Multifaceted Roles of SPL9 in Plant Growth, Development, and Stress Responses: A Review
Yang Gao, Yu Chen, Jingxia Zhang, Le Zhang, Zhangqiang Song, Furong Wang, Shengli Wang, Jun Zhang

TL;DR
This review explores how the SPL9 protein influences plant growth, development, and stress responses, and its potential for improving crops.
Contribution
The paper provides a comprehensive review of SPL9's regulatory mechanisms and its multifaceted roles in plant biology.
Findings
SPL9 regulates plant morphogenesis, reproductive processes, and crop yield.
SPL9 plays a role in plant responses to both abiotic and biotic stresses.
SPL9 integrates into broader regulatory networks affecting plant development.
Abstract
The SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) family, a group of plant-specific transcription factors, plays a key role in plant growth and development through complex regulatory networks. Within this family, SPL9 has been identified as a key regulator of diverse biological processes. In this review, we summarize the current knowledge on SPL9, focusing on its expression regulatory mechanisms and roles in plant development, such as morphogenesis, reproductive processes, and crop yield determinations. We further describe its role in plant responses to abiotic and biotic stresses and its integration into broader regulatory networks. We also outline future research priorities and discuss potential applications of SPL9-based strategies in molecular design breeding to increase crop productivity and stress resistance.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
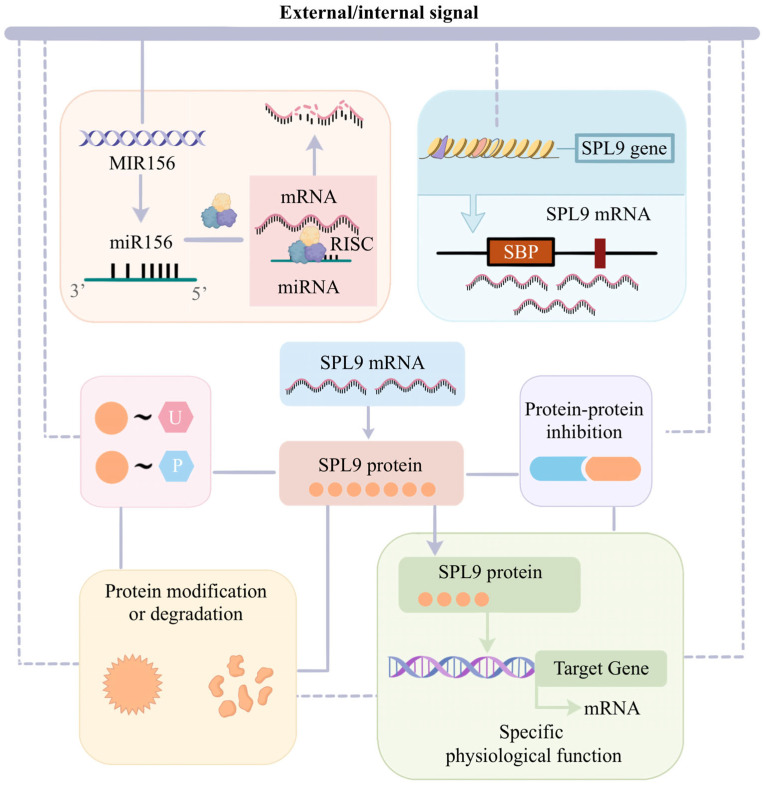 Figure 1
Figure 1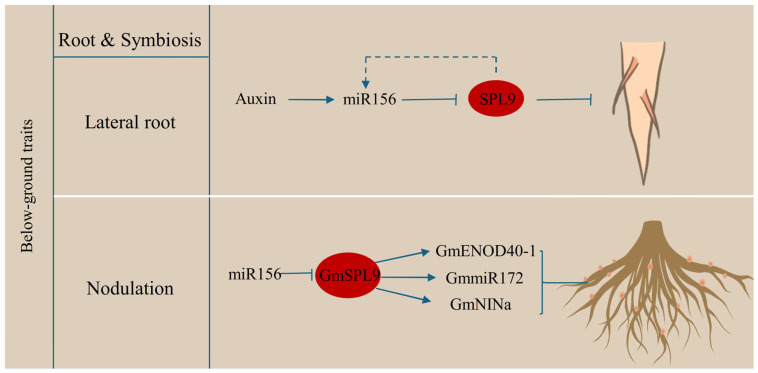 Figure 2
Figure 2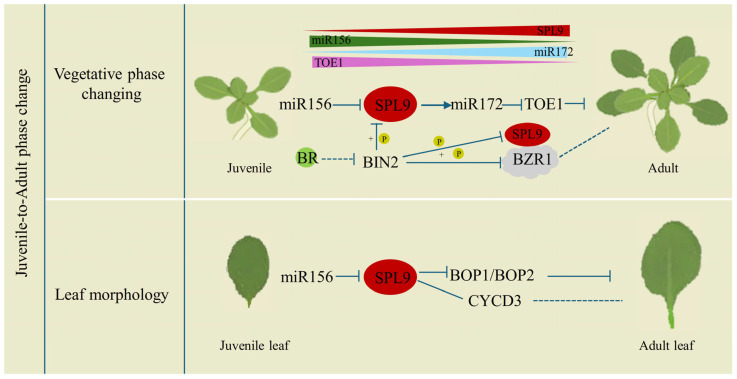 Figure 3
Figure 3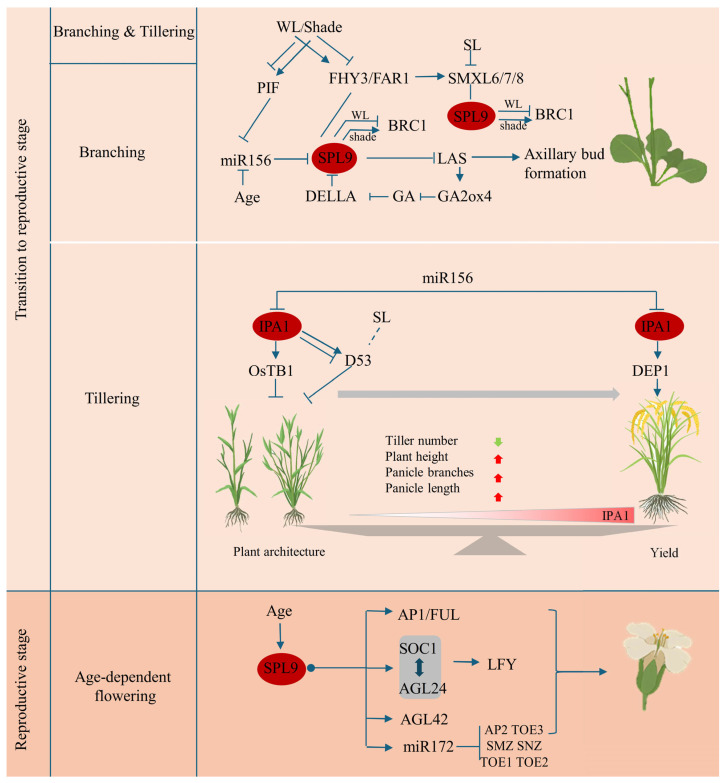 Figure 4
Figure 4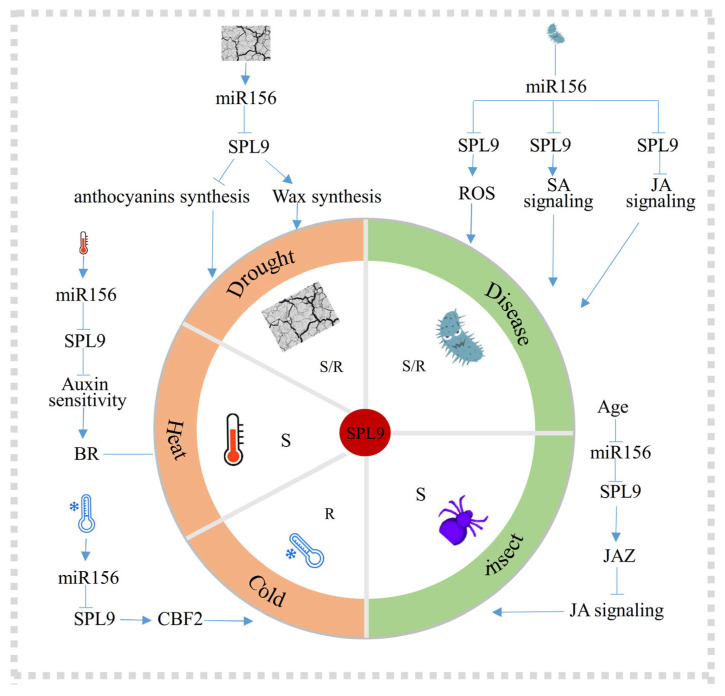 Figure 5
Figure 5- —The Key Research and Development Project of Shandong Province
- —Natural Science Foundation of Shandong Province
- —The Science and Technology Innovation Project of Shandong Academy of Agricultural Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicrotubule and mitosis dynamics · Lipid metabolism and biosynthesis · Plant Molecular Biology Research
1. Introduction
Plant development is governed by complex regulatory networks that integrate hormonal, signaling pathways, transcription factors (TFs), and various other molecular regulators. TFs binding to specific DNA sequences can either activate or repress gene expression, modulating diverse plant growth, developmental processes, and stress responses [1,2]. Among the various TF families, the SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) family plays key roles in numerous plant biological processes, including organ development [3,4], plant architecture formation [5], hormone signaling [6,7], and stress responses [8].
The SPL genes AmSBP1 and AmSBP2 were first identified in Antirrhinum majus L., where they were shown to regulate early stages of flower development [9]. After more genomic data can be used, numerous SPL genes have been identified in both monocotyledonous [10,11,12,13,14] and dicotyledonous species [15,16,17,18,19,20]. SPL proteins are evolutionarily conserved in land plants and consist of a conserved SBP domain of about 79 amino acids. This domain has two distinct Zn^2+^-binding motifs, Cys-Cys-His-Cys and Cys-Cys-Cys-His, as well as a putative nuclear localization signal [21,22,23]. The SBP domain is necessary and sufficient for binding to the TNCGTACAA sequence in the promoters of target genes, with the GTAC sequence serving as its core motif [23,24].
In Arabidopsis thaliana, 17 AtSPL genes have been identified, and 10 of which are post-transcriptionally regulated by microRNA156 (miR156) [25,26]. Among them, SPL9 is known to regulate developmental transitions such as the juvenile-to-adult phase change and the onset of reproductive growth [7,26,27]. Subsequent studies have shown that SPL9 plays a critical role throughout almost the entire Arabidopsis life cycle, influencing the development of vegetative organs such as lateral roots [28], stem branches [29] and leaves [30], as well as flower meristem activity during the reproductive stage [31,32]. In addition, SPL9 contributes to plant responses to abiotic [33,34] and biotic stresses [35,36], partly through and accompanied by the regulation of the hormone signaling pathway [37].
Notably, studies on SPL9 have extended beyond Arabidopsis. In rice, 19 OsSPL genes have been identified and systematically renamed according to their chromosomal positions [38]. Among them, the OsSPL14- and OsSPL17-encoded proteins cluster within the same phylogenetic clade as AtSPL9 and share significant protein domain similarity, suggesting conserved functions. OsSPL14, also known as IPA1, (Ideal Plant Architecture 1), was independently identified through quantitative trait locus (QTL) analyses: one focused on plant architecture [39], and another linked the WFP (WEALTHY FARMER’s PANICLE) locus to primary panicle branching [40]. It has been confirmed that IPA1 enhances rice plant architecture by reducing tillering and improving yield, while also increasing disease resistance [41,42,43]. Its broad impact has led to the recognition of IPA1 as a potential new “Green Revolution” gene for rice improvement [44].
Due to its pleiotropic roles across tissues and developmental stages, hormone signaling, and responsiveness to environmental stresses, SPL9 represents a key regulatory node in plant biology. Research on SPL9 not only deepens our understanding of fundamental plant developmental mechanisms but also provides valuable insights for crop improvement. In this text, we summarize the current literature on expression regulation, biological function and stress role of SPL9 as well as its regulatory networks. And we discuss the potential applications of SPL9 in future crop breeding programs.
2. Coordinated Multilayer Regulation of SPL9 Expression and Activity
As a key transcription factor involved in plant growth and development, SPL9 is expressed in multiple tissues, including roots, stems, leaves, and floral organs. Based on the AtGenExpress expression data, AtSPL9 exhibits high expression in the shoot apex during inflorescence and transition, as well as in early floral organs and carpels; moderate expression is observed throughout the rosette following the transition to flowering; it is also expressed in the seedling hypocotyl, roots and various other organs [45]. The functional output of SPL9 is governed by a sophisticated, multi-layered regulatory mechanism. It integrates transcriptional control, post-transcriptional fine-tuning by miR156, and post-translational regulation of protein activity and stability. In detail, transcriptional regulation defines the potential for SPL9 production, miR156-mediated post-transcriptional regulation quantitatively limits the translatable mRNA pool, and post-translational mechanisms act as a final functional valve, determining the activity, specificity, and stability of the SPL9 protein (Figure 1).
2.1. Transcriptional Regulation of SPL9 Expression
Regulations at the transcriptional level of SPL9 are mediated by specific transcription factors and epigenetic mechanisms at key developmental transitions. These regulated changes in SPL9 transcriptional levels establish the foundation for any subsequent downstream regulation. For instance, during the vegetative phase transition, MYB33 directly binds to a specific region (−585 to −690 bp) of the AtSPL9 promoter and enhances its transcription [46]. This transcriptional upregulation increases the substrate mRNA pool that is subject to subsequent fine-tuning by miR156 at the post-transcriptional level. During sucrose-induced juvenile-to-adult transition, PAP1 activates AtSPL9 expression by directly binding to the MYB-binding motif (MYB-BM) in its promoter sequence in Arabidopsis, which occurs in conjunction with glucose-mediated repression of MIR156 [47].
In rice, a 54 bp deletion in the promoter of IPA1 (a rice ortholog of AtSPL9) leads to increased transcriptional activity and improved agronomic traits, including increased panicle number and size [48]. This deletion removes An-1 binding to the GCGCGTGT cis-element and frees IPA1 expression from An-1 mediated repression, which increases the functional protein that regulates downstream target genes and thus improves key traits [48]. Whereas in alleles with an intact An-1 binding site, IPA1 transcription is suppressed. This results in robust vegetative growth similar to high miR156 conditions [39]. In addition, epigenetic mechanisms also influence the transcriptional control of SPL9. For example, high expression of IPA1 is associated with DNA hypomethylation at GHH-type cytosine sites within the 800 bp genomic region upstream of the ATG start codon in rice [42]. This indicates that stable epigenetic modifications can influence the basal transcription level of SPL9 orthologs.
2.2. Post-Transcriptional Regulation of SPL9 by miR156
Based on transcriptional control, the conserved miR156 determines SPL9 protein accumulation by targeting and degrading SPL9 mRNA. SPL9 mRNA contains the miR156 response element in the 3′ untranslated region (UTR), making it susceptible to miR156-guided RNA-induced silencing complex (RISC) [8,25]. Base-pairing with its target mRNA guides the RNA-induced silencing complex (RISC) to direct AGO protein-mediated transcript cleavage and degradation [49]. Even without complete degradation, the RISC bound to SPL9 transcripts blocks their translation, forming a translational barrier to limit the functional SPL9 protein production. It thus provides an important upper limit on SPL9 protein accumulation.
The expression of MIR156 genes is also precisely controlled in order to provide the signal integration node. The abundance of miR156 is dynamically regulated by a series of internal and external cues, which in turn fine-tune the miR156-mediated post-transcriptional repression of SPL9 across diverse developmental programs and stress responses [50]. In the context of the vegetative phase change, miR156 is not only regulated by developmental age but is also profoundly influenced by metabolic signals. For instance, glucose represses miR156 transcription and facilitates the degradation of its primary transcripts through a pathway involving Hexokinase 1 (HXK1) [47]. The age-dependent decline in miR156 is also reinforced by epigenetic modifications. Complexes such as PRC1, PRC2 and the PKL-HDA9 chromatin remodeling machinery establish a repressive chromatin state at specific loci such as MIR156A and MIR156C, thereby ensuring the irreversibility of the transition from juvenile to adult phase [51].
Precise regulation by miR156 fine-tunes the abundance of a suite of downstream SPL transcription factors. In Arabidopsis, at least ten SPL genes are validated targets of miR156, which can be functionally classified according to their roles in different developmental stages. For example, SPL9, SPL10, SPL13 and SPL15 regulate the vegetative phase change. Although functional redundancy exists among these members, SPL9 exhibits a uniquely broad and central role. It expands its functional scope by directly targeting and regulating key genes responsible for adult traits. During leaf development, SPL9 directly binds to the promoters of leaf morphology regulators such as BOP1, BOP2, and CYCD3 to control their expression [30]. While the SPL family collectively serves as an integrator of flowering signals, SPL9 adopts a distinct strategy compared to other members such as SPL3, SPL4, and SPL5, which directly activate flowering integrator genes [31], and functions as a developmental timer by activating the expression of miR172, which regulates downstream targets to flower [52]. Similarly, in the control of axillary bud outgrowth and branching, although SPL9 and SPL15 show redundant functions in certain light signaling pathways, only SPL9 has been demonstrated to uniquely integrate the miR156 module with the gibberellic acid (GA) signaling pathway to fine-tune branching architecture [29,37,53]. Collectively, these findings position SPL9 as a leader among the miR156-targeted SPLs, playing an indispensable and overarching role in multiple core developmental processes. Subsequent chapters will address how SPL9 regulates its target genes and the physiological mechanisms in plant development.
2.3. Protein Interaction and Post-Translational Modifications
The functional output of SPL9 is ultimately regulated by protein–protein interactions and post-translational modifications (PTMs). SPL9 transcriptional activity can be suppressed through direct or indirect suppression of protein–protein interactions, which maintain SPL9 in an inactive state until appropriate signals are perceived. DELLA proteins, plant-specific regulatory proteins lacking a DNA-binding domain, act as growth repressors by interacting with other TFs, including SPL9 [54,55,56]. They are usually bound to the C-terminal region of AtSPL9, which inhibits its ability to activate downstream genes in response to gibberellin signaling [57]. The critical link to upstream regulation is that GA signaling triggers DELLA degradation, thereby releasing the pre-existing suppressed SPL9 protein [37]. Other repressors such as DEWAX and strigolactone signaling components D53/SMXLs directly interact with SPL9 in the form of non-functional complexes, storing the TF and preventing target gene activation [29,58].
Post-translational modifications (phosphorylation and ubiquitination) provide a rapid and reversible means to fine-tune the activity, stability, and functional specificity of the SPL9 protein pool. Phosphorylation can inhibit or redirect SPL9 function. For example, BIN2 kinase phosphorylates AtSPL9 to suppress its role in the vegetative phase transition [7]. Pathogen-induced phosphorylation of IPA1 at Ser163 alters its DNA-binding specificity, enabling it to activate the immunity gene WRKY45 instead of its growth targets [59]. Ubiquitination also regulates SPL9 abundance, which is essential for maintaining a balance between growth and stress resistance [60,61,62]. The RING-finger E3 ligase IPA1 INTERACTING PROTEIN1 (IPI1) mediates the tissue-specific ubiquitination of IPA1 by means of different polyubiquitin chains, thereby modulating IPA1 complex stability and maintaining appropriate protein levels across tissues [62].
3. The Diverse Role of SPL9 in Plant Growth and Development
Plant growth and development are highly ordered and regulated processes involving numerous regulatory mechanisms and complex signaling networks. Among the main regulators, SPL9, a plant-specific TF targeted by miR156, forms a core regulatory module that integrates diverse internal and external cues to coordinate various developmental programs. SPL9 functions in each of these developmental pathways are determined by multilevel regulatory mechanisms that limit its expression and activity. In this section, we highlight the indispensable roles of SPL9 across multiple stages of development covering both below-ground and above-ground traits. Above-ground traits are subdivided into juvenile-to-adult phase change, shoot branching and tillering, and age-dependent flowering. Table 1 summarizes the main mechanisms for SPL9 activity, along with its key targets and their functions across these physiological processes.
3.1. SPL9 Regulates Lateral Root Development and Nodulation
The root system is essential for absorbing water and nutrients as well as for sensing changes in the soil environment. Lateral roots, as an important component of root architecture, originate from founder cells within the pericycle, located opposite the xylem poles. Following several rounds of division, these cells give rise to lateral root primordia from the primary root [28,69,70], which are regulated by hormones, miRNAs, and TFs [64,70,71,72]. The conserved miR156-SPL module acts antagonistically, in which miR156 promotes lateral root formation, while SPL9 and its homologs suppress it [63]. Both miR156 and its targets (SPL9 and SPL10) are responsive to auxin signals, with SPL9/SPL10 mediating mir156 levels to create feedback loops that stabilize lateral root development and integrate auxin into miR156/SPL module in Arabidopsis (Figure 2). The evolutionary conservation of SPL9’s role in lateral root development is illustrated by the influence of the miR156-StSPL9 module on both plant height and root architecture in potato [71].
In addition to developmental regulation, SPL9 mediates environmental responses in roots, such as symbiotic nodulation in legumes under low-nitrogen conditions or rhizobial infection (Figure 2). In soybean, the miR156b-GmSPL9d module is involved in nodulation. GmSPL9 is downstream of rhizobia-suppressed miR156, leading to increased protein abundance. GmSPL9d binds to the promoters of miR172 and nodulation markers such as GmNINa and GmENOD40-1, thereby integrating nodulation signaling with root development [64]. However, excessive nodulation is harmful because of the maintenance of nodules consumes considerable energy. In Medicago sativa (alfalfa), MsSPL9 suppresses nodulation in response to nitrate signals. Silencing MsSPL9 can alleviate nitrate stress and disrupt the expression of nitrate-responsive and nodulation genes [73].
3.2. SPL9 Modulates Vegetative Phase Change
Postembryonic ontogeny is characterized by distinct phase transitions, with the juvenile-to-adult vegetative shift being crucial for growth and reproduction. miR156 is a key regulator of this process, and its target gene, SPL9, plays a crucial role [26,74]. The level of miR156 is high in the juvenile phase and decreases with increasing age, while the expression of SPL9 increases. AtSPL9 promotes the transcription of miR172b and accelerates the transition to the adult phase [65]. A nonsynonymous mutation in the nuclear localization signal (NLS) of the SBP domain in SPL9 delays the vegetative phase change in Arabidopsis [75]. Similarly, the loss-of-function mutation dwf5 results in a delayed phase transition due to decreased AtSPL9 and miR172 levels and increased accumulation of TARGET OF EAT1 (TOE1) [7]. BIN2, a GSK3-like kinase that negatively regulates BR signaling [76,77,78], interacts with and phosphorylates both AtSPL9 and TOE1, promoting their degradation. Reduced BR levels increase BIN2 activity, destabilize AtSPL9 and TOE1, downregulate miR172, and delay phase transition [7]. SPL9 functions as a central integrator within the miR156-SPL9-miR172-TOE1 module, linking BR signaling to developmental timing (Figure 3).
The transition to adult vegetative growth in plants is often accompanied by heteroblastic changes in leaf morphology and arrangement [79,80]. SPL9 plays a key role in this process [26,81]. Together with its homolog AtSPL13, AtSPL9 promotes leaf blade elongation, resulting in larger, more elongated leaves [30]. They bind directly to the promoter regions of BOP1 and BOP2 to suppress their expression, which prolongs blade development while retarding petiole development in Arabidopsis [30] (Figure 3). Although AtSPL9 and AtSPL13 share certain binding sites within the genomic regions of BOP1 and BOP2, they also display distinct binding profiles, which may account for the shared and unique phenotypic traits of rSPL9 and rSPL13 in Arabidopsis.
Notably, AtSPL9 is absent in juvenile leaves, which have small and round blades distinctly separated from the basal petiole. However, SPL9 is expressed in adult leaves, which are larger, oval-shaped, and feature a more gradual transition between the blade and petiole in Arabidopsis [27]. Cytologically, AtSPL9 prolongs leaf morphogenesis by maintaining cell proliferation and delaying the transition to endoreduplication and differentiation [27]. AtSPL9 directly targets CYCD3 genes to regulate the cell cycle, thus influencing leaf heteroblasty [27] (Figure 3).
3.3. SPL9 Controls Branching and Tillering
SPL9 plays a significant role in regulating the development of lateral organs, such as branches in Arabidopsis and tillers in rice (Figure 4). In Arabidopsis, SPL9 fine-tunes branching by integrating age, hormone, and light signals. Under shade conditions, the accumulation of phytochrome-interacting factors (PIFs) directly binds to the MIR156 promoter to suppress its expression [53]. This suppression reduces the post-transcriptional regulation of miR156 on its target genes. Light and hormone signaling converge on AtSPL9 and AtSPL15, with both light-sensitive factors FHY3/FAR1 and strigolactone-associated SMXL6/7/8 physically interacting to inhibit their transcriptional activation of BRC1 [29]. Reduced FHY3 and FAR1 release SPL9/SPL15 from repression by FHY3/FAR1 and SMXL6/7/8, enabling them to activate BRC1 expression and suppress branching. Furthermore, PIF4 physically interacts with AtSPL9 to activate BRC1 and modulate branching [82]. Beyond its overlapping function with SPL15, SPL9 plays a distinct role in modulating axillary bud formation. It directly represses LATERAL SUPPRESSOR (LAS) by binding to its promoter, thereby inhibiting axillary bud formation [37]. This repression is counteracted by regulatory inputs from the gibberellin signaling pathway. DELLA proteins, key components of gibberellin signaling, interact with AtSPL9, diminishing its repressive effects on LAS and promoting axillary bud formation. This process operates through a GA-responsive feedback loop involving the DELLA-SPL9-LAS-GA2ox4 regulatory module [37].
IPA1 in rice regulates tillering and yield, primarily through its involvement in the strigolactone (SL) signaling pathway. In this pathway, IPA1 and D53 (a known SL signaling repressor) form a feedback loop [58,83]. By binding to the D53 promoter, IPA1 activates its expression, while the D53 protein interacts with IPA1 to inhibit its transcriptional activity. This interaction is modulated by SL perception, which induces D53 degradation and consequently releases IPA1 to suppress tillering [58]. Additionally, IPA1 directly activates OsTB1, a gene from the TCP (TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR) family that inhibits axillary bud outgrowth, and binds to the promoter of DEP1 (DENSE AND ERECT PANICLE 1), influencing both panicle architecture and grain yield [41,66]. Thus, IPA1 reduces tillering, increases plant height, and enhances panicle branching, contributing to an ideal plant architecture. Moreover, IPA1 recognizes novel cis-elements, such as the TGGGCC/T motif, through its interaction with PCF1 and PCF2, thereby expanding its regulatory targets [41].
3.4. SPL9 Roles in the Age-Dependent Flowering Pathway
The age-dependent flowering pathway relies primarily on intrinsic developmental signals, rather than external environmental cues [31,84]. The miR156-targeted SPL gene module regulates the vegetative-to-reproductive transition and flowering time by coordinating the expression of flowering-related genes across the age gradient [31,74,84,85]. As a key target of miR156, SPL9 has highly conserved flowering regulatory functions across multiple species [85,86,87]. In Arabidopsis, the levels of AtSPL9 and AtSPL3 in the shoot apices increase with age. The overexpression of these genes accelerates flowering, whereas elevated miR156 delays flowering by suppressing SPL activity [74,81,88]. Increased SPL9 levels promote flowering by directly binding to the GTAC core motif in the regulatory regions of positive floral regulators, including FUL, SOC1, AGL24 and AGL42, activating their expression [31,32]. Concurrently, SPL9 acts upstream of miR172, directly promoting its transcription. The upregulation of miR172 establishes a crucial indirect route for SPL9 to regulate flowering. miR172 in turn post-transcriptionally represses a suite of its own target genes, which encode AP2-like transcriptional repressors of flowering (e.g., TOE1, TOE2, TOE3, SMZ, SNZ) [52]. The suppression of these repressors further facilitates the floral transition. Thus, through this coordinated miR156-SPL9-miR172 regulatory module, SPL9 integrates direct gene activation with indirect de-repression to ensure a timely and precise onset of flowering (Figure 4).
3.5. Additional Developmental Functions of SPL9
In addition to its well-established roles in vegetative and reproductive development, SPL9 plays critical regulatory roles in seemingly disparate processes such as trichome initiation and cuticular wax biosynthesis.
In Arabidopsis, trichome initiation and development are regulated by positive regulators (e.g., TTG1, GL1, GL3, EGL3) [89,90,91,92] and by negative regulators from the R3 MYB factors (e.g., TRY, TCL1/2) [93,94,95,96,97,98]. AtSPL9 suppresses trichome initiation by activating the expression of TRY and TCL1 [99]. This pathway functions independently of the GIS-dependent trichome pathway [100,101]. LOM (LOST MERISTEMS) proteins (LOM1/2/3), which are targeted by timing miR171, interact with AtSPL9 to promote trichome formation and delay flowering [67]. In addition, TOE1, a downstream target of the miR156-SPL9 module, represses GL1 expression, thereby inhibiting trichome development [50].
SPL9 also promotes cuticular wax deposition in Arabidopsis via the DEWAX/SPL9-CER1 [68,102]. AtSPL9 expression follows a circadian rhythm, peaking during the light period and declining in darkness [68]. Under light, SPL9 directly influences the expression of wax biosynthesis genes, such as ECERIFERUM1 (CER1), and indirectly affects CER4 [68]. At night, DEWAX protein accumulates and directly interacts with SPL9, inhibiting its DNA-binding activity and suppressing wax synthesis [68]. Two MYB-SHAQKYF TFs, MYS1 and MYS2, function upstream of the DEWAX-SPL9 module, repressing DEWAX expression while promoting CER1 expression [102]. Although SPL9 and SPL13 play redundant roles in regulating wax biosynthesis under drought conditions, SPL13 is not involved in the DEWAX-mediated diurnal control of wax production [34].
4. The Multifaceted Regulatory Roles of SPL9 in Plant Stress Responses
Plants are constantly exposed to a wide array of environmental stresses that threaten their growth, development, and survival. Recent studies have suggested that SPL9 is a multifunctional modulator, coordinating plant responses to both biotic and abiotic stresses. Central to this regulation is the involvement of the miR156-SPL9 module, which integrates pathways related to secondary metabolite production and phytohormone signaling (see Table 2). The following sections summarize the roles of SPL9 in mitigating abiotic stresses, such as drought and temperature extremes, as well as biotic stresses, including pathogen infections and insect herbivory. We focus on the dual regulatory function of SPL9 as a transcriptional activator and repressor to fine-tune defense responses while balancing the growth-defense trade-offs.
4.1. Drought Stress
In response to drought stress, SPL9 negatively modulates antioxidant defense mechanisms and positively stimulates cuticular wax biosynthesis (Figure 5). One of its key functions is regulating anthocyanin synthesis in response to drought stress [103,106,107]. Anthocyanins are strong antioxidants, and their synthesis follows a pathway with the enzyme DIHYDROFLAVONOL-4-REDUCTASE (DFR) as the core. The expression of DFR is controlled by the MYB-bHLH-WD40 complex [103]. AtSPL9 interacts with PAP1, a core MYB component of the MYB–bHLH–WD40 complex, and negatively regulates DFR expression, thereby suppressing anthocyanin synthesis. This competitive interaction prevents PAP1 binding to TT8 (a bHLH factor), thereby destabilizing the transcriptional activation complex and reducing DFR expression. Thus, SPL9 fine-tunes anthocyanin synthesis, resulting in the negative regulation of drought stress responses. The contribution of SPL9 to drought tolerance through its antioxidant defense mechanism has been evaluated in some species. In cassava, MeSPL9-RNAi plants have high anthocyanin levels with improved drought resistance [108]. In rice, IPA1 positively regulates drought tolerance by activating the expression of SNAC1 [109], an NAC TF that plays a key role in reactive oxygen species (ROS) homeostasis [110]. This mechanism enhances drought resistance by promoting ROS scavenging.
SPL9 also acts as a positive regulator of cuticular wax deposition, which forms a critical barrier against non-stomatal water loss. Here, SPL9 functions downstream in the MYS1/MYS2-DEWAX-SPL9 module. The upstream regulators MYS1 and MYS2 are likely responsible for activating or stabilizing SPL9, which in turn promotes the expression of wax biosynthesis genes [102] (Figure 5). Overexpression of AtSPL9 rescues the drought hypersensitivity of mys1c mys2 mutants. Both AtSPL9 and AtSPL13 are involved in cuticular wax biosynthesis under drought stress [34]. Plants overexpressing AtSPL9 or AtSPL13 presented reduced transpiration rates, whereas single and double mutants showed increased water loss.
4.2. Temperature Stress
Extreme temperatures, both heat and cold, are major stressors that impact plant growth and development. miR156 expression is induced by high temperature and serves as a key regulator by targeting SPLs to mediate thermal responses. Among these targets, SPL9 has also been identified as a key member that mediates thermal responses [111,112] (Figure 5). The essential role of miR156 in thermomorphogenesis is demonstrated by the reduced hypocotyl elongation observed in miR156-knockdown (MiM156) plants [104]. Conversely, elevated AtSPL9 inhibits hypocotyl elongation in both MiM156 and rSPL9-overexpressing lines. The miR156-SPL9 module acts antagonistically to regulate hypocotyl elongation during thermomorphogenesis, enabling plants to adapt their growth patterns through the modulation of auxin sensitivity in Arabidopsis [104].
SPL9 also plays a vital role in cold stress responses (Figure 5). SPL9 expression is induced by low temperatures across various species [33,113]. Despite a concurrent increase in miR156 expression under cold stress, the induction of SPL9 appears to operate through a pathway that is independent of miR156-mediated targeting [33]. SPL9 cooperates with other TFs to enhance cold tolerance. Notably, SPL9 may bind to the promoter of CBF2, a member of the C-REPEAT-BINDING FACTOR/DRE-BINDING FACTOR1 (CBF/DREB1) family, to activate its expression [33,114]. In rice, the OsSAPK6-IPA1-OsCBF cascade is activated in response to cold stress [105]. IPA1 is phosphorylated by OsSAPK6 at Ser213, which stabilizes IPA1 and promotes cold adaptation. Furthermore, IPA1 activates OsCBF3 expression by binding to the GTAC core motif within its promoter, confirming that SPL9 is a regulator of cold tolerance.
4.3. Biotic Stress
SPL9 is also implicated in plant immunity against pathogens and insects (Figure 5). For instance, AtSPL9 promotes ROS accumulation and increases the expression of SA-responsive genes, including SA-induced pathogenesis-related (PR) genes PR1 and PR2 [35]. The miR156-SPL9 module further regulates phytohormone-responsive genes to trigger innate immunity [35,36]. Salicylic acid (SA) functions as a central defense hormone that activates immune responses against bacterial pathogens [115]. Arabidopsis plants in which miR156 was suppressed or SPL9 was overexpressed showed milder symptoms upon infection with Psedomonas. syringae pv. tomato DC3000, along with the upregulation of SA-dependent defense genes [35]. The role of SPL9 in biotic stress responses is notably context-dependent, varying with the type of pathogen involved [35,36]. During infection by the necrotrophic fungus Botrytis cinerea, the miR156-SPL9 module links the function of SQUINT (SQU), which is an ortholog of the immunophilin cyclophilin 40 (Cyp40), with jasmonate (JA) signaling in Arabidopsis [36]. SQU promotes miR156 accumulation to repress SPL9, thereby relieving the SPL9-mediated suppression of JA signaling [36].
Under insect attack, SPL9 functions as a negative regulator of JA-mediated defense responses [36,108]. AtSPL9 directly suppresses the JA signaling pathway, diminishing the plant’s resistance to insect attack [6]. Experimental evidence demonstrates that overexpression of miR156 represses SPL9, leading to the elevated expression of JA-responsive genes such as LOX2 and VSP2 [6]. These findings further underscore SPL9’s role as a suppressor of JA-dependent defense. Specifically, SPL9 interacts with JA ZIM-domain (JAZ) proteins, promoting and stabilizing their accumulation, which represses JA signaling and weakens JA-dependent insect resistance [6].
5. Conclusions and Perspectives
SPL9 is a key transcription factor involved in regulating plant growth, developmental phase transitions, and biotic and abiotic stress responses. Its expression is tightly controlled by miR156, forming a conserved miR156-SPL9 regulatory module across multiple crop species [39,64,71]. SPL9 acts as a transcriptional regulatory hub in complex networks, interacting with multiple genes and pathways. It primarily modulates downstream targets by binding to the GTAC core motif and/or TGGGCC/T motif, as demonstrated in the regulation of age-dependent flowering [31,32]. The transcriptional activity of SPL9 is dynamically influenced by developmental timing, environmental cues, and hormone signaling. For instance, during inflorescence stem branching in Arabidopsis, AtSPL9 expression is modulated by age-dependent pathways, light conditions, and hormonal inputs, highlighting its integrative role in multiple regulatory signals [29,37]. However, the molecular mechanisms involved in cross-talk between SPL9 and these pathways remain poorly understood.
We summarize the existing knowledge on SPL9 in terms of vegetative growth, reproductive development, transitions of developmental stages, and stress responses. By fine-tuning its spatiotemporal expression, SPL9 selectively regulates downstream genes to direct organ formation at precise stages and locations, a mechanism critical for achieving optimal plant architecture. SPL9 also serves as a mediator and regulator of stress responses by modulating secondary metabolic and phytohormone pathways. Currently, it is still not fully understood how SPL9 might serve as a convergence point for developmental and environmental signals to regulate the trade-off between growth and defense.
Research on SPL9 has been extensively conducted in model species such as Arabidopsis thaliana and rice, but the orthologous genes in other crops are largely unexplored (Table S1). Future research should focus on elucidating the conserved and divergent functions of SPL9 orthologous genes, as well as revealing the molecular mechanisms that govern their spatiotemporal expression and integration of developmental and environmental signals. Simultaneously, modern biotechnological tools such as CRISPR-based genome editing, high-throughput phenotyping, and systems-level omics are used to target the regulation of SPL9 and analyze regulatory networks, and there is a potential to optimize agronomic traits in molecular design breeding. These efforts lay the foundation for the regulatory networks mediated by SPL9 and also create practical strategies for improving agricultural productivity and enhancing stress resistance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Long Y.C. Scheres B. Blilou I. The logic of communication: Roles for mobile transcription factors in plants J. Exp. Bot.2015661133114410.1093/jxb/eru 54825635110 · doi ↗ · pubmed ↗
- 2Han X. Kumar D. Chen H. Wu S. Kim J.Y. Transcription factor-mediated cell-to-cell signalling in plants J. Exp. Bot.2014651737174910.1093/jxb/ert 42224347464 · doi ↗ · pubmed ↗
- 3Giaume F. Fornara F. SPL transcription factors prevent inflorescence reversion in rice Mol. Plant 2021141041104310.1016/j.molp.2021.06.00934133977 · doi ↗ · pubmed ↗
- 4Yang B.J. Sun Y.B. Minne M. Ge Y.H. Yue Q.R. Goossens V. Mor E. Callebaut B. Bevernaege K. Winne J.M. SPL 13 controls a root apical meristem phase change by triggering oriented cell divisions Science 2024386 eado 429810.1126/science.ado 429839541454 PMC 7616863 · doi ↗ · pubmed ↗
- 5Wang L. Zhang Q. Boosting Rice Yield by fine-tuning SPL gene expression Trends Plant Sci.20172264364610.1016/j.tplants.2017.06.00428647102 · doi ↗ · pubmed ↗
- 6Mao Y.B. Liu Y.Q. Chen D.Y. Chen F.Y. Fang X. Hong G.J. Wang L.J. Wang J.W. Chen X.Y. Jasmonate response decay and defense metabolite accumulation contributes to age-regulated dynamics of plant insect resistance Nat. Commun.201781392510.1038/ncomms 1392528067238 PMC 5233801 · doi ↗ · pubmed ↗
- 7Zhou B.Y. Luo Q. Shen Y.H. Wei L. Song X. Liao H.Q. Ni L. Shen T. Du X.L. Han J.Y. Coordinated regulation of vegetative phase change by brassinosteroids and the age pathway in Arabidopsis Nat. Commun.202314260810.1038/s 41467-023-38207-z 37147280 PMC 10163027 · doi ↗ · pubmed ↗
- 8Jerome Jeyakumar J.M. Ali A. Wang W.M. Thiruvengadam M. Characterizing the role of the mi R 156-SPL network in plant development and stress response Plants 20209120610.3390/plants 909120632942558 PMC 7570127 · doi ↗ · pubmed ↗