Endogenous Multilayer Control of Cambial Stem Cells and Its Consequences for Wood Formation
Yun-Jing Bao, Fang-Jing Fan, Ying-Gao Liu, Fu-Yuan Zhu

TL;DR
This paper reviews how complex biological networks control stem cells in wood formation, offering insights into improving wood production.
Contribution
The paper synthesizes current knowledge on multilayer regulation of cambial stem cells and its impact on wood formation.
Findings
Cambial stem cell activity is regulated by peptides, transcription factors, and phytohormones.
Chromatin state, protein stability, and non-coding RNAs add regulatory complexity.
Single-cell and spatial transcriptomics help predict wood formation dynamics.
Abstract
The vascular cambium serves as the fundamental meristem for wood formation. It determines wood biomass and structural properties by balancing self-renewal with the bidirectional production of xylem and phloem. This process is controlled by a complex network of peptides, transcription factors, and phytohormones. These regulatory networks coordinate cambial stem cell activity, balancing cell division and differentiation. Additionally, layers of regulation such as chromatin state, protein stability, and non-coding RNAs add significant complexity to these networks. Emerging single-cell and spatial transcriptomics, together with quantitative modeling, now resolve cambial heterogeneity, predicting the dynamic characteristics of wood formation. This review synthesizes current knowledge of cambial regulation, highlighting how feedback loops, spatial gradients, and dynamic signaling networks…
Click any figure to enlarge with its caption.
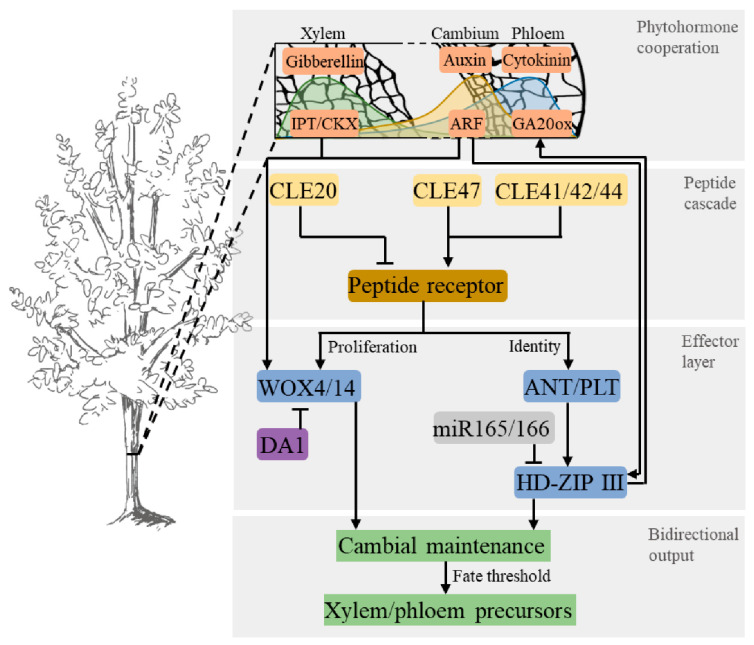 Figure 1
Figure 1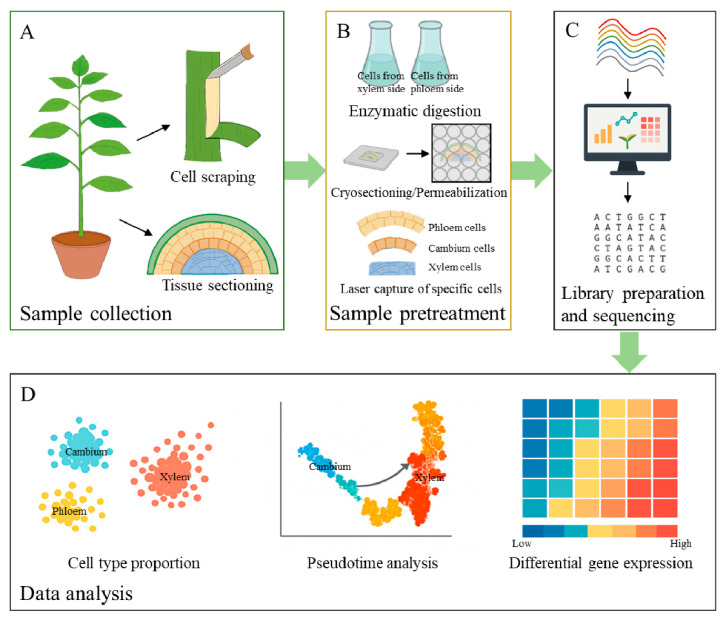 Figure 2
Figure 2- —National Natural Science Foundation of China
- —Biological Breeding-National Science and Technology Major Project
- —National Key Research & Development Program of China
- —Postgraduate Research & Practice Innovation Program of Jiangsu Province
- —Natural Science Foundation of Jiangsu Province
- —National Key Research and Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Polysaccharides and Plant Cell Walls · Plant Gene Expression Analysis
1. Introduction
Wood formation is sustained by secondary growth driven by the vascular cambium, which is essential for plant fitness by providing mechanical support and facilitating long-distance transport. Additionally, wood supports diverse industrial applications, including construction, paper production, and bioenergy [1]. Wood development proceeds through coordinated modules, including the maintenance of cambial cells, differentiation of xylem/phloem precursor cells, cell expansion, secondary cell wall (SCW) thickening, and programmed cell death [2,3]. The rate and spatial precision of cambial regulation determine the upstream supply of differentiating cells, driving radial expansion to produce thicker stems and roots, and establishing efficient vascular transport systems that support sustained growth [4,5,6,7]. This positions the regulation of cambial activity at the foundation of secondary growth.
Plant meristems are classified into apical meristems, which drive longitudinal growth, and lateral meristems, which drive radial growth [8]. The vascular cambium serves as the key tissue for secondary vascular development in woody plants [9]. It is a cylindrical lateral meristem formed by the fascicular cambium and the interfascicular parenchyma cells that close into a spatially continuous cylinder along the stem axis [9,10]. Through coordinated periclinal and anticlinal divisions, cambial cells balance self-renewal with bidirectional output. Specifically, periclinal divisions drive radial growth while anticlinal divisions ensure circumferential continuity as the stem increases in diameter [11]. This precise orientation of cell divisions allows the cambium to maintain its structural integrity while supplying the precursors for wood development [12]. Therefore, endogenous signals are particularly important for localizing and regulating cambial activity in woody species.
Recent research highlights three interacting regulatory layers related to cambial development, including peptide receptor signaling [13], transcriptional control [14], and phytohormone integration [15]. These layers work together to regulate cambial activity, ensuring a balance between stem cell proliferation, differentiation, and maintenance, which ultimately determines the output of xylem and phloem. Unlike primary meristems, which are primarily governed by intrinsic developmental programs, cambial stem cells are uniquely regulated by the integration of dynamic feedback loops, precise temporal control, and environmental cues [16,17]. This complex regulatory network facilitates secondary growth and enables the continuous and adaptive wood formation throughout the lifespan of a tree. We propose a conceptual model that illustrates how these regulatory pathways jointly define the cambial stem cell zone and determine bidirectional wood output (Figure 1). While these pathways are widely conserved, their interaction within the cambium is uniquely tailored to meet the specific requirements of secondary growth. Integrating these layers provides insights into how cambial stem cells maintain their identity, which informs predictive breeding and the rational engineering of biomass and material properties.
In recent years, model species such as Arabidopsis have provided significant contributions to understanding the molecular mechanisms underlying cambial activity, particularly during primary growth. These models have been instrumental in defining the signaling networks and phytohormonal pathways that control stem cell fate, division, and differentiation within meristems. Current research has highlighted conserved molecular frameworks shared between herbaceous and woody plants, such as the core CLE-WOX signaling modules [18,19]. However, there remain limitations in applying these findings directly to woody plants due to the complexities of secondary growth, the prolonged lifespan of tree species, and various environmental influences [20,21,22]. While many conserved mechanisms have been identified, particularly in juvenile plants like Populus, these findings are not fully applicable to mature woody plants as structural characteristics differ [23]. Specifically, mature woody plants possess more expansive secondary vascular systems and higher degrees of lignification maintained over decades [24]. Furthermore, while core components like CLE peptides and WOX transcription factors are evolutionarily conserved, their regulatory mechanisms and functional integration often diverge [25]. These functional adaptations highlight the necessity of integrating findings from both model species and mature woody plants to develop a more comprehensive understanding of cambial activity and wood formation.
In this review, we synthesize evidence on cambial regulation with an emphasis on woody plants, using Arabidopsis as a reference. We discuss peptide receptor signaling, transcriptional modules, and phytohormone integration. Furthermore, we explore how emerging single-cell and spatial transcriptomic approaches resolve cambial regulation at cellular resolution, which might guide future research into the vascular cambium.
2. Maintenance of Cambial Stem Cell Activity
2.1. Cross-Layer Peptide Signaling
A conserved peptide receptor system positions and sustains cambial activity through non-cell-autonomous communication from the phloem to the procambium [26]. Phloem-expressed CLAVATA3/EMBRYO SURROUNDING REGION 41/42/44 (CLE41/42/44) genes encode proteins that are processed into TRACHEARY ELEMENT DIFFERENTIATION INHIBITORY FACTOR (TDIF) peptides. These peptides are secreted into the apoplastic space and transported across tissue layers to the procambium through apoplastic diffusion [13,27]. This apoplastic transport is facilitated by the specific cellular anatomy of the cambial zone. During active growth phases, the absence or limited presence of plasmodesmata in the cambial initial zone prevents symplastic movement. Consequently, TDIF peptides must diffuse through the extracellular space to reach and bind with receptors on their target cells [18,28]. Loss of CLE41 reduces the number of cambial cell layers while leaving sensitivity to exogenous TDIF intact, indicating that CLE41 constitutes the principal endogenous TDIF source, with partial redundancy from CLE44 [13]. Notably, additional tissue-specific inputs diversify this axis. Xylem-derived PtrCLE20 acts on the cambium through long-distance transport, inhibiting cell division and forming a negative feedback regulation [29]. In contrast, PttCLE47 is expressed within the cambium and promotes cambial proliferation. Consistently, RNAi-mediated silencing of PttCLE47 results in fewer cambial cell layers and a significant narrowing of the secondary xylem region, with little effect on the phloem [25]. Furthermore, overexpression of PttCLE41 significantly increases the biomass of wood [29,30], underscoring the dosage-dependent control of radial growth. Together, the diverse tissue origins and action mode of CLE ligands enrich the regulatory network and provide distinct strategies for defining the cambial region.
TDIF peptides are recognized by the leucine-rich repeat receptor-like kinase PHLOEM INTERCALATED WITH XYLEM (PXY) on the plasma membrane of cambial cells, triggering a cascade of intracellular events that maintain the cambial stem cell identity [13]. Mutants of pxy fail to respond to TDIF and develop discontinuities in the cambial cylinder during secondary growth [31], demonstrating the essential role of this receptor in maintaining circumferential continuity.
Downstream of PXY, WUSCHEL HOMEOBOX RELATED 4 (WOX4) is a key regulator of cambial division and acts partly redundantly with WOX14. Notably, wox4/wox14 mutants retain residual cambial activity [14]. The rapid upregulation of WOX4 by TDIF in a PXY-dependent manner highlights the integration of peptide signaling with transcriptional regulation in maintaining cambial activity. Furthermore, wox4 mutants show reduced procambial proliferation but remain sensitive to the TDIF-mediated inhibition of xylem differentiation. Since overexpression of WOX4 is insufficient to replicate the proliferation induced by TDIF in the hypocotyl, additional WOX4-independent pathways are likely required [13]. This points to a more complex network in which genetic interactions reveal parallel determinants of cambial activity. For example, cambial activity is nearly abolished in the double mutants of wox4 and KNOTTED-like from Arabidopsis thaliana 1 (knat1), revealing KNAT1 alongside WOX4 as a major determinant of division [32]. In parallel, an identity branch composed of CAMBIUM-EXPRESSED AINTEGUMENTA-LIKE (CAIL) transcription factors operates downstream of TDIF-PXY. Consistent with this, mutants of PLETHORA 3/5/7 (PLT3/5/7) exhibit reduced cambial cell numbers and compromised secondary growth [33].
The CLE41-PXY-WOX pathway exhibits dynamic activity patterns, orchestrating cambial activity by integrating internal developmental stages with external environmental cues [34]. In woody species, the sequential phases of cambial cell development, from initial activation to peak proliferation and eventual maturation, are fundamentally synchronized with the seasonal cycle. For instance, in Betula pendula, the expression of BpCLE41/44a peaks during the initial proliferation phase, which coincides with the early spring reactivation of cambial initials. As development progresses, BpPXY expression reaches its maximum during the mid-differentiation phase, supporting the intensive mid-season production of secondary xylem. Additionally, while BpWOX4 typically maintains a high correlation with the rate of cell division, its expression can decouple in cases of abnormal xylogenesis, leading to premature cessation of activity during late-season maturation phases [35]. This intrinsic developmental clock is further modulated by broader seasonal variations across the annual growth cycle. Similarly, in Populus, WOX4-like genes regulate the rate of cambial division in response to these seasonal shifts, whose overexpression prolongs active growth phases, while mutants show phased declines in proliferation as the tree prepares for winter dormancy [34]. Computational models of the poplar cambium further synthesize these layers, illustrating how CLE-PXY signaling dynamically maintains homeostasis. Early-phase peptide release triggers the onset of spring cambial proliferation, while mid-to-late summer feedback loops prevent over-differentiation, thus ensuring sustained and balanced wood formation throughout the annual growth cycles [36].
Together, the cambium integrates peptides originating from distinct vascular tissues to promote division while preventing over-proliferation, thereby defining narrow, continuous stem cell domains and coordinating balanced and bidirectional production of xylem and phloem [35]. The TDIF peptides, via PXY signaling, combine with auxin gradients and transcription factors such as WOX4 and HD-ZIP III to establish a unique feedback system that ensures balance between proliferation and differentiation for sustained wood formation.
2.2. Cell Cycle Drivers of Cambial Proliferation
Cell cycle regulators are important drivers of cambial activity. The CYCLIN D3 (CYCD3) module is key to accelerating periclinal divisions within the cambial zone, thereby directly increasing the production of vascular precursor cells [37]. Recent studies demonstrate that CYCD3;1 acts as a critical downstream effector of cytokinin (CK) signaling to promote secondary growth, providing a conserved mechanism for regulating the rate of cambial cell proliferation [38]. In woody species, this module has expanded and diversified to meet the complex demands of perennial growth. In Populus trichocarpa, for instance, PtoCYCD3;1 maintains its role in promoting radial growth via CK signaling, while PtoCYCD3;3 has evolved additional functions in branching [21], illustrating how conserved G1 drivers are recruited for diverse woody architectural requirements.
These functional specializations are further refined by a sophisticated regulatory network, where specific CYCD and CYCLIN-DEPENDENT KINASE (CDK) family members exhibit distinct interaction preferences at the genome level to fine-tune vascular development in poplar [39]. CDKs are highly expressed in vascular domains including the cambium [40]. Furthermore, CDKA;1 not only regulates cell proliferation but also couples division control to SCW biosynthesis by interacting with enzymes like UDP-glucose 6-dehydrogenase (UGDH). This interaction ensures that the supply of SCW precursors is precisely synchronized with the rate of cell production [41,42]. This integrated framework demonstrates how vascular-specific signals and hormonal cues modulate cyclin-CDK activity to coordinate tissue proliferation with the structural development of woody plants.
2.3. Phytohormonal Control of Cambial Activity
Phytohormones play a pivotal role in regulating cambial activity, controlling key processes such as stem cell division, fate determination, and differentiation during the secondary growth. The main phytohormones involved include auxin, CK, gibberellin (GA), ethylene (ETH), and brassinosteroid (BR), each contributing in a specific manner to the maintenance and growth of the cambium.
Auxin is a key phytohormone that transmits signals between cells in meristematic tissue, regulating various developmental behaviors [15,43]. The concentration of auxin is elevated in the cambium relative to surrounding tissues, forming a gradient that is essential for defining the organizer identity [44]. Disrupting the apical sources of auxin suppresses cambial activity and secondary xylem formation [3], underscoring the dependence of auxin gradients in the vascular pattern [45]. As the cambium grows, the position of the auxin peaks shift. PIN-FORMED (PIN) family transporters, which are highly expressed around the cambium, play a key role in setting auxin distribution [46]. Specifically, PIN5 regulates intracellular levels of free indole-3-acetic acid (IAA) [46,47], controlling the availability of auxin in the cambium. Overexpression of PdePIN5b promotes the transport of auxin into the endoplasmic reticulum, reducing the concentration of IAA in the cytoplasm and thereby significantly suppressing cambial proliferation [47]. Auxin interacts with TDIF-PXY-WOX signaling to regulate the polarity of the stem cells [48]. For example, auxin upregulates PXY expression via ARF5, while TDIF-PXY signaling negatively regulates auxin response, creating a feedback loop that ensures proper cambial activity and prevents over-proliferation [15,36]. Furthermore, auxin regulates WOX4 expression, thereby coordinating auxin fluxes and defining the stem cell organizer [14,48,49]. These dynamic gradients help maintain a narrow stem cell domain while ensuring continuous bidirectional output. Auxin signaling is also involved in regulating HD-ZIP III, as PmHB1 is regulated by auxin homeostasis to affect meristem activity in Prunus mume [50]. Specific ARF members, such as ARF6 and ARF8, play a more targeted role in promoting xylem formation over phloem [51], further indicating the complexity and specificity of auxin signaling components in regulating cambial activity.
CK also directly influences cambial activity by regulating cell division and radial growth. The CK biosynthesis gene Isopentenyl transferase (IPT) and the catabolic gene Cytokinin oxidase 2 (CKX2) modulate cambial activity in predictable directions [52,53]. For instance, mutants of ipt show a loss of cambial cells, while overexpression of PtIPT7 significantly increases the number of vascular cambial cells [52,54]. Conversely, overexpression of CKX2 inhibits cambial activity and restricts radial growth [52], emphasizing the essential role of CK homeostasis in cambial activity. Besides that, CK receptors such as Histidine Kinase 3 (HK3) and Cytokinin Response 1 (CRE1) are enriched in dividing cambial cells, further supporting their role in regulating cambial proliferation [52]. Additionally, CK induces the expression of PHLOEM EARLY DOF (PEAR), which promotes procambium proliferation and patterning [55]. Downstream of cytokinin, a set of LATERAL BOUNDARY DOMAIN (LBD) transcription factors act to initiate secondary growth [56,57]. For instance, LDB11 forms a feedback loop with reactive oxygen species (ROS) to regulate cambial proliferation, where ROS over-accumulation suppresses LBD11 transcription, thereby ensuring balanced cambial activity [58]. This mechanism highlights the interplay between cytokinin and ROS signaling.
Auxin and cytokinin do not operate in isolation but instead interact synergistically to regulate cambial activity. A high concentration of auxin promotes xylem differentiation, while CK enhances stem cell division toward the phloem, together shaping the differentiation pattern of the cambium [59]. The interaction is underpinned by shared regulatory mechanisms, such as the Monopteros/Auxin response factor 5 (MP/ARF5)-Lonesome highway (LHW)-Target of monopteros 5 (TMO5) pathway, which directly drives cambial establishment and division by activating PIN1 to establish a positive feedback loop that stabilizes polar auxin transport [12,60,61,62]. A parallel mechanism involves LHW-TMO5 regulating CK, which is critical for cell proliferation and procambium patterning [63]. This interaction highlights the complementary roles of auxin and CK in regulating cambial proliferation and differentiation. Auxin signaling via ARFs controls cell cycle genes such as CYCD3 and regulates lateral root initiation and cell fate changes through AUX/IAA partners [64], linking primary and secondary growth. CK further promotes cambial activity by inducing CYCD3 expression [65], reinforcing the connection between phytohormone input and cell cycle entry. Interestingly, the role of ARF factors exhibits developmental context dependency. During primary vascular development, ARF5 activates LONELY GUY 3/4 (LOG3/4) to boost CK biosynthesis and promote radial division [63,66]. In contrast, during secondary growth, ARF5 suppresses cambial activity, whereas ARF3 and ARF4 act positively [15]. This functional difference highlights how the same transcription factor can be reprogrammed to perform distinct roles across different developmental stages and tissue environments.
GA promotes secondary growth by increasing the accumulation of dividing cambial cells [67]. GA regulates xylem formation through the degradation of the GA signaling repressors GIBBERELLIN INSENSITIVE (GAI) and REPRESSOR OF GA1-3 (RGA), promoting the differentiation of xylem vessels [51]. The promoting effect depends on PIN1-mediated polar auxin transport and ARF signaling, as its effect is inhibited in pin1 and arf7/arf19 mutants [44]. This demonstrates the interdependence of GA and auxin signaling. Additionally, the interaction between GA and auxin is facilitated by the direct activation of WOX4 by PtoARF7, forming a ternary complex with DELLA and AUX/IAA to regulate cambial proliferation [68]. Besides this, the HD-ZIP III transcription factor PHB, expressed in the vascular tissues, reduces GA activity by increasing GA2ox2 expression, thereby triggering endodermal accumulation of CYCD6;1 that drives the formative asymmetric divisions [69].
Similarly, ETH enhances cambial activity through canonical signaling components and cross-talk with downstream transcriptional regulator [15,70]. Furthermore, BR increases cambial proliferation, whereas BR biosynthesis inhibition dampens secondary vascular differentiation [71], placing BR as a tunable growth amplifier in woody stems.
The phytohormonal regulation of cambial activity is dynamically modulated depending on the phase of cambial growth, integrating seasonal and environmental cues to control activation, proliferation, and senescence in woody plants. For instance, phase-dependent phytohormonal shifts are particularly evident in species like Populus, where phytohormones such as abscisic acid (ABA), BR, jasmonic acid (JA), and GA are upregulated during active growth phases to balance proliferation and differentiation. The expression of genes related to these phytohormones peaks mid-season, coinciding with maximal xylem production, and declines in later phases to prevent overgrowth and ensure proper tissue maturation [72]. These phytohormonal gradients interact with the TDIF-PXY-WOX pathway, maintaining cambial homeostasis and adapting to environmental cues such as temperature and photoperiod [73]. In Populus, Pag Jasmonate ZIM-domain 5 (PagJAZ5), acting as a core repressor of JA signaling, coordinates CK signaling with TYPE-A RESPONSE REGULATORS (ARR) to regulate phase-specific cambial activity. During the early proliferation phase, PagJAZ5 enhances CK sensitivity to support rapid division, while in mid-to-late phases, it promotes differentiation by repressing PagARR, ensuring a balanced transition between cell division and maturation and preventing premature senescence [74].
Together, these phytohormonal signals create a multilayered regulatory network that coordinates the cambial response to positional cues, proliferation signals, and growth-promoting factors. This integration is unique to cambial stem cells due to their unique role in sustaining radial growth and generating both xylem and phloem tissues in a continuous manner. Phytohormones directly regulate cambial cell division and differentiation through spatial gradients while interacting with downstream transcription factors. The crosstalk adapts cambial transitions from reactivation through peak growth to dormancy, aligning with annual cycles for both longevity and environmental resilience, thereby maintaining tissue organization and aligning the balance between self-renewal and bidirectional output required for sustained secondary growth. The temporal and spatial regulation of these pathways in response to environmental cues further sets cambial stem cells apart from other meristems in the plant.
2.4. Fate Determination and Differentiation
Cambial fate thresholds emerge from the interplay of spatial hormone gradients, feedback stabilized auxin transport, and gatekeeper transcription factors. Phytohormones are distributed across the radial axis, with GA enriched in developing xylem, auxin peaking in the cambium, and CK accumulating toward the phloem, thereby imparting positional information to dividing and differentiating cells [75]. Through periclinal divisions, cambial stem cells undergo radial expansion and produce daughter cells whose fates are determined by their spatial positioning relative to an organizing center [76]. This center activates ARF5 and promotes HD-ZIP III, inducing proliferation in adjacent stem cells and determining the direction of cambial differentiation [77,78]. For instance, PtrHB4 regulates the formation of the interfascicular cambium, with Ptrhb4 mutants exhibiting a discontinuous cambium arrangement resembling a protovascular bundle pattern, which negatively affects xylem differentiation [10,79]. Similarly, PtrHB7 is highly expressed in the cambium and modulates the balance between xylem and phloem outputs in a dosage-dependent manner. Overexpression of PtrHB7 promotes excessive xylem formation at the expense of phloem, while the mutant displays the completely opposite phenotype [80]. In addition, KNOTTED-LIKE HOMEOBOX (KNOX) factors provide a complementary regulatory layer. While AtKNOX1 maintains cambial stem cell potential in Arabidopsis [81], its poplar homolog PtKNOX1 has acquired an additional function in promoting xylem fiber differentiation [20], illustrating functional innovation within woody species.
2.5. Cross-Meristem Commonalities
Comparative genetics across meristems reveals a conserved regulatory mechanism in which non-cell-autonomous peptide cues define the stem cell domain, and organizer transcription factors maintain stem cell identity. Both primary meristems and the cambium rely on similar signaling modules that consist of CLE family signaling. In the shoot apical meristem (SAM), the CLE peptide CLAVATA3 (CLV3) restricts the organizer factor WUSCHEL (WUS). In turn, WUS promotes CLV3 expression, establishing a dosage-sensitive feedback loop that controls the size of the stem cell domain [82,83]. A similar regulatory mechanism operates in the root apical meristem (RAM). CLE40 signals through the receptors CLV1 and ACT DOMAIN CONTAINING PROTEIN 4 (ACR4) to position the quiescent center and limit stem cell accumulation. Meanwhile, WOX5 functions as a root organizer analogous to WUS [17,84]. In contrast, the cambium exhibits more complex peptide receptor signaling to maintain stem cell identity [85]. The CLE41 peptide promotes cambial cell division and xylem formation [86], while CLE40 and CLV3 inhibit stem cell division [17,82], highlighting the functional diversity of CLE signaling across different tissues.
WUSCHEL family transcription factors play a pivotal role in defining stem cell identity across these meristems. While these transcription factors share functional similarities in stem cell regulation, they operate in distinct contexts [85]. In the SAM, WUS is regulated by low auxin levels, and its role is primarily to promote stem cell proliferation and maintenance [85]. Conversely, in the cambium, WOX4 is induced by high auxin concentrations, where it plays a critical role in maintaining cambial stem cell activity and supporting radial growth [48]. This differential auxin regulation underscores a fundamental distinction in cambial regulation, where WOX4 is not only crucial for stem cell maintenance but also for coordinating continuous wood formation and the bidirectional production of xylem and phloem.
While both primary and secondary meristems employ conserved CLE peptide and WOX transcription factors for stem cell regulation, their functions diverge to serve different developmental objectives. In primary meristems, the CLV3-CLV1-WUS pathway operates through negative feedback loops, where CLE peptides inhibit WUS expression to maintain a stable stem cell reservoir [82,87]. In contrast, the TDIF-PXY-WOX4 pathway works as a positive regulatory mechanism in the vascular cambium to promote sustained radial expansion [34,78]. The CLE peptides stimulate WOX4 expression to drive stem cell division and support sustained radial growth [88]. This integration of peptide signals with auxin and other cues allows the cambium to adapt its activity over time, facilitating the continuous and bidirectional production of xylem and phloem throughout the life of the plant.
AIL/PLT transcription factors supply a further insight into the regulatory mechanisms shared across meristems. In roots, a graded distribution of PLT, established by ARF-dependent transcription, maintains high doses of stem cels and promotes transit amplifying division at lower doses, thereby repressing differentiation. The PLT gradient is further constrained by peptide growth factors and their receptors, reflecting similar peptide signaling axes found in the cambium [89]. In contrast, AIL/PLT functions similarly to drive proliferation and delay differentiation but operate without a global protein gradient, with ANT being dominant and other PLTs acting unequally across space and time [89]. Moreover, ANT is enriched in cambial stem cells, mutants reduce radial growth, and expressing ANT or PLT5 from the PXY promoter restores cambial activity in pxy mutants, establishing ANT/PLT as a regulator of cambial activity in the lateral meristem [33]. These findings reveal the function of PLT in maintaining stem cell identity and differentiation across both primary and secondary meristems. A similar pattern is observed in the PHYTOCHROME-INTERACTING FACTOR (PIF) family. In primary growth, PIF3/4/5 act as negative regulators of vascular patterning and xylem differentiation for auxin-mediated cell elongation but suppressing SCW formation [90,91]. In contrast, PtoPIF3.1/3.2 positively regulate stem elongation and secondary xylem development through the conserved PIF-auxin module, while extending to modulate cambial markers like PtoWOX4 and PtoHB7/8 [22,92]. This functional shift highlights how gene family expansion in woody perennials adapts conserved pathways for sustained radial growth and wood formation.
Together, the conserved mechanisms across the SAM, RAM, and vascular cambium highlight both shared principles and unique adaptations in meristem regulation. CLE peptides, auxin signaling, and WUSCHEL family transcription factors are central to regulating stem cell identity and maintenance across these tissues. Additionally, the functional diversity detected in woody perennials, arising from the expansion and functional differentiation of specific gene families, has enabled the evolution of secondary growth (Table 1). These findings provide critical insights into the evolutionary adaptations that have allowed for the development of complex woody tissues and sustained perennial growth.
2.6. Maintenance of Cambial Stem Cells in Mature Trees
As trees age, the activity of cambial stem cells generally declines, typically resulting in reduced cell division rates and limited wood formation [100]. However, the regulatory mechanisms governing cambial aging exhibit significant interspecific variation, particularly in long-lived species where endogenous signals act to delay senescence. For instance, maintaining sufficient levels of IAA is crucial for sustaining cambial longevity [101]. In Populus euphratica, the upregulation of ABA, BR, and GA further assists in balancing cell proliferation with maturation [102]. Conversely, altered expressions of cell cycle regulators, such as CYCD3, have been shown to accelerate cambial aging in Eucalyptus [100]. Beyond molecular signaling, structural and functional adaptations also support sustained activity; for example, bark thickening provides thermal insulation to protect the cambium [103], while physiological shifts toward wider and longer vessels enhance hydraulic efficiency and cavitation resistance in aging stems [104].
While the ancestral bifacial cambium remains a conserved feature across most woody seed plants for the continuous production of secondary xylem and phloem, its operational longevity varies markedly between lineages. In long-lived gymnosperms such as Ginkgo biloba, the cambium exhibits remarkable mitotic stability that can persist for centuries. This stability is maintained through a complex interplay of reduced IAA, increased ABA, and epigenetic modulations including DNA methylation of DAL1 and miR166-mediated signaling, which collectively prevent a precipitous decline in cell division [105]. Consequently, although the seasonal duration of xylem formation may shorten as these trees mature, their fundamental capacity for wood formation remains intact [24].
In contrast, angiosperms are characterized by more dynamic aging mechanisms and a broader diversity of secondary growth patterns [106]. While angiosperms like Populus euphratica maintain cambial homeostasis by coordinating cell expansion genes such as XYLOGLUCAN ENDOTRANSGLUCOSYLASE/HYDROLASES (XTH) and lignin biosynthesis pathways such as TRANS-CINNAMATE 4-MONOOXYGENASE (C4H) [102], other lineages have evolved anomalous secondary growth to adapt to specific ecological niches. For example, the development of polycyclic eusteles and successive cambia in Commicarpus scandens [107], along with the isolated vascular cylinders in Manekia [108], enable rapid biomass accumulation and architectural flexibility. These diverse strategies indicate that angiosperms utilize high degrees of transcriptional and miRNA-mediated plasticity to modulate cambial activity and longevity throughout their life cycles [102,109,110].
3. Xylem Development Under Cambial Regulation
3.1. Advances in Differentiation of Xylem
Following periclinal division, cambial daughters enter the xylem lineage through a program set by transcription factors and tuned by phytohormones [111]. For example, Pag UNFERTILIZED EMBRYO SAC 12 (PagUNE12) promotes differentiation toward xylem and supports secondary vascular development [112]. Conversely, PagMYB31, expressed in the cambium and the early xylem, functions as a negative regulator, suppressing xylem differentiation [98]. PttCLE47 mutants exhibit secondary xylem width and xylem cell numbers while leaving phloem largely unchanged, suggesting impaired cambial proliferation and export of cambial progeny [25]. Meanwhile, the HD-ZIP III family plays a pivotal role in cambium establishment and differentiation direction, with Ptr Homeobox 4 (PtrHB4) ensuring cambial ring continuity to support normal xylem formation [10,79]. PtrHB7 and PtrHB8 regulate the differentiation of cambial cells into secondary xylem, and their overexpression promotes the differentiation of cambial cells towards xylem [10,113]. Furthermore, CAILs regulate cambial activity and determine xylem/phloem differentiation. For example, PLT5 rapidly inhibits the expression of xylem and phloem differentiation markers such as CASCULAR-RELATED NAC-DOMAIN 6 (VND6) and PEAR1, while its mutants lead to the reduction in cambial cells and arresting of secondary growth [33].
Phytohormone pathways intersect with these transcriptional modules to set early differentiation thresholds. Overexpression of IPT simultaneously increases the contents of CK and IAA, which further promotes xylem cell output [114]. GA signaling activates IAA signaling factors such as ARF5/ARF7/ARF19 that adjust the xylem/phloem ratio [44]. In addition, meristem identity genes enrich cell differentiation mechanisms associated with phytohormone signaling. For instance, PtoIAA9 interacts with PtoARF5 to inhibit the expression of PtoHB7/8, thereby inhibiting the differentiation of cambium cells into xylem [115]. Together, this coordination of transcription factors and phytohormonal signals ensures the proper differentiation and expansion of xylem cells, setting the stage for wood formation.
3.2. Xylem Precursor Cell Expansion
As cambial stem cells enter the xylem lineage, they progress into an elongation phase whose control is already set by earlier division and fate decisions [116,117]. Phytohormones play a central role in coordinating this transition. Auxin effector SMALL AUXIN UP RNA (SAUR) significantly promotes cell growth, highlighting a direct role for auxin in xylem expansion [118]. Similarly, GA promotes xylem precursor cell elongation, increasing vessel diameter and fiber number. This effect is consistent with phenotypes of Gibberellin 20-oxidase 1 (GA20ox1) overexpression and DELLA degradation [119,120]. ETH further enhances xylem expansion through increasing the activity of its biosynthesis enzyme Pt 1-AMINOCYCLOPROPANE-1-CARBOXYLIC ACID OXIDASE GENE 1 (PtACO1) and activating downstream targets such as XTH and EXPANSINS (EXP) through the Pt ETHYLENE INSENSITIVE 3-LIKE (PtEIN3) signaling pathway, leading to larger vessel and tracheid diameters [121,122]. In addition, phytohormones participate in the expansion of xylem cells by regulating the expression of cell wall-related genes and metabolic pathways to construct a multi-dimensional regulatory network. For example, ARF5 modulates the expression of cell wall remodeling genes EXPA1, linking identity signaling to expansion capacity [123]. GA and auxin signals also influenced cambial cell proliferation through WOX4, ensuring a balanced shift between stem cell activity and differentiation [67,68].
Expansion then depends on targeted remodeling of the primary wall. Several enzymes related to the cell wall contribute to this process, with PttEXPA1 specifically promoting radial expansion in xylem, and mutants resulting in radial growth defects [123]. OsEXPA8 also controls endodermal anisotropy and thus influences the growth of organs [124]. Additionally, XTH22 catalyzes xyloglucan remodeling [125]. PECTIN METHYLESTERASES (PME) further modifies wall mechanics. For example, PME5 restricts elongation, whereas the loss of PME35 reduces wall strength [126,127]. In parallel, REDUCED WALL ACETYLATION 2 (RWA2)-mediated xylan acetylation fine-tuned wall chemistry, with rwa2 mutants impairing fiber development and limiting xylem expansion [128].
In summary, differentiation and expansion of xylem cells arise as downstream outcomes of signals initiated in the cambium, with phytohormone inputs coupling to wall-modifying enzymes to convert early positional and identity cues into sustained cell enlargement and wood growth. This underscores the central role of cambial cell activity throughout the entire process of wood formation.
3.3. Secondary Cell Wall Deposition
SCW thickening plays a crucial role in translating developmental decisions into the mechanical and hydraulic properties of wood. In perennial woody plants, continuous lignin accumulation strengthens tissue and supports long-distance water transport, which together maintain growth and stability [129,130,131]. Lignin is essential for maintaining cell wall rigidity, while cellulose and hemicellulose provide tensile strength and flexibility. Specifically, cellulose microfibrils oriented in a layered pattern within the SCW impact the mechanical properties of wood [132].
Upstream cues that set cambial fate and division rates also shape SCW programs. For instance, the peptide signal PtrCLE20 produced by xylem inhibits cambial activity in aspen, thereby indirectly affecting the number of cells that entered the wall thickening program [29]. This peptide signal restricts the initiation of SCW biosynthesis by limiting the pool of cells that can differentiate into SCW-forming xylem cells.
Several transcription factors that influence cell fate also impact SCW deposition. For example, PagUNE12 indirectly impacts SCW thickening by influencing the timing of cambial differentiation [112]. Furthermore, loss of PtrHB4 disrupts cambial ring continuity and compromises SCW deposition [10]. Additionally, the Eg HISTONE VARIANT 1.3 (EgH1.3) and EgMYB1 prevent premature lignification in the cambium and early xylem cells, thereby maintaining a non-lignified identity in mature parenchyma cells [133].
Three primary components of SCW are tightly regulated to ensure the mechanical and hydraulic properties of wood. For instance, PtoMYB31 directly represses lignin biosynthesis genes, including Caffeoyl-CoA-O-methyltransferase (CCoAOMT), Caffeic acid-O-methyltransferase (COMT), and Cinnamoyl alcohol dehydrogenase (CAD), thereby reducing lignin deposition [98]. This regulation impacts mechanical properties by reducing the rigidity and water transport capacity of wood. Furthermore, ARF3 and ARF5 modulate the expression of Cellulose synthase (CesA), which is involved in cellulose biosynthesis during cambial differentiation [134]. Additionally, WOX4 modulates the expression of hemicellulose-related genes, coordinating the deposition of hemicellulose in the secondary wall during cambial differentiation [135].
Therefore, SCW deposition functions as a late-stage developmental program that precedes programmed cell death. It remains tightly coupled to earlier cambial regulation, where upstream signals modulate cambial activity and influence cell fate decisions, propagating forward to determine wall thickness, composition, and ultimately wood formation. These regulatory processes ensure that cellulose, hemicellulose, and lignin are precisely coordinated to optimize the physical properties of wood. The regulatory genes involved in wood formation in woody plants are summarized in Table 2, which highlights key genes responsible for regulating cambial activity, cell differentiation, and secondary wall biosynthesis.
4. Frontiers and Future Directions
4.1. Single-Cell, Spatial Transcriptomics, and Quantitative Modeling
Traditional bulk transcriptomics and histology have faced challenges, including low resolution in capturing cellular heterogeneity, spatial gradients, and dynamic interactions, often masking rare subpopulations or transient states critical for fate decisions [137]. Emerging single-cell and spatial transcriptomic approaches provide high resolution insights into cambial heterogeneity and differentiation trajectories in woody species (Figure 2). These advanced approaches provide new empirical data on spatial gradients and cell subpopulations, which bridge molecular inputs to cambial outputs.
Single-cell RNA sequencing in the SAM vascular differentiation captures low-abundance transcripts in the cell nucleus, revealing differentiation pathways in the xylem and phloem and linking TDIF-PXY-WOX4 signaling to asymmetric fate determination during cambial division, while identifying specific cell subpopulations essential for vascular tissue specialization and wood formation [138]. Similarly, a transcriptional landscape of stems identifies distinct cell clusters and further reveals cellular heterogeneity and lineage progression at single-cell resolution [139]. These approaches have revealed the occurrence and maintenance of cell heterogeneity within the cambium. Complementing cellular resolution, high-spatial-resolution transcriptome profiles have consecutive radial sections across the phloem, cambium, and xylem, providing a positional reference for wood-forming domains, thus identifying key markers and their expression gradients, and further clarifying the regional differences in the activity and differentiation of the cambium [140]. Spatial transcriptome analyses identify two distinct types of meristematic-like cell pools within secondary vascular tissues, thereby directly linking positional information to developmental progression and refining models of cambial organization and differentiation [141]. Furthermore, combining single-cell RNA sequencing and spatial transcriptome analyses localizes cambial sub-domains and expression gradients of key regulators, revealing distinct expression patterns of key factors like WOX4 and HD-ZIP III across different cambial subpopulations [142,143]. This integration has revealed distinct patterns of gene expression that are crucial for regulating cambial proliferation and differentiation, linking cellular behaviors to spatial positioning within the cambial zone. It also demonstrates how signal gradients and transcriptional programs are spatially organized to guide tissue patterning, offering potential solutions for cambial dynamics. Additionally, a stem-differentiating xylem (SDX) protoplast workflow, coupling RNA-seq with ChIP-seq, provides a strategy to detect hierarchical regulatory networks, linking upstream signals like ARF5 to downstream xylem development [144].
Laser capture microdissection (LCM) combined with transcriptomes has been pivotal for further interpreting the information that constitutes the map in specific tissues. LCM-guided analyses show that high CK levels in the phloem promote cambial proliferation, confirming its key role in cambial development [52]. Meanwhile, an auxin maximum centers on the cambium, providing quantitative inputs for fate and division models [59]. Focusing LCM on defined cambial cell layers enables discovery of tissue-restricted gene programs. Notably, 95 cambium-specific transcription factors are identified, which illustrates the precision of LCM in isolating gene expression patterns [78]. The tissue-specific analysis provides a tool for dissecting the complex molecular mechanisms that underlie wood formation, bridging the gap between cellular resolution and tissue-level functional analysis.
Moreover, quantitative modeling provides a mechanistic bridge from measured inputs to predicted cambial behaviors, converting inter-molecular interactions into direct developmental outcomes. Mathematical models integrate the complex interactions between PXY and MP signaling in cambial development, revealing how diverging interactions between these pathways impact stem cell activity and tissue patterning during growth [145]. Additionally, with the detection of more high-resolution data, frameworks that parameterize peptide signals, phytohormone signals, and transcription factor thresholds can predict the width of the cambial stem cell domain and the boundary of division and differentiation, as well as simulate cambial dynamics under various physiological conditions through running perturbations [33].
Integrating single-cell, spatial, and LCM-derived transcriptome with quantitative models improves prediction of cambial dynamics and guides experimental design and trait targeting. These advanced techniques not only map gene expression but also enable reconstruction of regulatory networks and dynamic gradient formation [78,142]. Coupled with mathematical models that simulate feedback loops and diffusion processes, they provide a mechanistic understanding of how gene families expand and diversify to generate phenotypic variation in wood development [98], informing strategies to optimize wood formation.
4.2. Epigenetic Regulation
Chromatin state and protein modifications work together to fine-tune wood formation. In Eucalyptus, a linker histone and TF complex, EgH1.3-EgMYB1, directly represses lignin biosynthetic genes, prevents premature lignification in the cambium and early xylem, and maintains a non-lignified identity in mature parenchyma [133]. Cambium identity is also set by chromatin state. PtrVCS2 modulates histone acetylation at PtrWOX4a, thereby adjusting the cambial layer number and secondary growth [78]. Beyond chromatin, post-translational control limits cambial division. DA1-type ubiquitin-associated proteins reduce WOX4 stability and restrain cambial proliferation, establishing a protein-level brake on stem cell activity [146]. Methodologically, applying chromatin profiling assays such as an assay for transposase-accessible chromatin with high throughput sequencing (ATAC-seq) and cleavage under targets and tagmentation (CUT&Tag) to cambium-enriched tissues should connect chromatin accessibility and histone marks to stem cell potential, identity gating, and lineage bias, thereby complementing transcriptional atlases [144].
4.3. Non-Coding RNAs
Non-coding RNAs add regulatory layers that interface with hormone pathways and transcription factor networks to shape cambial programs. A general splicing factor, CELL DIVISION CYCLE 5 (CDC5), promotes pri-miRNA transcription and processing, with mutants reducing multiple miRNAs including miR165, miR166, and miR167 [147]. This places miRNA dosage upstream of core developmental transcription factors and suggests system-wide consequences for cambial regulation. Within this framework, miR165/166 dosage modulates the width and position of the cambial identity zone by adjusting HD-ZIP III activity [143]. A second module links miRNA to phytohormone through miR167 targeting ARFs. In cdc5-1 mutants, derepressing of ARF3/8 illustrates the sensitivity of the auxin-response machinery to miRNA dosage [147]. As ARF5 promoting CK biosynthesis and CK-induced CYCD3 drives cambial proliferation, miRNA167-mediated control of ARF dosage provides a route to couple the miRNA layer with multiple phytohormone signaling pathways and transcription factor regulatory networks [15,37,65].
Long non-coding RNAs (lncRNAs) appear to operate with miRNAs in a complementary manner. Mechanistically, lncRNAs act in cis and in trans, linking positional information to stable changes in gene activity that are distinct from miRNA-mediated mRNA cleavage or translational control [148]. A multi-tissue transcriptome catalogs 2988 high-confidence lncRNAs with tissue-oriented expression, pointing to organ- and tissue-specific developmental programs relevant to cambial identity and differentiation patterning [149].
Together, non-coding RNAs constitute a regulatory layer that interfaces with peptide, phytohormone, and transcription factor networks. Their dosage and spatial distribution likely influence stem cell domain size and differentiation output, with direct consequences for wood quantity and quality.
5. Conclusions and Perspective
Cambial stem cell activity is a dynamic process integrated through peptide signaling [31], transcription factor networks [59], and phytohormone gradients [46]. In woody plants, these elements operate within a cylindrical lateral meristem that must preserve circumferential continuity while delivering balanced xylem and phloem output [10,150]. The cambium is not a static structure but a responsive system where upstream signals continuously define the localization and maintenance of stem cells, while downstream programs determine the expansion of precursors and the deposition of SCW during cell differentiation [33,151]. This dynamic balance is essential for maintaining both the growth and functional adaptability of trees in response to fluctuating environmental conditions. Additionally, epigenetic regulators, protein turnover, and non-coding RNAs provide additional layers of complexity, refining the decisions that govern cambial behavior across diverse developmental stages [146,147].
However, a significant challenge remains in bridging the gap between molecular insights obtained from herbaceous models like Arabidopsis and their application to woody plants. The morphology and activity of xylem in woody plants are different from those in Arabidopsis. Notably, woody plants exhibit lineage-specific expansion and functionalization of key gene families (Table 1), which enriches the complexity of secondary development of woody plants. Furthermore, as perennial plants, trees face unique temporal constraints. For example, cambial activity varies dramatically with the seasons, with the number of cambial cell layers reaching dozens in summer but retreating to only a few in winter [100,105,152]. Moreover, the age difference in trees significantly influences cambial activity [105]. Deciphering how the cambium maintains its stem cell reservoir and adjusts its output in response to these seasonal and age-related variations remains a critical frontier for further investigation.
With the advancement of single-cell and spatial omics, integrative epigenomics, and quantitative modeling, we can enable a transition from merely describing cambial dynamics to predicting and designing cambial behavior in woody plants [153]. These emerging technologies hold great potential for guiding rational breeding strategies to enhance wood biomass accumulation.
In summary, the multilayered regulation of cambial stem cells, driven by dynamic feedback mechanisms, is the key to optimizing wood formation. Future studies should focus on fine-tuning these regulatory networks, including adjusting peptide signaling, phytohormone levels, and transcriptional factors, to balance proliferation and differentiation for improved wood properties [33,154]. The layered mechanism in Figure 1 and the analytical workflow in Figure 2 form a synergistic loop: the cell type profiles, spatial gradients, and regulatory nodes identified via high-resolution omics (Figure 2) calibrate the peptide hormone effector thresholds (Figure 1), ultimately providing a roadmap for optimizing traits for sustainable material production and bioenergy applications.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhu Y. Li L. Wood of trees: Cellular structure, molecular formation, and genetic engineering J. Integr. Plant Biol.20246644346710.1111/jipb.1358938032010 · doi ↗ · pubmed ↗
- 2Schuetz M. Smith R. Ellis B. Xylem tissue specification, patterning, and differentiation mechanisms J. Exp. Bot.201364113110.1093/jxb/ers 28723162114 · doi ↗ · pubmed ↗
- 3Fischer U. Kucukoglu M. Helariutta Y. Bhalerao R.P. The dynamics of cambial stem cell activity Annu. Rev. Plant Biol.20197029331910.1146/annurev-arplant-050718-10040230822110 · doi ↗ · pubmed ↗
- 4Groover A. Robischon M. Developmental mechanisms regulating secondary growth in woody plants Curr. Opin. Plant Biol.20069555810.1016/j.pbi.2005.11.01316337827 · doi ↗ · pubmed ↗
- 5Baucher M. El Jaziri M. Vandeputte O. From primary to secondary growth: Origin and development of the vascular system J. Exp. Bot.2007583485350110.1093/jxb/erm 18517898423 · doi ↗ · pubmed ↗
- 6Mellerowicz E.J. Sundberg B. Wood cell walls: Biosynthesis, developmental dynamics and their implications for wood properties Curr. Opin. Plant Biol.20081129330010.1016/j.pbi.2008.03.00318434240 · doi ↗ · pubmed ↗
- 7Caño-Delgado A. Lee J.Y. Demura T. Regulatory mechanisms for specification and patterning of plant vascular tissues Annu. Rev. Cell Dev. Biol.20102660563710.1146/annurev-cellbio-100109-10410720590454 · doi ↗ · pubmed ↗
- 8Wang X. MäkiläR. Mähönen A.P. From procambium patterning to cambium activation and maintenance in the Arabidopsis root Curr. Opin. Plant Biol.20237510240410.1016/j.pbi.2023.10240437352651 · doi ↗ · pubmed ↗