Spatio-Temporal Variability Description of the Rare Species Lilium martagon L. in Different Habitat Conditions
Tomasz Wójcik, Kinga Kostrakiewicz-Gierałt, Maria Ziaja
TL;DR
This study examines how the rare Martagon Lily grows differently in three forest habitats in Southern Poland, finding that one population is healthier and more likely to survive.
Contribution
The study provides new insights into the spatio-temporal variability of Lilium martagon in different forest habitats and its implications for conservation.
Findings
Population 3 in Hrabeński Forest showed the highest number of Lilium martagon stems and better growth conditions.
Lack of juvenile stems in population 3 contrasts with populations 1 and 2, which had immature and generative stems.
Weather conditions in 2019 positively influenced Lilium martagon growth, but site-specific variability was observed.
Abstract
The investigations of the spatio-temporal variability of the rare species Lilium martagon L. were conducted in three populations: population 1 (located in Wolski Forest), population 2 (in Mount Chełm), and population 3 (in Hrabeński Forest). All aforementioned localities were situated in Southern Poland. The field studies were conducted in the years 2018–2023 in permanent study patches that differed regarding habitat conditions, as well as population abundance, structure, and individual traits. The greatest number of Lilium martagon stems and their substantial dimensions were recorded in population 3. The lack of juvenile stems was found in population 3, while in less abundant populations—1 and 2—juvenile, immature, virginile, and generative stems were found. The investigations performed suggest the favourable impact of weather conditions during the meteorological spring and summer of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1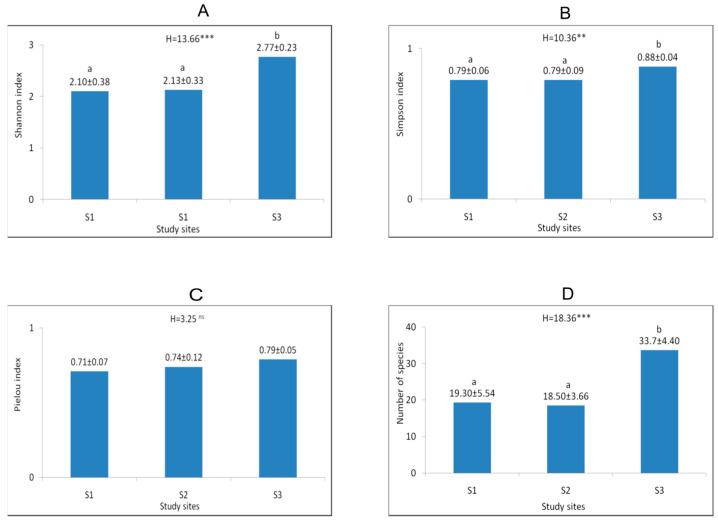 Figure 2
Figure 2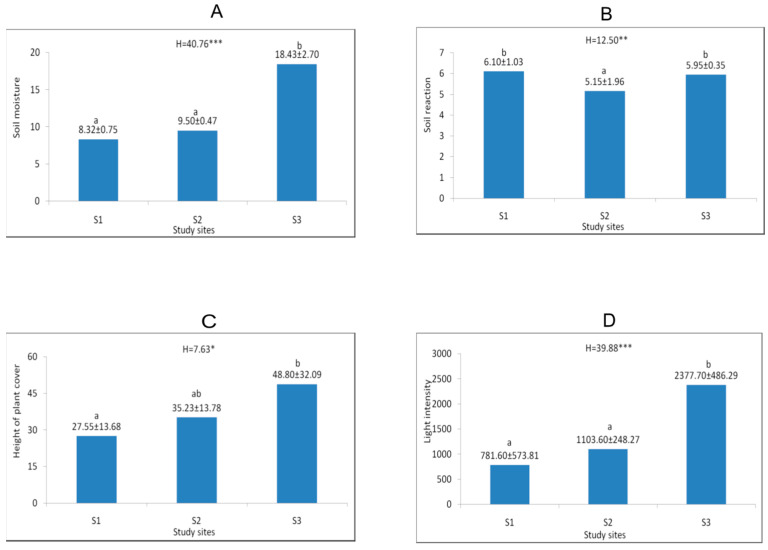 Figure 3
Figure 3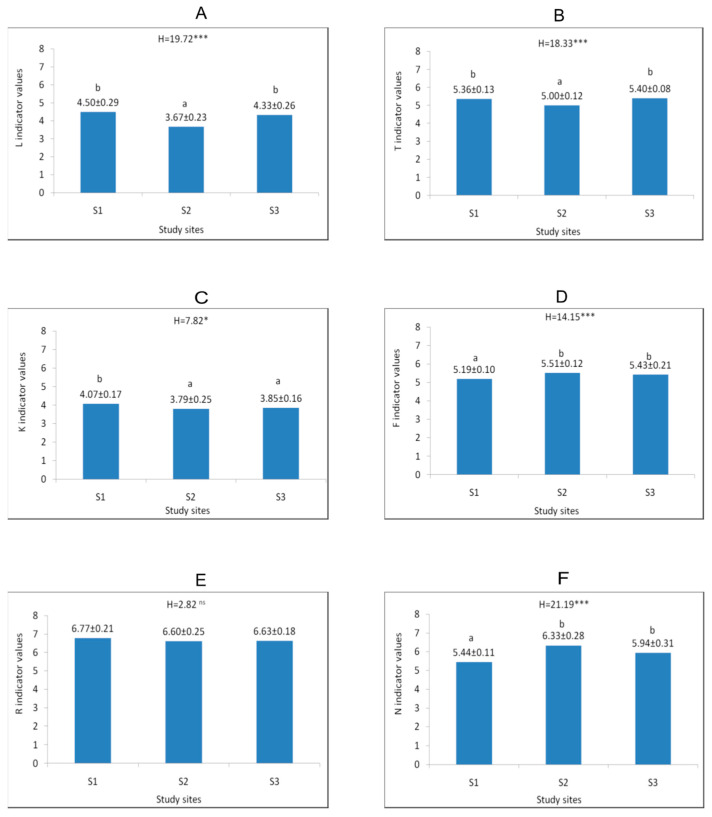 Figure 4
Figure 4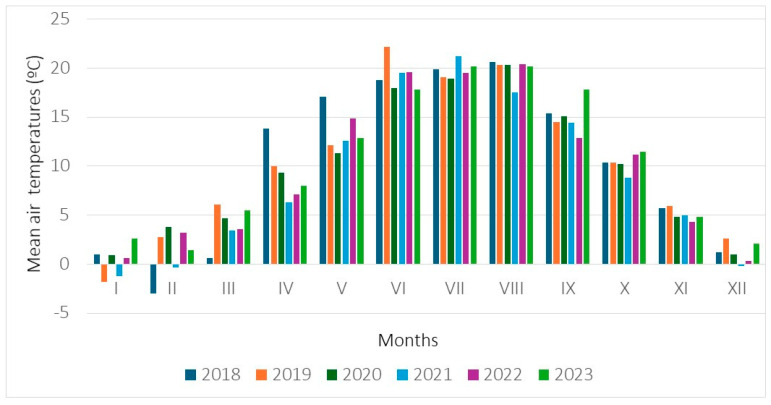 Figure 5
Figure 5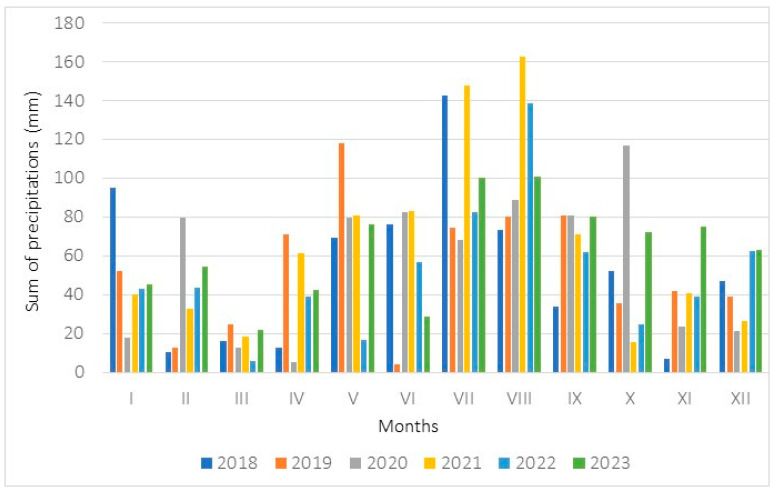 Figure 6
Figure 6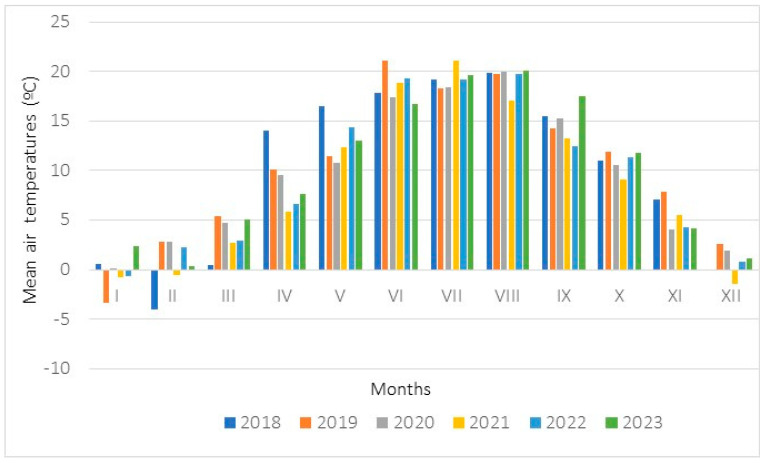 Figure 7
Figure 7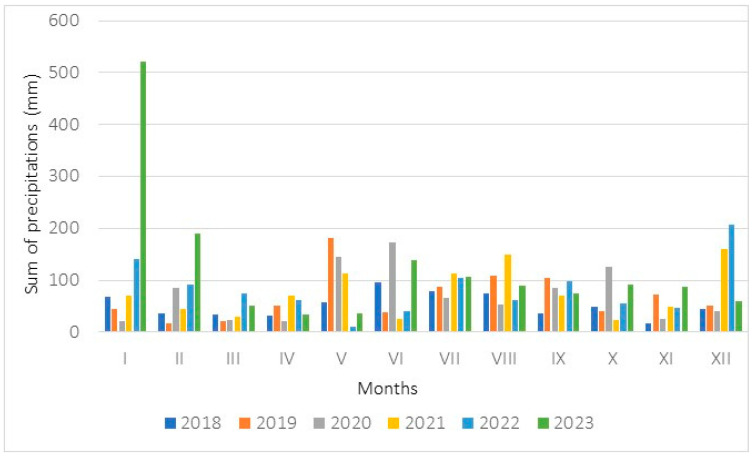 Figure 8
Figure 8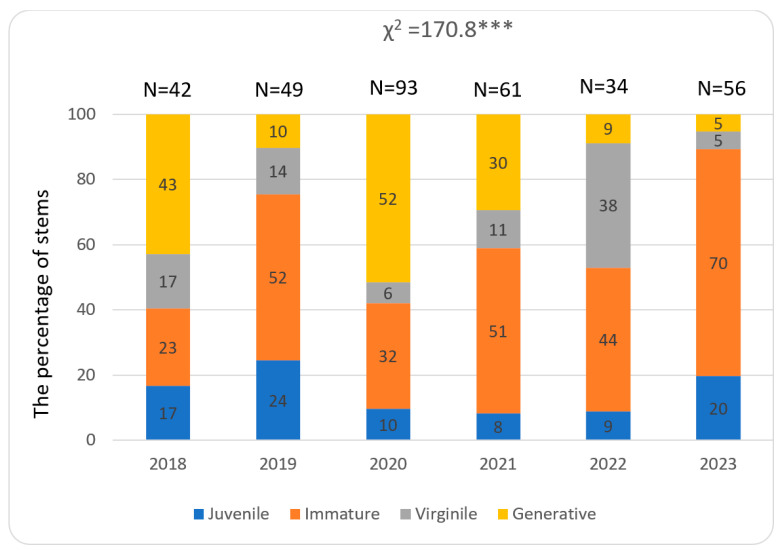 Figure 9
Figure 9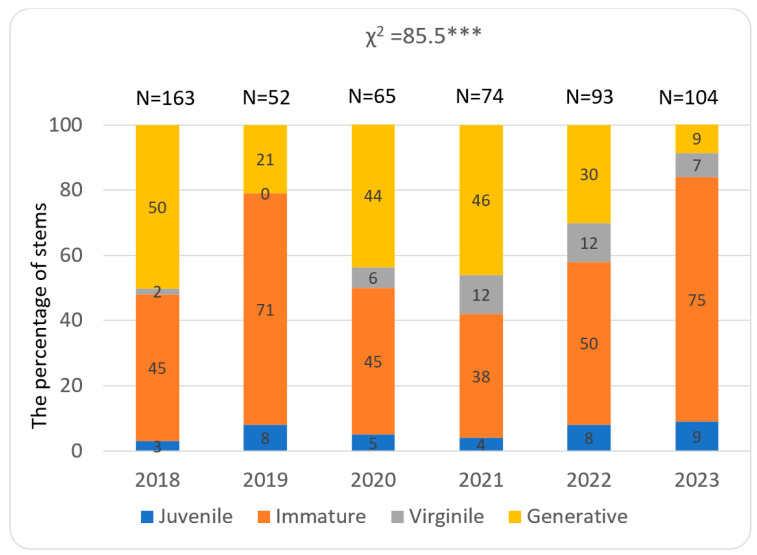 Figure 10
Figure 10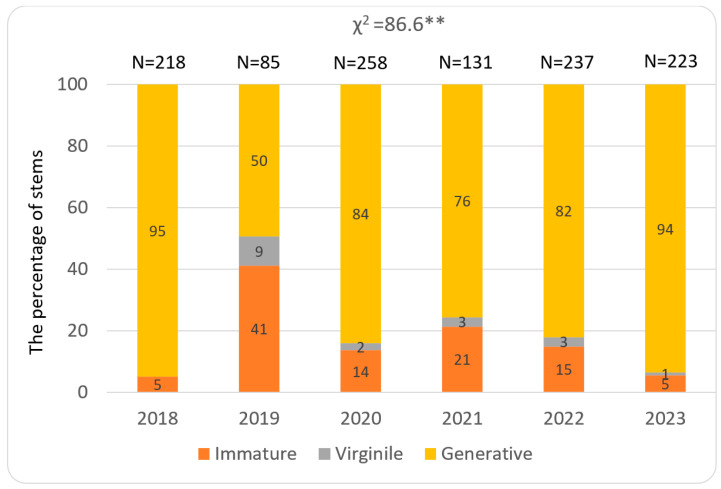 Figure 11
Figure 11- —Department of Nature Protection and Landscape Ecology, University of Rzeszów
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMorphological variations and asymmetry · Ecology and Vegetation Dynamics Studies · Species Distribution and Climate Change
1. Introduction
The genus Lilium consists of approximately 100 species and thousands of cultivars. These plants are found in temperate areas such as East Asia, Europe, and North America in the northern hemisphere, including 22 species in Europe. The genus is divided into seven sections, according to their bulb characteristics, leaf inflorescence, flower pattern, and origin, including Martagon, Archelirion, Sinomartagon, Daurolirion, Leucolirion, Pseudolirium, and Liriotypus [1,2]. Lilium is widely cultivated as an ornamental plant in Europe, North America, and Eastern Asia, playing a special role in garden design [3,4].
Lilium martagon L. (Martagon Lily) belongs to the Martagon section of the large Liliaceae family [5]. It is a 40–150 cm tall perennial with an underground golden-yellow, scaly bulb. In the central part of the stem, the leaves are arranged in characteristic whorls, usually one, sometimes two or three, while the remaining single oblong-ovate leaves, 8–12 cm long and 2–5 cm wide, are arranged spirally. It blooms from June to July. The fragrant, pendulous flowers are gathered in loose clusters at the top of the stem, numbering 3–12. The perianth tepals are 3–4 cm long, recurved upward, and pinkish-purple with darker spots. The fruit is an elongated, ribbed capsule bursting with three slits. The seeds are flat, broadly winged. It reproduces both generatively and vegetatively by producing new bulbs at the base of the mother bulb [5,6]. Lilium martagon is a geophyte, preferring partial shade and thermal conditions ranging from moderately warm to moderately cold. It grows in fresh, eutrophic soils with neutral and alkaline pH [7,8].
The aforementioned species has a wide range, covering temperate regions of Europe and Asia, extending from western, central, and eastern Europe to Mongolia (except Great Britain, Belgium, the Netherlands, the Scandinavian countries, and northern Russia) [9,10,11,12]. In Poland, it occurs quite often, except in the northwestern and western parts of the country [13]. Due to its wide distribution range, Lilium martagon grows in numerous plant communities. It is most frequently recorded in eutrophic and mesotrophic deciduous forests. In Poland, it is widespread in the Carpathians and Sudetes, from the foothills to the subalpine zone [14,15,16,17,18,19,20,21,22], and is less common in the lowlands [23,24]. It is a characteristic species of meso- and eutrophic deciduous forests of the order Fagetalia sylvaticae and a distinctive species of the Bupleuro–Calamagrostietum arundinaceae association, a group of high-mountain herbaceous plants from the Calamagrostion alliance [25]. Moreover, it also occurs in beech forests in the Eastern Alps, the Bohemian Massif and the Western Carpathians [26], subalpine beech forests in the Julian Alps, and in the northern Dinaric Alps [27], as well as in southeastern Europe in the Balkan Peninsula, from the southeastern areas of the Alps in Slovenia, through Croatia, Bosnia and Herzegovina, Serbia, Montenegro, and the Republic of Macedonia to northern and northeastern Greece and Bulgaria (alliance Aremonio-Fagion and Fagion moesiacae) [28]. Furthermore Lilium martagon occurs in oak-hornbeam forests, including the subcontinental oak-hornbeam forest Tilio cordatae–Carpinetum betuli [14,21,29] and the Central European Galio silvatici–Carpinetum [23], and is recorded in riparian forests of the Alno–Ulmion alliance [19,30,31] and in the subcontinental oak-hornbeam forest association Potentillo albae–Quercetum [32]. Additionally, it was noted in Carpathian mountain grasslands in the associations Festucetum saxatilis, Ranunculo platanifolii–Adenostyletum alliariae, Hyperico alpigeni–Calamagrostietum villosae [33,34], and Alpine pastures [35]. This species can also be found sporadically in anthropogenic habitats, such as old parks and cemeteries [36,37].
In Poland, Lilium martagon is under strict species protection [38] and classified in various threat categories depending on the region. It is considered as an endangered species (EN) in the Suwałki Lakeland [39], a vulnerable species (VU) in Western Pomerania [40] and the South Podlasie Lowland [41], whilst a near threatened species (NT) in Gdańsk Pomerania [42] and in the Silesian Voivodeship [43]. Moreover, Martagon Lily is considered as a species of least concern (LC) in Lower Silesia [44], in the Opole Voivodeship [45], and in Greater Poland [46], and as a low-risk species (LR) in the Proszowice Plateau [47]. It is also listed as endangered in neighbouring countries. Lilium martagon is included in the Red List of Vascular Plants of the Czech Republic with the NT category [48], the Red List of Vascular Plants of the Ukrainian flora with the LC category [49], the Red List of Vascular Plants of the Carpathians with the LC category [50], and the Red Book of Vascular Flora of Croatia with the VU category [51]. It has also been included in the European Red List of Medicinal Plants [52].
To date, the majority of investigations have focused on taxonomic and phylogenetic relationships among particular taxa representing the genus Lilium [11,14] or have addressed the issue of reproduction or development of individuals [53,54,55,56,57,58,59].
Due to the threat of extinction of Lilium martagon, studies of its population abundance and structure, as well as observations of individual traits, are particularly important because they allow for an evaluation of the prospects for persistence of the population in the occupied habitat and are the basis for undertaking specific protective actions. So far, such studies have been conducted among others in Poland [23,24,60,61,62,63,64,65,66], Ukraine [67], Russia [68], and Latvia [10,69]. Nevertheless, the aforementioned studies addressed solely selected individual traits and were conducted over one [16,23,29,60,65], two [62,66], or three [10] growing seasons in a low number of study sites. Longer observations lasting at least four years were carried out by Balode [69] and Pindel [63]. The longest six-year investigations of individual traits of Lilium martagon were carried out by Miciniak and Zątek [24]. However, the aforementioned research was conducted in selected years of a six-year period. All these authors analysed the influence of habitat conditions, such as light intensity and soil conditions as well as floristic and phytocoenotic variability of ecosystems on the state of Lilium martagon. The influence of weather conditions on Martagon Lily population was investigated exclusively by Truchan and Sobisz [64]. Based on three-year observations conducted at an anthropogenic locality, they confirmed the negative effect of precipitation on population abundance. Despite growing interest in the aforementioned issues, the present state of knowledge is still insufficient. Therefore, the presented six-year study was undertaken to evaluate the spatial and temporal variability of population and individual traits of Lilium martagon. The specific aims focused on assessing: (i) habitat conditions at all study sites, (ii) weather conditions in the study period, (iii) abundance and structure of the studied populations of Lilium martagon, and (iv) selected traits of stems of Lilium martagon representing different developmental stages.
2. Materials and Methods
2.1. Study Area
The investigations were conducted in three Lilium martagon localities in Southern Poland. Site 1 is located in Wolski Forest in the western part of Kraków (50.057617° N, 19.885717° E), within the Bielańsko-Tyniecki Landscape Park. In terms of physico-geographical regionalisation, this area is located in the Kraków Bridge mesoregion of the Kraków Gate macroregion [70]. A characteristic feature of this area is its varied relief, within which various landforms can be distinguished. The highest parts of the hills are occupied by a plateau, covered with loess deposits, on which a Pino–Quercetum mixed coniferous forest has developed. The slopes are cut by deep valleys; brown soils and rendzinas occurring here are covered by Tilio cordatae–Carpinetum betuli and Dentario glandulosae–Fagetum associations. On limestone rocks and in their surroundings, xerothermic grasslands representing the Festuco–Brometea class and shrubs of the Rhamno–Prunetea class can be found [71,72]. The observed Lilium martagon population (called in this study population 1) occurs in the Tilio cordatae–Carpinetum betuli association at an altitude of 243 m above sea level in the eastern part of Wolski Forest.
Site 2 is located on Mount Chełm in the village of Jaszczurowa (49.888957° N, 21.559002° E). This area is situated within the Mount Chełm Nature Reserve and the Czarnorzecko-Strzyżowski Landscape Park. In terms of physico-geographical regionalisation, site 2 is located in the Strzyżów Foothills mesoregion, in the Mid-Beskidy Foothills macroregion [70]. Mount Chełm is composed of Carpathian flysch, with thick-bedded sandstones in the substrate. The aforementioned reserve protects an extrazonal site of the Dentario glandulosae–Fagetum association, an area of springs with the Carici remotae–Fraxinetum association, the Tilio cordatae–Carpinetum betuli association, and an abandoned quarry [73,74]. The population of Lilium martagon (labelled population 2) occurs in the top part of the Mount Chełm (520 m above sea level) in a Dentario glandulosae–Fagetum association.
Site 3 is located in Hrabeński Forest in the village of Besko (49.594234° N, 21.920799° E), situated in the Jasło–Krosno Basin mesoregion, in the Mid-Beskidy Foothills macroregion [70] and is protected within the Natura 2000 Hrabeński Forest PLH180039 site. The substrate consists of Carpathian flysch composed of a series of alternating shales and sandstones. Hrabeński Forest occupies the hills between the Wisłok and Tabor river valleys, where the elevation differences reach up to 100 m. In the lowest part of the forest, on gentle slopes, the Tilio cordatae–Carpinetum betuli association is found. In the higher parts of the hills, on slopes with greater inclination and varied exposure, Carpathian beech forest occurs in the rank of two subassociations: Dentario glandulosae–Fagetum typicum and Dentario glandulosae–Fagetum lunarietosum [21]. The population of Lilium martagon (called population 3) occurs throughout the whole forest complex; however, it is most abundant in the Tilio cordatae–Carpinetum betuli association at an altitude of 255 m above sea level.
2.2. Data Collection
In 2018, the investigations of plant cover and habitat conditions were conducted at three study sites: Wolski Forest, Mount Chełm, and Hrabeński Forest. At each site, 10 phytosociological relevés covering an area of 100 m2 were taken using the Braun-Blanquet method [75]. The names of vascular plants are given according to Mirek et al [76]. The description of vegetation includes species covering more than 5% of a given plant layer. Syntaxonomic classification was based on the Matuszkiewicz system [25]. For each phytosociological relevé, the Shannon–Wiener diversity index [77], Pielou index [78], Simpson index [79], and number of species were calculated using JUICE 7.1 software [80]. Then, mean values of diversity indices for plant communities occurring in the three study sites were calculated.
Subsequently, within each study site one permanent research patch measuring 20 m × 20 m was established. Altogether, in each permanent patch, 10 measurements of soil pH, 20 measurements of soil moisture (%), and herbaceous vegetation height (cm), as well as 30 measurements of light intensity (lx) were taken. Soil pH was measured with a soil acidimeter (RIM KOWALCZYK SP.J., Janki, Poland) using Hellig’s solution, soil moisture with an Extech MO750 hygrometer (Extech Instruments Corporation, Maastricht, Netherlands), and light intensity at ground level with a TES1335 luxmeter (TES Electrical Electronic Corp., Taipei, Taiwan). The aforementioned measurements were conducted from 10 to 15 June 2018, from 12:00 to 15:00 in full sunlight. Additional analyses of habitat conditions were conducted using Ellenberg indicators [81]. Ellenberg indicators were calculated for each phytosociological relevé in JUICE programme [80] and then mean values for the plant communities occurring in particular study sites were computed.
In order to describe the temporal variability of weather conditions in the study period, monthly sum of precipitation [mm] and monthly mean temperature [°C] data from Kraków and Krosno stations (situated near the Mount Chełm and Hrabeński Forest sites) were taken from the website TuTiempo (Accessed on 21 March 2025) [82].
The investigations of populations of Lilium martagon were conducted in the years 2018–2023 within the aforementioned permanent plots measuring 20 m × 20 m. In each study year, all stems of Lilium martagon occurring within the permanent patches were counted. Taking into account the fact that the accurate determination of age of individuals in clonal, strictly protected populations of Lilium martagon in Poland [38] is possible only on the basis of bulb morphology, instead of age, the developmental stage of the stems was determined on the basis of the number of leaves [29,61,62]. Considering this, each stem was assigned to one of the following stages: (i) juvenile stage (stem with 1–2 leaves), (ii) immature stage (3–12 leaves), (iii) virginile (mature vegetative) stage (more than 12 leaves and lack of inflorescence), or (iv) generative stage (with developed inflorescence).
Next, observations of selected individual traits of the stems of Lilium martagon were carried out. The height, number of leaves in the whorl, and length and width of the longest leaf in the whorl were observed in immature stems. The height, number of leaves in the lower whorl, and length and width of the longest leaf in this whorl were observed in virginile stems. The height, number of leaves in the lowest whorl, length and width of the longest leaf in this whorl, number of spiral leaves, and length and width of the spiral leaf placed above the highest whorl, as well as the number of flowers, were recorded in generative stems. When the number of stems representing a particular stage reached no more than 30, all stems were measured; meanwhile, if they occurred more abundantly (at least 31 stems), the number of examined stems was limited to 30. The stems were randomly selected based on 30 throws with an iron rim, 20 cm in diameter. Each time, one stem occurring in the centre of the rim was measured and labelled.
2.3. Statistical Analysis
The non-parametric Kruskal–Wallis H test for multiple comparisons was applied to check the statistical significance of differences in values of:
- -Shannon index, Simpson index, Pielou index, number of species, soil moisture, soil pH, height of plant cover and light intensity at ground level, as well as values of Ellenberg indices among study sites;
- -Individual traits of immature stems of Lilium martagon (height, number of leaves in the whorl, and dimensions of the longest one) among populations in the years 2018–2023;
- -Individual traits of virginile stems (height, number of leaves in the lower whorl, and dimensions of the longest one) among populations in the years 2020–2023;
- -Individual traits of generative stems (height, number of flowers, number of whorls, number of leaves in the lowest whorl and dimensions of the longest one, number of spiral leaves, and dimensions of spiral leaf placed above the highest whorl) among populations in the years 2018–2023.
The non-parametric Mann–Whitney U test was used to check the statistical significance of differences in values of height, number of leaves in the lower whorl and dimensions of the longest leaf in virginile stems among populations in the years 2018 and 2019. Moreover, the non-parametric Kruskal–Wallis H test for multiple comparisons was applied to check the statistical significance of differences in values of individual traits of stems in the study years. Additionally, the d-Cohen coefficient was calculated to check the scale of practical significance of differences among pairs of data referring to habitat conditions at the study sites, as well as individual traits of stems representing different populations. The scale of practical significance of differences based on absolute values of the d-Cohen coefficient was as follows: <0.2—negligible, 0.21–0.49—small, 0.5–0.79—moderate, and >0.8—substantial difference. The χ^2^ test was used to test the statistical significance of spatial and temporal differences in the share of stems at various developmental stages. The analyses were conducted using Statistica 13.3 software and an interactive χ^2^ test calculator [83].
3. Results
3.1. Habitat Conditions
Analysis of phytosociological relevés showed that the studied sites differed in terms of vegetation cover. At sites 1 (Wolski Forest) and 3 (Hrabeński Forest), the Tilio cordatae–Carpinetum betuli oak-hornbeam forest association was observed, while at site 2 (Mount Chełm) the Dentario glandulosae–Fagetum mountain beech forest association was recorded (Table 1, Figure 1). At all sites, the tree layer was well developed, reaching an average cover of 77 to 88%. At site 1, the tree layer was dominated by Tilia cordata and Quercus robur, at site 2 Fagus sylvatica and Acer pseudoplatanus prevailed, while at site 3 Carpinus betulus, Acer pseudoplatanus, and Quercus robur were represented most abundantly. The shrub layer was relatively well developed in the Tilio cordatae–Carpinetum betuli association, where its average cover was 34% (site 3) and 45% (site 1). At site 2, the shrub layer was poorly developed, with an average cover of only 5%. The herbaceous vegetation layer achieved high, but varied, coverage across sites. It was most developed at site 3 (91% coverage), where as many as eight species achieved coverage exceeding 5%, including Lilium martagon and Stellaria holostea, a characteristic species of the oak-hornbeam forest. At site 2, the herbaceous vegetation was dominated by Dentaria glandulosa, a characteristic species of the Dentario glandulosae–Fagetum association, but Galeobdolon luteum and Rubus hirtus also had significant cover. Herbaceous vegetation at site 1 was slightly less developed (68%). Five species achieved high coverage in this layer, including Stellaria holostea, a species of the oak-hornbeam forest. The highest species richness was observed at site 3, where an average of 34 species per phytosociological relevé was recorded. This site also had the highest values of the Shannon–Wiener, Simpson, and Evenness indices (Figure 2), as confirmed by the values of the d-Cohen coefficient (Table A1).
The study sites differed in terms of habitat conditions. The highest values of soil moisture, light intensity at ground level, as well as height of plant cover were noticed at site 3. The values of the d-Cohen coefficient confirmed the substantial differences, especially in the case of soil moisture and light intensity (Table A1). At sites 1 and 3, located in the oak-hornbeam forests, the soil pH was neutral, while at site 2, situated in a beech forest, it was slightly acidic (Figure 3). The substantial differences in soil reaction between the oak-hornbeam and beech forests were confirmed by the d-Cohen coefficient values (Table A1). Analysis of Ellenberg’s climatic indicators revealed significantly higher L and T values at sites 1 and 3 than at site 2. In the case of edaphic indicators, significant differences were observed for F and N. They reached much greater values at sites 2 and 3 than at site 1. Additionally, the K indicator was greater at site 1 than at the sites 2 and 3 (Figure 4). All aforementioned differences were confirmed by the values of the d-Cohen coefficient (Table A2).
3.2. The Selected Weather Conditions in the Study Period
Weather conditions in the study site where population 1 occurred were evaluated on the basis of data from the Kraków station, where measurements were taken (Figure 5 and Figure 6). The annual sum of precipitation ranged from 615.1 mm (in the year 2022) to 781.5 mm (2021). The mean annual air temperatures achieved values ranging from 8.9 °C (in the year 2021) to 10.4 °C (2019, 2023). During the meteorological spring occurring from 1 March till 31 May, the sum of precipitation ranged from 61.5 mm (in the year 2022) to 214.1 mm (2019), while the mean air temperatures ranged from 7.4 °C (in the year 2021) to 10.5 °C (2018). During the meteorological summer (from 1 June to 31 August), the sum of precipitation rose from 159.3 mm (2019) to 394.0 mm (2021), while the air temperatures achieved values ranging from 19.1 °C (2020) to 20.5 °C (2019).
Weather conditions at the study sites where populations 2 and 3 occurred were evaluated on the basis of data from the Krosno station, where measurements were taken (Figure 7 and Figure 8). The annual sum of precipitation rose from 623.6 mm (in the year 2018) to 1483.6 mm (2023). The mean annual air temperatures achieved values ranging from 8.7 °C (2020) to 12.0 (2018). During the meteorological spring, the sum of precipitation ranged from 123.0 mm (2023) to 254.5 mm (2019), while the mean air temperatures ranged from 6.4 °C (2023) to 9.0 °C (2019). During the meteorological summer, the sum of precipitation rose from 207.5 mm (2022) to 334.7 mm (2023), while the measured mean air temperatures ranged from 17.9 °C (2020) to 19.7 °C (2019).
3.3. Spatio-Temporal Variability of Abundance and Structure of Developmental Stages of Populations
The abundance of stems of Lilium martagon in the study patches and structure of their developmental stages showed both spatial and temporal. The total number of stems was the greatest in population 3, where it ranged from 85 to 237 in particular study years. A much lower number of stems, ranging from 52 to 163, was recorded in population 2, while in population 1 34 to 93 stems were noted (Figure 9, Figure 10 and Figure 11).
During the whole study period in populations 1 and 2, juvenile, immature, virginile, and generative stems occurred. In population 1, the greatest share of juvenile stems (comparing other populations) especially in the year 2019 was observed, while the contribution of stems representing the remaining stages differ in particular years (Figure 9). In population 2, a share of immature stems greater than at other study sites was noted. Also, the percentage of generative stems was substantial during almost all study years (Figure 10). In population 3, juvenile stems were absent and the contribution of generative stems was much greater than in other populations (Figure 11). The temporal variability among the populations was statistically significant and ranged from χ^2^ = 47.00 (p < 0.001) in 2021 to 266.4 (p < 0.001) in 2023.
3.4. The Spatial Variability of Selected Individual Traits of Lilium martagon
The mean height of immature stems increased from population 1, where it achieved from 11.02 cm to 17.56 cm, via population 2 (11.53–17.12 cm), to population 3 (13.71–18.77 cm). The mean number of leaves in the whorl ranged from 5.58 to 9.40 in population 1, from 6.63 to 7.85 in population 2, and from 7.30 to 8.95 in population 3. Such phenomena were confirmed by the values of the Kruskal–Wallis H test for multiple comparisons (Table 2) and the d-Cohen coefficient (Table A3). The mean length of the longest leaf exhibited the lowest values (94.15–119.14 mm) in population 3 and was the greatest (97.90–139.28 mm) in population 1. Also, the mean width of the aforementioned leaf was the lowest (32.96–38.77 mm) in population 3 and the greatest (35.70–39.75 mm) in population 1 (Table 2). These tendencies were also confirmed by the statistical analysis (Table 2 and Table A3).
The mean height of the virginile stem was the lowest (13.14–22.86 cm) in population 1 and the greatest (22.17–31.25 cm) in population 3. The mean number of leaves in the lower whorl was the lowest (5.33–7.80) in population 1 and the greatest (8.29 to 11.00) in population 3. The differences among the populations were confirmed by small, moderate, or substantial values of the d-Cohen coefficient (Table A4). The mean length of the longest leaf ranged from 103.43 mm to 128.71 mm, while the mean width of this leaf varied from 29.40 mm to 45.57 mm, with limited variation among the populations confirmed by the Kruskal–Wallis H test (Table 3). Interestingly, the substantial values of the d-Cohen coefficient indicated a remarkable size effect in some study years (Table A4).
The mean height of generative stems achieved the lowest values (55.27–72.93 cm) in population 2 and was the greatest (58.67–97.00 cm) in population 1 (Table 4). The significant differences between populations 1 and 2 were confirmed by the d-Cohen coefficient values (Table A5). The mean number of flowers ranged from 2.17 to 2.68 in population 2, from 3.03 to 4.58 in population 3, and from 1.67 to 6.20 in population 1. The mean number of whorls reached the lowest values (1.40–1.82) in population 2, whereas the greatest number (1.44–3.00) was recorded in population 1. In the case of the number of flowers and whorls, the statistical analysis confirmed the moderate level of the d-Cohen coefficient between populations 1 and 2 (Table A5). The mean number of leaves in the lowest whorl had the lowest values in population 1 (5.57–7.00) and the greatest values in population 3 (8.00–9.47). This phenomenon was confirmed by the substantial values of the Kruskal–Wallis H test (Table 5) and the d-Cohen coefficient (Table A6).
The mean length of the longest leaf in the lowest whorl was the lowest (109.67–126.13 mm) in population 3 and the greatest (119.57–147.80 mm) in population 1. The mean width of the longest leaf achieved the lowest values (35.03–39.63 cm) in population 3 but was the greatest (34.93–43.00) in population 2. The mean number of spiral leaves ranged from 3.73 to 5.20 in population 2, from 4.30 to 5.26 in population 3, and from 4.00 to 8.50 in population 1. The values of the Kruskal–Wallis H test (Table 5) and the d-Cohen coefficient (Table A6) indicated small or moderate differences in the aforementioned leaf dimensions among the populations.
The mean number of spiral leaves reached the lowest values (3.59–5.20) in population 2 but was the greatest (4.00–8.50) in population 1 (Table 5). This phenomenon was confirmed by the values of the Kruskal–Wallis H test (Table 5) and the d-Cohen coefficient (Table A6).
The mean length of the spiral leaf above the highest whorl achieved the lowest values (81.83–94.68 mm) in population 3 and was the greatest (97.67–113.33 mm) in population 1. The mean width of the aforementioned leaf exhibited a similar tendency, ranging from 16.73 mm to 23.68 mm in population 3, from 21.10 mm to 25.13 mm in population 2, and from 20.20 mm to 30.53 mm in population 1. The greatest leaf dimensions observed in population 1 were confirmed by the values of the Kruskal–Wallis H test (Table 5) and the d-Cohen coefficient (Table A6).
3.5. The Temporal Variability of Selected Individual Traits of Lilium martagon
The height of immature stems in population 1 presented significantly lower values in the years 2018–2020 than in subsequent years, while in population 2 and in population 3 the highest stems were noted in the year 2019 (Figure S1, Table S1). The number of leaves in the whorl was significantly greater in the year 2019 than in other study seasons solely in population 1, while in the remaining populations this trait did not show temporal variability. The length of the longest leaf was significantly greater in the year 2019 than in other study seasons solely in population 1. In population 2, the length was lower in the years 2018–2020 than in the years 2021–2023, whereas in population 3 much lower values were observed in the years 2020, 2021, and 2023 than in other study seasons. The width of the longest leaf in population 1 was much lower in the years 2020 and 2022 than in other years; in population 2, it was much lower in the years 2018–2020 than in subsequent study seasons, while in population 3 the aforementioned trait did not present temporal variability.
The individual traits of virginile stems did not present temporal variability in population 2 or in population 3. In population 1, the height of virginile stems and the length of the longest leaf in the lower whorl reached the greatest values in the year 2021 and the lowest values in the year 2018 (Figure S2). The moderate and substantial values of the d-Cohen coefficient indicate remarkable differences in the height of stems and leaf dimensions, particularly in population 1 (Table S2).
The height of generative stems in population 1 achieved much greater values in the year 2019 than in remaining study seasons (Figure S3). In population 2, the greatest values of the aforementioned trait were observed in the years 2022, while in population 3 this trait reached its greatest values in the years 2022 and 2023. The number of flowers and whorls showed temporal variability in population 1, where this trait reached its greatest values in the year 2019, as well as in population 3, where it achieved the greatest values in the years 2022 and 2023. The substantial values of the d-Cohen coefficient regarding the height, number of flowers, and number of generative whorls indicate significant differences between 2019 and the other years (Table S3).
The number of leaves in the lowest whorl presented temporal variability solely in population 3, where it achieved the greatest values in the year 2019. This phenomenon was confirmed by the values of the Kruskal–Wallis H test (Figure S4) and the d-Cohen coefficient (Table S4). The length of the longest leaf in the lowest whorl showed temporal variability in population 2 and in population 3. In population 2, it reached the greatest values in the years 2022 and 2023, while in population 3 the greatest values were observed in the year 2019. This phenomenon was confirmed by the small and moderate values of the d-Cohen coefficient (Table S4). The width of the aforementioned leaf in population 2 was significantly greater in the years 2021 and 2022 than in other study seasons, whereas in other populations the values of this trait were similar in individual years. The number of spiral leaves showed temporal variability in population 1 and in population 2 (Figure S5, Table S4). The lowest values were observed in the year 2023 in the case of population 1 and in the year 2020 in the case of population 2. The length and width of the spiral leaf did not exhibit temporal variability.
4. Discussion
4.1. Habitat Conditions
Lilium martagon occurs at various geographical latitudes, both in the warmer regions of southern and western Europe and in central Asia, where the climate is distinctly continental [9,11], indicating that it is a species neutral to continentality with a wide range of climatic tolerance. In cooler regions, it prefers partially wooded ecosystems [68,84], whereas in sunnier and drier areas, it finds optimal growth conditions in dense and shaded forests [14,27,28]. The investigated populations are located within forest communities with diverse light conditions. In Hrabeński Forest, light intensity measured at the ground was more than twice as high as at the other sites. This was the most numerous population, characterised by a high proportion of flowering and fruiting individuals in all years of the study. Urbaniec and Bach [16] and Pindel [63] reported the positive effect of light on the development of Lilium martagon, finding that optimal conditions for the development of this species occur at light levels approaching 60%. In turn, according to Jańczyk-Węglarska and Węglarski [61], the optimal conditions for the occurrence of this species range from 5 to 20% of light reaching the herbaceous vegetation layer. According to Pindel [63], increasing stand density and reduced light access negatively impact the flowering process, and at light levels below 5% the process is completely inhibited. We observed a similar relationship at the other two sites, where lower light intensity may have negatively impacted the number of flowering individuals.
According to the literature on the subject, Lilium martagon can occur on various soil types (including podzolic soils, brown soils, leached brown soils, gley soils, and rendzinas) and on various formations (including loess formations and calcareous clay soils), but their common feature is a high humus content and high or moderate nitrogen content [10,16,23,61,69,85]. Such findings are consistent with the results of the presented investigations, particularly Ellenberg N index values suggesting soils moderately rich in nitrogen. However, the content of available compounds can reach diverse values. Moreover, Lilium martagon has a wide amplitude in relation to soil pH and it occurs in habitats ranging from very acidic to alkaline. However, many authors emphasise that the optimal pH for this species ranges from 5 to 7 [16,23,58,60,61,62,63]. The aforementioned findings are supported by the performed investigations showing that average pH value of the study sites ranged from 5.15 to 6.10.
Analysis of moisture conditions showed twice as high humidity in Hrabeński Forest, whose habitat can be described as moderately moist. At the other two sites, the soils were fresh, and during prolonged periods without rainfall in the summer, they dried out. Some authors have pointed out that individuals of Lilium martagon develop best in fresh habitats [7,8], but other literature data indicate that it can also occur in seasonally moist habitats, such as riparian forests [19,30,31,69]. At the same time, it is worth mentioning that excessive humidity in spring, resulting from high rainfall totals and the formation of ponds, can lead to bulb rot and the decline of entire populations [63]. A similar relationship in this type of habitat was also observed in the case of the Arum alpinum species [73,86].
In the temperate climate zone, individuals of Lilium martagon have optimal conditions for development in fertile and mesotrophic deciduous forests; therefore, this species is recognised as a characteristic species of the order Fagetalia [25,27]. In the lowlands and foothills, it occurs mainly in oak-hornbeam forests, including in the Tilio cordatae–Carpinetum betuli and Galio sylvatici–Carpinetum betuli associations [14,23,29,60,85,87], while it grows much less frequently in pine forests and mixed forests [69,78,88], light oak forests, [24,32] and riparian forests [19,30,31]. In the montane zone and extrazonally in the foothills, it is a frequent component of beech forests, especially the Dentario glandulosae–Fagetum association [14,15,16,26,27,62,85], while in higher locations it occurs in alpine grassland communities [16,33,34]. Interestingly, this species can establish itself in anthropogenic habitats, such as manor parks and cemeteries, where it was intentionally introduced [10,36,37,69]. Our study results confirm the literature data regarding the preferences of Lilium martagon for the phytocoenoses it occupies. Two of the sites we studied were located in the oak-hornbeam forest Tilio cordatae–Carpinetum betuli, while the third site was in the mountain beech forest Dentario glandulosae–Fagetum.
Natural forest phytocoenoses with Lilium martagon are characterised by high species richness, and a particularly high share of characteristic species from the order Fagetalia. In such communities, Lilium martagon populations are usually more numerous and stable [23,30,32,85,87]. We observed similar relationships in our studies. The highest species richness and the highest diversity indices were recorded in Hrabeński Forest, where well-developed and properly preserved natural habitats are protected [21]. The smaller number of species and lower diversity indices on Mount Chełm can be explained by the habitat type. The Carpathian beech forest Dentario glandulosae–Fagetum occurs here, which, unlike oak-hornbeam forests, is characterised by lower species richness [20]. The lowest Shannon–Wiener diversity index in the case of Wolski Forest can be explained by the high intensity of anthropopressure at this site, resulting in heavy trampling of the forest floor and the penetration of synanthropic species [89]. Other authors have also pointed out the negative impact of anthropopressure manifesting itself with a decrease in abundance or complete disappearance of the population of Lilium martagon [58,62].
4.2. The Spatial and Temporal Variability of Abundance and Structure of Population Lilium martagon
The greatest abundance of population of Lilium martagon was found at site 3 (Hrabeński Forest), where the highest values of soil moisture, light intensity at ground level, and height of plant cover were noticed. Despite the substantial share of generative stems in population 3 during whole study period, exceeding those recorded in populations 1 and 2, as well as many others [23,24,60,61,62,63], a lack of juvenile stems was noted. Considering the successful germination of the seed of Lilium martagon in laboratory conditions [90], the absence of juvenile stems in population 3 presumably might be caused by the dense vegetation cover and the absence of gaps in continuous plant cover acting as safe sites for seedling recruitment sensu Harper et al. [91]. The presence of juvenile stems in population 1 and population 2, exceeding values recorded by other authors [29,65], might be linked to the occurrence of openings in the plant cover, causing, among other things, the death of senile plants or ramets (in the case of clonal species), as well as disturbances caused by the natural activity of forest mammals, such as burrowing and digging. Furthermore, the presence of natural gaps at site 2 might be linked to the occurrence of shallow soil with a significant content of skeletal elements, such as gravel, cobbles, stones, and rock fragments, whose spontaneous subsidence causes the removal of plants. Additionally, the high tourist pressure documented at site 1 (Wolski Forest) by Kostrakiewicz-Gierałt et al. [89] might contribute to gap appearance. The aforementioned authors argued that trampling by people frequently accompanied by dogs might affect the creation of gaps in the plant cover of the forest floor.
The recorded temporal differences in the abundance of populations and their structure might be connected, on the one hand, with the recruitment of new individuals in a generative or vegetative way and their subsequent growth, and, on the other hand, with the prolonged bulb dormancy or mortality of individuals. The latter might be a consequence of the natural senescence process of individuals or be triggered by external factors such as people or herbivorous animal activity. The recreational activities of people, such as walking, jogging or biking, as well as divergence from formal trails and spontaneous creation of informal pathways, recorded among others in Wolski Forest [89], might contribute to damage or/and mortality of juvenile stems of Lilium martagon. Additionally, the blooming stems of Lilium martagon might be picked, while the bulbs might be dug out by visitors due to the ornamental value. Furthermore, herbivorous animals including wild boars, whose occurrence has been documented in Wolski Forest [92], might contribute to bulbs being dug up, affecting the mortality of individuals of the studied species. Moreover, deer, whose presence has been confirmed on Mount Chełm and in Hrabeński Forest (personal observation) may browse on the stems of Lilium martagon, contributing to a decline in the population abundance. The damage and mortality of individuals of Lilium martagon might also be caused by the activity of the scarlet lily beetle (Lilioceris lilii), traces of whose feeding activity were recorded in the studied populations, as well as in many other locations [23,65].
Furthermore, the recorded temporal differences in the abundance of populations and their structure might be linked to weather conditions. The greatest share of juvenile stems in population 1 observed in the year 2019 comparing to other study seasons might be connected with favourable weather conditions, such as considerable precipitation during the meteorological spring (especially in May) and substantial values of air temperatures during the meteorological spring and summer (particularly in June). Such observations remain in accordance with the findings of Truchan and Sobisz [64], who claimed that substantial precipitation and air temperature from May to July favour the growth and development of Lilium martagon individuals. On the other hand, in the year 2019 in populations 2 and 3, the lowest number of stems in the whole study period were recorded despite substantial precipitation and considerable air temperatures. Additionally, the share of juvenile stems in population 2 was rather similar to the share noted in other study years.
4.3. Spatial Variability of Traits of Stems of Lilium martagon
The highest number of immature and virginile stems with the greatest number of whorl leaves, as well as the substantial height of generative stems and number of whorl leaves observed in population 3 might be the result of growing in conditions of lateral shading by adjacent plants. As stated by Fiorucci and Fankhauser [93], shading by neighbouring herbaceous species reduces energy for photosynthesis and contributes to stem elongation. Under crowding conditions, the height increment is advantageous because it allows neighbours to be overtopped and contributes to more effective light interception. At the same time, it should be noted that gaps in tree canopy enabled successful light assimilation by overgrowing adjacent plants of Lilium martagon stems and might explain the lowest dimensions of whorl and spiral leaves in population 3. The lowest height of immature and virginile stems recorded in population 1 and generative stems noticed in population 2 might be caused by their over shading by the canopy of surrounding trees, which might contribute to elongation suppression. Such findings seem to correspond with the observations of Miciniak and Zątek [24], who demonstrated that individuals of Lilium martagon growing in the dry-mesic oak forest reached greater height on average than those occurring in the oak-hornbeam forest. Also, the findings of Pindel [63] demonstrated that the stems of Lilium martagon are much shorter in localities where light availability is slight than in not shaded stands.
Although the increase in flower number observed from population 2 via population 3 to population 1 increased the visibility of inflorescences and chances for successful pollination and seed production, it should be pointed out that the recorded mean number of flowers in the studied populations was rather low and corresponds with the observations of Bednorz [23] and Gazda and Gazda [29], while the greater number of flowers observed in the year 2019 in population 1 was close to mean values which were noted by Truchan [65] in a manor park. At the same time, it is worth adding that in populations occurring in manor parks [64] and in grasslands [10,69], the mean values of number of flowers per stems are much greater.
4.4. The Temporal Variability of Traits of Stems of Lilium martagon
The significantly greater values of selected individual traits observed in the year 2019 in immature stems (height of stems in populations 2 and 3, number, length and width of whorl leaves in population 1) and in generative stems (height of stems, number of flowers and whorls in population 1, number and length of whorl leaves in population 3) might suggest the favourable impact of weather conditions (specifically substantial precipitation and air temperatures) during the meteorological spring and summer on the growth of Lilium martagon stems. The aforementioned findings seem to correspond with the experimental studies of Khashroum et al. [94], who demonstrated the positive effect of substantial water irrigation among others on the length of Lilium martagon stems and flower number.
On the other hand, performed investigations showed that several traits did not exhibit temporal variability at least in one population. The lack of a unified trend in the studied populations might suggest the occurrence of site-specific temporal variability of individual traits. Micro-environmental soil conditions, such as nutrient availability (determined by the interplay of various nutrients and soil characteristics such as pH, texture and organic matter) might fluctuate during the growing season and may differ among sites. The seasonal fluctuations in nutrient availability were evidenced by Díaz-Raviña et al. [95], while variability among study sites was confirmed among others in pine forests [96] and beech forests [97].
5. Conclusions
The observed spatial variability of the studied populations allows the conclusion to be drawn that substantial soil moisture, considerable light intensity at ground level, as well as slightly acidic soil pH and moderate nitrogen content favour the development and growth of Lilium martagon individuals. On the other hand, factors negatively affecting its growth are represented by the lack of microsites suitable for seedling recruitment, over shading of stems by the tree canopy, the presence of shallow soils, as well as human and herbivorous animal activity contributing to damage of the above- and/or belowground parts of individuals. The study observations suggested that considerable precipitation during the meteorological spring (especially in May) and substantial values of air temperatures during the meteorological spring and summer (particularly in June) contribute to the growth of Lilium martagon stems. However, the lack of a unified trend in all the studied populations might indicate the occurrence of site-specific temporal variability of individual traits.
Moreover, the obtained results allow us to evaluate the state of the observed populations. The most abundant population, population 3, located in an oak-hornbeam forest Tilio cordatae–Carpinetum betuli in Hrabeński Forest, presents the greatest prospects for persistence in the occupied site. However, it is highly necessary for artificial openings to be made in the dense plant cover to create suitable microsites for seedling recruitment and development of juvenile individuals. Although stems representing all developmental stages were noticed in population 1, situated in the oak-hornbeam forest Tilio cordatae–Carpinetum betuli in Wolski Forest, and in population 2 in the beech forest Dentario glandulosae–Fagetum in Mount Chełm, the state of these populations seems unsatisfactory due to their much lower abundance. Furthermore, in more shaded areas, it is recommended to remove individual trees and shrubs to ensure 5–25% light availability for the herb layer.
All studied populations are located in protected areas, although their protection regimes vary. In the case of Mount Chełm Nature Reserve and Hrabeński Forest (Natura 2000 Hrabeński Forest PLH180039 site), passive conservation protection appears to be an effective strategy for the preservation of the Lilium martagon population, especially since these sites are located far from built-up areas. However, in Wolski Forest (Bielańsko-Tyniecki Landscape Park), a low protection regime is ineffective in counteracting the increased human impact. Unfortunately, the proximity of a large urban area contributes to the excessive trampling and destruction of vegetation, including the Lilium martagon population. Therefore, it is necessary to implement constant monitoring and channel tourist traffic through appropriately designated paths and trails that would bypass the Lilium martagon site.
Considering the obtained results, it can be concluded that long-term (covering at least several years) observations of populations of Lilium martagon should be continued. Such laborious, demanding, and time-consuming investigations provide the essential data supplementing the current state of knowledge and are crucial for better protection of this rare species. Of particular value would be comparisons of population and individual traits of Lilium martagon occurring in different habitats, including forests, grasslands, and anthropogenic localities.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhou J. An R. Huang X. Genus Lilium: A review on traditional uses, phytochemistry and pharmacology J. Etnopharmacol.202127011385210.1016/j.jep.2021.11385233485985 · doi ↗ · pubmed ↗
- 2Zhou N. Miao K. Liu C. Jia L. Hu J. Huang J. Ji Y. Historical biogeography and evolutionary diversification of Lilium (Liliaceae): New insights from plastome phylogenomics Plant Divers.20244621922810.1016/j.pld.2023.07.00938807906 PMC 11128834 · doi ↗ · pubmed ↗
- 3Fornaris A. Chiavazza P.M. Devecchi M. The importance of lilium in garden design: History and future Acta Hortic.2011900596410.17660/Acta Hortic.2011.900.5 · doi ↗
- 4Marasek-Ciolakowska A. Sochacki D. Marciniak P. Breeding Aspects of Selected Ornamental Bulbous Crops Agronomy 202111170910.3390/agronomy 11091709 · doi ↗
- 5van Tuyl J.M. Arens P. Shahin A. Marasek-Ciołakowska A. Barba-Gonzalez R. Kim H.T. Lim K.-B. Lilium Ornamental Crops, Handbook of Plant Breeding Van Huylenbroeck J. Springer Berlin/Heidelberg, Germany 201848151210.1007/978-3-319-90698-0_20 · doi ↗
- 6Piękoś-Mirkowa H. Mirek Z. Flora Polski. Atlas Roślin Chronionych Multico Oficyna Wydawnicza Warszawa, Poland 20031584
- 7Zarzycki K. Trzcińska-Tacik H. Różański W. Szeląg Z. Wołek J. Korzeniak U. Ecological Indicator Values of Vascular Plants of Poland Biodiversity of Poland Mirek Z. W. Szafer Institute of Botany, Polish Academy of Sciences Krakow, Poland 2002 Volume 21183
- 8ChytrýM. TichýL. Dřevojan P. Sádlo J. ZelenýD. Ellenberg-type indicator values for the Czech flora Preslia 2018908310310.23855/preslia.2018.083 · doi ↗