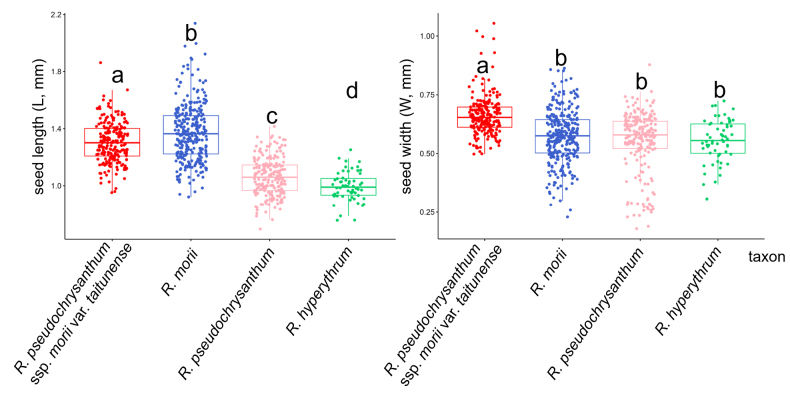 Figure 8
Figure 8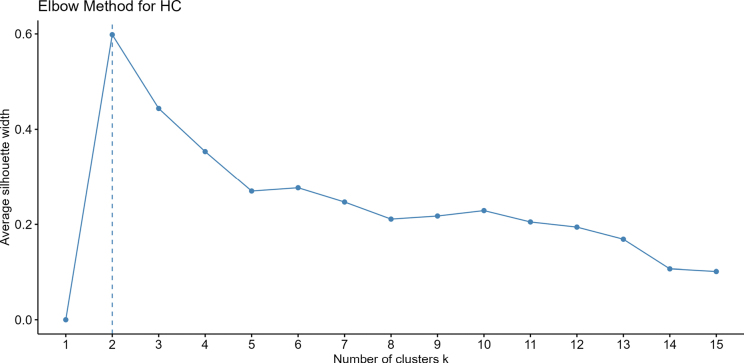 Figure 9
Figure 9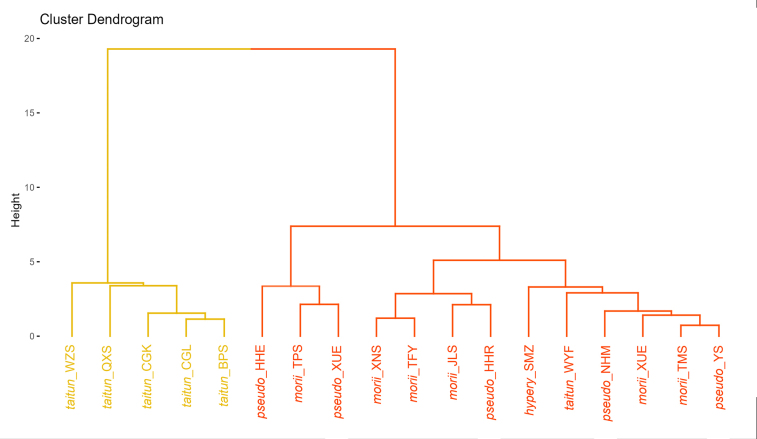 Figure 10
Figure 10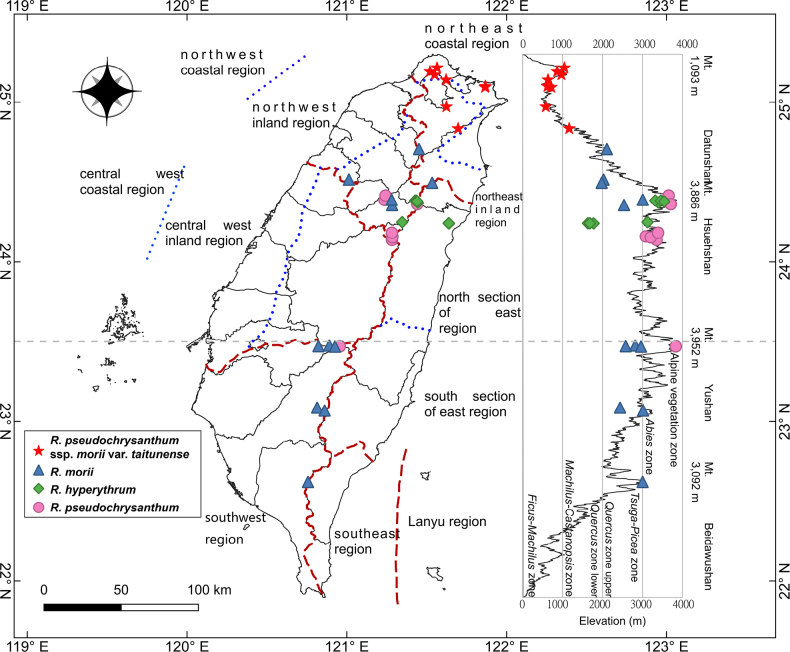 Figure 11
Figure 11| taxon | location | code | lat. (°N), long. (°E) | altitude (m) | Specimens examined |
|---|---|---|---|---|---|
| Mt. Qixing | taitun_QXS |
| 972 | ZYR298 | |
| Mt. Wuzhi | taitun_WZS |
| 635 | ZYR257 | |
| Mt. Bijia | taitun_BJS |
| 580 | ZYR266 | |
| Mt. Caigongkeng | taitun_CGK |
| 864 | ZYR271 | |
| Mt. Banping | taitun_BPS |
| 621 | ZYR332 | |
| Mt. Canguangliao | taitun_CGL |
| 717 | ZYR270 | |
| Mt. Xin | taitun_XS |
| 481 | ZYR335 | |
|
| Mt. Jiali | morii_JLS |
| 2,026 | ZYR30 |
| Yushan Trail 5 km | morii_YS |
| 2,963 | ZYR280 | |
| Tataka | morii_TTK |
| 2,820 | ZYR275 | |
| Mt. Taman | morii_TMS |
| 2,109 | ZYR222 | |
| Mt. Xinan | morii_XNS |
| 2,439 | ZYR345 | |
| Beinanzhu | morii_BNS |
| 2,551 | ZYR337 | |
| Tefuya | morii_TFY |
| 2,580 | ZYR375 | |
| Mt. Xue 4 km | morii_XUE |
| 3,007 | ZYR69 | |
| Mt. Taiping | morii_TPS |
| 1,979 | ZYR66 | |
|
| Hehuan East Peak | pseudo_HHE |
| 3,353 | ZYR297 |
| Hehuan Road Side | pseudo_HHR |
| 3,084 | ZYR350 | |
| Mt. Xue | pseudo_XUE |
| 3,596 | ZYR113 | |
| Yushan Main Peak | pseudo_YS |
| 3,834 | ZYR86 | |
| Nanhu Main Peak | pseudo_NHM |
| 3,709 | ZYR366 | |
|
| Nanhu Cirque | hypery_NHC |
| 3,400 | ZYR364 |
| Nanhu 18.6 km | hypery_NH18k |
| 3,460 | ZYR367 | |
| Nanhu 17 km | hypery_NH17k |
| 3,320 | ZYR370 | |
| Shenmazheng | hypery_SMZ |
| 3,457 | ZYR362 | |
| Wuyan Peak | hypery_WYF |
| 3,544 | ZYR369 |
| taxon |
|
|
| |
|---|---|---|---|---|
| traits | ||||
| Leaf length | 9.482 ± 1.203a | 9.264 ± 1.641a | 5.272 ± 0.761b | 5.370 ± 0.988b |
| Leaf width | 2.465 ± 0.589ab | 2.713 ± 0.484b | 2.080 ± 0.408c | 2.371 ± 0.393a |
| Leaf length/width | 4.141 ± 1.186a | 3.454 ± 0.367b | 2.634 ± 0.307c | 2.287 ± 0.383b |
| Leaf lateral vein pairs | 10–15 | 12–19 | 12–15 | 11–14 |
| Leaf shape | narrowly elliptic- oblanceolate | narrowly elliptic- lanceolate | narrowly ovate | narrowly ovate |
| Leaf margin | strongly revolute | flat to revolute | revolute | revolute |
| Leaf base | obtuse-acute | obtuse-acute | obtuse-acute | obtuse-cordate |
| Leaf trichome | hairless | tufted hair on abaxial midrib | glandular hair on abaxial midrib | branched hair on leaf abaxial |
| Flower bud length | 3.615 ± 0.372a | 2.170 ± 0.288b | 2.019 ± 0.141b | 1.928 ± 0.178b |
| Flower bud width | 1.554 ± 0.240a | 1.260 ± 0.083b | 1.165 ± 0.096c | 1.215 ± 0.069b |
| Flower bud length/width | 2.367 ± 0.331a | 1.722 ± 0.190bc | 1.742 ± 0.162b | 1.592 ± 0.173c |
| Style hair type | glandular hairs | glandular or puberulent | glandular or puberulent | glandular or puberulent |
| Ovary hair type | glandular hairs | glandular or puberulent | glandular or puberulent | glandular or puberulent |
| calyx | lobes 5, triangular, with glandular hairs | lobes 5, triangular, with glandular hairs | lobes 5, triangular, with glandular hairs | lobes 5, triangular, with glandular hairs |
| Pedicel hair type | glandular hairs | glandular or puberulent | glandular or puberulent | glandular or puberulent |
| Locule number | 6–10 | 6–8 | 6–7 | 6–7 |
| Stamen number | 10–12 | 10–12 | 10–12 | 10–12 |
| taxon |
|
|
| |
|---|---|---|---|---|
| traits | ||||
| Tetrad diameter (D) | 45.090 ± 1.096a | 40.988 ± 0.896b | 40.369 ± 0.882bd | 40.243 ± 0.848d |
| Polar length (P) | 21.804 ± 0.638a | 20.050 ± 0.569b | 19.525 ± 0.465c | 20.325 ± 0.403b |
| Equatorial diameter (d) | 32.462 ± 1.622a | 29.894 ± 1.139b | 29.469 ± 1.464b | 29.621 ± 0.675b |
| Colpus length (2f) | 19.723 ± 0.764a | 16.122 ± 0.746b | 15.991 ± 1.332b | 14.035 ± 1.362c |
| P/d ratio | 0.675 ± 0.048ab | 0.673 ± 0.034ab | 0.666 ± 0.038a | 0.687 ± 0.020b |
| D/d ratio | 1.393 ± 0.062a | 1.374 ± 0.045a | 1.376 ± 0.050a | 1.363 ± 0.042a |
| 2f/D ratio | 0.439 ± 0.019a | 0.394 ± 0.016b | 0.397 ± 0.032b | 0.350 ± 0.037c |
| apocolpial exine sculpture | gemmate | gemmate | gemmate | gemmate |
| taxon |
|
|
| |
|---|---|---|---|---|
| traits | ||||
| Seed length | 1.303 ± 0.070a | 1.371 ± 0.146b | 1.062 ± 0.095c | 0.996 ± 0.102d |
| Seed width | 0.662 ± 0.058a | 0.574 ± 0.092b | 0.552 ± 0.121b | 0.552 ± 0.094b |
| Seed shape | narrowly elliptic | narrowly elliptic | elongate | elongate |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBioactive Natural Diterpenoids Research · Plant Diversity and Evolution · Biological and pharmacological studies of plants
Introduction
Rhododendron subg. Hymenanthes (Blume) K.Koch belongs to the genus Rhododendron L. in the family Ericaceae Juss. They are also known as elepidote rhododendrons and they account for approximately one-quarter of the species diversity within the genus. This subgenus is classified into one section and 24 subsections (Chamberlain 1982), comprising around 270 species worldwide. It is primarily distributed in Asia, with additional occurrences in Europe and North America. China has 259 Rhododendron subg. Hymenanthes species (Fang et al. 2005). Most phylogenetic studies indicate that this subgenus forms a monophyletic group (Kron 1997; Kurashige et al. 1998, 2001; Milne 2004).
Rhododendron rubropunctatum Hayata was described by Hayata (1913) based on a specimen collected by S. Sasaki from Chihsingshan in Northern Taiwan. This taxon was later reported from Taiwan by several authors, including Kanehira (1917), Sasaki (1928), Masamune (1936), Kanehira (1936), and Masamune (1954). However, Wilson (1925), Li (1963), Hsu (1973), Ying (1976), and Li (1978) observed that both R. rubropunctatum Hayata and R. hyperythrum Hayata possess prominent glandular structures on the abaxial leaf surface, and consequently treated R. rubropunctatum Hayata as a synonym of R. hyperythrum Hayata.
In 1981, Yamazaki noted that a Rhododendron species occurring in the Tatun Volcanic Group of Northern Taiwan, including on the slopes of Chihsingshan and Tatunshan, was initially published as R. rubropunctatum by Hayata (1913). He pointed out that this name is a later homonym of R. rubropunctatum H. Lév. & Vaniot (1911), and thus, illegitimate. Yamazaki also noted that Wilson (1925) replaced the name with R. hyperythrum Hayata, which led to further taxonomic confusion (Yamazaki 1981). Nevertheless, due to the revolute leaf margins distinct from other taxa, Yamazaki renamed the taxon as R. pseudochrysanthum Hayata ssp. morii (Hayata) Yamazaki var. taitunense Yamazaki.
Later, Lu and Yang (1989) and Li et al. (1998) considered the reddish glands on the abaxial leaf surface to lack taxonomic significance and treated R. rubropunctatum Hayata, R. morii Hayata, and R. pseudochrysanthum Hayata ssp. morii (Hayata) Yamazaki var. taitunense Yamazaki as synonyms of R. pseudochrysanthum Hayata. In the same treatment, Li et al. (1998) recognized only three taxa in the Rhododendron subgenus Hymenanthes in Taiwan: R. formosanum Hemsl., R. hyperythrum Hayata, and R. pseudochrysanthum Hayata.
Furthermore, due to continuous morphological variation and the absence of clear genetic differentiation among R. pseudochrysanthum Hayata ssp. morii (Hayata) Yamazaki var. taitunense Yamazaki, R. morii Hayata, R. pseudochrysanthum Hayata, and R. hyperythrum Hayata, these four taxa have been collectively referred to as the Rhododendron pseudochrysanthum species complex (Lu and Yang 1989; Chung et al. 2007; Hsieh 2007; Liang 2011; Chen et al. 2014; Cao et al. 2022).
Pollen morphology is considered to exhibit a high degree of genetic conservatism, with intrinsic features such as shape, surface ornamentation, and the number of germination pores commonly used for plant classification and identification (Zhou et al. 2008). Moreover, it is informative for both taxonomic delimitation and phylogenetic analysis within certain groups of Ericaceae (Kron et al. 2002; Sarwar 2007). Within a single subgenus, pollen characters can be particularly valuable for the recognition of sections and subsections, for instance, in Rhododendron sect. Ponticum G. Don, although all taxa produce tetrad pollen grains, variations in pollen size and exine sculpture have been reported, reflecting meaningful interspecific differentiation (Huang 1972; Vasanthy and Pocock 1987; Fuhsiung et al. 1995; Mao et al. 2000; Terzioğlu et al. 2001; Gao et al. 2002; Wang et al. 2006; Zhang et al. 2009; Miyoshi et al. 2011). In Rhododendron species, seed morphology, size, and the presence or absence of appendages are closely related to plant habits, primarily reflecting the systematic differentiation among taxa within the genus (Geng 2014). Moreover, the width and shape of seed wings vary significantly across different groups (Ding et al. 1995; Wang et al. 2007).
Previous studies on the Rhododendron pseudochrysanthum species complex have focused primarily on external morphological descriptions. In this study, we re-examine morphological differentiation within the complex by incorporating a broader set of macromorphological characters, as well as pollen and seed morphological traits.
Materials and methods
Samples collection and morphological comparison
The materials used in this study were primarily collected from wild populations of the R. pseudochrysanthum species complex in Taiwan (Table 1). Fieldwork included photography, GPS point recording, and fixation of fresh leaves and flowers. External morphological traits, such as leaves, flowers, floral buds, and seeds, were observed and compared among the different taxa. Macroscopic features were documented using a digital camera equipped with a macro lens (Nikon D7500 + Nikon AF-S DX NIKKOR 40 mm f/2.8G Micro). At the same time, finer structures were examined under a high-magnification stereomicroscope (KEYENCE VHX-7020).
Table 1.: Materials collected of the R. pseudochrysanthum species complex.
Herbarium resources
Herbarium acronyms from Index Herbariorum were used in this study (Thiers 2026, continuously updated). Voucher specimens collected for the current study were deposited in the herbarium of the Department of Forestry, National Chung-Hsing University, Taiwan (TCF). Physical and digital specimens in several herbaria were also examined; physical specimens: Taiwan Forestry Research Institute (TAIF), Herbarium of Taiwan Biodiversity Research Institute, Taiwan (TAIE), National Taiwan University, Taiwan (TAI), National Chung-Hsing University, Taiwan (TCF); and digital specimens: Department of Forestry and Natural Resources, National Chia-Yi University, Taiwan (CHIA), Provincial Pingtung Institute, Taiwan (PPI), the Royal Botanic Gardens, Kew (K), Herbarium of Academia Sinica, Taiwan (HAST), and Herbarium of the University of Tokyo, Japan (TI).
Pollen morphology
Fresh stamens were collected and fixed in 70% ethanol. During the collection, pollen was extracted from the anthers and placed into a 1.5 ml centrifuge tube containing 70% ethanol. After a single centrifugation step, the supernatant was removed, and a mixture of 30 ml 2,2-dimethoxypropane (DMP) and 1 drop of 1M HCl was added. The sample was incubated for approximately 2 hours, followed by an acetone substitution for at least 1 hour. Subsequently, the samples underwent critical point drying (CPD) (Halbritter 1998). The mounted samples were then gold-coated for 100 seconds at 15 mA using a Quorum SC7620 sputter coater (Quorum Technologies, Laughton, UK). Finally, observations and imaging were performed using a scanning electron microscope (Hitachi S-3400N) at 15 kV.
The morphological characteristics of pollen were described with reference to Erdtman (1952), Sarwar (2007), and Sarwar and Takahashi (2013). Observations included the shape and size of tetrad pollen, as well as the following measurements (in µm): Tetrad diameter (D), Polar length (P), Equatorial diameter (d for tetrads; E for monads), Colpus length (2f for tetrads; L for monads), P/d ratio (polar to equatorial diameter ratio), D/d ratio (tetrad diameter to equatorial diameter ratio), 2f/D ratio (colpus length to tetrad diameter ratio), and Apocolpial exine sculpture.
Seed morphology
Mature dehiscent capsules of each taxon were collected from various sampling sites. A total of 3–5 capsules were placed in an oven at 50 °C for at least 48 hours. Thirty mature seeds were randomly sampled for each taxon and observed under a high-magnification dissecting microscope (KEYENCE VHX-7020) to examine and document seed size and appendages. The morphological characteristics of the seeds were described with reference to Barthlott (1981, 1990) and Geng (2014), and the following measurements (in mm) were noted: seed length (L) and seed width (W).
Distribution map
The collection site information of this study, including GPS coordinates and elevation, was integrated. The distribution map was adapted from Lin (2018), and the distribution points of each taxon were plotted using QGIS ver. 3.4. Additionally, the study referred to the geographical and climatic regions of Taiwan as defined by Su (1985) and the montane vegetation zones to analyze the vertical and horizontal distribution patterns of each taxon.
Data analysis
Quantitative traits were measured using ImageJ software. Statistical analyses were conducted in R 4.3.2 using the stats package (R Core Team 2024). First, normality was assessed using the Shapiro-Wilk test (shapiro.test) to determine whether the quantitative trait values followed a normal distribution across taxa (p < 0.05 indicates non-normal distribution). If traits followed a normal distribution, one-way ANOVA was performed using the aov function, followed by a Tukey HSD post-hoc test (TukeyHSD, p ≤ 0.05) to assess significant differences among taxa. If traits did not follow a normal distribution, nonparametric analysis was conducted using the Kruskal-Wallis test (kruskal.test) to determine differences among taxa, followed by a pairwise Wilcoxon rank-sum test (pairwise.wilcox.test, p ≤ 0.05) for post-hoc comparisons. Boxplots were generated using the ggboxplot function from the ggplot2 package (Wickham 2016) to visualize data distribution.
To investigate the clustering results of taxa based on characteristic variables such as pollen and seeds, cluster analysis was conducted using R 4.3.2. The stats package (R Core Team 2024) was used with the dist function to construct a distance matrix based on Euclidean distance to determine the proximity between data points. Hierarchical cluster analysis was then performed using the hclust function with Ward’s minimum variance method. Finally, a dendrogram was plotted using the plot function from the graphics package (R Core Team 2024).
Results and discussion
Macromorphological differences
The leaf morphology of the R. pseudochrysanthum species complex is highly variable (Fig. 1, Table 2), exhibiting shapes such as narrow elliptic, lanceolate, oblanceolate, and narrow ovate. Leaf base morphology is also diverse, and many traits show considerable intraspecific variation. Although leaf length differs significantly among some taxa (Fig. 4, Table 2), leaf size and shape can be influenced by habitat conditions or climate, and even within a single individual, variation may occur due to light exposure (Geng 2014). Therefore, leaf size, shape, and base morphology alone cannot be used as reliable taxonomic criteria. In terms of epidermal appendages, glands were present across all four taxa, consistent with findings by Lu and Yang (1989), indicating that this trait is not a valid diagnostic character. However, trichomes exhibited high diversity among the four taxa, suggesting that this macromorphological feature may serve as a useful characteristic for taxonomic identification.
Leaf morphology of Rhododendron pseudochrysanthum species complex. A. R. pseudochrysanthum Hayata ssp. morii (Hayata) Yamazaki var. taitunense Yamazaki; B. R. morii Hayata; C. R. pseudochrysanthum Hayata; D. R. hyperythrum Hayata.
Table 2.: Macromorphological differences within the Rhododendron pseudochrysanthum species complex.
Within the R. pseudochrysanthum species complex, traits such as the indumentum on the style, ovary, calyx, pedicel, and stamens, as well as the number of ovary locules, exhibit variation among individuals within each taxon. Floral color also varies depending on environmental conditions. However, R. pseudochrysanthum ssp. morii var. taitunense consistently possesses a higher number of ovary locules compared to the other taxa. (Fig. 3, Table 2). Therefore, some of these characters are not suitable as diagnostic features for species delimitation within the species complex. However, in terms of floral bud size, R. pseudochrysanthum ssp. morii var. taitunense displays significantly greater bud length, width, and length-to-width ratio (p < 0.05) compared to the other taxa (Figs 2, 4). In contrast, R. morii, R. pseudochrysanthum, and R. hyperythrum show no significant differences in floral bud dimensions (Fig. 4, Table 2).
Flower bud morphology of Rhododendron pseudochrysanthum species complex. A. R. pseudochrysanthum Hayata ssp. morii (Hayata) Yamazaki var. taitunense Yamazaki; B. R. morii Hayata; C. R. pseudochrysanthum Hayata; D. R. hyperythrum Hayata.
Flower morphology of Rhododendron pseudochrysanthum species complex. A. R. pseudochrysanthum Hayata ssp. morii (Hayata) Yamazaki var. taitunense Yamazaki; B. R. morii Hayata; C. R. pseudochrysanthum Hayata; D. R. hyperythrum Hayata: 1, flower; 2, style; 3, ovary; 4, ovary locules; 5, calyx; 6, pedicel.
Boxplot comparing the leaf and flower bud characteristics of Rhododendron pseudochrysanthum species complex. Different letters in each boxplot indicate statistically significant differences (p < 0.05).
Pollen morphology
The pollen grains of the R. pseudochrysanthum species complex are all tetrads, arranged in a tetrahedral configuration, and exhibit heteropolarity (Fig. 5). The average diameter of the tetrads ranges from 40.243 ± 0.848 µm to 45.090 ± 1.096 µm; the average polar axis length ranges from 19.525 ± 0.465 µm to 21.804 ± 0.638 µm; the average equatorial diameter ranges from 29.469 ± 1.464 µm to 32.462 ± 1.622 µm; and the average colpus length ranges from 14.035 ± 1.362 µm to 19.723 ± 0.764 µm (Table 3). The P/d ratio ranges from 0.666 ± 0.038 to 0.687 ± 0.020; the D/d ratio ranges from 1.363 ± 0.042 to 1.393 ± 0.062; and the 2f/D ratio ranges from 0.350 ± 0.037 to 0.439 ± 0.019 (Table 3). According to Erdtman (1952), based on the classification of pollen grain size and shape, these pollen grains are categorized as medium-sized (Mediae, 25–50 µm) and oblate in shape. They are 3-colporate, with a circular outline in polar view.
Pollen morphology of the Rhododendron pseudochrysanthum species complex. A. R. pseudochrysanthum Hayata ssp. morii (Hayata) Yamazaki var. taitunense Yamazaki; B. R. morii Hayata; C. R. pseudochrysanthum Hayata; D. R. hyperythrum Hayata: 1, tetrads polar plane; 2, tetrads equatorial plane; 3, tetrads polar plane; 4, apocolpial exine sculpture.
Table 3.: Pollen morphological differences of the R. pseudochrysanthum species complex.
Overall, R. pseudochrysanthum ssp. morii var. taitunense exhibits significant differences (p < 0.05) in all quantitative pollen traits (except for the P/d and D/d ratios) compared to the other three taxa within the R. pseudochrysanthum species complex (Fig. 6). According to the study by Sarwar and Takahashi (2013), the apocolpial exine sculpture of pollen in the R. pseudochrysanthum species complex is consistently of the gemmate type (Fig. 5).
Boxplot comparing the pollen grain characteristics of the R. pseudochrysanthum species complex. Different letters in each boxplot indicate statistically significant differences (p < 0.05).
Seed morphology
The fruit of the R. pseudochrysanthum species complex is a septicidal capsule, and the seeds are elongate or narrowly elliptic in shape (Fig. 7). The average seed length ranges from 1.062 ± 0.095 mm to 1.371 ± 0.146 mm, and the average seed width ranges from 0.552 ± 0.121 mm to 0.662 ± 0.058 mm (Table 4). Within the R. pseudochrysanthum species complex, R. pseudochrysanthum ssp. morii var. taitunense shows significant differences (p < 0.05) in both seed length and width when compared to R. morii, R. pseudochrysanthum, and R. hyperythrum (Fig. 8). However, no significant differences in seed width were observed among R. morii, R. pseudochrysanthum, and R. hyperythrum.
Seed morphology of the Rhododendron pseudochrysanthum species complex. A. R. pseudochrysanthum Hayata ssp. morii (Hayata) Yamazaki var. taitunense Yamazaki; B. R. morii Hayata; C. R. pseudochrysanthum Hayata; D. R. hyperythrum Hayata.
Boxplot comparing the seed characteristics of the R. pseudochrysanthum species complex. Different letters in each boxplot indicate statistically significant differences (p < 0.05).
Table 4.: Seed morphological differences of the R. pseudochrysanthum species complex.
As elevation increases, wind becomes stronger, facilitating more effective seed dispersal. Therefore, Rhododendron species at higher elevations often possess narrower seeds and seed wings. Since stronger winds at high altitudes can effectively disperse seeds, there is less selective pressure to develop elaborate dispersal structures (Howe and Smallwood 1982). Nonetheless, R. pseudochrysanthum and R. hyperythrum, both distributed at high elevations, still exhibit prominent wing-like seed appendages (Fig. 7).
Cluster analysis
To determine the optimal number of clusters, we employed the Elbow Method in conjunction with average silhouette width as an evaluation metric, using hierarchical clustering (HC) analysis. The results indicated that when the number of clusters was set to two, the average silhouette width reached its maximum value (approximately 0.8; Fig. 9), suggesting the highest intra-cluster similarity and inter-cluster dissimilarity at this point. Therefore, based on this analysis, dividing the samples into two clusters was considered the optimal solution, further supporting a clear distinction between R. pseudochrysanthum ssp. morii var. taitunense and the other taxa in terms of pollen and seed characteristics (Fig. 10). This result indicates that R. pseudochrysanthum ssp. morii var. taitunense is distinct from the other three taxa within the R. pseudochrysanthum species complex in terms of reproductive traits.
Elbow Method for Determining Optimal Number of Clusters in Hierarchical Clustering. Number of clusters k = 2.
Cluster analysis of reproductive traits. The yellow lines are R. pseudochrysanthum ssp. morii var. taitunense, and the orange lines are R. morii, R. pseudochrysanthum, and R. hyperythrum.
Distribution and habitat
Within the Rhododendron pseudochrysanthum species complex, R. pseudochrysanthum ssp. morii var. taitunense shows a distinct geographic distribution compared with other taxa, occurring in the Datun Volcanic Group, Keelung Volcanic Group, Shiding District, and Wulai District at elevations of 440–1,200 m (Fig. 11). Currently, it is known from Mt. Qixing and Mt. Wuzhi in Taipei City, as well as Mt. Zhuzi, Mt. Caigongkeng, Mt. Xin, Mt. Canguangliao, Mt. Banping, Mt. Bijia, and Mt. Hongludi in New Taipei City. This taxon primarily grows along ridges, cliff edges, or in open forest habitats, whereas the other three taxa are mainly distributed in mid- to high-elevation mountainous regions of Taiwan.
Distribution map of Rhododendron pseudochrysanthum species complex in Taiwan.
Conclusions
In summary, based on the literature review, the geographical distribution, and morphological analyses, we found that R. pseudochrysanthum ssp. morii var. taitunense is clearly distinguishable from other taxa within the R. pseudochrysanthum species complex by its strongly revolute mature leaves, larger flower buds with a long conical shape (over 3 cm), and larger pollen grains. Furthermore, the restricted and localized distribution of this taxon solely to the low-elevation mountainous areas of Northern Taiwan further supports its distinctness.
In addition, we found that R. morii, R. pseudochrysanthum, and R. hyperythrum are difficult to delineate taxonomically based on certain morphological characteristics, such as floral morphology, pollen, and seed. This complexity is compounded by the occurrence of intermediate individuals between R. pseudochrysanthum and R. hyperythrum in high-elevation areas, making species delineation challenging. Therefore, more in-depth studies on these remaining taxa are planned for the future, including expanding sampling efforts and incorporating molecular evidence.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barthlott W (1981) Epidermal and seed surface characters of plants: Systematic applicability and some evolutionary aspects. Nordic Journal of Botany 1: 345–355. 10.1111/j.1756-1051.1981.tb 00704.x · doi ↗
- 2Cao JJ, Li YS, Chang CT, Chung JD, Hwang SY (2022) Adaptive divergence without distinct species relationships indicate early stage ecological speciation in species of the Rhododendron pseudochrysanthum complex endemic to Taiwan. Plants 11: 1226. 10.3390/plants 11091226 PMC 910153035567227 · doi ↗ · pubmed ↗
- 3Chamberlain DF (1982) A revision of Rhododendron II subgenus Hymenanthes. Notes from the Royal Botanic Garden Edinburgh 39: 270–486. 10.24823/nrbge.1982.3252 · doi ↗
- 4Chen CY, Liang BK, Chung JD, Chang CT, Hsieh YC, Lin TC, Hwang SY (2014) Demography of the upward-shifting temperate woody species of the Rhododendron pseudochrysanthum complex and ecologically relevant adaptive divergence in its trailing edge populations. Tree Genetics & Genomes 10: 111–126. 10.1007/s 11295-013-0669-x · doi ↗
- 5Chung JD, Lin TP, Chen YL, Cheng YP, Hwang SY (2007) Phylogeographic study reveals the origin and evolutionary history of a Rhododendron species complex in Taiwan. Molecular Phylogenetics and Evolution 42: 14–24. 10.1016/j.ympev.2006.06.02717070712 · doi ↗ · pubmed ↗
- 6Ding BY, Wu HX, Zhang HM, Fang YY (1995) Seed morphology of Rhododendron (Ericaceae) from Zhejian and its taxonomic significance. Acta Botanica Boreali-Occidentalia Sinica 15(6): 36–42.
- 7Erdtman G (1952) Pollen Morphology and Plant Taxonomy-Angiosperms. Almqvist and Wiksell, Stockholm, 539 pp. 10.1080/11035895209453507 · doi ↗
- 8Fang MY, Fang RZ, He MY, Hu LZ, Yang HB, Chamberlain DF (2005) Rhododendron. Flora of China. http://www.efloras.org/flora_page.aspx?flora_id=2