A Comparison of Volatile Components Across Native Australian Mentha (Lamiaceae)
Trevor C. Wilson, Paul I. Forster, Daniel J. Duval, Joseph J. Brophy

TL;DR
This study compares the volatile components of all native Australian mint species, revealing significant variation in their chemical profiles.
Contribution
The paper provides the first comprehensive documentation of volatile oil variation across all Australian Mentha species.
Findings
Mentha australis and M. diemenica had high concentrations of menthone and pulegone.
Mentha satureioides showed a distinct chemotype in eastern New South Wales with limonene and 1,8-cineole.
Mentha atrolilacina had a unique oil profile with linalool and β-caryophyllene as main components.
Abstract
Mentha are historically important regarding their volatile oils. Since limited accounts exist for Australian species, we document the variation in volatiles across all Australian Mentha species, using the GC/MS of pentane extractions from leaf samples of replicate populations for all known species. Oil yields were consistently poor (<0.2% w/w) for freshly dried and herbarium specimens. Many species uniformly had high percentages of volatiles characteristically attributed to Mentha (viz. Menthone, Pulegone); yet, others—consistently or variably—lacked them. Mentha australis had the highest concentrations of menthone (25%), isomenthone, (9%) and pulegone (24%), and M. diemenica had menthone (32.5%) and pulegone (29.8%). Extracts from M. grandiflora from herbarium specimens produced weak traces, high in menthone and pulegone. Mentha satureioides had the highest menthone (20–30%) and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2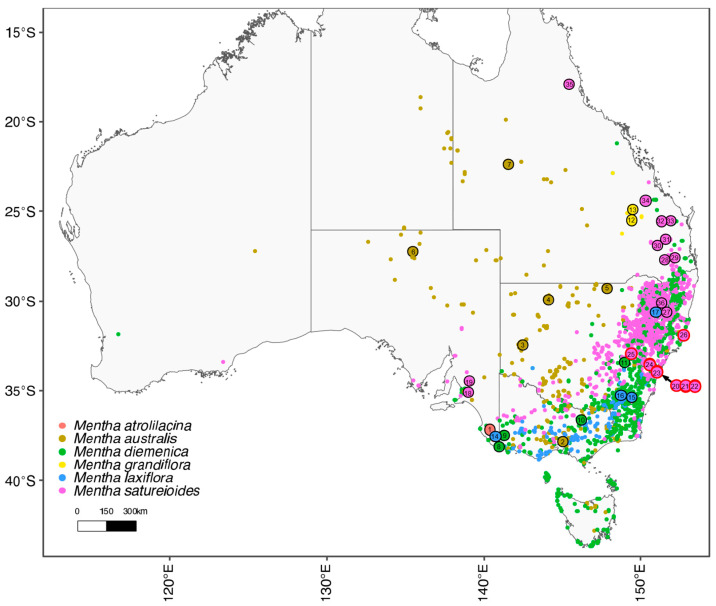 Figure 3
Figure 3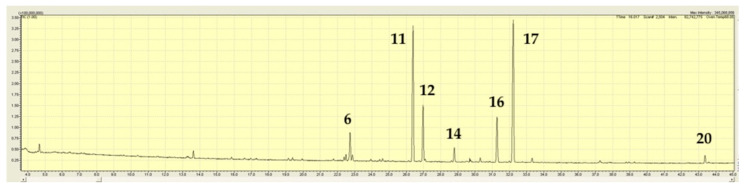 Figure 4
Figure 4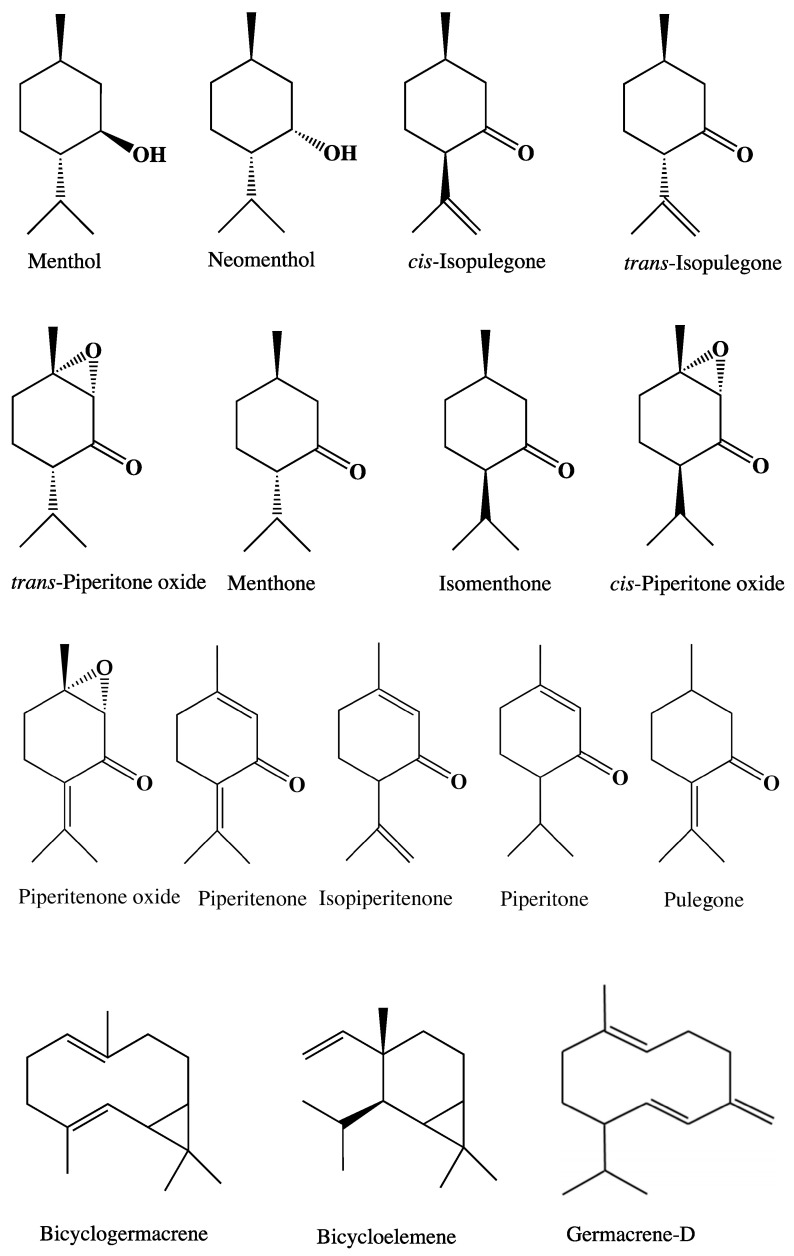 Figure 5
Figure 5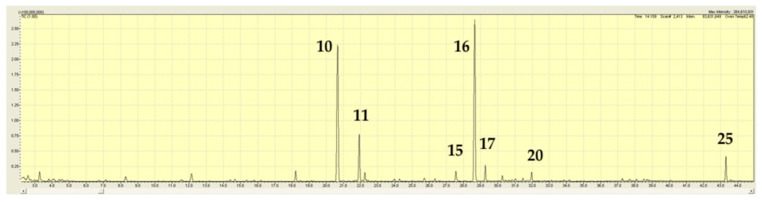 Figure 6
Figure 6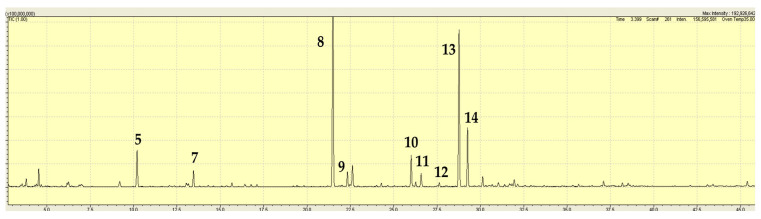 Figure 7
Figure 7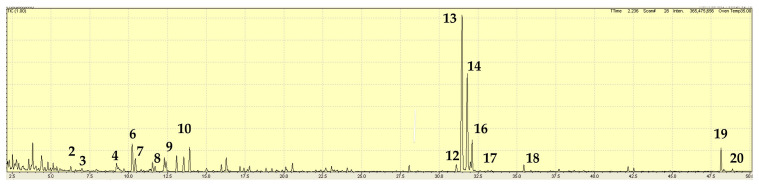 Figure 8
Figure 8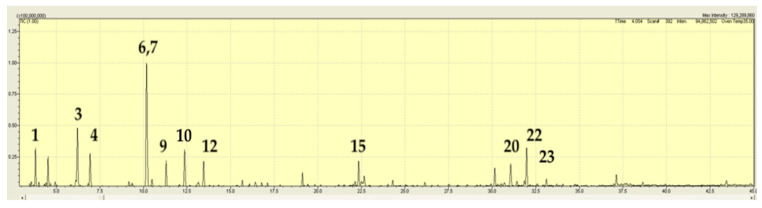 Figure 9
Figure 9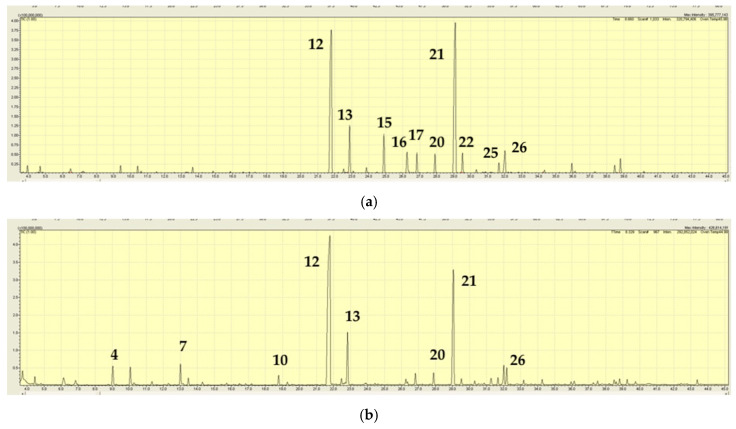 Figure 10
Figure 10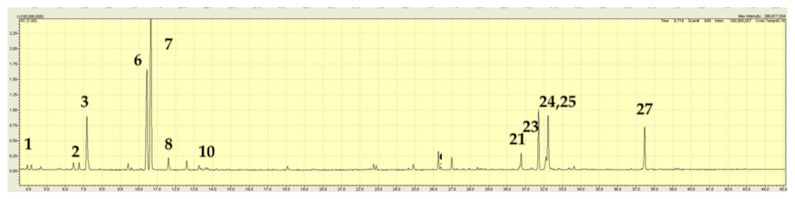 Figure 11
Figure 11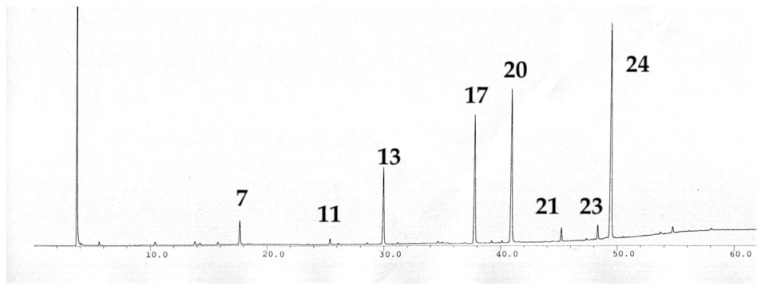 Figure 12
Figure 12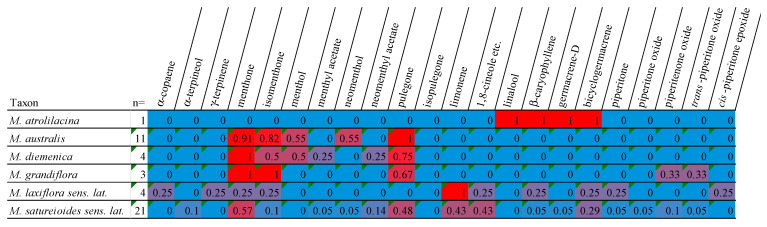 Figure 13
Figure 13Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEssential Oils and Antimicrobial Activity · Plant biochemistry and biosynthesis · Edible Oils Quality and Analysis
1. Introduction
Lamiaceae, or the ‘mint family’, are a well-recognised group useful for culinary, medicinal, and ornamental uses. Likely the most celebrated of its members is Mentha L., commonly known as ‘Mint’, which are fragrant rhizatomous herbs of generally damp, open habitats [1]. The aromatics in Mentha are most celebrated for flavouring food and drinks, but extracts from these plants have a broad range of applications including as antimicrobials, biocides, and medicines (Figure 1) [2,3,4,5,6,7,8,9]. Commercial oil production is largely restricted to only a few species or cultivated hybrids, including M. citrata Ehrh. (M. aquatica L. var. citrata (Ehrh.) Fresen), M. canadensis L., Mentha × piperita L. (Peppermint), M. pulegium L. (Pennyroyal), M. spicata L. (Native Spearmint), and M. × gracilis Sole (Scotch Spearmint). The popular flavours these plants produce (e.g., mint, pennyroyal, peppermint, and spearmint) is due to their high concentrations of menthols and pulegone, menthofurans, and carveols [10].
The taxonomy of Mentha has had a legacy of instability [11], with 25 species and 15 hybrids currently recognised [12]. Widespread polyploidy associated with cryptic species appears to have complicated an accurate delineation of the component taxa. It is being revealed through genomic tools how a mismatch between the genotype and morphotype has undermined traditional species identification and complicated the extent of our knowledge about its biodiversity [13,14]. Within Australia, there are six naturally occurring, endemic Mentha species (Figure 2): M. atrolilacina B.J.Conn & D.J.Duval, M. australis R.Br., M. diemenica Spreng., M. grandiflora Benth., M. laxiflora Benth., and M. satureioides R.Br [15,16,17]. These species form a distribution that spans the eastern half of the continent (Figure 3). Mentha pulegium is a seventh species naturalised throughout southeastern Australia. Although this exotic species is morphologically similar to M. diemenica and M. satureioides (i.e., ‘M. satureioides group’ sensu Conn [15]), it can readily be distinguished by the densely flowered verticillasters (vs. sparsely flowered verticillasters) and hairy inner corolla tube (vs. glabrous inner corollas tube) [15,18]. According to the most recent phylogenetic study of Mentha infrageneric relationships, Australian species (i.e., M. diemenica and M. satureioides) and the New Zealand M. cunninghamii Benth. form a clade that is sister to Eurasian and American species [19]. One study has so far provided empirical morphological evidence about the distinctiveness between M. australis and M. diemenica [20], but no other test of species boundaries or phylogenetic study for Australasian species has been made.
The most comprehensive treatment for Australian Mentha is shared in the Flora of New South Wales and Flora of Victoria [15,16]. Both of these flora account for four of the six Australian species (i.e., M. australis, M. diemenica, M. laxiflora, and M. satureioides). Relative to the ‘M. satureioides group’, M. australis and M. laxiflora are readily identified by having more or less spreading hairs on the inner surface of their calyx (vs. villous hairs) and have few- to many-flowered inflorescences (i.e., up to seven per axil vs. three per axil). Mentha grandiflora is similar to M. australis, but has broader leaves and larger flowers [21,22], and M. atrolilacina is similar to the ‘M. satureioides group’ [18] but has cordate leaves (vs. ovate or elliptic) and flowers with inserted anthers (vs. exserted anthers).
A persisting challenge in the taxonomy of Australian Mentha continues to be the ‘M. satureioides group’ (sensu Conn [15,16]), as it is unclear whether it represents distinct species or extremes of a morphological continuum. Declaring this ambiguity, for the interim, Conn [15,16] distinguished M. satureioides from M. diemenica as having nearly glabrous branches (vs. hairy branches), glabrous outer calyx (vs. hairy calyx), narrow-elliptic to linear-elliptic leaves (vs. ovate, often angular-ovate and rarely narrow-ovate or elliptic leaves) with an entire margin (vs. sometimes having a margin with distant shallow teeth, if not entire). Conflict in the recognition of species for this group is apparent between state floras. For example, the National Herbarium of NSW (NSW) identifies specimens collected from Queensland as either M. satureioides and M. diemenica; yet, the Queensland Herbarium (BRI) recognises only M. satureioides as indigenous to the Queensland state.
Unsurprisingly, use of Australian Mentha has been similar to that in the northern hemisphere. Decoctions of M. australis were considered by indigenous Australians as beneficial for coughs and colds as well as for an abortifacient [23]. After the arrival of Europeans to Australia, accounts of M. australis (or ‘River Mint’) identified the scent of its oil as similar to peppermint, but coarser [2]—it has since been used for flavouring (e.g., desserts and drinks), cold and cough medicine, insect repellent, and antioxidant or anti-inflammatory compounds for skincare [3]. The characterization of phenolics (higher-molecular-weight components, not volatile ones) [24] and antioxidants [25] have been intensively investigated for this species; yet, the oil chemistry has been quantified by only one study using ex situ material in Saudi Arabia [26]. The essential oil of M. laxiflora was recorded as similar to that of peppermint, and that of M. grandiflora is strongly fiery, bitter, and unpleasantly nauseous to taste [2]. Mentha satureioides (syn. Mentha gracilis) was originally described as similar to peppermint, and reported to have a slight admixture of pennyroyal and be quite disagreeable and acrid [2]—its uses have included being a diuretic, diaphoretic, insect repellent, or for regulating menstrual cycles [2]. For this species, the most intensive essential oil study of any Australian Mentha distilled 100 lbs. of ‘wilted’ M. satureioides from Dalby Qld, obtaining 93 g of oil (0.2%) that was described to resemble the “pennyroyal oils of commerce” [27]. Later, the essential oil composition of M. satureioides (originally identified as M. diemenica—see taxonomy above) from Didcot, Qld (approximately 100 km north of the Dalby location) was reconfirmed as having significant amounts of menthone and pulegone [28].
The examination of replicate material in the quantification of volatiles is a sensible idea, granted that individuals even within a population of the same species can have a different oil composition. Early investigations of eucalyptus essential oils suggested that the composition of the oil was typically constant within the same species [29]—the typical leaf oil make up for Eucalyptus [30], as well as Melaleuca [31], can be defined as including 1,8-cineole (~60–70%), α-pinene (~10%), limonene (~8%), and α-terpineol (~5%). However, it has been discovered that trees of the same species, even growing beside one another, can produce different compounds (in several families including Myrtaceae), and thereby are described as separate chemotypes (e.g., [31,32,33]). Nonetheless, distinct oils and oil profiles continue to assist with the identification and description of new taxa (Eucalyptus and others) [27,34,35,36,37], and the miniaturization of methods (working with mgs of leaf material) to quantify and qualify them is enabling other uses of herbarium samples to assist with species description [38,39].
In this study, we aim to provide the first comprehensive study of oil quantification for all Australian Mentha—we reanalyse previously sampled species, as well as provide the first account of oil composition for M. atrolilicana and M. laxiflora. Using GC/MS (gas chromatography/mass spectrometry) Total Ion Current (TIC) traces, we sampled the fresh material of most species, comparing relative rather than absolute quantities of compounds. Furthermore, sampling was augmented using herbarium material to include several populations across a species’ distribution in recognition of the unresolved taxonomy of Mentha [15,16], and the likelihood that several chemotypes may exist within a species. Additionally, through sample provision from multiple Australian horticulture suppliers, the wild type was also compared with industry stock. We ask the following questions: what are the chemotypes for each species, do they vary within species, and do they compare between samples sourced from commercial suppliers and with a known provenance?
2. Results
2.1. Mentha atrolilacina
Mentha atrolilacina GC/MS TIC traces demonstrated a mixture of volatiles containing four major components, the majority being sesquiterpene hydrocarbons. These were linalool (21.2%), β-caryophyllene (14.1%), germacrene-D (13.3%), and bicyclogermacrene (23.7%), (Figure 4; Table 2). Also present in amounts between 1–5% were γ-elemene, α-copaene, aromadendrene, δ-cadinene, and germacrene-D-4-ol (see Figure 5 for conformation of some of these compounds). There were more components present in amounts < 1%.
2.2. Mentha australis
The major volatile components in the fresh M. australis extract were menthone (32%), isomenthone (11%), and pulegone (31%); see Table 3 and Figure 6, below. These components were accompanied by lesser amounts of α-pinene and β-pinene (1–1.5%), hex-2-enal (1.5%, isomer not determined), 3-hexene-1-ol (2.2%, isomer not determined), neomenthol (2.3%), menthol (3.1%), bicyclogermacrene (1.9%), and germacrene-D-4-ol. One individual plant from Melbourne, Victoria (JB6180) contained less menthone (20%) than pulegone (70%). From the extracts of herbarium samples, menthone, pulegone, and isomenthone were detected as prominent components in the extracts (Table 1).
2.3. Mentha diemenica
Fresh extracts of M. diemenica from Honans Native Forest Reserve, SA (JB6209) (see Table 4, Figure 7) contained menthone (32.5%) and pulegone (29.8%) in high amounts, and a somewhat high amount of menthol (9.7%) (Table 4, Figure 7). Herbarium extracts similarly had the highest amounts of menthone and pulegone, except one population from Victoria (JB6204) which was missing pulegone. Also matching the fresh extract were other compounds detected from some herbarium specimens including isomenthone, neomenthyl acetate, menthone, isomenthone, and menthol (Table 1).
2.4. Mentha grandiflora
Our extractions were solely based on herbarium specimens. Our results identified the major components of pulegone and menthone (Table 1).
2.5. Mentha laxiflora
The volatiles, as major components in M. laxiflora, differed among the extracts of fresh specimens and herbarium specimens. The extraction of fresh material from Tallaganda National Park (JB6231) showed piperitone (24.9%) and cis-piperitone epoxide (30.4%) as major components. Other significant components were trans-piperitone epoxide (7.4%), limonene (9.0%), 1,8-cineole (3.5%), p-cymene (4.1%), thymol (5.2%), and bicyclogermacrene (2.1%) (Table 5, Figure 8).
The extract from the South Australian M. sp. aff. laxiflora (JB6182) was dominated by limonene (27.4%), with significant amounts of the monoterpenes α-pinene (5.8%), β-pinene (11.3%), sabinene (6.3%), γ-terpinene (7.2%), and the olefinic 2-hexenal (4.4%, isomer not identified), and the principal sesquiterpenes being germacrene-D (4.5%) and bicyclogermacrene (7.1%) (Table 6, Figure 9).
Extracts from both herbarium specimens provided limonene; however, plants from the Goodradigbee River population also contained α-copaene (similar to Dry Creek) and β-caryophyllene, whereas, from the Warrabah National Park, the population showed 1,8-cineole, menthone, and isomenthone being present (Table 1).
2.6. Mentha satureioides
Mentha satureioides in Queensland and South Australia (Table 7, Figure 10) generally represent opposite ends of the species distribution. Comparing the fresh extracts from both ends of the distribution, the major components were menthone (22.5, 36%) and pulegone (22.5, 12.1%), respectively. These were accompanied by lesser amounts of isomenthone (8.7, 6.7%), neomenthyl acetate (7.2%), menthyl acetate (4.7%), cis-isopulegone (3.5%), trans-isopulegone (1.4%), piperitone (5.3, 2.8%), bicyclogermacrene (2.2%), and piperitenone (2.4%). Most other fresh and herbarium extractions were similar in their major volatiles, including menthone, and, to a lesser extent, pulegone—in herbarium specimens from northern New South Wales and northern Queensland, menthone was accompanied with either piperitone oxide and piperitenone oxide (former), or neomenthyl acetate and menthyl acetate (latter) instead.
The fresh extracts sourced from Sydney, New South Wales (NSW) yielded a different set of volatiles, representing a second chemotype (Table 8, Figure 11). The extract was dominated by limonene (17.3%) and 1,8-cineole (19.2%), with sabinene (9.4%), α-terpineol (8.1%), and bicyclogermacrene (9.1%) being the other major components. Neomenthyl acetate and menthyl acetate (totaling 3.7%) were the only components in common with the fresh extracts of the first chemotype demonstrated above. Other fresh extracts from Sydney populations, Taree, all commercial supplies of unknown provenance, and herbarium specimens from Glenbrook and near Kerrs Creek similarly had major amounts of limonene and 1,8-cineole (Table 1).
3. Discussion
This is the first comprehensive account of the volatile compounds for Australian Mentha species, and the first account of the volatile compounds for M. atrolilacina and M. laxiflora. We found that the levels of menthone or pulegone (or close derivatives), which are characteristic economically useful volatiles attributed to the genus, were not consistently high across all species (e.g., M. atrolilacina), nor even within some species (e.g., M. satureioides). This alludes to the potential for chemotypes in Australian Mentha species, or, perhaps, to the idea that there are species that have not yet been identified.
3.1. Species with High Menthone and Pulegone Oil Profiles
We found that, across our replicated sampling of populations, M. australis, M. diemenica, and M. grandiflora are consistently high in levels of menthone (25–40%) and pulegone (25–40%) as dominant components. In addition to menthone and pulegone, there are lesser amounts of related compounds, both in the oxidation state and functional groups. For the profiles of M. grandiflora and M. diemenica, these are supplemented by trans-piperitone oxide and piperitenone oxide and other oxidised compounds based on piperitone (in all of these structures, there is oxidation at the carbon adjacent to the isopropyl group).
The uniform oil profile of M. australis with distinctively high amounts of menthone and pulegone documented in all wild populations sampled in this study corresponds to the early reports (i.e., the 1861 Dublin Exhibition) that mention the species has ‘pennyroyal’ smells [2]. Fresh extracted commercially supplied samples, which have the highest amounts of menthone followed by pulegone, match closely with all samples we analysed from herbarium specimens. Fresh wild-sourced material from the Melbourne population [JB6180] does have a disproportionately larger amount of pulegone than menthone. However, such a difference in the oil profile is comparably small in relation to the only other study of isolated extracts from ex situ M. australis samples [26], which found distinctively high amounts of linalool, and little to no trace of menthol. The fact that our relatively consistent results from a widespread Australian sampling also matches the earliest anecdotal evidence of M. australis volatiles suggests the anomalous account is due to a misidentification. However, this is difficult to test given that the previous study did not disclose the herbarium records or the provenance of its material.
The high levels of menthone and pulegone match previous studies made for M. grandiflora from Precipice National Park, Queensland (see Table 9, Figure 12). From our only sample of M. grandiflora—a herbarium specimen collected in Palmgrove National Park—we obtained two extremely small peaks, which were of comparable intensity to the impurities in our pentane solvent, matching the two distinct menthone and isomenthone peaks shown by [22]. The poor signal from our extraction likely did not exhibit other volatiles similar to those found in smaller amounts from the previous study (which were obtained by steam distillation on a much larger amount of fresh leaf) [22], such as the piperitenone oxide (36.2%), trans-piperitone oxide (21.4%), pulegone (19.1%), menthone (9.7%), piperitenone (1.7%), bicyclogermacrene (2.1%), and limonene (3.5%) (Table 9, Figure 12).
The high levels of menthone and pulegone we recorded from fresh M. diemenica (JB6209) match the results of previous extractions of cultivated material originally sourced from Canberra, ACT [28]. Similar to this study, we also note, in lower quantities (yet still above 1%), neomenthyl acetate, isomenthone, cis-isopulegone, and myrcene. However, unlike this study, we also detected limonene, cis-isopulegone, and myrcene. Despite being weak in signal, herbarium specimens similarly yielded the highest levels of menthone and pulegone.
3.2. Two Putative Chemotypes for Mentha satureioides
Most M. satureioides populations we examined have high levels of menthone and pulegone, in the same reasonably high quantities as M. australis, M. grandifolia, and M. diemenica. This includes all our sampling from Queensland and South Australia, and one from New South Wales (i.e., JB6207), whose quantities range around approximately 32% menthone and 25–44% pulegone. A previous GC-MS study [28] of M. satureioides (identified as M. diemenica) that also examined samples from Didcot Queensland identified high levels of menthone (32%) and pulegone (25%), with neomenthyl acetate (18%) and menthyl acetate (6%) also found in substantial but lower quantities. Similarly, an earlier study using pure oil extract [27] showed that Dalby Queensland populations (only 30 km south of our JB6185—Jimbour West population) have high pulegone (40%) and l-menthone (20–30%), although our extraction demonstrated nearly equal quantities of menthone and pulegone (Table 7).
The remainder of samples we extracted from the New South Wales Central Coast, Central Tablelands, and (in part) North Coast botanical districts show a remarkably different chemotype. This second M. satureioides chemotype is distinct because it is consistently dominated by limonene (~17%) and 1,8-cineole (~19%), whereas menthone and pulegone were either not detected, or detected in low concentrations (<1%). Material from all commercial suppliers (2) that we sampled from exhibited this second chemotype.
3.3. Mentha atrolilacina Oil Profile
Mentha atrolilacina has the most distinctive profile of volatile oils compared to any other Australian Mentha species (Figure 13). Bicyclogermacrene is the principal component highest in quantity (24%) and found in lesser quantities across all other species of Mentha we examined. This species also has a nearly similar concentration of linalool (21%), a compound found highest in concentration in some non-Australian species such as M. citrata [40]. This chemical is not found in high quantities in any other Australian species from our result, and from previous studies [22,28]. One exception is that of ex situ material of M. australis from which linalool was identified as the most dominant compound [26].
3.4. Mentha laxiflora Oil Profile
Mentha laxiflora (like M. satureioides) also has variability in its main volatile compounds across the range we examined. The fresh material we examined from Tallaganda National Park in the Southern Tablelands of New South Wales (JB6231) has, as main components, piperitone and cis-piperitone epoxide, the latter also found in higher quantities for M. grandiflora [22]. Similar to menthone and pulegone, such compounds equally have a smell traditionally considered as ‘minty’. Hence, although no chemical compounds have previously been analysed, early reports about the ‘minty’ smell of M. laxiflora extracts at the 1861 Dublin Exhibition [2] correspond with our results. However, the population we examined in South Australia (JB6224) has a notably distinct oil profile with limonene (28%) as a principal component (with the next highest component being β-pinene, 12%). Our broader screening across the known distribution of M. laxiflora using herbarium specimens did indeed identify that limonene is a major component consistent across all populations; yet, this was accompanied by a combination of other compounds unique to each sample site. Hence, given the poor signal delivered in analyses of herbarium specimens, a reanalysis of fresh material would be necessary to confirm the exact quantities across this variation. Menthone was the only other component we detected that would produce a ‘minty’ smell corresponding with the early anecdotal evidence, but we only found it in the Goodradigbee River specimen (JB2618), proximally located to the sample from Tallaganda National Park in the Southern Tablelands of New South Wales.
3.5. Taxonomic Insights and Conclusion
We found variations in the dominant essential oils across Australian Mentha through sampling replicates for most species, and that not all of them have the main oil components corresponding to ‘mint-like’ smells as traditionally thought (e.g., menthone or pulegone). All replicates of M. australis, M. diemenica, and M. grandiflora yield such a ‘mint-like’ profile, whereas not all populations of M. atrolilacina, M. laxiflora, and M. satureioides do. Mentha laxiflora and M. satureioides have limonene in higher quantities for all replicate populations. However, other compounds in a high quantity vary between M. laxiflora populations, whereas they form two consistent chemotypes in M. satureioides.
Two distinct chemotypes within M. satureioides might merely be variations in the oil composition, since the quantities of oil may be expressed within different individuals of a species based on the genetic variation, ontogenetic processes, and environmental variation [34,41]. Despite some level of obfuscation, chemovariation can often align with phylogenetic signals such as interspecific boundaries [37]. Hence, such a discrete difference in the chemotypes of M. satureioides could indicate different taxa, especially since the high menthone and pulegone chemotype is found in similar woodland habitats as the high limonene chemotype, but, yet, is found in a broader, overlapping climatic envelope that also spans the well-recognised biogeographic barrier of the Great Dividing Range [42]. Considering the number of ambiguities currently associated with the ‘M. satureioides group’, any taxonomic conclusion is premature and a robust phylogenetic investigation is needed before the future identification of chemotaxonomic markers can be accomplished. Such an examination would require genomic data with a population-based sampling approach for all species, with special attention towards representing the morphological and chemical variation of the ‘M. satureioides group’ and M. laxiflora, and any instances of sympatry (e.g., M. diemenica and M. atrolilacina at Honans NFR) that could potentially reveal the levels of gene flow and reproductive isolation.
For the industry actively searching for Australian sources of ‘mint-like’ flavouring, our results so far support that M. australis is the best option. We confirm high quantities of menthone and pulegone are in M. australis, though the toxic nature of pulegone should be taken into account, and this is consistent across all wild populations and the wide number of plants we obtained from the horticultural trade. Populations currently identified as M. diemenica or M. satureioides might be useful as well; yet, we are hamstrung by the current lack of a robust taxonomy that would otherwise assist us with diagnosing phylogenetic patterns in chemovariation. Such a phylogenetic study would permit a more informed approach towards selecting plants for desired flavours (e.g., by pinpointing species highest in menthones), or low toxicity (e.g., by pinpointing species with lower levels of pulegones). A sound taxonomic framework is fundamental to equipping such downstream research endeavours, just as it is a necessity for the conservation of our biological heritage.
4. Materials and Methods
4.1. Plant Material
Morphology of plants was examined in the field and in cultivation. Specimens were consulted at the National Herbarium of NSW (NSW). Leaf material was collected from freshly collected, air-dried plants, or from herbarium specimens (Table 1). Herbaria acronyms are listed here according to Index Herbarium [43], and we identify botanical regions in New South Wales [44,45] according to the defined areas based on the National Interim Biogeographic Regionalisation for Australia system IBRA v7 [46]. Since similar studies have sampled from herbarium samples and shown the success of the technique [38,39,47], we sampled from herbarium samples at NSW that ranged from between 36 and 50 years old. Mentha australis and M. satureioides leaf oil were each extracted from fresh samples obtained through three different commercial suppliers, with the provenance of only one sample (JB6187) known to be from near Taree, New South Wales.
Robert Brown’s type material of M. satureioides (BM 000838674) was collected from Parramatta, New South Wales [48] and has mostly elliptic to ovate leaves that often have a slightly toothed margin—this is consistent with all NSW-held samples of the species collected from Queensland and the New South Wales Central Coast botanical district, as well as other specimens from the South Coast and North Coast botanical districts. However, specimens identified as M. satureioides that are found across this distribution and that resemble the type material can vary in the characters used to distinguish them in the Mentha key of the NSW and Victoria flora [15,16]. That is, there is a gradient in hair density of branches between individuals (i.e., from glabrous, or nearly so, to hairy), leaf lamina can range between linear and ovate, and the leaf margin can be entire to shallowly toothed on the same plant. Such morphological variation complicates the final couplet distinguishing the two taxa in the Mentha key in the NSW and Victoria flora [15,16].
There are also complications with diagnostic characters used in identification keys as revealed by our examination of the M. diemenica holotype (syn. M. gracilis R.Br.) made also by Robert Brown (BM 001041075). Though the NSW and Victoria Flora accounts describe M. diemenica as having ovate to angular-ovate leaves that correspond with Brown’s type material, their identification of plants with glabrous stems (c.f., hair) and often-toothed leaf margins is inconsistent with this, as well as with other material from New South Wales, Victoria, South Australia, and Tasmania.
For this study, we abandon the use of the stem indumentum as a diagnostic character and rely solely on matching leaf shape and marginal teeth on the leaf lamina with original material of M. diemenica (ovate, and often angular-ovate) (Figure 2g). For the purpose of this paper, we use M. satureioides sens. lat., referring to specimens that have linear leaves with an entire margin to ovate leaves with a shallowly toothed margin (e.g., Figure 2c–e).
A number of specimens have been identified as M. laxiflora on both sides of the South Australia–Victorian Border, the western-most extent of the species’ distribution [49]; yet, they do not closely resemble the type specimen made by Hooker (an unknown locality of Australia; K 000494937). For instance, our specimen JB6182 from this area (Figure 2h) in comparison to material resembling the type specimen (Figure 2i), has leaves less than 10 mm long (c.f., up to 30 mm long) and an entire to shallow-toothed margin (c.f. serrate). It furthermore has calyx lobes whose inner surface is glabrous, rather than having villous or spreading hairs as indicated by [15,16]. Hence, due to the distinctively different morphology, we identify this specimen as M. sp. aff. laxiflora.
4.2. Extraction of Volatile Components
Extraction of the volatiles was from the leaves. A small amount of leaf material (three or four leaves, between 0.05 g and 0.1 g fresh weight, or 0.02 g and 0.05 g dry weight) was placed in a two ml screw top vial, and then immersed in one ml pentane and sealed. The solution was kept at room temperature for at least two days and shaken at least once a day, and then the pentane solution was analysed by GC/MS as one sample of a batch run. As well as the extract, samples also included a solvent blank and a sample containing a solution of C10–C25 alkanes, to allow linear retention index (LRI) data to be calculated. From the strength of the TIC traces, the oil yields appeared to be poor.
4.3. Analysis of the Pentane Extract
The chief aim of this study was solely to acquire and compare relative rather than absolute amounts of volatile organic compounds and to identify the chief compounds in each Mentha specimen’s oil profile. The response from the Total Ion Current (TIC) trace is similar for all compounds found in this work because they, with very few exceptions, are all aliphatic/olefinic organic compounds. Only two aromatic compounds (thymol and carvacrol) were found in the whole of this work, and these were present at low levels. Aromatic compounds, because of the stability of the ions readily formed in the mass spectrometer ion source, tend to give a larger signal than those from aliphatic or olefinic compounds. Therefore, the small TIC signals for thymol and carvacrol indicate that they were present in very low levels. We found the mass spectrometer response to be similar among the compounds detected in this survey. We think it valid to show and produce the relative integrated area counts on all the TIC peaks. The presence of γ-terpinene in some samples and the lack of its oxidised product, p-cymene, indicates that the samples were not oxidised.
Analysis of the extract was carried out by combined gas chromatography/mass spectrometry (GC/MS) on a Shimadzu GCMS-2020 mass spectrometer, operating at 70 eV ionisation energy, in the e.i. mode. The GC column used was a BP-20 column (30 m × 0.25 mm × 0.25 μm), programmed from 35 °C to 220 °C at 3 °C/min with helium (1 mL/min) as carried gas, and injector temperature was 250 °C. Then, 1 mL of pentane solution was injected with a 5:1 split ratio. Mass spectra were recorded in the electron impact (EI) mode at 70 eV, scanning the 41–450 m/z range. Interface and source temperatures were 250 °C and 220 °C, respectively, with a 1 scan/sec cycle rime. Compounds were identified by their mass spectra, comparison with Adams and Wiley libraries, and linear retention indices (LRI). The hydrocarbon standard solution (C10–C25) was made from individual alkanes, purchased from Aldrich Chemical Co. USA and Tokyo Kasai Chemical Co. Tokyo. They were dissolved in pentane, ~10 mg/10 mL. As the number of volatiles in the herbarium samples was usually very small, the associated peaks, as opposed to the peaks from the impurities in the pentane solvent, were of comparable height. In these cases, the peaks in the TIC runs were examined individually to ensure that only compounds arising from the sample were reported on relative to n-alkanes [50,51] and by comparison of their mass spectra with either known compounds or published spectra [52] and Wiley libraries.
Every sample was analysed by GC/MS. For each species, these TIC traces were visually compared to find traces that lacked significant solvent impurity peaks. The chosen TIC trace was then manipulated to remove the response caused by the solvent impurity (even though the peak still remained on the TIC trace). The original TIC trace was then printed, but the peak areas reported from the result of removal of the impurity. The chart is characteristic of the TIC traces of that species or of the chemotype of that species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Harley R.M. Labiatae VII. 2. Tribe Mentheae Dumort (1827)The Families and Genera of Vascular Plants, Vol. VII Flowering Plants. Dicotyledons—Lamiales (Except Acanthaceae, Including Avicenniaceae) Harley R.M. Atkins S. Budantsev A.L. Cantino P.D. Conn B.J. Grayer R. Harley M.M. de Kok R. Krestovskaja T. Morales R. Springer Berlin/Heidelberg, Germany 2004230250
- 2Maiden J.H. The Useful Plants of Australia (Including Tasmania)Turner and Henderson Sydney, Australia 1889194, 280281
- 3Lassak E.V. Mc Carthy T. Australian Medicinal Plants Methuen Australia Sydney, Australia 19831240
- 4Lawrence B.M. The composition of commercially important mints Mint: The Genus Mentha Lawrence B.M. CRC Press Boca Raton, FL, USA 2007217324
- 5Mamadalieva N.Z. Hussain H. Xiao J. Recent advances in genus Mentha: Phytochemistry, antimicrobial effects, and food applications Food Front.2020143545810.1002/fft 2.53 · doi ↗
- 6Kimbaris A.C. González-Coloma A. Andrés M.F. Vidali V.P. Polissiou M.G. Santana-Méridas O. Biocidal Compounds from Mentha sp. essential oils and their structure-activity relationships Chem. Biodivers.201714 e 160027010.1002/cbdv.20160027027770481 · doi ↗ · pubmed ↗
- 7Brahmi F. Khodir M. Mohamed C. Pierre D. Chemical composition and biological activities of Mentha species Aromatic and Medicinal Plants—Back to Nature El-Shemy H. Intech Open London, UK 20174779
- 8Tafrihi M. Imran M. Tufail T. Gondal T.A. Caruso G. Sharma S. Sharma R. Atanassova M. Atanassov L. Valere Tsouh Fokou P. The wonderful activities of the genus Mentha: Not only antioxidant properties Molecules 202126111810.3390/molecules 2604111833672486 PMC 7923432 · doi ↗ · pubmed ↗