A new cryptic species of Chelidurella Verhoeff, 1902 (Dermaptera, Forficulidae) from the Italian Alps: molecular evidence reveals hidden diversity in a high-altitude refugium

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
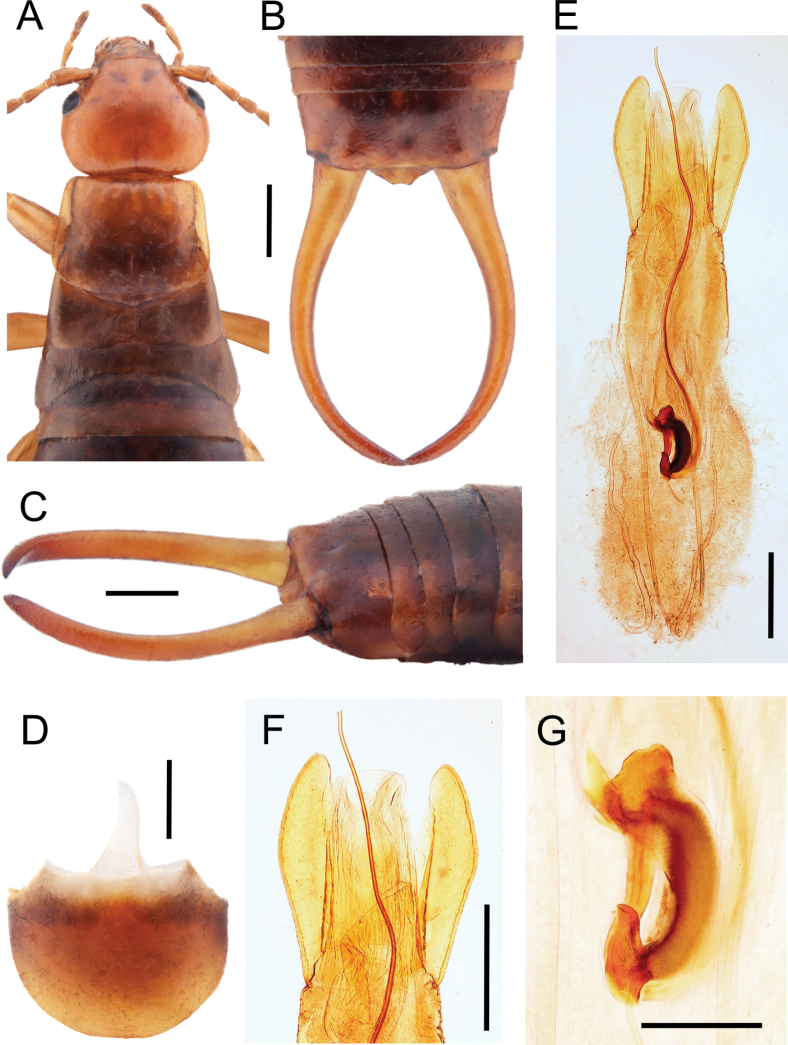 Figure 1
Figure 1 Figure 2
Figure 2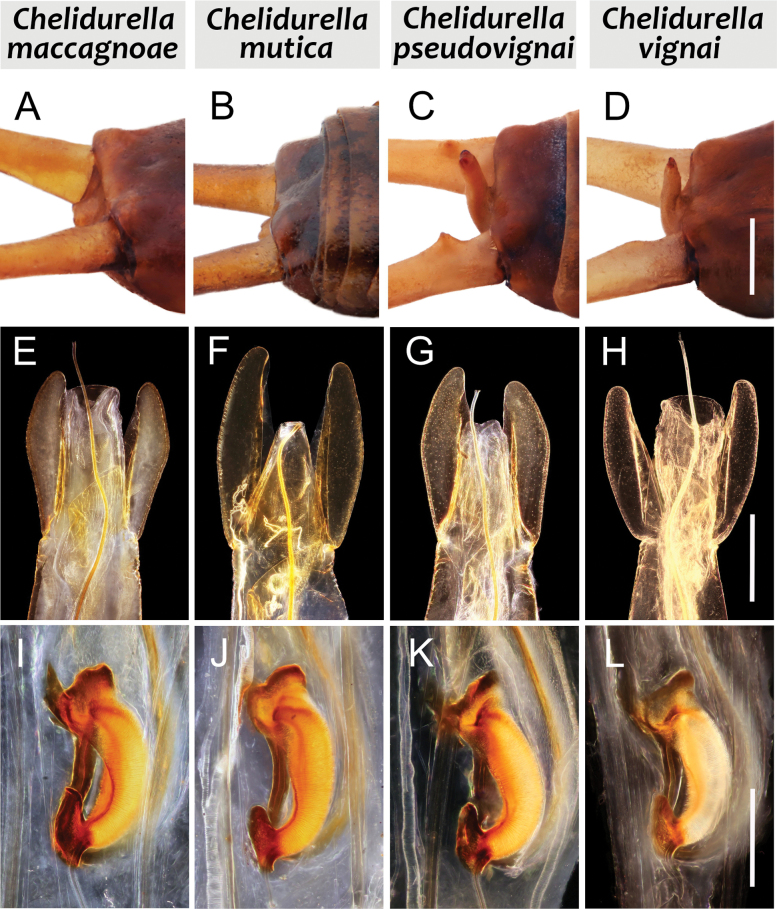 Figure 3
Figure 3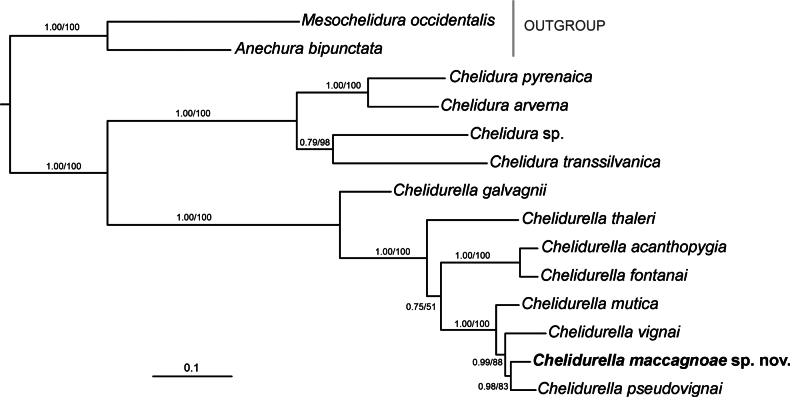 Figure 4
Figure 4| Gene | Primer name | Primer sequence 5’ | PCR protocol | Primer reference |
|---|---|---|---|---|
|
| jgLCO1490 | TITCIACIAAYCAYAARGAYATTGG | 2 min 30 s at 94 °C, [30 s at 90 °C, 1 min at 51 °C, 1 min at 72 °C] 40×, 10 min 30 s at 72 °C |
|
| jgHCO2198 | TAAACTTCAGGGTGACCAAAAAATCA | |||
|
| SR-N-14594 | AAACTAGGATTAGATACCC | 3 min at 95 °C, [30 s at 95 °C, 1 min at 52 °C, 1 min at 72 °C] 10×, [30 s at 95 °C, 1 min at 52 °C, 1 min at 72 °C] 25×, 7 min at 72 °C |
|
| SR-J-14199 | TACTATGTTACGACTTAT | |||
|
| CAS5p8sFc | TGAACATCGACATTTYGAACGCACAT | 1 min at 94 °C, [15 s at 94 °C, 15 s at 53 °C, 15 s at 72 °C] 35×, 6 min at 72 °C |
|
| CAS28sB1d | TTCTTTTCCTCCSCTTAYTRATATGCTTAA |
| Analysed taxa | Locality and year of collection | Isolate / Voucher No. (deposition) |
| 12S |
|
|---|---|---|---|---|---|
|
| |||||
|
| |||||
| France: Chalmazel, 2016 | C_pyr (OU) |
|
|
| |
| Spain: Girona, 2011 | 230 (OU) |
|
|
| |
| Romania, Chișcău, 2022 | 392 (OU) |
|
|
| |
| Italy: Lombardy (Sondrio), 2019 | 251 (OU) |
|
|
| |
|
| |||||
| Czech Republic: Ostrava, 2014 | 24CZ (OU) |
|
|
| |
| Italy: Genoa, 2015 | 091T (OU) |
|
|
| |
| Austria: Karnten, 2015 | 01AU (OU) |
|
| N/A | |
| Italy: Passo di Crocedomini, 2024 | 481 ( |
|
|
| |
| Italy: Veneto, Monte Baldo, 2023 | 444 (OU) |
|
|
| |
| Austria: Carinthia, 2015 | 04AU (OU) |
|
|
| |
| Italy: Bolzano, 2015 | 07IT (OU) |
|
|
| |
| Italy: Trento, 2015 | 05IT (OU) |
|
|
| |
|
| |||||
| Mongolia: Ikh-tamir, 2016 | A_bip (OU) |
|
| N/A | |
| Portugal: Monchique, 2017 | M_occ (OU) |
|
| N/A | |
| 1 | Pygidium small, rounded. Viewed from the back, it does not reach the top line of the 10th tergite (Fig. |
|
| – | Pygidium triangular, pointed at the end or truncated, and when viewed from the back, pygidium reaches or exceeds the upper line of the 10th tergite (Fig. |
|
| 2 | Penis lobe nearly as long as parameres, robust, only slightly tapering towards the top (Fig. | |
| – | Penis lobe narrower, conspicuously trapezoidal, reaching maximally to 2/3 of parameres (Fig. |
|
| 3 | Pygidium long, elongated; viewed from the back, pygidium extends over the upper edge of the 10th tergite ≤ 0.3× the body thickness measured from pygidium base to the upper line of the 10th tergite |
|
| – | Pygidium short, trigonal; viewed from the back, pygidium either does not reach the upper edge of the 10th tergite, or it extends only to 0.3× the body thickness measured from pygidium base to the upper line of the 10th tergite |
|
| 4 | Pygidium very long; viewed from the back, pygidium extends over the upper edge of the 10th tergite ≤ 2× the body height measured from pygidium base to this line. Distal 2/3 of pygidium narrow with parallel margins; pygidium is not strongly S-shaped viewed from the side |
|
| – | Pygidium long, viewed from the back, pygidium extends over the upper edge of the 10th tergite 0.3–0.8× the body height measured from pygidium base to this line. Distal 2/3 of pygidium narrow with convergent margins; pygidium is typically S-shaped viewed from the side |
|
| 5 | Parameres broadly rounded externally, not angulated in distal third. Basal vesicle as in Fig. |
|
| – | Parameres slightly but clearly angled externally in distal 1/3. Basal vesicle as in Fig. |
|
| 6 | Antennomeres short and robust. Length/width ratio of antennomere 4 is maximally 1.5 |
|
| – | Antennomeres long and thin. Length/width ratio of the antennomere 4 is ≤ 1.6 |
|
| 7 | Pygidium with shape of an isosceles triangle; viewed from the back, pygidium barely reaches the upper edge of the 10th tergite, or it extends over the edge maximally 0.25× the body thickness measured from the pygidium base to the upper edge of tergite 10 |
|
| – | Basal part of pygidium triangular; distal part with the narrow projection with nearly parallel sides. Apex of pygidium truncated with two protuberances |
|
| 8 | Subgenital plate with trigonally tapering distal 1/3 |
|
| – | Subgenital plate nearly parallel-sided on proximal 2/3, with rounded sides on distal 1/3 |
|
| 9 | Penis lobe relatively long; paramere ≤ 2× longer than maximal width of pronotum; distal width of penis lobe ≤ 2× broader than maximal width of paramere |
|
| – | Penis lobe relatively short; paramere maximally 1.6× longer than maximal width of pronotum; distal width of penis lobe approximately as broad as maximal width of paramere |
|
| Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
|---|---|---|---|---|---|---|---|---|---|
|
|
| 0.031 | 0.169 | 0.119 | 0.128 | 0.122 | 0.128 | 0.146 | |
|
|
| 0.051 | 0.161 | 0.113 | 0.122 | 0.114 | 0.131 | 0.137 | |
|
|
| 0.203 | 0.215 | 0.122 | 0.128 | 0.126 | 0.136 | 0.155 | |
|
| 0.181 | 0.189 | 0.183 | 0.028 | 0.028 | 0.115 | 0.071 | ||
|
|
| 0.190 | 0.193 | 0.176 | 0.086 | 0.039 | 0.127 | 0.082 | |
|
|
| 0.186 | 0.200 | 0.167 | 0.067 | 0.086 | 0.122 | 0.063 | |
|
|
| 0.174 | 0.165 | 0.183 | 0.180 | 0.164 | 0.183 | 0.139 | |
|
|
| 0.178 | 0.180 | 0.182 | 0.074 | 0.080 | 0.071 | 0.174 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFossil Insects in Amber · Collembola Taxonomy and Ecology Studies · Scarabaeidae Beetle Taxonomy and Biogeography
Introduction
The genus Chelidurella Verhoeff, 1902 represents a complex of nine morphologically similar earwig species distributed across Europe. Majority of these flightless forficulids are restricted to high-altitude habitats, with most species showing narrow endemic distributions in the Alps (Galvagni 1997; Kirstová et al. 2021).
The taxonomic complexity of Chelidurella has long been recognised, with individual species traditionally delimited by subtle combinations of variable characters on the pygidium and forceps that often overlap between taxa (Capra 1982; Vigna Taglianti 1993; Galvagni 1994, 1995, 1996, 1997). This morphological similarity has led to considerable confusion in species identification and likely to the underestimation of true diversity within the genus. Recent molecular phylogenetic analyses have confirmed the existence of cryptic diversity, as demonstrated by the discovery of C. pseudovignai Kočárek & Kirstová, 2021, which is morphologically nearly identical to C. vignai Galvagni, 1995, but genetically well-differentiated and geographically separated (Kirstová et al. 2021).
The Alpine region has been recognised as a major centre of speciation and endemism for numerous arthropod groups, with Quaternary climatic oscillations promoting allopatric divergence in montane refugia (Schmitt 2009; Pauls et al. 2013). The restricted distributions and low dispersal ability of Chelidurella species make them particularly susceptible to such geographic isolation, potentially leading to cryptic speciation in isolated mountain massifs.
Chelidurella mutica (Krauss, 1886) is one of the most distinctive species in the genus, characterised by its small, broadly truncated pygidium (Kirstová et al. 2021). The species was described from the central-southern Alps of Italy (Monte Baldo), with some additional records from Austria and Switzerland (Galvagni 1997; Haas 2007). However, the true diversity within what has been considered “C. mutica” has never been thoroughly investigated using molecular tools.
During recent collecting efforts in the Italian Alps, we discovered a population of Chelidurella specimens from Passo Crocedomini that is morphologically very close to C. mutica but show distinct molecular divergence. Preliminary molecular analyses reveal that this population represents a cryptic species that, despite its morphological similarity to C. mutica, is phylogenetically closer to the species pair C. pseudovignai /C. vignai. This finding adds further evidence to the growing recognition of hidden diversity within Chelidurella and highlights the importance of integrative taxonomic approaches in revealing the true extent of Alpine biodiversity.
Here we describe this new cryptic species based on molecular phylogenetic analyses combined with detailed morphological examination, discuss its biogeographic implications for understanding Quaternary diversification in the Alps, and provide updated identification tools for the genus.
Materials and methods
Taxon sampling and processing
Specimens of describing species has been collected in Italy: Passo di Crocedomini, Brescia (BS), and, and preserved in 96% ethanol until tissue extraction for DNA isolation. For DNA isolation, we used leg and thoracic muscles. After that, the studied specimens were dissected and mounted on a label, with male genital armatures placed in small vial with glycerine and pinned to the card with the specimen. For the morphological comparisons, the specimens from the following localities were used: Chelidurella mutica: Italy: Veneto, Monte Baldo, 45°46'27.383"N, 10°52'5.160"E, 6.ix.2023, P. Kočárek leg.; Chelidurella pseudovignai: Italy: Carnia env., 550 m a.s.l., 46°23'06"N, 13°09'53"E, 20.ix.2015, sweeping of bushes, M. Kirstová leg.; Chelidurella vignai: Italy: Trento, 46°07'11"N, 11°15'40"E, loc. 7, 1031 m a.s.l., 12.ix.2015, M. Kirstová leg.
The specimens used for visualisation were photographed with a Leica Z16 APO macroscope (Leica Microsystems, Wetzlar, Germany) equipped with a CANON 6D Mark II camera. Micrographs of 10 to 20 focal layers of the same specimen were combined with Helicon Focus software and finally processed with Adobe Photoshop CS6 Extended (Version 13). The terminology used for morphological characteristics follows that of Steinmann (1986); the terminology used to describe the terminalia and genitalia follows that of Kamimura (2014). The nomenclature used follows that of Hopkins et al. (2025).
The material treated in this study is deposited in the following collections:
FMCR the Fondazione Museo Civico di Rovereto, Italy
UO the University of Ostrava, Department of Biology and Ecology, Czech Republic
DNA extraction, amplification, and sequencing
Total genomic DNA was extracted from five Forficulidae taxa via the DNeasy Blood & Tissue Kit (QIAGEN, Inc.) following the manufacturer’s protocol. Partial sequences of three genetic markers (one nuclear and two mitochondrial) were amplified and sequenced. These include internal transcribed spacer 2 (ITS2), cytochrome c oxidase subunit I (COI), and 12S rRNA. Polymerase chain reactions (PCRs) were performed in 20-µl volumes containing 1 µl of DNA template, each primer at 0.4 µM, distilled water, and 1× PCRBIO HS Taq Mix Red (PCR Biosystems, London, UK). Primer pairs and PCR conditions used for amplification are listed in Table 1. The PCR products were visualised via 1% agarose gel electrophoresis with MIDORI Green Advance (NIPPON Genetics EUROPE) to confirm amplification. For purification of the amplified DNA was used a Gel/PCR DNA Fragments Extraction Kit (GENAID, Taiwan). Sanger sequencing reactions were performed at Eurofins Genomics (Konstanz, Germany). The chromatograms were visually checked via ChromasPro v2.1.10.1 software (Technelysium, Brisbane, Australia). Details of analysed taxa including isolation numbers, localities and GenBank accession numbers are indicated in Table 2.
Phylogenetic analysis
The sequences were aligned using MAFFT with default parameters implemented via the EMBL-EBI sequence analysis tools (Madeira et al. 2022). Pairwise genetic distances using the Kimura 2-parameter model were calculated for the COI and 12S alignments in MEGA v12 (Kumar et al. 2024).
For phylogenetic reconstruction, we used partial sequences of three genetic markers: COI (657 bp), 12S rRNA (~617 bp), and ITS2 (~457 bp). The protein-coding gene (COI) was translated into amino acids to check for potential stop codons within the open reading frames. The multigene dataset of 1,731 bp was concatenated using Concatenator v0.2.1 (Vences et al. 2022). The best-fitting partitioning schemes and molecular evolution models were selected under the corrected Akaike information criterion via PartitionFinder v2.1.1 (Lanfear et al. 2017). Bayesian inference (BI) and maximum likelihood (ML) analyses were performed to infer phylogenetic relationships, and both analyses were run on the CIPRES Science Gateway v3.3 (Miller et al. 2010). Bayesian analysis was conducted with MrBayes v3.2.7a in XSEDE (Ronquist et al. 2012) via the Markov chain Monte Carlo (MCMC) method. Two independent MCMC runs of four chains were run for 5 million generations, and 25% of the trees were discarded as burn-in. The convergence of BI analysis was confirmed in Tracer v1.7.2 (Rambaut et al. 2018). The maximum likelihood analysis based on the GTR+G+I nucleotide model was conducted via RAxML-HPC BlackBox v8.2.12 (Stamatakis 2014). The obtained trees were rooted by outgroup taxa (Anechura bipunctata and Mesochelidura occidentalis) and were displayed in iTOL (interactive Tree of Life) v6.8.1 (Letunic and Bork 2021).
Results
Taxonomy
Chelidurella
Taxon classificationAnimaliaDermapteraForficulidae
Genus
Verhoeff, 1902
70333C86-3D42-5A69-B857-A0A4720AF51D
Chelidurella Verhoeff, 1902: 187. Type species: Forficula acanthopygia Gené, 1832 by subsequent designation in Kirby 1904: 43. Type locality: Alpes-Maritimes.
Included species.
C. acanthopygia (Gené, 1832); C. caprai Vigna Taglianti, 1993; C. fontanai Galvagni, 1996; C. galvagnii Kirstová & Kočárek, 2021; C. mutica (Krauss, 1886); C. poggii Capra, 1982; C. pseudovignai Kočárek & Kirstová, 2021; C. thaleri Harz, 1980; C. vignai Galvagni, 1995; C. maccagnoae Kočárek & Fontana, sp. nov.
Distribution.
Europe.
Chelidurella
maccagnoae
Taxon classificationAnimaliaDermapteraForficulidae
Kočárek & Fontana sp. nov.
4A6995FB-A40F-5EEE-9188-958EEEA03693
https://zoobank.org/1DAB9F3B-5ED2-47E0-B989-5F3B6B2F146A
Material.
Holotype. Italy • 1 ♂; Passo di Crocedomini (Breno) BS, 1900 m; 45°54'19"N, 10°24'33"E; 19 Aug. 2024; Paolo Fontana leg.; GenBank: COI: PX585052, 12S: PX640177, ITS2: PX640172 (the Fontana collection in FMCR). Paratypes. Italy • 1 ♀; Passo di Crocedomini (Breno) BS, 1900 m; 45°54'19"N, 10°24'33"E; 19 Aug. 2024; Paolo Fontana leg. (the Fontana collection in FMCR); Italy • 2 ♀; Passo di Crocedomini (Breno) BS, 1900 m; 45°54'28.7"N, 10°24'33.6"E, 5 Sep. 2023, Paolo and Carlotta Fontana leg.; ID: 454, 455 (UO).
Diagnosis.
Chelidurella maccagnoae Kočárek & Fontana, sp. nov. and C. mutica share shortened pygidium without upcurved projection and this character clearly separates these two species from the remaining described species of Chelidurella. Chelidurella maccagnoae sp. nov. and C. mutica are externally indistinguishable, but differ in the morphology of male genitalia. The penis lobe of C. maccagnoae sp. nov. is long, reaches almost the tip of the parameres, is nearly parallel sided and truncated distally; the penis lobe of C. mutica is shorter, reaches up to two-thirds of the length of parameres, sloping to the apex that is truncate. The parameres of C. maccagnoae sp. nov. are more angular along the outer margin than those of C. mutica. Both species differ also in the morphology of basal vesicle, in the shape of proximal expansion. Proximal and distal corners of the proximal expansion of basal vesicle are pointed in C. maccagnoae sp. nov., but broadly rounded in C. mutica.
Description.
Body yellowish-brown (Figs 1, 2); head orange-yellow, legs and forceps ochre. Cuticle punctured, shiny. Tegmina of Chelidurella type, rudimentary, rugose, with a short section overlapping medially; wings entirely absent. Pygidium ochre, short, without upward projection. Measurements of holotype male: Total length without forceps 11.8 mm, length of forceps 4.6 mm. Maximal head width 2.18 mm, maximal head length 2.1 mm, maximal pronotum length/width 1.7/2.2 mm, maximal tegmina length 2.49 mm, length of 1^st^ antennomere (scapus) 0.7 mm, length of 2^nd^ antennomere (pedicel) 0.1 mm, length of 3^rd^ antennomere 0.4 mm. Total length of male genitalia 3.2 mm (with exclusion of exceeding virga), basal vesicle length 0.4 mm. Measurements of paratype female: Total length without forceps 11.3 mm, length of forceps 2.3 mm in females.
Chelidurella maccagnoae Kočárek & Fontana, sp. nov., habitus of holotype male. A. Dorsal view to head and thoracal segments; B. Ultimate tergite and cerci from dorsal view; C. Ultimate tergite and cerci from lateral view; D. Penultimate sternite, ventral view; E. Dorsal view on the genital of holotype; F. Detail of the tip of genital; G. Basal vesicle, ventral view. Scale bars: 1.0 mm (A–D); 0.5 mm (E, F); 0.2 mm (G).
Chelidurella maccagnoae Kočárek & Fontana, sp. nov., live habitus of holotype male (A) and paratype female (B). Passo di Crocedomini (Breno), 1900 m. Photograph: PF 19 Aug. 2024.
Holotype male. Head (Fig. 1A) transverse; frons gibbose convex, postfrontal and coronal sutures distinct, posterior margin concave in middle. Eyes small, as long as 3^rd^ antennomere. Antennae with 13 antennomeres; 1^st^ antennomere narrowed in basal third, widened distally, as long as 3^rd^ and 4^th^ antennomeres combined; 2^nd^ antennomere short, quadratic; 3^rd^ antennomere longer than 4^th^, both subconical; antennomeres 5–13 cylindrical, narrowed at extreme base. All antennomeres shortly pubescent.
Pronotum (Fig. 1A) transverse, as wide as head, slightly expanded distally and broadly rounded posteriorly. Disc of pronotum flat, median sulcus fine but distinct. Tegmina rugose, rudimentary, with a short section overlapping medially, lateral and internal edges broadly rounded. Mesonotum dorsally visible as distally broadly rounded mesoscutellum; metanotum transverse, wider than long, posteriorly broadly emarginate. Sternal plates typical for genus. Legs unicolourous ochre; femora stout; tibiae clad with thick and fine setae; 1^st^ metatarsomere cylindrical, longer than 2^nd^ and 3^rd^ tarsomeres combined, pulvilli not developed. Claws simple, symmetrical; arolium absent.
Abdomen (Fig. 1B, C) somewhat widened in middle, lateral glandular folds present on tergites 3 and 4, tergites 4–6 rugose-striate at sides. Ultimate tergite transverse, slightly sloping posteriorly, more than twice as wide as long, median part somewhat depressed near posterior margin, disc with a pair of rounded tubercles near the forceps bases. Ninth sternite (penultimate) broadly rounded posteriorly (Fig. 1D). Pygidium broad and short, without upward projection, lateral sides sloping to central truncated part with small tubercules at corners. Forceps (Fig. 1B, C) arcuate, without any tooth, branches cylindrical and regularly tapering to the apex.
Genitalia (Figs 1E–G, 3E, 3I) with only slightly narrowed penis lobe distally with truncated apex. Parameres thin, 4× longer than wide, broadened in middle and narrowed apically, external margin convex, internal margin gently emarginated. Basal vesicle thin, 2× shorter than length of paramere, reniform. Proximal expansion more robust than distal expansion, distal expansion transverse, with acute corners. Virga sinuated, well sclerotised, elongated ~ 3× longer than paramere.
Comparison of male pygidiums, penis lobes, parameres, and basal vesicles. A, E, I. Chelidurella maccagnoae Kočárek & Fontana, sp. nov.; B, F, J. C. mutica (Krauss, 1886); C, G, K. C. pseudovignai Kočárek & Kirstová, 2021; D, H, L. C. vignai Galvagni, 1995. Scale bars: 1.0 mm (A–D); 0.5 mm (E–H); 0.2 mm (I–L).
Female. Agrees with male in all characters except female forceps have simple and straight with contiguous branches (Fig. 2B).
Molecular identification.
We obtained partial COI sequence (657 bp) from the holotype male specimen of Chelidurella maccagnoae Kočárek & Fontana, sp. nov. as DNA barcode for molecular identification of the species. The barcode is deposited in GenBank under accession number PX585052 (DNA isolate number: 481).
Etymology.
The new species is dedicated to the Italian zoologist Teresita Maccagno Paulucci (1900–1999), who, with her seminal works, made a decisive contribution to the study of Dermaptera by pioneering the comparative examination of male genitalia in the classification of these insects (Maccagno 1926, 1933). Her life spanned a century, but, like many women in science, she never received the recognition she deserved.
Biology.
The few specimens collected (during intense searches) were identified under stones of various sizes at the edges of pastures near the Cricedomini Pass. Specimens were collected (in very small numbers) between the end of August (2024) and the beginning of September (2023). The species was also the subject of research in mid-July 2025, but without success. During the same field trip conducted at the end of August 2024, some juvenile stages of Dermaptera were identified in addition to the adult male and female collected, apparently attributable to the new species. This very limited data does not allow us to outline the phenology of this species, which appears to be present in a very small population.
Distribution.
Known only from type locality in Adamello-Presanella Alps in Italy, the Crocedòmini Pass. This is an Alpine pass in the Brescia Prealps, located in the province of Brescia at 1,892 m a.s.l., south of the Adamello Park in Lombardy. From an orographic perspective, the pass separates the Southern Rhaetian Alps from the Brescian and Garda Prealps. It is located just north of the point where the Three Brescian Valleys meet: Val Trompia, Val Camonica, and Val Sabbia.
Key to males of Chelidurella (modified from Kirstová et al. 2021)
**: **
Phylogenetic analyses
The analyses contain eight species of the genus Chelidurella, including the new species described in this study, and four species of the genus Chelidura Latreille, 1825. The dataset of three molecular markers (COI, 12S, and ITS2) comprised 1,731 bp. The trees based separately on the BI and ML methods produced identical topologies (Fig. 4). The genus Chelidurella forms a monophyletic clade with 100% support in both analyses in a sister position to the genus Chelidura.
Phylogenetic tree resolved by Bayesian inference based on the combined dataset for three molecular markers (COI, 12S rRNA, and ITS2). Bayesian posterior probabilities (PP) and RAxML bootstrap support (BS) values are given at each node in the following order: PP/BS.
Chelidurella mutica, C. vignai, C. pseudovignai, and C. maccagnoae sp. nov. form a monophyletic clade that is sister to the clade comprising C. acanthopygia and C. fontanai. The new species, C. maccagnoae, is the sister species to C. pseudovignai. The pairwise genetic distances (Kimura 2-parameter model) between these two species were 6.7% for COI and 2.8% for 12S. The pairwise genetic distances for all analysed Chelidurella species (COI and 12S) are presented in Table 3.
Discussion
The cryptic diversity of the genus Chelidurella has long been recognised, but small morphological differences and high intraspecific variability have complicated species identification (Kirstová et al. 2021). While Italian entomologists Capra, Vigna Taglianti, and Galvagni made significant taxonomic contributions (Capra 1982; Vigna Taglianti 1993; Galvagni 1994, 1995, 1996, 1997), recent molecular revisions not only confirmed most described species but revealed additional cryptic taxa like C. pseudovignai (Kirstová et al. 2021). The discovery of C. maccagnoae sp. nov. through molecular verification of an isolated C. mutica population exemplifies this hidden diversity. Surprisingly, despite sharing an extremely shortened pygidium with C. mutica, C. maccagnoae sp. nov. is phylogenetically closer to the C. vignai/C. pseudovignai complex, indicating convergent evolution (homoplasy) rather than shared ancestry. This pygidial shortening likely represents adaptation to high-altitude conditions, potentially following Allen’s rule, though in ectothermic insects such morphological variation may reflect allometric growth under different climatic conditions rather than thermoregulation (Bidau and Martí 2008; Shelomi and Zeuss 2017).
Quaternary climatic oscillations have profoundly shaped Alpine invertebrate communities, particularly affecting flightless and brachypterous insects (Ikeda et al. 2012; Hunt et al. 2013). While classical European phylogeography emphasised three Mediterranean refugia (Iberian, Italian, Balkan) with northward postglacial expansion (Taberlet et al. 1998; Hewitt 2000), molecular studies reveal more complex patterns including peripheral refugia, nunatak refugia within mountain ranges (ice-free rocky peaks), and extra-Mediterranean refugia (Schönswetter et al. 2005; Schönswetter and Tribsch 2019; Stewart and Lister 2019). Flightless arthropods demonstrate pronounced Quaternary diversification effects due to limited dispersal. Studies of ground beetles reveal strong genetic differentiation with divergence times extending 1.6–2.5 million years (Raupach et al. 2013), while flightless species retain twice the speciation rates of flying relatives (Ikeda et al. 2012). The endemic bush-cricket Anonconotus italoaustriacus exemplifies Alpine flightless arthropod dynamics, with genomic evidence supporting survival in multiple peripheral refugia and subsequent southeastern recolonisation (Schönenberger et al. 2022).
The earwig genera Chelidura and Chelidurella exemplify contrasting evolutionary responses to Quaternary diversification. Both are completely flightless with rudimentary tegmina, severely limiting dispersal (Fontana et al. 2021; Kirstová et al. 2021). However, while Chelidurella species inhabit diverse environments from lowlands to upper forest boundaries, Chelidura species are exclusively high-altitude specialists (1400–2300 m), persisting above the tree line in harsh alpine conditions. Recent genetic analysis of Chelidura aptera species complex revealed substantial cryptic diversity among Alpine populations indicating long-term isolation in multiple refugia and significant taxonomic underestimation (Fontana et al. 2021). Their ecological requirements for alpine grasslands with rocky outcrops align perfectly with nunatak refugia scenarios, where ice-free peaks provided suitable refugial environments during Pleistocene glaciations. This ecological contrast drives different diversification patterns. Chelidurella shows four strongly supported molecular clades with probable Pleistocene diversification, yet even this ecologically flexible genus exhibits cryptic species pairs like C. vignai and C. pseudovignai in alpine environments (Kirstová et al. 2021). Notably, only high-altitude specialists C. mutica and the newly described species approach Chelidura’s ecological niche at ~2000 m elevation.
Both Chelidura and Chelidurella diversity patterns align with broader phylogeographic paradigms. Their restriction to high-altitude habitats, flightless condition, and specialised ecological requirements have promoted allopatric speciation within isolated mountain refugia. Molecular phylogenies indicate relatively recent, likely Pleistocene diversification, consistent with other Alpine endemic lineages showing Quaternary rather than Tertiary origins (Kadereit et al. 2004; Schmitt 2007).
These phylogeographic patterns have significant conservation implications under ongoing climate change. Historical Alpine refugia are becoming increasingly important as potential climate refugia for cold-adapted species, yet current protected area networks may inadequately cover these future refugia (Wilkes et al. 2022, 2023). High-altitude specialists like Chelidura species face particular vulnerability, as their narrow ecological tolerances and restricted distributions limit upward range shift capacity in response to warming temperatures.
Supplementary Material
XML Treatment for Chelidurella
XML Treatment for Chelidurella maccagnoae
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bidau CJ, Martí DA (2008) A test of Allen’s rule in ectotherms: the case of two south American Melanopline Grasshoppers (Orthoptera: Acrididae) with partially overlapping geographic ranges. Neotropical Entomology 37(4): 370–380. 10.1590/S 1519-566X 200800040000418813738 · doi ↗ · pubmed ↗
- 2Capra F (1982) Sulla Chelidurella acanthopygia (Gené) e su una nuova specie dell’Alto Adige (Insecta: Dermaptera). Memorie della Società Entomologica Italiana 60: 111–118.
- 3Fontana P, Pedrazzoli F, Malagnini V, Ruzzier E, Marangoni F, Kočárek P (2021) Toward a revision of the genus Chelidura Latreille, 1825: designation of the Neotype for Chelidura aptera (Megerle in Charpentier, 1825) (Dermaptera: Forficulidae). Memorie della Società Entomologica Italiana 97: 279–302. 10.4081/memoriesei.2020.279 · doi ↗
- 4Galvagni A (1994) Chelidurella guentheri specie nuova dell’Europa centrale e della Norvegia Sud-orientale (Insecta Dermaptera Forficulidae). Atti della Accademia Roveretana degli Agiati 3(B): 347–370.
- 5Galvagni A (1995) Chelidurella vignai specie nuova delle Alpi Sud-orientali (Insecta Dermaptera Forficulidae). Annales Museo Civico di Rovereto 10: 379–397.
- 6Galvagni A (1996) Identificazione e variabilitá della Chelidurella acanthopygia (Gené, 1832) con istituzione della Chelidurella fontanai sp. n. (Insecta Dermaptera). Atti della Accademia Roveretana degli Agiati 6(B): 5–45.
- 7Galvagni A (1997) Contributo alla conoscenza del genere Chelidurella Verhoeff, 1902, in Italia e territori limitrofi (Insecta Dermaptera). Atti della Accademia Roveretana degli Agiati 7: 5–61.
- 8Geller J, Meyer C, Parker M, Hawk H (2013) Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Molecular Ecology Resources 13(5): 851–861. 10.1111/1755-0998.1213823848937 · doi ↗ · pubmed ↗