A simplified co-culture reveals altered cardiotoxic responses to doxorubicin in hPSC-derived cardiomyocytes in the presence of endothelial cells
Marcella Brescia, James Gallant, Andrea Chatrian, Paul Keselman, Elsa Sörman Paulsson, Mervyn P.H. Mol, Rickard Sjögren, Karine Raymond, Valeria Orlova, Kalpana Barnes, Richard Wales, Jonas Austerjost, Michael W. Olszowy, Christine L. Mummery, Berend J. van Meer, Richard P. Davis

TL;DR
This study shows that endothelial cells respond early to doxorubicin and increase heart cell death, suggesting new ways to test drug toxicity.
Contribution
The study introduces a machine learning-based method to monitor cardiotoxicity in co-cultures without labels.
Findings
Endothelial cells are more sensitive to doxorubicin than cardiomyocytes or fibroblasts.
Co-culture with endothelial cells increases doxorubicin-induced apoptosis in cardiomyocytes.
Inhibiting nitric oxide signaling reduces caspase activity in co-cultures.
Abstract
Cardiotoxicity is a significant challenge in cancer therapies, particularly with doxorubicin, a widely used anthracycline. More predictive in vitro models are needed to understand doxorubicin-induced cardiac damage and patient-specific responses. Here, human pluripotent stem cell (hPSC)-derived cardiomyocytes (hPSC-CMs), cardiac fibroblasts (hPSC-cFBs), and endothelial cells (hPSC-ECs) were cultured in mono- or multi-cell-type formats and repeatedly treated with doxorubicin to mimic cumulative clinical exposure. A machine learning-based tool enabled continuous quantification of the early toxicity marker caspase-3/7 and accurately identified hPSC-CMs within mixed cultures. Notably, hPSC-ECs were more sensitive to doxorubicin than hPSC-CMs or hPSC-cFBs, with nitric oxide signaling contributing to the elevated cardiomyocyte toxicity observed in co-culture. These results question the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
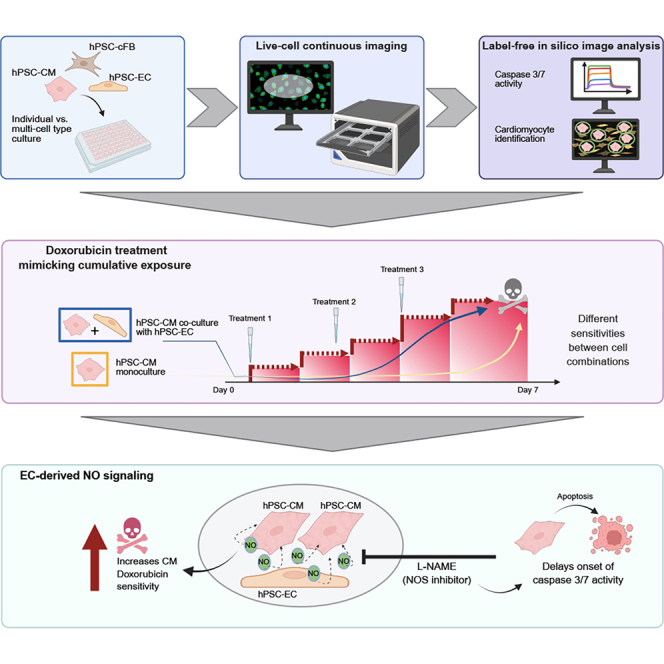 Figure 1
Figure 1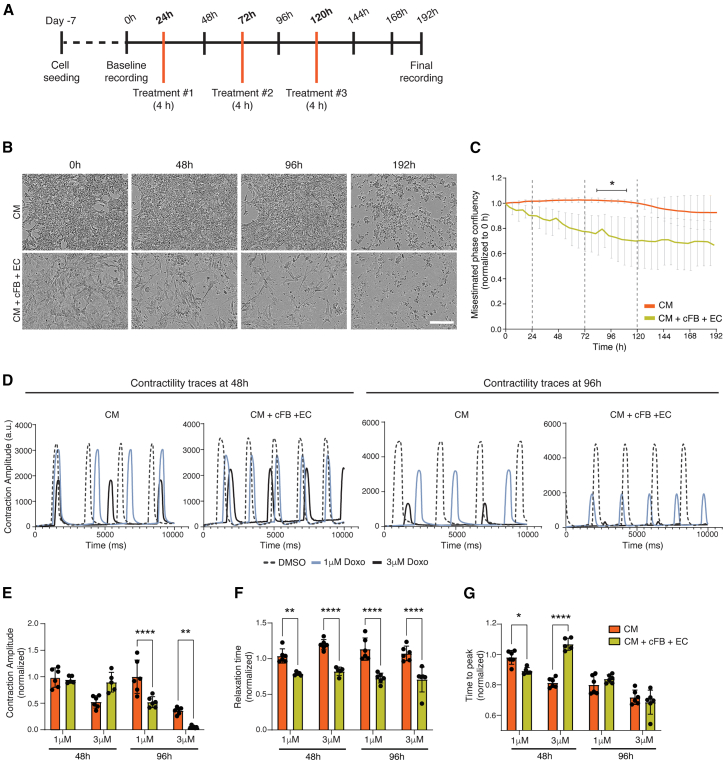 Figure 2
Figure 2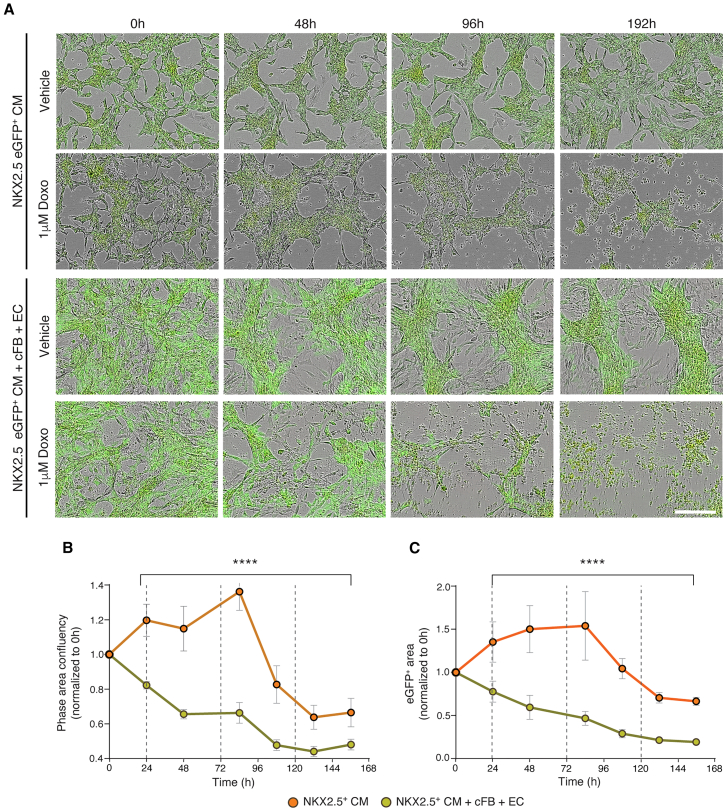 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5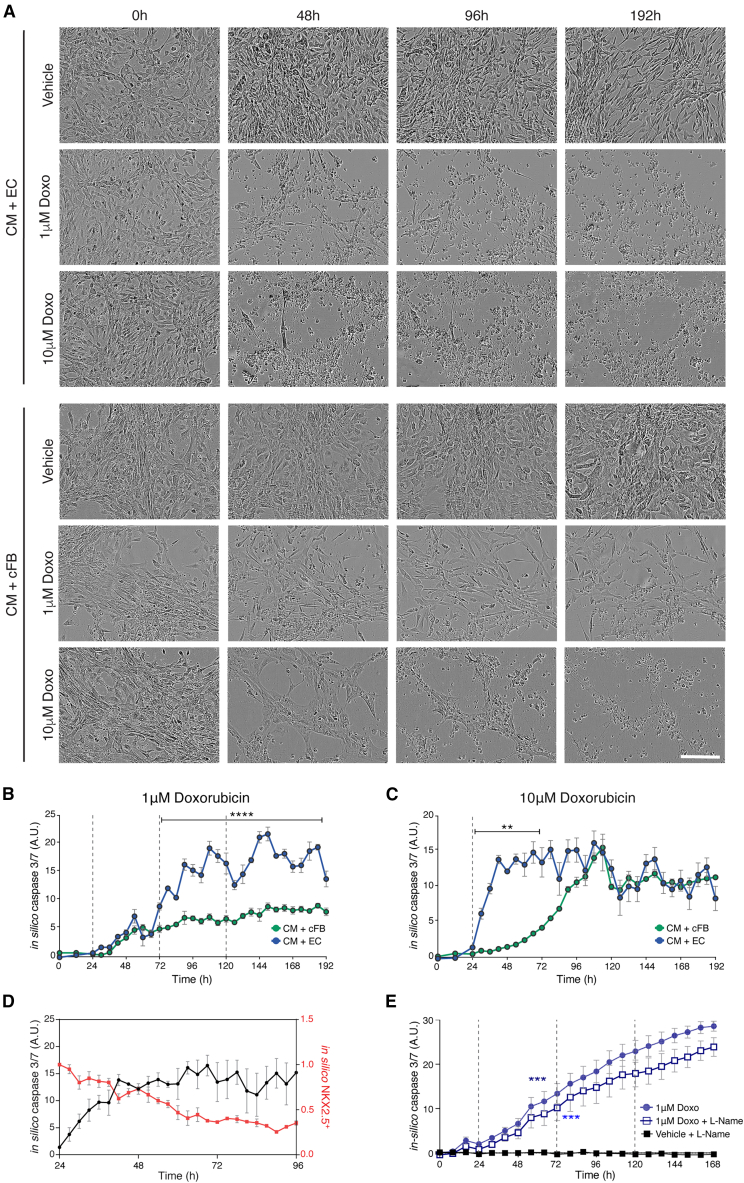 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsChemotherapy-induced cardiotoxicity and mitigation · Cardiac electrophysiology and arrhythmias · Congenital heart defects research
Introduction
Predicting patient-specific drug responses and adverse reactions remains a significant challenge in healthcare, particularly in the context of cardiotoxicity. This is most notable with chemotherapeutics, which can have well-known life-threatening side effects with late heart failure evident in up to 10% of patients (van der Pal et al., 2012). The variability in drug efficacy and safety, even among patients with identical diagnoses, complicates the prediction of such adverse effects (Schwach et al., 2024). The heart is one of several organs for which regulatory authorities mandate comprehensive cardiotoxicity assessments throughout the drug development process (Kettenhofen and Bohlen, 2008). Conventionally, these assays have predominantly relied on ex vivo assays and in vivo studies in animals. However, the physiological and anatomical differences between humans and most animal models often limit the translatability of such studies, contributing to high failure rates (∼90%) of drug candidates in clinical trials due to unforeseen toxicities and lack of efficacy (Mak et al., 2014; Zaragoza et al., 2011). Contemporary regulatory initiatives, exemplified by the FDA Modernization Act 2.0, are driving a paradigm shift away from reliance on animal models toward the adoption of cell-based assays and advanced data analysis methodologies (Zushin et al., 2023). Human pluripotent stem cells (hPSCs) are emerging as a promising alternative, potentially offering more predictive models for evaluating drug-induced cardiotoxic effects and reducing reliance on animals (Saleem et al., 2020). Moreover, these cells are particularly valuable for personalized screening and improving understanding of why individual patients respond differently to cardiotoxic compounds (de Korte et al., 2020).
Doxorubicin (Doxo), a widely used anthracycline, exemplifies the delicate balance between therapeutic efficacy and cardiotoxic risk. Despite its proven effectiveness in treating a variety of cancers, including acute leukemia, lymphomas, and various solid tumors in both adults and children, it is also associated with significant adverse effects, most notably cardiotoxicity (Linders et al., 2024). This cardiotoxic effect, characterized by a spectrum of cardiac dysfunctions such as congestive heart failure, arrhythmias, and reduced left ventricular ejection fraction (Csapo and Lazar, 2014), manifests in a cumulative and dose-dependent manner and severely limits its repeated or long-term clinical use. The mechanisms involved are complex and not fully understood, but include mitochondrial dysfunction, calcium overload, DNA damage, and apoptosis following caspase activation (Karabulut et al., 2021; Michihiko et al., 2006; Rawat et al., 2021).
hPSC-derived cardiomyocytes (hPSC-CMs) have become a mainstream tool for cardiotoxicity testing by both the pharmaceutical industry and academia, owing to their scalability and suitability for high-throughput safety assessments (Raniga et al., 2024; Stebbeds et al., 2023). However, despite their widespread adoption, these models still present limitations in accurately predicting clinical outcomes. Increasing their complexity by incorporating additional hPSC-derived cardiac cell types such as endothelial cells (ECs) and cardiac fibroblasts (cFBs), has been shown to improve their physiological relevance and consequently, their clinical translatability (Raniga et al., 2024). This improvement is particularly evident in 3D cultures, for example, microtissues or engineered heart tissues (EHTs), which promote not only the maturation of hPSC-CMs but also the predictiveness of drug responses (Giacomelli et al., 2020; Saleem et al., 2020), including the effects of Doxo on contraction dynamics (Qiao et al., 2020; Schwach et al., 2024).
However, it currently remains challenging to investigate cell type-specific responses in three-dimensional (3D) models, particularly in real time. In this context, we investigated the potential of two-dimensional (2D) multi-cell-type cultures to offer a more predictive model than 2D hPSC-CM monocultures, while remaining more accessible than 3D cardiac models for both drug exposure and imaging. To analyze the dynamics of cardiotoxicity over prolonged and repeated exposures to Doxo and quantitatively compare the effects between different cell types, we applied a machine learning (ML)-based analysis tool to assess caspase-3/7 activity as a marker of early toxicity. This in silico tool could also identify hPSC-CMs in multi-cell-type cultures, allowing us to track cardiomyocyte apoptosis in different cellular environments over time. Our findings revealed that the presence of specific cell types, notably hPSC-ECs, influenced the sensitivity of hPSC-CMs to Doxo and may be a key driver of cardiomyocyte toxicity. This study not only sheds light on Doxo-induced cardiotoxic mechanisms but also underscores the potential of AI tools to advance high-throughput analysis and personalized drug screening, paving the way for safer therapeutic interventions.
Results
hPSC-CMs co-cultured with other cardiac cell types appear more sensitive to Doxo
To emulate the in vivo pharmacodynamics of Doxo (Barpe et al., 2010; Pang et al., 2013), an in vitro treatment protocol was used in which the hPSC-derived cells were exposed to Doxo for 4-h intervals every 48 h (Figure 1A). Because the free diffusible Doxo concentration in vivo varies between 20 nM and 2 μM (Greene et al., 1983), we examined a range of concentrations (0.01–10 μM), with representative phase contrast images and analysis presented in Figure S1. We selected 1 μM Doxo as a proxy for the cardiotoxic effect seen in vivo since this concentration induced cumulative toxicity without immediate cell death in hiPSC-CMs.Figure 1. Cumulative Doxo treatment effect on hPSC-derived cardiac cell cultures(A) Schematic of treatment protocol timeline, with orange lines marking the time points of 4-h Doxo application to the cells.(B) Representative phase contrast images of monoculture hiPSC-CMs (top) or co-cultured with hiPSC-cFBs and -ECs (bottom) treated with 1 μM Doxo. Images were acquired at time points corresponding to baseline (0 h), 24 h after treatments 1 and 2 (48 and 96 h, respectively), and the final time point (192 h). Scale bar, 100 μm.(C) Quantification of phase confluency area calculated from live cell imaging, and normalized to baseline (0 h), for conditions shown in (B), highlighting challenges in accurately determining confluency. Dotted lines indicate treatment time points with 1 μM Doxo. The asterisk and black bar indicate the time points where there were statistically significant differences between the monoculture and the multi-cell-type culture setups; ∗p < 0.05.(D) Representative contraction traces of hiPSC-CMs in both monoculture and multi-cell-type conditions at 48 and 96 h (24 h after treatments 1 and 2, respectively). Treatments were either DMSO (vehicle control) or 1 or 3 μM Doxo.(E–G) Graphs comparing contraction amplitude (E), relaxation time (F), and time to peak (G) of hiPSC-CMs in either monoculture or multi-cell-type culture conditions at the indicated time points. Each was normalized to their respective vehicle control. Statistical significance was determined by two-way ANOVA analysis. Analysis is based on 3 biological replicates, each with 3 technical replicates, with error bars representing SEM; ∗p < 0.05, ∗∗p < 0.01, ∗∗∗∗p < 0.0001. See also Figures S1 and S2.
When hiPSC-CMs were treated in co-culture with hiPSC-cFBs and hiPSC-ECs, Doxo induced toxicity more rapidly than in monocultures of hiPSC-CMs. In phase contrast images, cell death was immediately evident after a single 1 μM Doxo treatment in triple cultures, while monocultures of hiPSC-CMs required 3 cycles for visible cell death (Figure 1B). Through frequent imaging, morphological changes indicating of apoptosis became evident, including cytoplasmic shrinkage, cell shape changes, and cell detachment. However, when attempting to quantify the extent of toxicity, conventional phase microscopy was unable to distinguish live from dead cells (Figure S2A) due to cellular debris and clumping, which affected the ability to apply size- or eccentricity-based threshold filters. This resulted in discrepancies with, for example, confluency measurements indicating only 20% loss in the co-culture condition at 96 h (Figure 1C), despite most cells appearing dead (Figure 1B). Nevertheless, we could distinguish differences in the toxicity dynamics between triple cultures and hiPSC-CM monocultures, with triple cultures exhibiting a more rapid onset of cell death following Doxo treatment (Figure 1C). The composition of the triple culture (70% hPSC-CMs, 15% hiPSC-ECs, and 15% hiPSC-cFBs) prior to Doxo treatment was confirmed (Figures S2B–S2D).
From video recordings of the hiPSC-cardiac cultures acquired during their treatment, differences in the beat rate of the hiPSC-CMs between the mono- and triple culture conditions were also observed (Figure 1D). Contraction traces were analyzed using CardioMotion software 24 h after the first and second treatments with either 1 or 3 μM Doxo (Stebbeds et al., 2023). Contractility was not assessed after the third cycle of Doxo treatment due to significant cell death. Although contraction amplitudes did not initially differ significantly between culture types, co-cultures had notably lower amplitudes after the second Doxo treatment (Figure 1E). Additionally, analysis of contraction duration parameters, specifically relaxation time and time to peak (Figures 1F and 1G), revealed significantly shorter relaxation times in hiPSC-CMs within the triple cultures after 1 round of Doxo treatment, suggesting more pronounced cardiotoxicity in the presence of ECs and cFBs.
To visualize the effect of Doxo on the hPSC-CMs directly, we used an NKX2.5-eGFP human embryonic stem cell (hESC) reporter line in which cells differentiating to cardiomyocytes express eGFP (Elliott et al., 2011). This confirmed the increased sensitivity of triple cultures to Doxo compared to monocultures, with overall cell confluency significantly decreasing after the first Doxo treatment (Figures 2A and 2B). The decrease in GFP area (corresponding to NKX2.5^+^ hESC-CMs) paralleled the decrease in phase confluency, indicating that the hESC-CMs were also dying sooner in response to Doxo when co-cultured with the other cardiac cell types (Figure 2C). However, again here it was challenging to accurately quantify cell death with conventional phase microscopy analysis. For example, in cultures exposed to multiple treatment rounds of 1 μM Doxo, the monocultures appeared to proliferate due to increased cell spreading before a decrease in confluency after 80 h was observed (Figures 2B and 2C). Furthermore, while visual inspection confirmed 100% cell death at the last time point, quantitative analysis based on confluency only indicated an ∼60% reduction in both phase confluency and GFP area.Figure 2. Cumulative Doxo treatment effect on hESC-CMs in mono- or tri-cellular cultures(A) Representative merged phase and green fluorescence images of NKX2.5-eGFP^+^ hESC-CMs in monoculture or co-cultured with hiPSC-cFBs and -ECs, treated with either 1 μM Doxo or vehicle control (DMSO) following the cumulative treatment protocol. Images were acquired at time points corresponding to baseline (0 h), 24 h after treatments 1 and 2 (48 and 96 h, respectively), and the final time point (192 h). Scale bar, 100 μm.(B and C) Quantification of phase (B) and green (NKX2.5^+^ hESC-CMs, C) confluency areas calculated from live cell imaging and normalized to baseline (0 h), for cell culture conditions outlined in (A) treated with 1 μM Doxo. Dotted lines indicate the treatment time points. Statistical analysis between cell culture conditions employed two-way ANOVA analysis for each time point. Analysis is based on 3 biological replicates, each with 3 technical replicates, with error bars representing SEM; ∗∗∗∗p < 0.0001. See also Figure S2.
We investigated then whether we could quantify caspase activation as a measure of Doxo-mediated apoptosis using a fluorescence assay. Although this was possible when hPSC-CMs were treated with a single high dose (10 μM) of Doxo (Figures S2E and S2F), the utility of the assay was limited in the cumulative treatment protocol with abrupt peaks in the fluorescence signal observed in all analyses performed (Figure S2G). This suboptimal fluorescence quantification was likely due to artifacts introduced with the re-addition of the dye after each medium replacement and the loss of labeled cells with washes.
Our findings thus indicated that hPSC-CMs in multi-cell-type cultures of cFBs and ECs were more sensitive to Doxo-induced toxicity than hPSC-CMs cultured alone. However, due to the complexity of the treatment protocol, accurate quantification was not possible using standard image processing assays indicating the need for alternative analytical techniques, which we next sought to address.
In silico prediction software can accurately detect and quantify hPSC-CMs and caspase activation
We investigated whether the deep neural network (DNN) tools could facilitate and improve the quantification of toxicity evident in the phase images, as well as identify specific cell types. When comparing fluorescence images of multi-cell-type cultures that contained NKX2.5-eGFP^+^ hESC-CMs and hiPSC-cFBs and -ECs, and exposed to different treatments, with images in which the NKX2.5-expressing cells were determined using the DNN, we observed a mean accuracy of 74.4% across all predictions (Figures 3A and 3B). Further analysis confirmed that in silico NKX2.5 predictions closely matched actual NKX2.5-eGFP measurements in multi-cell-type cultures treated with either a lethal concentration of Doxo (10 μM) or DMSO (Figure 3C).Figure 3. Validating the DNN tools for cardiomyocyte and caspase activity prediction(A) Matched images comparing actual NKX2.5-eGFP^+^ hESC-CMs (ground truth) merged with phase contrast to the corresponding in silico-predicted NKX2.5-eGFP^+^ cardiomyocytes derived from phase contrast imaging. Scale bar, 200 μm.(B) Analysis evaluating the distribution of the accuracy of predicted in silico masks against a total of 7,200 actual NKX2.5-eGFP^+^ images.(C) Graph depicting the overlap between in silico NKX2.5^+^ predictions and actual NKX2.5^+^ measurements over time, normalized to baseline (0 h). Lines represent control cells treated with DMSO (green) and those exposed to single 10 μM Doxo treatment (purple).(D) Matched images comparing actual caspase-3/7 expression (ground truth) merged with phase contrast to the corresponding in silico-predicted caspase-3/7 expression derived from phase contrast imaging. Scale bar, 200 μm.(E) Analysis evaluating the distribution of the accuracy of predicted in silico masks against known caspase-3/7 fluorescent dye values from a total of 8,537 images.(F) Time-course comparison of caspase-3/7 activity, contrasting in silico predictions with quantification obtained for caspase-3/7 dye-labeled cells treated with 10 μM Doxo.(G and H) In silico quantification of caspase-3/7 activity (G) and NKX2.5-eGFP^+^ hESC-CMs, normalized to baseline (0 h) (H), in monoculture versus multi-cell-type culture conditions undergoing cumulative treatment with 1 μM Doxo (dotted lines). Asterisks and black bars indicate the time points where there are statistically significant differences between the monoculture and the multi-cell-type culture setups. Statistical analysis was performed with two-way ANOVA for each time point. Analysis is based on 3 biological replicates, each with 3 technical replicates, with error bars representing SEM; ∗∗∗p < 0.001, ∗∗∗∗p < 0.0001. See also Figure S3.
Additionally, we applied a DNN to identify cells undergoing caspase-3/7-mediated apoptosis from phase contrast images. Also here, the hPSC-derived cells that were predicted by the DNN analysis to express caspase-3/7 closely matched the actual labeling, with a mean accuracy of 77.3% across the various cell types, time points, and treatments analyzed (Figures 3D and 3E). Quantification using the DNN also appeared more reliable, detecting high levels of caspase activity over the entire duration of the experiment, compared to the caspase-3/7 dye in which the signal intensity waned over time, particularly after medium changes (Figure 3F).
We then reanalyzed the phase contrast and fluorescence imaging data collected of the mono- and triple cultures of the NKX2.5^eGFP^ hESC-CMs treated with multiple rounds of 1 μM Doxo (Figure 2) using the DNN tools. Quantification better reflected the visual observation, namely, that triple cultures were more sensitive to Doxo-mediated apoptosis (Figure 3G) and that the hPSC-CM numbers declined more quickly in these wells than in monocultures (Figure 3H). We also analyzed data collected from hPSC-CMs exposed to both toxic and non-toxic cardiac-relevant drugs using the DNN caspase-3/7 software (Figure S3). As expected, caspase-3/7 activity was only predicted in the cultures treated with either Doxo or ouabain, a Na^+^/K^+^-ATPase inhibitor also known for its cardiotoxicity (Sapia et al., 2010). Other compounds, such as isoprenaline and nifedipine, as well as DMSO, were not predicted to induce caspase-3/7 activity, consistent with observations in cultures labeled with the caspase-3/7 dye (Figure S3A).
Differential Doxo toxicity responses between isogenic cardiac and non-cardiac cell types
To investigate whether the higher sensitivity of cells in the triple cultures to Doxo might be due to differential sensitivity of one cell type, we separately exposed each cell type to the cumulative Doxo treatment. We also included hiPSC-derived dermal fibroblasts (dFBs), differentiated from the same hiPSC line, as a non-cardiac cell type for comparison (Figure S4A). Phase contrast images clearly indicated that ECs were affected after a single round of exposure to 1 μM Doxo, while cFBs showed delayed cumulative effects (Figure 4A). Interestingly, the hiPSC-dFBs were not only resistant to the cumulative Doxo treatment but also continued to proliferate.Figure 4. Differential Doxo sensitivity of individual isogenic hiPSC-derived cell types(A) Representative phase contrast images of monocultures of hiPSC-ECs (top), -cFBs (middle), and -dFBs (bottom) undergoing cumulative treatment with 1 μM Doxo. Images were acquired at time points corresponding to baseline (0 h), 24 h after treatments 1 and 2 (48 and 96 h, respectively), and the final time point (192 h). Scale bar, 100 μm.(B) In silico quantification of caspase-3/7 activity for all isogenic monocultures, as well as the tri-cellular culture condition, undergoing cumulative treatment with 1 μM Doxo (dotted lines). Color-coded asterisks indicate the initial time point at which caspase-3/7 activity is significantly higher (p < 0.05) than baseline (0 h) for each cell type. Statistical analysis was performed using a two-way repeated-measures ANOVA with Geisser-Greenhouse correction. Analysis is based on 3 biological replicates, each with 3 technical replicates, with error bars representing SEM; ∗p < 0.05.(C) In silico quantification of NKX2.5^+^ cells in monoculture hiPSC-CMs versus tri-cell-type culture conditions. The asterisk and black bar indicate the time points where there are statistically significant differences between the 2 cultures (∗∗p < 0.01). Statistical analysis was performed with two-way ANOVA. Analysis is based on 3 biological replicates, each with 3 technical replicates, with error bars representing SEM. See also Figures S4 and S5.
Quantitative evaluation using the DNN caspase-3/7 tool supported the above observations, as well as the differential response observed in Figure 1B with the hiPSC-CM monocultures and triple cultures from this cell line (Figure 4B). Distinct temporal toxicity profiles were detected for each cell type, indicating variable sensitivities to Doxo. Among the individual cell types, hiPSC-ECs were the most sensitive, displaying significant in silico caspase-3/7 activity within 75 h of the initial Doxo treatment. In contrast, both cFBs and dFBs showed overall less caspase activation, with dFBs expressing almost no caspase-3/7, reflecting observations in the phase contrast images. To determine if the toxicity resistance of the hiPSC-dFBs was Doxo specific, we treated these cells with carfilzomib, a proteasome inhibitor not only used clinically to treat multiple myeloma but also known to cause broad tissue toxicity (Georgiopoulos et al., 2023). In contrast to their Doxo response, dFBs were sensitive to carfilzomib, showing significant cell death and exhibiting high levels of in silico caspase-3/7 activity within 24 h of treatment (Figures S4B–S4D). Additionally, the caspase activity of triple cultures in which the hiPSC-cFBs were substituted for hiPSC-dFBs were compared (Figure S4E). DNN caspase-3/7 analysis indicated no significant differences in caspase levels between the triple cultures.
As previously seen with the NKX2.5-eGFP^+^ hESC-CMs (Figure 3G), caspase-3/7 activity was detected in the triple culture following the first treatment but only became apparent in the hiPSC-CM monoculture after the third round of Doxo exposure (Figure 4B). Further, in silico prediction of NKX2.5 confirmed that apoptosis in hiPSC-CMs occurred significantly faster when these cells were co-cultured with other cell types (Figure 4C). Detailed Doxo dose-response and time course analysis for in silico caspase-3/7 activity (Figure S5A) confirmed that repeated exposures to Doxo at concentrations below 1 μM resulted in minimal effects after 3 cycles across most conditions tested. An exception was monocultures of hiPSC-ECs, with noticeable caspase activity detected after 3 rounds of 0.03 μM Doxo treatment. At the higher concentration of 3 μM, Doxo induced significant caspase-3/7 activity from just 1 treatment cycle in hiPSC-CM and hiPSC-EC monocultures as well as in the multi-cell type culture, while hiPSC-cFBs required 2 cycles. Treatment with 10 μM Doxo quickly resulted in significant caspase-3/7 activity in hiPSC-EC and multi-cell-type cultures within 6 h of treatment, and within 2 days of the single treatment in monocultures of hiPSC-CMs and hiPSC-cFBs. In contrast, hiPSC-dFBs did not show consistent and significant caspase-3/7 activity at any Doxo concentration.
hiPSC-ECs amplify doxorubicin-induced cardiotoxicity in co-culture systems
Given the greater sensitivity of the hiPSC-ECs to Doxo, we hypothesized that these cells contributed to the increased caspase-3/7 activity observed in the multi-cell-type cultures. To test this, we treated dual-cell cultures of hiPSC-CMs with either ECs or cFBs to either a single exposure of 10 μM Doxo or the cumulative 1 μM treatment protocol. Phase contrast images showed that within 48 h of the initial Doxo treatment, many cells in the hiPSC-CM and hiPSC-EC co-cultures appeared apoptotic (Figure 5A). While cell death was also observed in the co-culture of hiPSC-CMs and hiPSC-cFBs, it was less pronounced and only evident under the 10 μM Doxo condition. This was quantitatively supported by the DNN caspase-3/7 analysis, which showed a significantly earlier caspase response in cultures containing hiPSC-ECs following 10 μM Doxo treatment and an overall significantly higher caspase activity signal in those exposed to the cumulative 1 μM Doxo treatment (Figures 5B and 5C).Figure 5. Contribution of hiPSC-ECs and -cFBs to Doxo-induced cardiotoxicity(A) Representative phase contrast images of hiPSC-CMs co-cultured with isogenic hiPSC-ECs or hiPSC-cFBs treated with 1 μM or 10 μM Doxo. Images were acquired at time points corresponding to baseline (0 h), 24 h after treatments 1 and 2 (48 and 96 h, respectively), and the final time point (192 h). 10 μM Doxo was only administered once. Scale bar, 200 μm.(B and C) In silico quantification of caspase-3/7 activity in either co-cultures of hiPSC-CMs with hiPSC-ECs or -cFBs and treated with either 1 μM (B) or 10 μM (C) Doxo. Dotted lines indicate the treatment time points, while the asterisks and black bars indicate the time points where there are statistically significant differences between the 2 co-cultures; ∗∗p < 0.01, ∗∗∗∗p < 0.0001.(D) Simultaneous in silico quantification of caspase-3/7 activity (black line) and NKX2.5^+^ cells (red line) in co-cultures of hiPSC-CMs and -ECs treated with 1 μM Doxo at 24 and 72 h time points, comparing the dynamics of apoptosis with rate of cardiomyocyte death.(E) In silico quantification of caspase-3/7 activity in co-cultures of hiPSC-CMs with hiPSC-ECs treated with 1 μM Doxo with or without 100 μM L-NAME. Dotted lines indicate the treatment time points. Color-coded asterisks indicate the initial time point at which caspase-3/7 activity is significantly higher than baseline (0 h) for each condition. Statistical analysis was performed with two-way ANOVA. Analysis is based on 3 biological replicates, each with 2 or 3 technical replicates, with error bars representing SEM; ∗∗∗p < 0.001. See also Figure S5.
Using the DNN tools, we also examined the specific effect on hiPSC-CMs within the co-cultures treated with 1 μM Doxo (Figure 5D). As previously observed, there was a sharp increase in caspase-3/7 activity (and general cell death) within the first 24 h of treatment that then plateaued. In contrast, the number of hiPSC-CMs steadily declined over the entire treatment period, suggesting that Doxo has an acute impact on hiPSC-ECs but more gradually affects the hiPSC-CMs. Overall, these results indicated interplay between the hiPSC-CMs and hiPSC-ECs in mediating Doxo-induced cardiotoxicity and suggest that ECs may play a pivotal role in exacerbating cardiomyocyte sensitivity to Doxo.
To investigate whether this EC-CM interplay involves paracrine signaling via nitric oxide (NO), we treated co-cultures of hiPSC-ECs and -CMs with Doxo in the presence or absence of the NO synthase (NOS) inhibitor L-NAME. DNN-based toxicity analysis showed that addition of 100 μM L-NAME to the 1 μM Doxo treatment delayed the onset of caspase-3/7 activation in hiPSC-CM-EC co-cultures by 24 h compared with Doxo alone (Figure 5E). A similar delay (44 h) was observed in hiPSC-EC monocultures, but not in hiPSC-CM monocultures (Figure S5B). This temporal shift in toxicity was also observed in co-cultures of ECs and CMs derived from independent hiPSC lines, where significant caspase-3/7 activation occurred 8 h later in cultures treated with L-NAME (Figure S5C).
Discussion
Our experiments demonstrate that hiPSC-ECs accelerate the cardiotoxic effects of Doxo on co-cultured hPSC-CMs, underscoring the critical role of cell-cell interactions in drug-induced cardiotoxicity. Using ML-based tools that quantify caspase-3/7 activity from phase contrast images and can identify hPSC-CMs within mixed cultures, we achieved continuous, dye-free quantification of apoptosis together with cell type-resolved readouts. The 2D format of our assay provided straightforward accessibility for imaging and analysis, allowing cumulative Doxo effects to be attributed to specific cell types and their interactions. Although this format does not replicate the full structural and physiological complexity of the human heart, it revealed paracrine-mediated crosstalk via NO signaling as a contributor to anthracycline toxicity.
Despite its effectiveness against a broad range of cancers, Doxo is well known for cardiotoxic side effects that can manifest either early or decades after treatment (Schimmel et al., 2004). The cumulative dose is a predictor of cardiac dysfunction, yet limiting the total dose primarily reduces instances of acute toxicity without diminishing the incidence of late-onset effects (Cappetta et al., 2018). The challenge of mitigating these effects, while maintaining therapeutic efficacy, underscores a pressing need for more accurate models to understand Doxo-induced cardiotoxicity and that of its analogs (van Gelder et al., 2023).
Advanced 3D cardiac models, including microtissues and EHTs, capture multicellular interactions and reveal rapid Doxo-induced functional impairment (Qiao et al., 2020; Schwach et al., 2024). However, they are less suited to real-time, cell type-specific readouts. Our 2D multi-cell-type culture complements these systems by isolating the contributions of key cardiac cell types to Doxo responses and enabling longitudinal, label-free quantification. Accordingly, we modeled cumulative exposure with repeated 1 μM, 4-h pulses every 48 h over 8 days. This scheme was based on the reported in vivo pharmacokinetics where the free-plasma concentration of Doxo rapidly drops from initial levels ranging between 20 nM and 2 μM and has a terminal half-life of 20–48 h (Barpe et al., 2010; Greene et al., 1983; Pang et al., 2013). Under these conditions, hPSC-CM monocultures required 3 treatment rounds for visible cell death, whereas co-cultures of hPSC-CMs, -ECs, and -cFBs showed signs of caspase-3/7 activation within 24 h of the first exposure, accompanied by earlier deterioration of contractile parameters. These findings demonstrate that intercellular interactions can modulate toxicity dynamics.
Our data are consistent with prior reports that cFBs and ECs exhibit early Doxo-induced stress responses. These studies identified a pro-fibrotic influence of Doxo on fibroblasts in rat hearts via TGF-β and SMAD3 signaling, triggering their transition to myofibroblasts (Cappetta et al., 2016). Furthermore, mouse cFBs exposed to Doxo were reported to become senescent leading to the secretion of pro-inflammatory cytokines, such as IL-1β (Espitia-Corredor et al., 2022). Likewise, clinical studies have associated Doxo cardiotoxicity with damage to the vascular endothelium (Bielak-Zmijewska et al., 2014; Wojcik et al., 2015), and a recent single-cell transcriptomic study reported a significant loss of ECs in mouse hearts following repeated intraperitoneal injections of Doxo (Huyan et al., 2024). Furthermore, both in vitro and animal studies have indicated that Doxo can compromise endothelial elasticity, increase cardiac microvasculature permeability, inhibit vascular network formation, and induce oxidative stress through altered reactive oxygen species and NO levels (Cappetta et al., 2018).
Consistent with ECs playing a role in Doxo-induced cardiotoxicity, pharmacologic NOS inhibition with L-NAME delayed the onset of caspase activity in hPSC-EC monocultures and in EC-CM co-cultures, but not in CM monocultures. This suggests that EC-derived NO (and/or downstream reactive nitrogen species) contributes to the priming of cardiomyocyte susceptibility to Doxo. Although L-NAME did not abolish toxicity, the shift in apoptosis timing supports NO as a contributing driver. These observations are in line with our previous work showing that EC-CM crosstalk via the NO pathway enhanced inflammatory responses and influenced contractility in hiPSC-based systems (Arslan et al., 2023). Together, these findings highlight EC-CM communication, and specifically NO-dependent signaling, as a modulator of anthracycline cardiotoxicity and demonstrate an experimentally accessible platform for further mechanistic dissection and intervention testing. Senescence and p53 signaling are also implicated in anthracycline cardiotoxicity (Linders et al., 2024). Although not assayed here, future investigations that incorporate senescence markers and targeted p53 perturbations into the label-free pipeline will clarify how these programs intersect with the NO-dependent EC-CM effects.
Live-cell imaging commonly relies on organic fluorophores or fluorescent proteins to monitor specific cellular processes or markers (Specht et al., 2017). However, dye signal decay, which can be exacerbated by culture medium changes, limits lengthy treatment protocols involving repeated dosing, as we observed with the caspase-3/7 reagent (Figure 2F). Tagging endogenous markers with fluorescent proteins can mitigate some of these issues but restricts assays to genetically engineered cell lines, thereby limiting the ability to assess patient-specific responses. To overcome these constraints, we applied a DNN trained for apoptosis analysis to quantify in silico caspase-3/7 activity from phase contrast images, enabling continuous label-free monitoring over extended periods. Moreover, the tool also segmented and classified hPSC-CMs within mixed cultures, facilitating cell type-resolved toxicodynamic analyses across multiple hPSC lines and co-culture contexts.
ML applications in cardiotoxicity assessment, particularly with hPSC-CMs, are advancing rapidly. Models trained on functional parameters, such as action potential, calcium cycling, or contraction properties, can distinguish cardiotoxic from non-cardiotoxic compounds (Maddah et al., 2020; Pang et al., 2022; Sala et al., 2018; Serrano et al., 2023), and high-content imaging pipelines can quantify structural toxicity in fixed hiPSC-CMs (Grafton et al., 2021; Maddah et al., 2020). Notably, toxicity from continuous low dose (0.3–0.6 mM) Doxo exposure could be detected in phase contrast images of live hiPSC-CMs with greater sensitivity than cell confluency assays (Maddah et al., 2020). Our approach extends these advances by applying ML to repeated pulse exposure protocols with medium exchanges and by resolving apoptosis dynamics within multi-cell type cultures without fluorescent labels.
In summary, while hPSC-CMs have traditionally been the focus of in vitro cardiotoxicity studies, our data demonstrate that including other cardiac cell types, in particular hiPSC-ECs, critically influences Doxo cytotoxicity responses. Coupling a multi-cell-type human co-culture system with label-free ML analysis revealed EC-derived NO signaling as a contributor to the enhanced cardiotoxicity that was observed in hPSC-CM and -EC co-cultures. These findings highlight endothelial pathways as potential targets for mitigating anthracycline cardiotoxicity and exemplify how scalable human stem cell models can advance mechanism-driven drug safety testing.
Methods
hPSC culture and differentiation
All hPSC lines were maintained as previously described (Brandao et al., 2020) in either Essential 8 medium (Thermo Fisher Scientific) or TeSR-E8 medium (STEMCELL Technologies) on vitronectin-coated plates. The following hPSC lines were used in this study: LUMC0020iCTRL-06 hiPSC line (RRID:CVCL_ZA25) (Zhang et al., 2014), alpha-actinin-2^mEGFP^ hiPSC line (RRID:CVCL_WM14), NCRM-1 hiPSC line genetically modified to constitutively express mCherry (Arslan et al., 2023), and HES-3 Mesp1^mcherry^-NKX2.5^eGFP^ hESC line (RRID:CVCL_A8JT) (Den Hartogh et al., 2015).
The hPSC lines were differentiated into CMs, ECs, cFBs and dFBs following previously established protocols (Campostrini et al., 2021; Orlova et al., 2014; Itoh et al., 2013). All experiments were performed with cryopreserved, differentiated hPSCs that were thawed as previously described (van den Brink et al., 2020; Campostrini et al., 2021). Additional details regarding thawing and plating of the cells are provided in the supplemental methods.
Flow cytometry and immunofluorescence
Details regarding the preparation of the cells and staining procedure are provided in the supplemental methods. Table S1 lists antibodies used in this study.
Compound treatments
All compounds were reconstituted in DMSO according to the manufacturer’s guidelines. Table S2 provides details of the drugs, including stock and final testing concentrations. Stock solutions were stored at −20°C, thawed only once on the day of use, and diluted in mBEL CM Maintenance Medium. For single treatments, hiPSC-CMs were exposed to the compounds for 30 min at the concentrations indicated in Table S2.
For the cumulative treatment protocol, Doxo was diluted in the culture medium appropriate for each cell type. Further details are provided in the supplemental methods. For NOS inhibition experiments, L-NAME remained present throughout the entire cumulative treatment period.
Live-cell imaging and dye-based caspase-3/7 detection
High-definition phase contrast images were captured at 3-h intervals using the Standard Adherent cell-by-cell scan type on an Incucyte S3 Live-Cell Analysis system (Sartorius). Per well, 5 areas were imaged, corresponding to ∼80% of the total well area. Where applicable, NKX2.5-eGFP^+^ hESC-CMs images were acquired using the green fluorescence channel (300 ms acquisition).
For dye-based apoptosis detection, cells were incubated with Incucyte Caspase-3/7 reagents (Sartorius) prior to compound exposure, and fluorescence imaging was performed in parallel with phase contrast imaging at 3-h intervals. Further details regarding analysis are provided in the supplemental methods.
Contractility analysis
Bright-field videos were acquired on an Incucyte S3 system with modified custom acquisition software (Sartorius), immediately prior to compound addition and at 24-h intervals thereafter. Recordings (10–20 s at 100 fps) were made using a 10× objective. For analysis, each field of view was divided into 5 regions and saved as separate videos to reduce noise prior to processing. Contractile parameters were obtained using CardioMotion software (Stebbeds et al., 2023).
Analysis using in silico models
A DNN model was generated to identify hPSC-CMs. Phase contrast images were paired with corresponding NKX2.5 fluorescence images as ground truth. Image segmentation was performed using a U-Net-based architecture (Falk et al., 2019). A similar methodology was applied to train a distinct model for caspase-3/7 detection. Further details are provided in the supplemental methods.
Archived phase contrast image datasets and associated plate maps were exported from the Incucyte system and analyzed using the caspase-3/7 and NKX2.5 DNN models (Sartorius). All images acquired during the experiments were included. Processing steps are described in the supplemental methods. In silico caspase-3/7 outputs were corrected to the vehicle control (DMSO) and normalized to 0 h. In silico NKX2.5^+^ values were normalized to the initial time point (0 h).
Each experiment consisted of 3 biological replicates performed independently. Within each biological replicate, 2 or 3 technical replicate wells were included per condition, as indicated in the figure legends. Analyses were performed independently per biological replicate before pooling data for visualization and statistical testing.
Statistics
Statistical analyses were performed using Prism 9 (GraphPad). Data are presented as mean ± SEM. Time-course datasets were analyzed using a two-way repeated-measures ANOVA, with culture condition as the between-subject factor and time as the within-subject (repeated) factor. The Geisser-Greenhouse correction was applied where the assumption of sphericity was violated. Unless stated otherwise, Sidak’s post-hoc test was used to adjust for multiple comparisons between conditions and across time points. Statistical significance was defined as p < 0.05 and represented as ^∗^p < 0.05, ^∗∗^p < 0.01, ^∗∗∗^p < 0.001, or ^∗∗∗∗^p < 0.0001.
Resource availability
Lead contact
Further information and resource requests should be directed to Dr. Richard Davis ([email protected]).
Materials availability
This study did not generate any new reagents.
Data and code availability
- •The raw datasets have been deposited in Zenodo (https://doi.org/10.5281/zenodo.17974612). All other requests should be directed to the lead contact ([email protected]).
Acknowledgments
We thank F.E. van den Hill for providing endothelial cells. Research support and funding for this study were provided by Sartorius AG, as acknowledged by authors C.L.M., B.J.v.M., R.P.D., and M.B. Additional funding support came from a research grant (10.13039/100018666CARMEN; LSHM20018) co-funded by the PPP Allowance made available by 10.13039/100016036Health∼Holland TKI-LSH to stimulate public-private partnerships, a 10.13039/100032285Novo Nordisk Foundation grant (NNF21CC0073729; reNEW), and an 10.13039/100002177NWO Gravitation project funded by the Ministry of Education, Culture, and Science of the government of the Netherlands (024.003.001). K.R. is Chargé de Recherche at the 10.13039/501100001677Institut National de la Santé et de la Recherche Médicale (INSERM). The graphical abstract was created in https://BioRender.com.
Author contributions
Conceptualization, M.B., C.L.M., B.J.v.M., R.P.D., R.W., and R.S.; methodology, M.B., B.J.v.M., R.P.D., R.S., and K.B.; software, E.S.P. and P.K.; formal analysis and investigation, M.B., J.G., R.S., E.S.P., and A.C.; resources, M.P.H.M., K.R., V.O., P.K., and K.B.; writing – original draft preparation, M.B. and R.P.D.; writing – review and editing, M.B., C.L.M., B.J.v.M., and R.P.D.; funding acquisition, C.L.M., B.J.v.M., R.P.D., and R.W.; supervision: C.L.M., B.J.v.M., R.P.D., R.W., J.A., and M.W.O.
Declaration of interests
C.L.M. has advisory roles in HeartBeat.bio AG, Angios GmBH, Mogrify Limited, Cellistic, and Sartorius AG. E.S.P., P.K., A.C., R.S., K.B., R.W., J.A., and M.W.O. were all employees of Sartorius AG at the time the study was conducted. A patent related to this work (EP4145385) was filed.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Arslan U.Brescia M.Meraviglia V.Nahon D.M.van Helden R.W.J.Stein J.M.van den Hil F.E.van Meer B.J.Vila Cuenca M.Mummery C.L.Vascularized hi PSC-derived 3D cardiac microtissue on chip Stem Cell Rep.1820231394140410.1016/j.stemcr.2023.06.001PMC 1036250837390826 · doi ↗ · pubmed ↗
- 2Barpe D.R.Rosa D.D.Froehlich P.E.Pharmacokinetic evaluation of doxorubicin plasma levels in normal and overweight patients with breast cancer and simulation of dose adjustment by different indexes of body mass Eur. J. Pharm. Sci.41201045846310.1016/j.ejps.2010.07.01520688160 · doi ↗ · pubmed ↗
- 3Bielak-Zmijewska A.Wnuk M.Przybylska D.Grabowska W.Lewinska A.Alster O.Korwek Z.Cmoch A.Myszka A.Pikula S.A comparison of replicative senescence and doxorubicin-induced premature senescence of vascular smooth muscle cells isolated from human aorta Biogerontology 152014476410.1007/s 10522-013-9477-924243065 PMC 3905196 · doi ↗ · pubmed ↗
- 4Brandao K.O.van den Brink L.Miller D.C.Grandela C.van Meer B.J.Mol M.P.H.de Korte T.Tertoolen L.G.J.Mummery C.L.Sala L.Isogenic Sets of hi PSC-C Ms Harboring Distinct KCNH 2 Mutations Differ Functionally and in Susceptibility to Drug-Induced Arrhythmias Stem Cell Rep.1520201127113910.1016/j.stemcr.2020.10.005PMC 766405133176122 · doi ↗ · pubmed ↗
- 5Campostrini G.Meraviglia V.Giacomelli E.van Helden R.W.J.Yiangou L.Davis R.P.Bellin M.Orlova V.V.Mummery C.L.Generation, functional analysis and applications of isogenic three-dimensional self-aggregating cardiac microtissues from human pluripotent stem cells Nat. Protoc.1620212213225610.1038/s 41596-021-00497-233772245 PMC 7611409 · doi ↗ · pubmed ↗
- 6Cappetta D.Esposito G.Piegari E.Russo R.Ciuffreda L.P.Rivellino A.Berrino L.Rossi F.De Angelis A.Urbanek K.SIRT 1 activation attenuates diastolic dysfunction by reducing cardiac fibrosis in a model of anthracycline cardiomyopathy Int. J. Cardiol.20520169911010.1016/j.ijcard.2015.12.00826730840 · doi ↗ · pubmed ↗
- 7Cappetta D.Rossi F.Piegari E.Quaini F.Berrino L.Urbanek K.De Angelis A.Doxorubicin targets multiple players: A new view of an old problem Pharmacol. Res.127201841410.1016/j.phrs.2017.03.01628336372 · doi ↗ · pubmed ↗
- 8Csapo M.Lazar L.Chemotherapy-Induced Cardiotoxicity: Pathophysiology and Prevention Clujul Med.87201413514210.15386/cjmed-33926528012 PMC 4508592 · doi ↗ · pubmed ↗