First molecular detection of Babesia caballi in camel associated ticks in Uzbekistan, with no evidence of Anaplasma or Rickettsia
Alisher Safarov, Noureddine Mechouk, Alireza Sazmand, Angela Monica Ionică, Ansorkhon Maksudov, Baurjan Kunisov, Georgiana Deak

TL;DR
Camels in Uzbekistan were found to host ticks carrying Babesia caballi, a parasite previously not reported in this region or in camels.
Contribution
This is the first molecular detection of Babesia caballi in camel-associated ticks in Uzbekistan.
Findings
Babesia caballi DNA was detected in two tick pools from camels in Uzbekistan.
No evidence of Anaplasma or Rickettsia was found in the ticks.
Ticks identified included species from the genera Hyalomma, Dermacentor, and Rhipicephalus.
Abstract
Dromedary camels (Camelus dromedarius) have an important role in food and textile production in arid and semi-arid regions of the world, yet data on their tick-borne pathogens (TBPs) is scarce, especially in Central Asia. This study aimed to: (i) identify the ticks infesting camels from different regions of Uzbekistan, (ii) molecularly detect TBPs in the collected ticks for the first time. In 2024, 290 camels from three regions (Karakalpakstan, Jizzakh, and Navoi) were examined for tick infestation. Ticks were collected, morphologically identified, pooled, and screened using PCR and sequencing for Babesia., Theileria., Anaplasma., and Rickettsia. Overall, 32.6% of camels harbored ticks, with 993 specimens collected, belonging to the genera Hyalomma, Dermacentor, and Rhipicephalus, comprising at least six species. Molecular analysis revealed the presence of Babesia caballi DNA in two…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
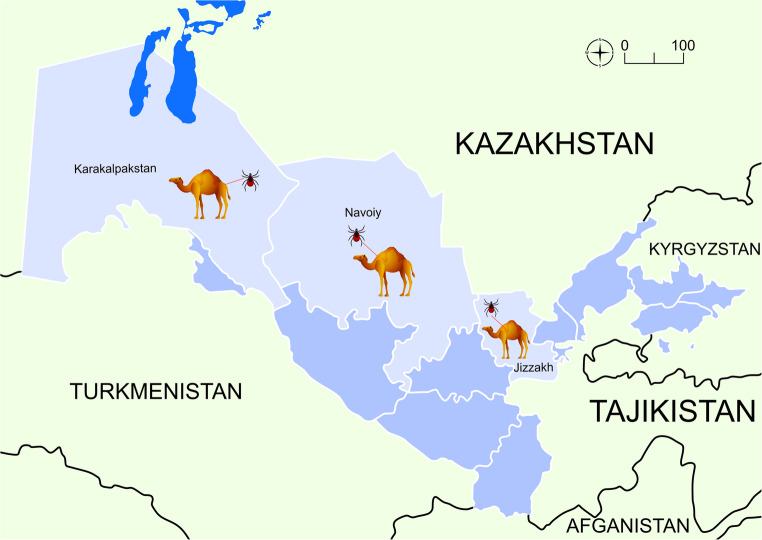 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6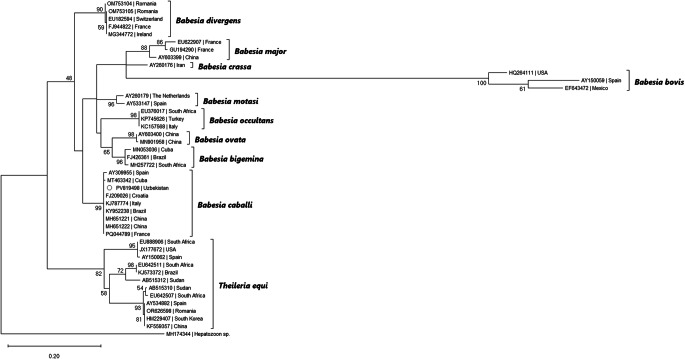 Figure 7
Figure 7- —the Ministry of Research, Innovation, and Digitization, CNCS-UEFISCDI
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Bartonella species infections research · Vector-Borne Animal Diseases
Introduction
Dromedary camels (Camelus dromedarius), with a world population of more than 41 million, are a valuable source of food, wool, skin, and labor in 47 countries around the world; however, there is little in-depth information about the parasites infecting them compared to other livestock (FAO, 2025). In particular, tick-borne pathogens that pose a serious threat to humans, domestic and wildlife animals, and have not been the focus of molecular studies until recently (Sazmand et al. 2019; Alanazi et al. 2020; Selmi et al., 2019).
Camels can be frequently infested by Ixodidae ticks, especially species of the genera Hyalomma, Dermacentor, Amblyomma, and Rhipicephalus, which are widely distributed in arid and semi-arid ecosystems (Ballweber 2009; Getange et al. 2021). Among these, Hyalomma spp. are considered the most frequent ticks in camels worldwide and are recognized as vectors of various pathogens affecting both animals and humans (El-Alfy et al. 2024; Soliman et al. 2024). Camels often share grazing areas with equines, cattle, sheep, and goats, creating ecological conditions favorable for tick exchange and interspecific transmission of some pathogens. Babesia, Theileria, Anaplasma, and Rickettsia are tick-borne pathogens with global distribution and infect blood cells of mammalian species, some being zoonotic (Uilenberg 1995; Young et al. 2019). Infections with these parasites lead to anemia, loss of productivity, and reduced production of milk and meat (Selmi et al. 2019). Several Theileria, Babesia, Anaplasma, and Rickettsia spp. are known to infect camels (Sazmand et al. 2019; Sazmand and Joachim, 2017; Alanazi et al. 2020; Selmi et al., 2019).
In dromedary camels from Egypt, molecular investigations demonstrated the presence of several piroplasmid species, including T. equi, B. caballi, B. bigemina, B. bovis, B. vulpes, as well as undetermined Babesia and Theileria spp., using molecular tools (Mahdy et al. 2023).This study provided the first molecular evidence of B. vulpes infection in camels, expanding the known host range of this piroplasmid species (Mahdy et al. 2023).
Molecular detection of Anaplasma spp. has been reported from camels in several countries in Africa, the Middle East, and Asia, with lower prevalence values than those reported in cattle (El-Alfy et al. 2024). Camels often exhibit mild or no clinical signs, raising the possibility that they act as subclinical carriers (Hekmatimoghaddam et al. 2012; Qablan et al. 2012; Sazmand et al., 2019).The genus Rickettsia (Rickettsiales: Rickettsiaceae) comprises obligate intracellular bacteria of major zoonotic concern that are transmitted by ticks through transstadial and, in some species, transovarial routes, enabling ticks to serve both as vectors and reservoirs (Kim 2022). Several Rickettsia spp. have been detected in ticks infesting camels, especially in Hyalomma spp., which are suitable vectors of pathogenic rickettsiae affecting humans and animals (Alanazi et al. 2020). Although camels are not considered primary hosts for Rickettsia spp., their role in sustaining infected tick populations and facilitating human exposure in pastoral systems should not be underestimated, mainly in arid environments with a close contact between humans, camels, and ticks.
In Central Asia, Uzbekistan ranks third in the camel population, with 18,400 heads, only after Turkmenistan and Kazakhstan (FAOSTAT, 2025). In the country, camel breeding is well developed in four regions: the Navoi region (44.3%), the Republic of Karakalpakstan (24.1%), the Bukhara region (11.9%), the Kashkadarya region (8.1%). In other regions, 11.6% of the total number of camels can be found. In Uzbekistan, camels are mainly used for milk, meat, and wool production (Safarov et al. 2024; Kunisov et al. 2024b). Although endoparasites of camels have been studied in Uzbekistan (Sultanov et al. 1973, 1975; Saparov 2006; Azimov et al. 2015; Kunisov et al. 2024a, b; Safarov et al. 2024), there is a shortage of information about tick-borne pathogens (TBPs), although Uzbekistan’s tick fauna comprises 35 species from two families: Ixodidae and Argasidae, distributed across nine genera (Rasulov 2007; Mechouk et al. 2025). In the only previous study, Esonboev et al. (2024) reported Hyalomma scupense from camels of the Jizzakh region in Uzbekistan. Likewise, there is no information about the infection of camels with TBPs, although theileriosis has been reported from cattle in several regions of Uzbekistan (Kuchkarova 2023). Considering the shortage of information about ticks and TBPs related to camels and the roles played by camels in the epidemiology of TBPs in Uzbekistan, this study aimed to: (i) identify the ticks infesting camels from different regions of the country, (ii) molecularly detect TBPs in the collected ticks.
Materials and methods
Sampling procedures
From July 10 to August 18, 2024, a total of 290 apparently healthy dromedary camels were inspected for tick infestation. The animals lived in three regions of Uzbekistan with different climatic conditions, i.e., Karakalpakstan (n = 122), Jizzakh (n = 52), and Navoi (n = 116) (Fig. 1). In most cases, camels shared the pasture with horses and cattle. When observed, ticks were collected in individual plastic containers and stored in 70% ethanol. Each tick was individually morphologically identified using valid keys (Estrada-Peña et al. 2018), and photographs were taken using an optical microscope (Olympus BX61) connected to a digital camera (DP72 with Cell∧F imaging software Olympus Corporation, Tokyo, Japan). Figures 2, 3, 4, 5, and 6.Fig. 1. Map showing the investigated areas of UzbekistanFig. 2Top: (A) Dermacentor niveus male ventral view; (B) Dermacentor niveus male dorsal view. Bottom: A. Dermacentor niveus female dorsal view. B. Dermacentor niveus female ventral viewFig. 3Top (A) Hyalomma anatolicum male dorsal view, (B) Hyalomma anatolicum female ventral view. Bottom: A. H. anatolicum female ventral view, B. H. anatolicum female dorsal viewFig. 4(A) Hyalomma dromedarii male dorsal view, (B) H. dromedarii male ventral viewFig. 5(A) Hyalomma scupense female ventral view, (B) H. scupens female dorsal viewFig. 6Top: (A) Hyalomma spp. female ventral view, (B) Hyalomma spp. female dorsal view. Bottom: (A) Rhipicephalus spp. female ventral view, (B) Rhipicephalus spp. dorsal view
DNA isolation from camel ticks, molecular analysis by PCR and sequencing
For DNA isolation, ticks were divided into 34 pools, each containing several ticks of the same species from animals from different locations. More specifically, from each geographical region, 4–5 ticks of the same genus were selected and considered an individual pool. When more than one genus/species was detected in one animal from a region, a separate pool was done. Ticks were selected based on their engorgement status, with preference given to less engorged individuals, as these are more likely to yield pathogen DNA with minimal host contamination. In cases where only non-engorged ticks were available, a single tick was selected for analysis (Supplementary file). Each tick was cut in half, and DNA was isolated using a commercial kit (ISOLATE II Genomic DNA Kit, Meridian Bioscience, London, UK), according to the instructions provided by the manufacturer. Specimens were screened by PCR for the presence of DNA of piroplasmids, Anaplasma spp., Ehrlichia spp., and Rickettsia spp. using previously published primers (Table 1) and protocols (Matei et al., 2021; Corduneanu et al. 2023; Mechouk et al., 2024). All PCR reactions included a no-template control consisting of PCR water and a positive control represented by pathogen DNA, previously confirmed by sequencing during other studies. The samples were visualized by electrophoresis (Supplementary file). DNA from individual ticks was not tested when pools were negative. PCR-positive products were purified using the/PCR DNA Fragments Kit (Geneaid Biotech, New Taipei, Taiwan) and bidirectionally sequenced using an external service (Macrogen Europe B.V., Amsterdam, The Netherlands). The sequences were assembled using Geneious software (Biomatters Ltd., New Zealand) and compared to other sequences available in the NCBI GenBank^®^ database by Basic Local Alignment Search Tool (BLAST) analysis.
Table 1. Table showing the primers used for the current studyPathogenTarget geneProduct size (bp)Forward primerReverse primerReferencePiroplasmids and Hepatozoon spp.(nested PCR)18 S rRNA561–620BTH-1 F: CTGAGAAACGGCTACCACATCTBTH-1R: TTGCGACCATACTCCCCCCA Hodžić et al. 2018GF2: GTCTTGTAATTGGAATGATGGGR2: CCAAAGACTTTGATTTCTCTC Bonnet et al. 2007SFG Rickettsia gltA 381Rsfg877: GGGGGCCTGCTCACGGCGGRsfg1258: ATTGCAAAAAGTACA GTGAACA Regnery et al. 1991Anaplasmataceae16 S rRNA345EHR16SF: GGTACCYACAGAAGAAGTCCEHR16SR: TAGCACTCATCGTTTACAGC Parola et al. 2000
The phylogenetic analysis was conducted using MEGA X software (Kumar et al., 2018). The sequence attained during the present study was aligned with 42 other Babesia spp. and Theileria equi sequences, and an additional Hepatozoon spp. sequence used as outgroup, downloaded from GenBank^®^, using the MUSCLE algorithm. The evolutionary history was inferred by using the Maximum Likelihood method and Tamura-Nei model (Tamura and Nei 1993), with a discrete Gamma distribution used to model evolutionary rate differences among sites (5 categories (+ G, parameter = 0.1581).
Statistical analysis
Prevalence (i.e., proportion of hosts infected with ticks), tick burden (i.e., arithmetic mean number of ticks per infected host), and pathogen infection rate were estimated using the Quantitative Parasitology program recommended by Rózsa et al. (2000). Confidence intervals (95% CI) were calculated. Differences among regions were assessed by chi-square testing and were considered significant at p < 0.005.
Results
Of the 290 camels examined, 88 (32.59%; 95% CI: 27.04–38.5) were infested by ticks, with a total of 993 specimens collected: 482 (48.54%, 95% CI: 45.4–51.6) males, 439 (44.2%, 95% CI: 41.1–47.3) females and 72 (7.2%, 95% CI: 5.8–9.03) nymphs. Ticks have been recorded on camels distributed across all three study regions: Karakalpakstan – 51 (41.8%; 95% CI: 32.9–51); Jizzakh – 21 (18.1%; 95% CI: 11.6–26.3); Navoiy – 16 (30.7%; 95% CI: 18.7–45.1). The prevalence of tick infestations was significantly higher in Karakalpakstan region when compared to Navoiy (p < 0.0001). The differences between Jizzakh and the other two regions were not statistically significant (p = 0.18 against Karakalpakstan and p = 0.08 against Navoiy). The highest intensity of infestation was recorded in Karakalpakstan, with a median of 11 ticks/camel (IQR: 10–16), followed by Navoiy with a median of 4 ticks/animal (IQR: 3–10), and Jizzakh, with a median of 3 ticks/camel (IQR: 1.5–7.5). The complete data about camels, including the collection date, age, sex, and locality, are shown in the Supplementary file.
All ticks were morphologically identified as belonging to three genera (Hyalomma, Dermacentor, Rhipicephalus) and at least six species (Table 2; Supplementary file). Some ticks were not identified to the species level due to their poor preservation and invisible morphological structures. Information on the age and sex of the affected camels and their origin is presented in the Supplementary file.Table 2. Tick species identified and their development stagesTick speciesCamels affectedTotalDevelopmental stagesN/88 (%)95% CIFemaleMaleNymphHyalomma anatolicum (Fig. 3)72 (81.82)72.16–89.24440533870Hyalomma dromedarii (Fig. 4)17 (19.32)11.68–29.12490490Hyalomma scupense(Fig. 5)1 (1.14)0.03–6.171100Hyalomma spp. (Fig. 6)71 (80.68)70.88–88.32386308672Dermacentor niveus (Fig. 2)3 (3.41)0.71–9.6410868400Rhipicephalus sanguineus s.l.7 (7.95)3.26–15.79900
Out of 34 tested pools (131 ticks), two were positive for piroplasms (Supplementary file). Sequencing revealed Babesia caballi in the two positive samples, one from a pool of H. anatolicum male ticks from Karakalpakstan and the second from a pool of D. niveus from Jizzakh, both showing a 100% similarity with over 30 other entries from GenBank, e.g., in the blood of equines worldwide, in France (PQ044790), Spain (AY534883), Brazil (KY952238), Cuba (MT463341), South Africa (EU642512). The sequence was deposited in GenBank under the accession number PV819498. Theileria spp. was not identified in none of the two positive samples for piroplasmids. The phylogenetic analysis also placed the herein obtained sequence in a clade including exclusively B. caballi isolates (Fig. 7).Fig. 7. Bootstrap consensus tree. The percentage of trees in which the associated taxa clustered together is shown next to the branches (values below 40% not shown). The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. This analysis involved Babesia nucleotide sequences, and one Hepatozoon sequence used as outgroup
All samples tested negative for Anaplasmataceae and Rickettsia spp.
Discussion
This study reports the first molecular identification of B. caballi in Uzbekistan, expanding current knowledge on tick-borne parasites in Central Asia. Of note, the detection of B. caballi in ticks collected from camels in Uzbekistan suggests the potential role of camels and their associated ticks in the epidemiology of equine piroplasms in Central Asia. Previously, in neighboring Kazakhstan, B. caballi was identified in Dermacentor marginatus collected from cattle (Chimkent − 5.6% and Saryagash district − 30%) and horses (10%, Karabulak district). In another study from Kazakhstan, Babesia caballi was not detected in Hyalomma asiaticum collected from camels and Rhipicephalus turanicus ticks collected from dogs, using molecular techniques (Chunli et al. 2021). Additionally, in Kyrgyzstan, B. caballi was reported in the blood of 8.4% (160/1900) of imported donkeys from the Osh region, southwestern Kyrgyzstan (Wu et al. 2024).
Although B. caballi is normally associated with equines, the identification in ticks from camels could represent an interspecific transmission, which might have occurred in mixed-grazing systems. In many parts of Africa and Asia, including Uzbekistan, camels often share grazing areas with other animals such as cattle and equines, particularly horses and donkeys (Alanazi et al. 2020). This practice of co-grazing could facilitate exposure to ticks, making cross-species transmission of TBPs possible. Babesia caballi was identified in the blood of camels using molecular tools in Iraq (39.5%, 31.6%; Hussein et al. 2015), Iran (19.3%; Mirahmadi et al. 2022), Jordan (6%; Qablan et al. 2012), and Egypt (5.4–8.3%; Mahdy et al. 2023; Amer et al. 2024) (Table 3). Theileria equi, the causative agent of equine theileriosis, has been identified in the blood of camels in Egypt (41%, 4.4%; Mahdy et al. 2023), Iraq (23.68% Hussein et al. 2015), Jordan (4%; Qablan et al. 2012), and Iran (2.8%; Bahrami et al., 2016). It is known that equine piroplasmosis (EP), caused by B. caballi and T. equi, causes significant economic losses in the equine industry.
Table 3. Table shows a revision of all studies investigating Babesia caballi and Theileria equi in camels or camel-associated ticksPathogenHostPrevalence % (positive/total)CountryReference Babesia caballi
Camelus dromedarius 5.9 (2/34)UzbekistanCurrent study Camelus dromedarius 6 (6/100)Jordan Qablan et al. 2012 Camelus dromedarius 5.4 (11/203)Egypt Mahdy et al. 2023 Camelus dromedarius 1.2 (3/248)Iran Bahrami et al. 2017 Camelus dromedarius 39.5 (15/38)Iraq Jasim et al. 2015 Camelus dromedarius 83.8 (62/74)Iraq Al-Obaidi et al. 2021 Camelus dromedarius 62.2 (51/82)Egypt Kuraa and Malek 2024 Camelus dromedarius 8.3 (15/181)Egypt Amer et al. 2024 Camelus dromedarius 19.3 (27/140)Iran Mirahmadi et al. 2022 Hyalomma dromaderii 0.2 (1/593)Nigeria Onyiche et al. 2020 Theileria equi
Camelus dromedarius 4 (4/100)Jordan Qablan et al. 2012 Camelus dromedarius 41 (84/203)Egypt Mahdy et al. 2023 Camelus dromedarius 4.4 (8/181)Egypt Amer et al., 2024 Camelus dromedarius 2.8 (7/248)Iran Bahrami et al. 2017 Camelus dromedarius 23.7 (9/38)Iraq Jasim et al. 2015
Economic losses include treatment costs, abortions, loss of activity, and death. Clinical signs of equine babesiosis and theileriosis vary, but share fever, anemia, inappetence, oedema, icterus, hepatomegaly, splenomegaly, and death in some cases (Onyiche et al. 2019). Dromedary camels infected with B. caballi and T. equi, including camels in this study, however, have been reported to be apparently healthy or have no clinical signs usually associated with piroplasmosis (Qablan et al. 2012). Furthermore, dromedaries mainly free-roam in arid and semi-arid areas where other livestock, including horses, cannot resist (Qablan et al. 2012), supporting the hypothesis that infection of camels with equid piroplasms could circulate in camel populations without frequent involvement of the equine hosts. This hypothesis is further supported by the transovarial and transstadial transmission of B. caballi and T. equi in Ixodidae ticks (De Waal and Potgieter 1987; De Waal 1990; Scoles and Ueti 2015; Nadal et al. 2022; Ravindran et al., 2023). (Nadal et al. 2022; Ravindran et al. 2023) Hence, the molecular detection of equine piroplasms in ticks from camels suggests inclusion of camels in epidemiological surveillance programs for piroplasmoses, especially in regions like Central Asia.
Prior to this study, in Uzbekistan, some faunistic researches were conducted on ticks infesting camels (Kuklina, 1976; Rasulov 2007). However, no data were provided on how many of these tick species are found on camels. Analysis of literature data shows that ticks of the genus Hyalomma are the most common species infesting camels worldwide, many of which can serve as vectors for piroplasms (El Kady 1998; Elghali and Hassan 2009; Fard et al. 2012; Shiri et al. 2024). In Kazakhstan, B. caballi was detected in Dermacentor marginatus (Chunli et al. 2021), while in the present study, it was identified only in Hyalomma sp. It is worth noting that the detection of a pathogen DNA in ticks does not confirm the host’s infection (Krupa et al. 2024). To address this, future studies should consider tick dissection and examination of salivary glands to distinguish between DNA carriage resulting from bloodmeal and active infection, correlated with blood screening of the same animals (Cabezas-Cruz et al. 2018). A possible explanation for the negative molecular results regarding Anaplasma and Rickettsia spp. could be the inherently low infection rates of these bacteria in camel-associated ticks in the studied regions, combined with the limited number of tested pools. Moreover, the pooling strategy and the use of conventional PCR may have reduced the sensitivity for detecting low-level or mixed infections. It is also possible that the local environmental conditions, such as ecological and climatic, and the predominance of Hyalomma spp. more commonly associated with piroplasmids than with Anaplasma or Rickettsia has contributed to the absence of positive findings. The absence of Anaplasma spp., Theileria spp., and Rickettsia spp. DNA in the tested ticks might either reflect an absence in Uzbekistan or might be due to the sample size and pooling strategy.
Considering the previous reports of Anaplasma spp. in camels and ticks in other regions (Selmi et al. 2019; Alanazi et al. 2020), ongoing monitoring remains important.
The present study had several limitations that should be mentioned. First, no pathogen testing was conducted on the blood of the same host animal. Such a thing would have made possible the assessment of pathogen transmission and could have correlated molecular findings in ticks with systemic infections. Secondly, the use of conventional PCR might have led to an underestimation of mixed piroplasmid infections, as its sensitivity is limited compared to more advanced techniques (Criado-Fornelio et al. 2003). Third, the amplification of a single target gene for each of the pathogens may have slightly reduced the reliability of detection results. Additionally, a proportion of collected ticks could only be identified to the genus level due to poor preservation, which might have restricted the precise ecological interpretation and showed imprecise definitive conclusions regarding species-specific vector competence and pathogen–vector associations.
Conclusion
This study, involving a large sample of 290 investigated camels and nearly 1000 associated ticks, provides the first molecular confirmation of B. caballi in Hyalomma spp. ticks collected from camels in Uzbekistan are expanding the current knowledge on tick-borne pathogens in Central Asia, and opening doors for further investigations into the epidemiological role of camels in the transmission of B. caballi, a pathogen traditionally associated with equines. None if the pooled tick samples were positive for Anaplasma or Ehrlichia. To elucidate this, parallel screening of equines in the region and comparison of the genetic characteristics of the parasites detected in two hosts are essential. Considering the co-grazing system is common in Central Asia, these results suggest that camels should be included in the regional screening programs for tick-borne pathogens.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1 (XLSX 2.65 MB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Al-Obaidi QT, Hasan SD, Alsaad KM (2021) Clinical, haematological and blood biochemical parameters in Arabian one-humped camels (Camelus dromedarius) with Babesia Caballi infection. Bulgarian J Veterinary Med, 24(3)
- 2Chunli S, Meihua Y, Bin X, Guangyuan L, Yicheng Y, Kairullayev K, Otarbayev B, Wurelihazi H, Sándor H, Yuanzhi W (2021) Tick distribution and detection of Babesia and Theileria species in Eastern and Southern Kazakhstan, Ticks and Tick-borne Diseases, Volume 12, Issue 6. 10.1016/j.ttbdis.2021.101817
- 3FAOSTAT., (2025) Food and Agriculture Organization of the United Nations Statistics Division. http://www.fao.org/faost
- 4Hussein JJ, Ghazi YA, Rasha MO (2015) Conventional and molecular detection of Babesia Caballi and Theileria equi parasites in infected camels in South of Iraq. Bas J Vet Res Vol. 14, 2.
- 5Kunisov BM, Maksudov AA, Safarov AA (2024 a) Cephalopina titillator (Clark 1797) in camels of Uzbekistan. Actual problems of treatment and prevention of diseases of young animals. Proceedings of the International scientific and practical conference dedicated to the 100th anniversary of the educational institution Vitebsk Order of the Badge of Honor State Academy of Veterinary Medicine, Vitebsk, November 4–6, P. 210–212
- 6Saparov KA (2006) Ecological and biological features of camel helminths - Сamelus dromedarius Linnaeus, 1758 in the conditions of Uzbekistan. Abstract. Ph.D Thesis, Tashkent [In Russian and Uzbek]
- 7Sultanov MA, Azimov DA, Gekhtin V, Muminov PA (1975) Helminths of camels (Camelus dromedarius) helminths of domestic mammals in Uzbekistan. Publ., pp 80–81. [In Russian]