A commercial bacteriophage cocktail failed to decolonize Zophobas morio larvae and promoted overgrowth of an OXA-48-producing Salmonella enterica
Claudia Aldeia, Edgar I. Campos-Madueno, Andrea Endimiani

TL;DR
A commercial phage treatment failed to remove a harmful Salmonella strain from mealworms and instead caused it to grow more, showing the need for caution with phage therapies.
Contribution
Demonstrates that a broad-spectrum bacteriophage cocktail can promote overgrowth of a carbapenem-resistant Salmonella strain in an in vivo model.
Findings
INTESTIbc treatment led to significant regrowth of OXA-48-producing Salmonella enterica in Zophobas morio larvae.
Phage administration reduced potential bacterial competitors of Salmonella, possibly contributing to its overgrowth.
Zophobas morio larvae are a viable in vivo model for studying intestinal colonization by Salmonella enterica.
Abstract
Effective decolonization strategies for intestinal carriers of carbapenem-resistant Enterobacterales are essential to prevent severe life-threatening infections. In this work, we established gut colonization in Zophobas morio larvae (ZmL) using an OXA-48-producing Salmonella enterica ST198 strain (Sk-1) and assessed the commercial INTESTI bacteriophage cocktail (INTESTIbc) for decolonization. ZmL were fed with food contaminated with Sk-1 (INTESTIbc-susceptible) for 3 days and then maintained on a non-contaminated diet until day 14 (T14). At T3, ZmL were grouped in untreated, dPBS- or INTESTIbc-treated (oral force-feeding on T3 and T5). At specified intervals, ZmL were sampled for quantification and characterization of Sk-1 (antibiotic/INTESTIbc susceptibility and whole-genome sequencing). ZmL microbiota was also investigated by 16S rRNA amplicon sequencing. ZmL were rapidly colonized…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
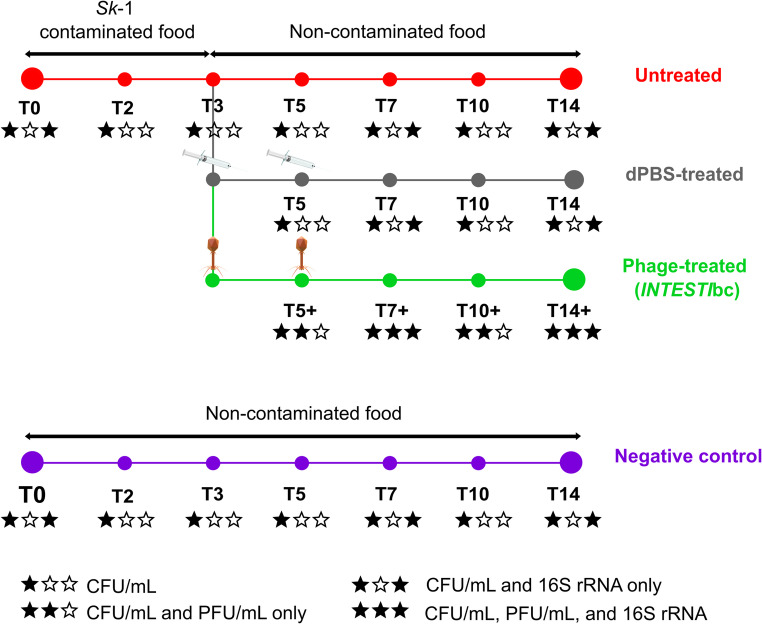 Figure 1
Figure 1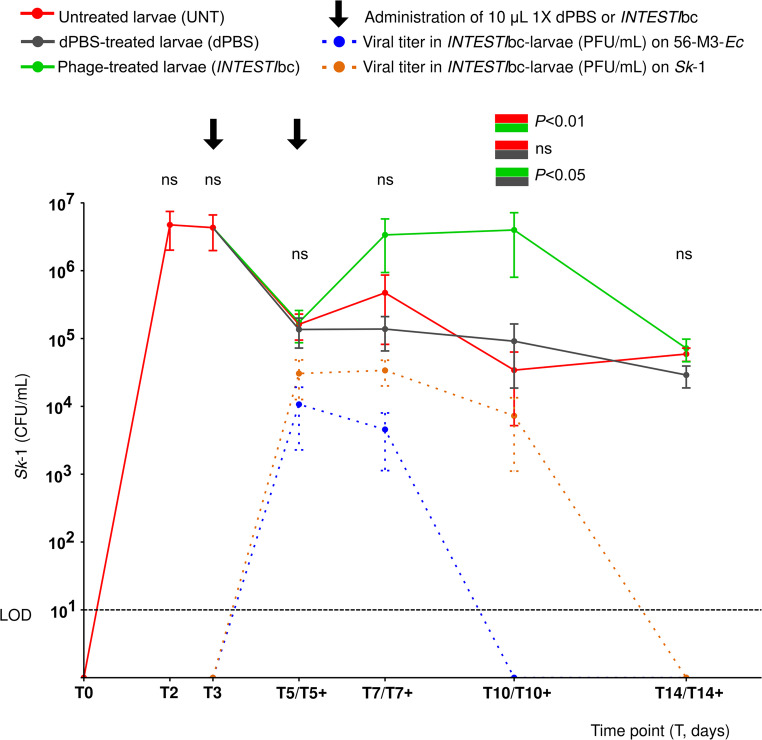 Figure 2
Figure 2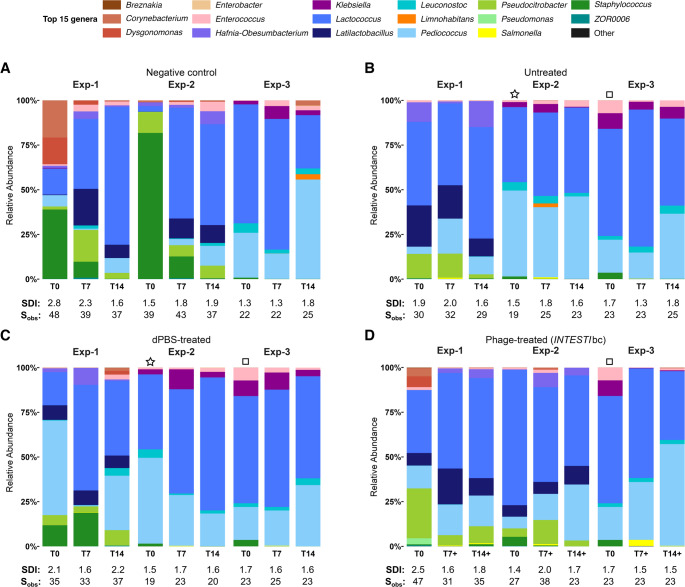 Figure 3
Figure 3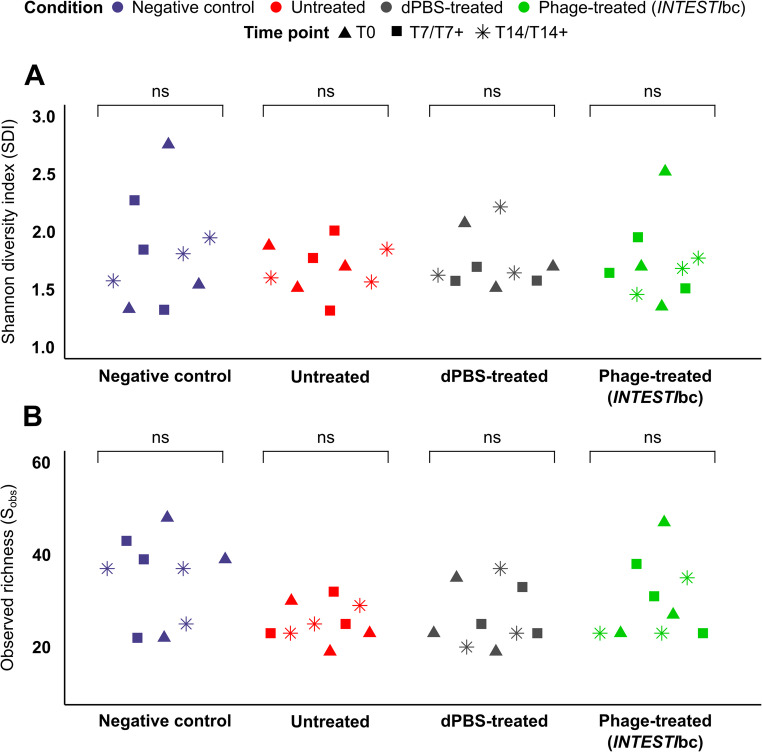 Figure 4
Figure 4- —University of Bern
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Aquaculture disease management and microbiota · Salmonella and Campylobacter epidemiology
Introduction
Salmonella enterica isolates are important pathogens primarily responsible for gastroenteritis, but may also cause invasive infections with high mortality rates. In this context, third-generation cephalosporins (3GCs) and fluoroquinolones are usually used for treatment [1–3]. However, the high prevalence of 3GC-resistant S. enterica strains that produce plasmid-mediated extended-spectrum β-lactamases (ESBLs) or AmpC represent a serious concern [4–6]. Even more alarming is the recent emergence of global hyperepidemic clones, such as sequence type (ST) 198 serovar Kentucky [7–11], which are capable of producing carbapenemase enzymes (e.g., OXA-48-like, KPC- and NDM-types) [12–15]. Overall, these 3GC-/carbapenem-resistant S. enterica strains can lead to difficult-to-treat infections, but they may also contaminate the food chain and colonize the intestinal tract of both humans and animals [9, 16, 17], contributing to their further spread and expansion among different settings [18, 19].
As for other Enterobacterales, finding strategies for decolonizing gut carriers of multidrug-resistant (MDR) S. enterica is therefore a public-health priority [20]. In the past, several approaches (e.g., bacteriophages, fecal microbiota transplantation, probiotics) have been explored and envisioned. Nevertheless, such suggestions were based on limited data that were mostly obtained with difficult to implement animal models (e.g., mouse, chicken) [21–24]. For instance, the gold-standard mouse model presents severe ethical (e.g., pain experienced by animals) and logistical limitations (e.g., requirement for skilled personnel, designated facilities), which can all together generate expensive and time-consuming investigations. In contrast, the use of an invertebrate model could provide a cheap, suitable and highly scalable alternative in agreement with the Replacement, Reduction and Refinement (3Rs) framework [25]. Such a model may also be useful for rapid in vivo testing of novel treatment and decolonization strategies, generating detailed preclinical information prior to human clinical trials.
Zophobas morio is an insect that belongs to the family of Tenebrionidae beetles. Its late-instar larvae (700 ± 50 mg and 45 ± 5 mm in length) possess a hard exoskeleton and may remain in this stage without pupating for > 6 months. Moreover, Z. morio larvae (ZmL) can be easily reared on a wide range of diets [26]. This last aspect determines the unusual richness and diversity of bacterial species that can populate their gut, resembling that of humans [27, 28]. Overall, due to these characteristics, ZmL could provide an advantageous model compared to other invertebrates (e.g., Galleria mellonella, GmL) for simulating the gut colonization [29].
Recently, we designed a new intestinal colonization model using ZmL [27]. We demonstrated that larvae could be persistently colonized with MDR Escherichia coli isolates belonging to hyperepidemic clones (e.g., ST131 producing the CTX-M-15 ESBL, ST410 producing the OXA-181 carbapenemase) after being fed contaminated food. We also showed that the use of the commercial INTESTI bacteriophage cocktail (INTESTIbc; Eliava BioPreparations) was able to fully decolonize ZmL when the MDR E. coli was INTESTIbc-susceptible in vitro [27].
In this work, ZmL were successfully colonized by a hyperepidemic carbapenemase-producing S. enterica strain (Sk−1) and then treated with the INTESTIbc. As a result, though Sk−1 was in vitro INTESTIbc-susceptible, the in vivo model showed that the bacteriophage treatment was ineffective, leading instead to an unexpected overgrowth of the pathogen. Microbiota analyses also revealed that INTESTIbc treatment resulted in a reduction in bacterial richness, including the relative abundance of putative competitors of Sk−1.
Materials and methods
Features of the colonizing strain (Sk-1)
Sk−1 was isolated in 2012 from a perianal screening culture of a patient. It was previously identified as a S. enterica subsp. enterica serovar Kentucky of ST198 possessing several antimicrobial resistance genes, including the blaOXA−48 carbapenemase- (IncL plasmid) and the blaVEB−8 ESBL- (chromosomally located) encoding genes [12]. Antimicrobial susceptibility testing (AST) performed with the Sensititre^™^ MIC microdilution ESB1F and GNX2F panels (Thermo Fisher Scientific) indicated that Sk−1 was resistant to all cephalosporins, piperacillin-tazobactam, ciprofloxacin, amikacin, tobramycin and ertapenem (Table 1) [12].
Table 1. Phenotypic and molecular characteristics of the original S. enterica subsp. enterica serovar Kentucky strain Sk−1 used for ZmL experimentsAntibioticMIC, µg/mL (interpretation)^a^Ampicillin> 16 (R)Piperacillin-tazobactam> 64 (R)Ticarcillin-clavulanate> 128 (R)Cefazolin> 16 (R)Cephalothin> 16 (NA)Cefpodoxime> 32 (R)Cefoxitin8 (NA)Ceftriaxone128 (R)Cefotaxime> 64 (R)Cefotaxime-clavulanate32 (NA)Ceftazidime> 128 (R)Ceftazidime-clavulanate> 128 (NA)Cefepime> 16 (R)Aztreonam> 16 (R)Imipenem≤ 0.5 (S)Meropenem≤ 1 (S)Ertapenem1 (R)Doripenem0.25 (S)Gentamicin≤ 1 (S)Tobramycin> 8 (R)Amikacin16 (R)Ciprofloxacin> 2 (R)Levofloxacin> 8 (R)Trimethoprim/sulfamethoxazole≤ 0.5 (S)Colistin≤ 0.25 (S)Polymyxin B0.5 (NA)Minocycline8 (NA)Doxycycline8 (NA)Tigecycline0.5 (S)Susceptibility to INTESTI bacteriophage cocktail++++ (S)blaOXA−48 (β-lactams)blaVEB−8 (β-lactams)aac(6)-Ib (aminoglycosides)tet(A) (tetracyclines)sul1 (sulfonamides)Antimicrobial resistance genes (targeted antibiotics)^b^mphA (macrolides)#1:** ~**63.5-Kb - IncL(blaOXA−48) #2:
- ~* 5.7-Kb
-
-
Col156
#3:
- ~* 2.1-Kb
-
-
Col(MP18) Plasmids#4: ~2-Kb - ColpVCSequence type (ST)ST198^a^According to the EUCAST criteria 2025 (v15): R, resistant; S, susceptible; NA, not applicable or not available. MIC values were obtained implementing the Sensititre ESB1F and GNX2F plates (Thermo Fisher Scientific)^b^Sk-1 also had amino acid substitutions in GyrA (Ser83Phe and Asp87Asn) and ParC (Ser80Ile) conferring resistance to fluoroquinolones
In the present study, susceptibility of Sk−1 to the INTESTIbc was tested using the double-layer agar method (DLA) [30]. Sk−1 also underwent whole-genome sequencing (WGS) with both Illumina NovaSeq 6000 (2 × 150 bp) and Nanopore MinION^™^ Mk1B (SQK-RBK004 rapid barcoding kit; FLO-MIN106D R9.4.1 flow cell, 48 h) technologies, as previously described [31, 32]. Briefly, short- and long-read adapter sequences were trimmed using Trimmomatic v0.39 and Porechop v0.2.4, respectively. Hybrid assembly was generated with Unicycler v0.5.0 and polished one round using Polypolish v0.6.0 and PyPolca v0.3.1. The full nucleotide dataset from the Virulence Factor Database (http://www.mgc.ac.cn/VFs/; downloaded on June 17th, 2025) was used to construct a Salmonella-specific database. Virulence factor (VF) genes were investigated by querying the hybrid assembly against the custom database via ABRicate v1.0.1 (https://github.com/tseemann/abricate).
ZmL colonization and experimental groups
Late-instar ZmL (BUGS-International GmbH) originating from two larval stocks (batch 1: August 2023; batch 2: February 2024) purchased from the same Swiss pet store were used for all experiments, as previously described [27]. Briefly, ZmL were reared and kept in polypropylene containers and on a daily diet consisting of fresh pear slices and dry cat food mixed in a substrate of oat flakes (Figure S1). Mortality was recorded as previously defined [33].
To induce gut colonization, ZmL were fed for 3 days [from day 0 (T0) to day 3 (T3)] with food contaminated with S. enterica Sk−1. To do so, colonies grown on a MacConkey II plate were incubated overnight (36 ± 1 °C) in 10 mL Luria-Bertani (LB) broth. This broth was then poured onto a sterile Petri dish containing the food and incubated (1 h, 36 ± 1 °C) before being given to ZmL (Figure S1). After T3, ZmL were randomly assigned to 3 experimental groups (i.e., untreated, INTESTIbc- and dPBS-treated) and transferred to separate cages, where they received non-contaminated food daily until T14. Three replicates were performed per condition, including three additional experiments for the negative controls that received only non-contaminated food from T0 to T14 (Fig. 1). Notably, experiments with ZmL do not require ethical approval (Art. 112 of the Swiss Animal Protection Ordinance; https://www.blv.admin.ch/blv/en/home/tiere/tierschutz.html).
Fig. 1. Overall experimental design for the Zophobas morio larvae (ZmL) assays. T, time point (day). CFU, colony forming unit. PFU, phage forming unit. At T3 and T5, symbols indicate oral administration of either 10 µL dPBS or INTESTI bacteriophage cocktail (INTESTIbc). Stars below each time point correspond to sampling type (see legend)
Treatment with bacteriophages
To attempt decolonizing from Sk−1, a randomly selected group of colonized ZmL received 10 µL of INTESTIbc orally via force-feeding on both T3 and T5, administered with a blunt 26s-gauge needle connected to a 250 µL Hamilton 1700 syringe. Another group of colonized ZmL was also subjected to force-feeding experiments in which on both T3 and T5 they received 10 µL of sterile 1X Dulbecco phosphate-buffered saline (dPBS) used as control (Fig. 1).
Notably, the INTESTIbc [~ 10^5–6^ plaque forming unit (PFU)/mL] targets E. coli, Shigella spp., Salmonella spp. (Paratyphi A, Paratyphi B, Typhimurium, Enteritidis, Cholerasuis, Oranienburg), Proteus vulgaris/mirabilis, Pseudomonas aeruginosa, Staphylococcus aureus, and Enterococcus spp [34]. All experiments were conducted using the same INTESTIbc lot (M2-1301).
Analysis of ZmL
ZmL were processed as previously described [27] (Fig. 1). In brief, for each experiment, 4 ZmL were randomly sampled at 7 time points (Tps): T0 (before administering contaminated food), T2, T3 and T5 (both before the corresponding force-feeding with INTESTIbc or dPBS), T7, T10, and T14. For simplicity, samples taken during experiments with the use of INTESTIbc will be hereafter referred to as T5+, T7+, T10+, and T14+. For each Tp, the 4 ZmL were placed inside a 50 mL Falcon tube, euthanized (1 h, −20 °C), and then disinfected (70% ethanol, 3 h, 4 °C). After that, ZmL were homogenized using the Precellys^®^ Evolution Touch tissue apparatus (Bertin Technologies) [27].
To detect Sk−1, final homogenized samples were properly diluted in dPBS and 100 µL aliquots were plated on selective ChromID^®^ ESBL agar (bioMérieux) that were incubated overnight (36 ± 1 °C). After species identification using the MALDI-TOF MS (Brucker), colony forming unit (CFU)/mL counting for Sk−1 was performed for all the Tps.
Bacteriophage titers were evaluated using the DLA method, as previously done [27, 35]. In brief, homogenized samples from INTESTIbc*-*treated ZmL were filtered using a 0.22 μm pore size PES syringe (Carl Roth Gmbh). Brain Heart Infusion (BHI) agar (1.5%) was distributed as a first layer in a Petri dish. Then, 100 µL of the filtrated tissues was added to a 15 mL tube containing 1 mL of BHI broth. Subsequently, 100 µL of the previously characterized INTESTIbc-susceptible E. coli strain 56-M3-Ec (++++; confluent lysis [36]) or Sk−1 were combined with 4 mL of 0.6% agar BHI overlay and poured over the surface of a dried underlay plate. Upon an overnight incubation (36 ± 1 °C), plaques (if any) were counted to determine viral titers (PFU/mL) from samples obtained at T3 (null control), T5+, T7+, T10+, and T14+.
Characteristics of Sk-1 strains recovered over time
Selected Sk−1 strains (n = 12) recovered during ZmL experiments across multiple Tps and experimental conditions underwent AST and DLA assays. Results were compared to those obtained from the original Sk−1 strain.
The 12 strains also underwent hybrid WGS. gDNA was extracted using the PureLink™ Microbiome DNA Purification kit (Thermo Fisher Scientific). WGS was performed on a NovaSeq 6000 (Illumina; 2 × 150 bp paired-end protocol), and MinION™ (Nanopore; SQK-RBK114.24 rapid barcoding kit 24 V14, FLO-MIN114 R.10.4.1 flow cell, 72 h) sequencing platforms. Read pre-processing and the pipeline used to generate the hybrid assemblies were performed as described above. Hybrid genome assemblies were used to generate whole-genome single nucleotide variants (SNVs) alignments (chromosome and plasmids) with snippy v4.6.0, using the original Sk−1 genome as a reference, as previously described [31, 32, 37]. The input assemblies were further analyzed for structural variations implementing SyRI v1.7.1 using paired (vs. reference) genome filtered alignments (delta-filter: ‘- 90 -l 100’) generated with nucmer v3.1 and the ‘--maxmatch’ argument.
Microbiota characterization
Microbiota characterization for ZmL was achieved through 16S rRNA amplicon sequencing analysis as previously done [27]. Samples obtained at T0, T7/T7+, and T14/T14 + for ZmL were analyzed (Fig. 1).
Briefly, gDNA was isolated from the homogenized tissues of ZmL using the QIAamp PowerFecal Pro DNA Kit (Qiagen) and purified with CleanNGS magnetic beads (CleanNA). DNA concentration and purity were assessed with Qubit™ 3.0 and NanoDrop™ One^C^ (Thermo Fisher Scientific), respectively. gDNA preparations were sent to Eurofins Genomics GmbH (Germany) for amplification and sequencing of the bacterial 16S rRNA gene (V4 region). Genomic libraries were prepared using a PCR amplification with the primers 515 F Parada and 806R Apprill [38]. All samples were sequenced on an Illumina MiSeq platform (2 × 300 bp read lengths) and the resulting read quality was assessed with FastQC v0.12.1.
Raw sequence data in FASTQ format were processed with the DADA2 pipeline v1.22.0 in R programming language v4.4.2 [parameters: truncLen f/r 235/190, maxN = 0, maxEE = c(2,2), truncQ = 2, rm.phix = TRUE, compress = TRUE, and multithread = TRUE] using the SILVA reference database v138.1 Nr99 as previously described [27, 39]. Agglomerated amplicon sequence variants (ASVs) at the genus level and their relative abundances were determined using the phyloseq v1.50.0 R package.
Alpha diversity metrics [Shannon diversity index (SDI) and Observed richness (S_obs_)] were calculated on non-normalized ASV counts with phyloseq. Additionally, differentially abundant genera were assessed using the DESeq2 v1.46.0 R package, adjusted for batch variation, and applied on non-normalized ASV counts, specifying comparisons within-groups across time points (P-adj < 0.05) via the contrast argument. Microbial compositional variation was determined after cumulative-sum scaling (CSS) normalization from metagenomeSeq v1.48.1 to consider differences in sequencing depth, while further batch-to-batch correction was performed with ConQuR v2.0 with a penalized fitting strategy. Beta diversity was assessed using the Bray-Curtis dissimilarity index (BCDI) and principal coordinate analysis (PCoA) of the Bray-Curtis distance matrix with phyloseq.
Statistical analyses
Statistical analyses were performed using R v4.4.2 as previously done [27, 33]. Data normality and homogeneity of variances was assessed using the Shapiro-Wilk and Levene’s tests. Growth curves (CFU/mL) were statistically compared using a two-way ANOVA on log-transformed data (log_10_) to assess the interaction between experimental conditions and time points (stats package v4.4.2). Post hoc comparisons were carried out with estimated marginal means, while P values were adjusted with the Tukey method (emmeans v1.11.1).
For microbiota, alpha diversity was analyzed over time within each experimental condition with the non-parametric Kruskal-Wallis test (stats package v4.4.2). The effect of batch on the community composition at T0 was assessed by permutational multivariate analysis of variance using distance matrices (PERMANOVA, adonis2 function; with 9999 permutations) from the vegan v2.6.10 R package using the Bray-Curtis distance matrix (CSS transformed and batch-corrected). Results were considered statistically significant when P value < 0.05. All packages were set to default parameters, unless otherwise indicated. Graphical analyses were visualized with the R package ggplot2 v3.5.2 and modified in Inkscape v1.4 for style.
Results
Further data about Sk-1
The tested S. enterica strain was phenotypically susceptible (“++++”: complete clearing) to the INTESTIbc (Table 1). The resulting genome consisted of a circular chromosome (4,841,872-bp) and 4 plasmids (including a ~ 63,5-Kb IncL-blaOXA−48).
A total of 265 putative VF genes were detected in Sk−1 (File S1), including sseK2 that is linked to biofilm formation and survival under difficult conditions [40]. Concerning the VF genes usually associated with S. enterica human infection, only pipB2 and sifA were detected, whereas Sk−1 lacked of grvA, sseI, sopE, sodCI, sopD2, sspH2, srfH, and shdA [17, 41–43].
ZmL colonization and treatment with bacteriophage
As depicted in Fig. 2 (crude data in Table S1), after providing 3 days of contaminated food, untreated ZmL remained persistently colonized with S. enterica Sk−1 strain (e.g., mean of 4.3 × 10^6^ and 5.9 × 10^4^ CFU/mL for T3 and T14, respectively). Moreover, administration of dPBS on T3 and T5 did not significantly influence Sk−1 load. On the other hand, treatment with two doses of INTESTIbc induced a statistically significant overgrowth of Sk−1 that reached 4.0 × 10^6^ CFU/mL on T10+ (P < 0.01 for ZmL treated vs. untreated and P < 0.05 vs. the control with dPBS). Finally, bacteriophages targeting E. coli 56-M3-Ec were recovered on T5 + and T7+ (titers 1.1 × 10^4^ and 4.6 × 10^3^ PFU/mL, respectively), while those against Salmonella Sk−1 were detected on T5+, T7+, and T10+ (3.1 × 10^4^, 3.4 × 10^4^, and 7.3 × 10^3^ PFU/mL, respectively).
Fig. 2. Gut colonization of Z. morio larvae (ZmL) by S. enterica strain Sk−1. Y axis: colony forming unit (CFU)/mL or phage forming unit (PFU)/mL [mean ± standard error (SEM; n = 3)]. X axis: time points (T, ZmL not receiving INTESTIbc; T+, ZmL receiving INTESTIbc). At T3, ZmL were randomly assigned to 3 groups: untreated, dPBS-treated and INTESTIbc-treated. dPBS and INTESTIbc groups received two oral force-feedings (10 µL) at T3 and T5. LOD, limit of detection. ns, not significant
Notably, ZmL mortality after administering food contaminated with Sk−1 was 0% at both T3 and T7. Moreover, mortality after force-feeding with INTESTIbc or dPBS was 12% and 7% at T7+/T7, respectively (Table S2).
Dynamics of ZmL microbiota
As shown in Fig. 3, the baseline microbiota (before administration of Sk−1 at T0) was commonly rich in Lactococcus, Pediococcus,* Staphylococcus*, and Latilactobacillus genera (relative abundance, range: 28–57%, 11–40%, 2–40% and 0.1-8%, respectively). Genera belonging to the Enterobacteriaceae family were also present, predominantly represented by Pseudocitrobacter (relative abundance, range: 2–11%), while Escherichia/Shigella were notably absent (File S2). These relative abundance patterns across groups were also supported by differential abundance analysis. In particular, Lactococcus,* Pediococcus*,* Staphylococcus*,* Latilactobacillus* and Pseudocitrobacter were shown to be significantly decreased at T7 compared to T0 (File S4).
Fig. 3. Comparison of the top 15 most abundant genera per sample across experimental groups. Relative community composition of ZmL gut microbiota was determined by 16S rRNA gene amplicon sequencing of the V4 region at the genus-level, based on three experiments across time points (T0, T7/T7+, and T14/T14+). Symbols star and square indicate experiments that share the same ZmL (T0). Panels show the top 15 genera identified from a total of 26 genera obtained from 292 amplicon sequencing variants (ASVs). All remaining genera are grouped under “Other”. SDI, Shannon diversity index. S_obs_, observed richness
Alpha diversity analysis indicated that the initial microbiota at T0 was heterogeneous (SDIs and S_obs_ of 1.3–2.8 and 19–48, respectively). Nevertheless, both parameters were not statistically significantly different between the ZmL belonging to the same experimental condition and over time (i.e., on T0, T7/T7 + or T14/T14+) (Fig. 4). Remarkably, beta diversity analysis indicated that the microbiota heterogeneity observed at T0 was mainly due to the compositional differences of the two distinct ZmL batches utilized for the experiments, as shown by the clear clustering separation by batch in the PCoA ordination (P = 0.0128; Figure S2).
Fig. 4. Alpha diversity analysis using the SDI (A) and S_obs_ (B). Each condition included three experimental replicates (Exp-1, Exp-2, Exp-3), with samples collected at T0, T7/T7 + and T14/T14+. Condition is color-coded and time points are indicated by symbol shape. Above each condition (inverted bracket) ‘ns’ represents no statistically significant differences over time within that group
After administering the two INTESTIbc doses, microbiota patterns showed a decrease in the relative abundance and number of genera (e.g., Enterococcus,* Staphylococcus* and Pseudocitrobacter), with SDIs and S_obs_ of 1.5–1.8 and 23–35, respectively at T14+. In contrast, it was evident the emergence of the Salmonella genus (relative abundance, range: 0.4-3% on T7+ and 0-0.6% on T14+) (Fig. 3D; File S2-F). Differential abundance testing further corroborated these genera-level differences, which identified a strong reduction in abundance of Enterococcus, Staphylococcus and Pseudocitrobacter at T7. On the other hand, a significant increase of Salmonella at T7+ and T14+ was recorded (File S4-D). However, alpha diversity analysis showed that these over time microbiota changes were not statistically significant (Fig. 4).
Features of Sk-1 recovered during experiments
Twelve representative S. enterica Sk−1 strains were recovered during different ZmL experiments at T7/T7+, T10/T10+, and T14/T14+. As summarized in Table S3, all 12 strains demonstrated consistent antibiotic and INTESTIbc susceptibility profiles as for the original Sk−1.
Genome alignment (98.5%) analysis indicated that each of the 12 strains possessed ≤ 3 SNVs when compared to the initial reference Sk−1 genome (Table S4). We also note that the analysis demonstrated small structural changes when compared to the reference. For instance, ≤ 22 and ≤ 21 SNVs and ≤ 3 and ≤ 2 insertions for Sk−1 strains recovered during experiments without and with INTESTIbc treatment, respectively (File S3).
Discussion
In agreement with the 3Rs guidelines, insect larvae may represent an alternative to complex animal models for studying the effects of gut bacterial pathogens [25]. In particular, GmL have been used in the past as an infection model to study the virulence of Salmonella enterica and potential novel treatment approaches (e.g [44–46]). Nevertheless, the use of GmL for studies focusing on intestinal colonization has limitations. First, GmL possess a very limited microbiota, dominated by Enterococcus spp. [47], which does not mimic the richness observed in the human gut. Second, microbiota manipulation via contaminated food administration is impracticable, as GmL do not feed voluntarily [29]. Third, GmL may undergo high mortality rates during experiments using Enterobacteriaceae, especially if the force-feeding approach is implemented [27, 48]. For instance, Nale JY *et al., *colonized GmL with S. enterica strains to demonstrate the therapeutic potential of a three-phage cocktail. However, many optimization experiments were necessary to determine the right oral gavage inoculum for each specific strain to avoid high mortality rates [49].
ZmL were stably colonized by Sk-1
In this study, we exploited the unique biological features of ZmL to study their susceptibility to colonization by a hyperepidemic ST198 S. enterica serovar Kentucky isolate (Sk−1) which produces the pandemic OXA-48 carbapenemase [7–15, 26]. As a result, untreated ZmL remained stably colonized for 14 days with Sk−1 (T14: ~10^4^ CFU/mL) after 3 days of feeding with contaminated food (Fig. 2). We emphasize that this approach resembles natural Salmonella infections, which are typically acquired via the oral route [1–3, 9]. On the other hand, we did not use the force-feeding administration - as we have previously done with MDR E. coli [33] - because it does not mimic the natural acquisition of S. enterica.
ZmL underwent low mortality
In the study of Nale YT et al., the mortality rates of untreated GmL that underwent force-feeding with 3 different S. enterica isolates (all serovar Typhimurium) were > 80% at 72 h [49]. In contrast, the mortality of untreated ZmL colonized with Sk−1 was zero at both T5 and T7 (Table S2). This positive effect may be due to i) the harmless oral route of administration via the diet, ii) the lower quantity of pathogen intake compared to the oral gavage, and/or iii) the superior physiologic features of ZmL compared to those of GmL [26]. Nevertheless, considering the results of the VFs analysis (File S1), it should also be hypothesized that Sk−1 is less virulent than the 3 Typhimurium strains tested by Nale YT et al. [49]. In particular, the pattern of VF genes detected in Sk−1 suggests that the strain is more prone to induce ZmL colonization rather than clinical illness [17, 40–43].
The mortality rates of colonized ZmL that received two force-feeding administrations (T3 and T5/T5+) with INTESTIbc or dPBS were also low (≤ 12% at T7+/T7; Table S2), as we have observed in a previous study [33]. Overall, these findings indicate that ZmL are particularly resilient to receive oral injections, supporting the suitability of this model for future experiments of similar design.
INTESTIbc treatment was ineffective
The INTESTIbc is a commercially available product used for the treatment or prophylaxis of enteric bacterial infections (per os: 10 mL 4 times per day for 5–6 days or 10–20 mL daily every 5 days over the course of 1 month, respectively), including those due to S. enterica (https://phage.ge/en/products/phago-intesti) [34]. However, to our knowledge, the activity of this specific bacteriophage cocktail against S. enterica colonization/infection and its impact on the microbiota have not yet been evaluated in vivo. Therefore, we used this product in our study to assess its capacity in decolonizing ZmL from S. enterica Sk−1. Notably, due to potential harm from repeated force-feeding, ZmL received only two doses of INTESTIbc, as in our previous studies [27, 33]. Nevertheless, we believe that this approach still provides valuable proof-of-concept data that may be translatable to the human clinical context.
In ZmL receiving INTESTIbc, bacteriophages were detected only between T5+ and T10+ (Fig. 2). We speculate that their absence on T14 + may reflect an inability to locate suitable bacterial hosts for replication and/or because the ZmL gut does not provide an optimal environment (e.g., pH, temperature) for viral stability [50]. In this context, the overgrowth of Sk−1 between T5+ and T10+ may partly account for the longer persistence of its specific active bacteriophages compared with those targeting E. coli 56-M3-Ec.
After the first dose of INTESTIbc, ZmL showed an initial decrease in Sk−1 concentration (T5: 2 × 10^5^ CFU/mL), but an unanticipated regrowth occurred following the second administration and continued through T10+ ( 4 × 10^6^ CFU/mL), showing a statistically significant difference compared with the control curves (Fig. 2). To explain this phenomenon, we initially hypothesized that Sk−1 underwent mutations conferring resistance to bacteriophages, as previously noted by others using mono-phage therapies in GmL colonized with S. enterica [24, 44, 49]. However, both phenotypic and genotypic analyses did not support this hypothesis, instead indicating the stability of Sk−1 during the 14 days of experiments (Table S3, Table S4 and File S3).
We therefore speculate that the overgrowth of S. enterica Sk−1 under INTESTIbc treatment was linked to the effect of the overall multiple lytic phages (cocktail) on the complex bacterial population residing within the ZmL gut. In particular, we hypothesize that there were specific bacterial competitors of Sk-1 in the intestinal tract of ZmL that could partially protect against its colonization (a phenomenon known as “colonization resistance”) [51]. Since the INTESTIbc may inhibit multiple species [34] - including the putative competitors - Sk-1 could overgrowth in ZmL following the peak of bacteriophages observed between T5+ and T7+ (Fig. 2). In this context, microbiota analyses have provided several clues about the possible Sk−1 competitors (see below).
Microbiota underwent small but critical changes
ZmL microbiota was characterized over time to assess the effect of INTESTIbc on the microbial community in larvae colonized with S. enterica Sk−1. Notably, our previous analysis of ZmL challenged with MDR E. coli strains (with or without administering INTESTIbc) revealed a non-significant reduction of the bacterial diversity during experiments [27].
In the present study, the starting ZmL microbiota patterns at T0 were heterogeneous, particularly given that two distinct batches of larvae were used in the experiments. Nevertheless, this variation reflects the natural diversity of their microbiota, with many bacterial genera also encountered in the human microbiota (Fig. 3 and Figure S2). Similarly, the human microbiota can vary substantially at the individual level due to various factors (e.g., diverse diet, age, ethnicity, travels) [52–54]. Therefore, these observations suggest that the dynamics of the ZmL microbiota recorded under the INTESTIbc challenge may mimic those that could occur in the human gut.
As anticipated, administration of INTESTIbc to ZmL colonized by Sk−1 induced significant overgrowth of the pathogen as confirmed by the 16S rRNA microbiota analysis of T7+ and T14+ samples (Fig. 3D, File S4-D). Simultaneously, INTESTIbc targets several genera that tend to decrease in abundance - or even disappear entirely - at T7+ and/or T14+. In particular, a specific negative effect against Pseudocitrobacter was noted [relative abundance range: T0: 4.69–27.89% vs. T14+: 0.49–9.46% (File S2-F); mean log_2_ fold change range: T7+: −23.6 vs. T14+: −23.7 compared to T0 (File S4-D)]. This genus - belonging to the Enterobacteriaceae - has been rarely found in cockroaches and in culture-based fecal samples of humans [well summarized in [55]. Nevertheless, the scarcity of data regarding Pseudocitrobacter - including those present in gut microbiome datasets - may just reflect its recent classification. It should also be noted that the routinely used MALDI-TOF MS or VITEK 2 systems may misidentify Pseudocitrobacter spp. as Pantoea spp. [55].
It should be emphasized that Enterobacteriaceae represent < 1% of the total bacteria detected in the gut microbiota of healthy individuals [56]. However, mouse model experiments have demonstrated that these bacteria - especially E. coli - play a major role in the colonization resistance against Salmonella infections [57, 58]. Therefore, similar to the effects of broad-spectrum antibiotics [20], INTESTIbc might negatively affect the competitive activity of some endogenous Enterobacteriaceae against S. enterica infection. This effect may occur in ZmL, animals, or humans, although the specific protective Enterobacteriaceae species affected by INTESTIbc are likely to differ between hosts (e.g., Pseudocitrobacter spp. in ZmL vs. E. coli in humans).
Conclusions
In this work, we demonstrated that ZmL can be easily and stably colonized with S. enterica without affecting larval survival. Therefore, this new in vivo model may represent an inexpensive, simple, fast, and reliable alternative to the gold-standard mouse model for studying several key aspects of this important enteric human pathogen (e.g., development of novel treatment or decolonization strategies) [9, 16, 17, 25].
As a proof-of-concept, we used the INTESTIbc to decolonize the ZmL carrying an hyperepidemic OXA-48-producing ST198 S. enterica [7–15]. Accordingly, the commercial bacteriophages cocktail was ineffective, likely because its disruptive impact on the ZmL microbiota led to pathogen overgrowth. This phenomenon may be associated to the inhibition of the Pseudocitrobacter genus [55], a member of the Enterobacteriaceae which appears not significantly present in the human microbiota, but may confer a protective effect against S. enterica colonization in ZmL.
Notably, the overgrowth of Sk−1 observed with our in vivo model was transient (i.e., the Sk−1 load returned to baseline at T14+) following the two consecutive INTESTIbc administrations. However, it is plausible that during prolonged treatment in humans or animals (e.g., 5–6 days as suggested) [34], repeated INTESTIbc doses may continue to promote the surplus of the pathogen at the intestinal level.
Regardless of host-specific differences in Enterobacteriaceae species that mediate colonization resistance, further in vivo studies are needed to assess the suitability of using bacteriophages to treat human/animal intestinal infections or colonization due to hyperepidemic MDR clones of S. enterica. In fact, while the mono-phage treatment is at risk of selecting a resistant phenotype [24, 44, 49], our results suggest that the use of broad-spectrum cocktails could promote a dangerous overgrowth of the targeted enteric pathogen. Therefore, the development and use of commercial formulations containing several lytic bacteriophages targeting only the specific enteric pathogen is envisioned for future in vivo studies followed by clinical trials [24, 44, 49].
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Clement M, Keller PM, Bernasconi OJ, Stirnimann G, Frey PM, Bloemberg GV, Sendi P, Endimiani A (2019) First clinical case of In vivo acquisition of DHA-1 plasmid-mediated Amp C in a Salmonella Enterica subsp. Enterica isolate. Antimicrob Agents Chemother 63(10):e 00992–1910.1128/AAC.00992-19PMC 676153531358582 · doi ↗ · pubmed ↗
- 2Biggel M, Horlbog J, Nuesch-Inderbinen M, Chattaway MA, Stephan R (2022) Epidemiological links and antimicrobial resistance of clinical Salmonella enterica ST 198 isolates: a nationwide microbial population genomic study in Switzerland. Microb Genom 8(10):mgen 00087710.1099/mgen.0.000877 PMC 967605236301086 · doi ↗ · pubmed ↗
- 3Samper-Cativiela C, Dieguez-Roda B, Trigo da Roza F, Ugarte-Ruiz M, Elnekave E, Lim S, Hernandez M, Abad D, Collado S, Saez JL, de Frutos C, Aguero M, Moreno MA, Escudero JA, Alvarez J (2022) Genomic characterization of multidrug-resistant Salmonella serovar Kentucky ST 198 isolated in poultry flocks in Spain (2011–2017). Microb Genom 8(3):00077310.1099/mgen.0.000773 PMC 917628035259085 · doi ↗ · pubmed ↗
- 4Rabiu AG, Fadiji O, Adesoji AT, Jimoh SO, Cho S (2025) A review of the environmental spread of Salmonella enterica serovars through water in Africa. Lett Appl Microbiol 78(2):ovaf 00510.1093/lambio/ovaf 00539848915 · doi ↗ · pubmed ↗
- 5Galan-Relano A, Valero Diaz A, Huerta Lorenzo B, Gomez-Gascon L, Mena R, Mf, Carrasco Jimenez E, Perez Rodriguez F, Astorga Marquez RJ (2023) Salmonella and Salmonellosis: an update on public health implications and control strategies. Animals (Basel) 13(23)10.3390/ani 13233666 PMC 1070559138067017 · doi ↗ · pubmed ↗
- 6Rumbos CI, Athanassiou CG (2021) The superworm, Zophobas morio (Coleoptera:Tenebrionidae): a ‘sleeping giant’ in nutrient sources. J Insect Sci 21(2)10.1093/jisesa/ieab 014PMC 803324733834209 · doi ↗ · pubmed ↗
- 7Aldeia C, Campos-Madueno EI, Endimiani A (2024) Genomic insights into Leminorella grimontii and its chromosomal class A GRI b-lactamase. Eur J Clin Microbiol Infect Dis 43(9):1855–186010.1007/s 10096-024-04888-7PMC 1134977238958811 · doi ↗ · pubmed ↗
- 8Worley MJ (2025) Salmonella type III secretion system effectors. Int J Mol Sci 26(6)10.3390/ijms 26062611 PMC 1194232940141253 · doi ↗ · pubmed ↗