Plant Non-Specific Lipid Transfer Proteins (nsLTPs): Comprehensive Functional Analysis and Defense Mechanisms
Bikram Giri, Dhirendra Kumar

TL;DR
This paper reviews plant nsLTP proteins, which help transport lipids and support plant defense and stress responses.
Contribution
The paper offers a comprehensive and updated analysis of nsLTP evolution, localization, and roles in plant defense.
Findings
nsLTPs are involved in lipid transport, stress responses, and signaling in plants.
nsLTPs have a conserved structure with eight cysteine residues and a hydrophobic cavity.
nsLTPs interact with signaling molecules to modulate stress-related gene expression.
Abstract
Non-specific lipid transfer proteins (nsLTPs) play a crucial role in transferring lipids across plant tissues and membranes. They are important for plant defense, particularly under stressful environmental conditions. They achieve this by coordinating signaling pathways and regulating the plant’s antioxidants and hormone systems. nsLTPs are broadly classified into two main groups, nsLTP1 and nsLTP2, based on their molecular size. However, their specific functions remain poorly understood. This paper provides a comprehensive review of nsLTPs, covering their evolution, localization in plants, and biological roles, providing a clear understanding of these important proteins. Non-specific lipid transfer proteins (nsLTPs) play a crucial role in lipid transport across membranes, contributing to cellular integrity and structural stability. These proteins are characterized by the presence of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
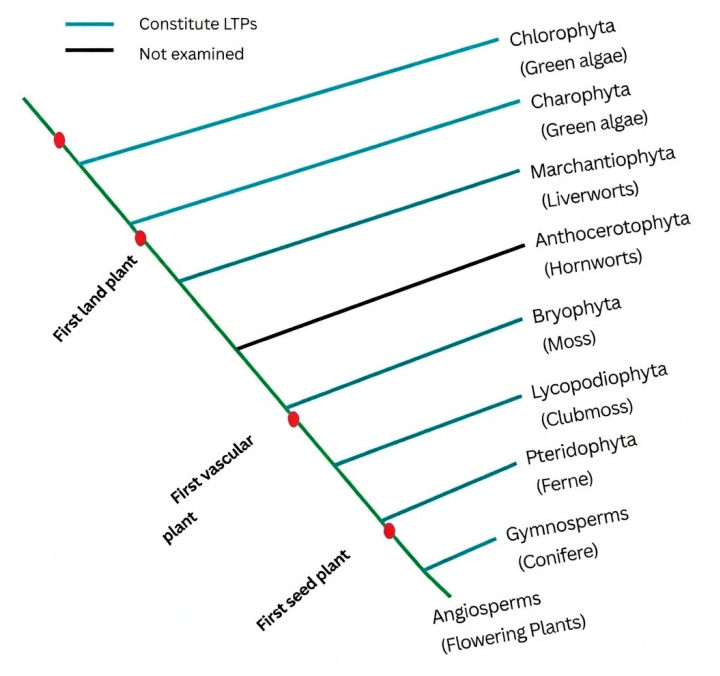 Figure 1
Figure 1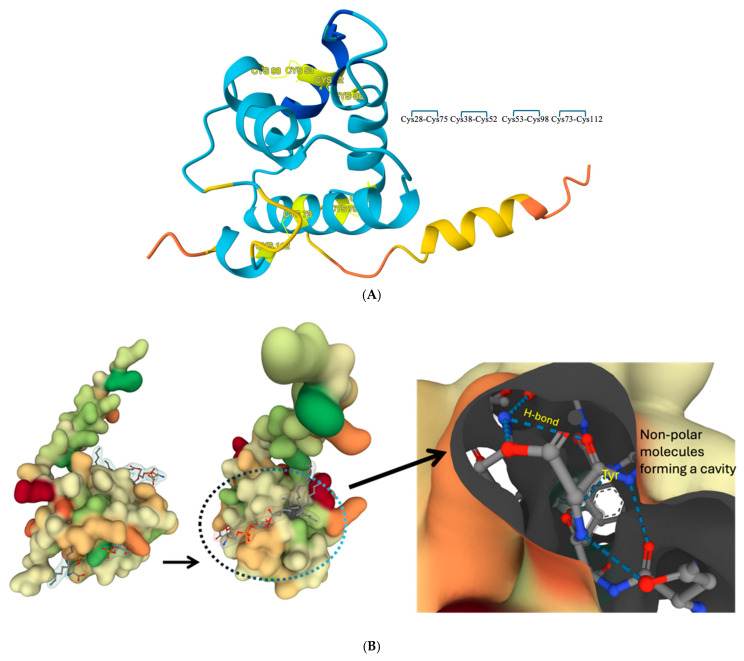 Figure 2
Figure 2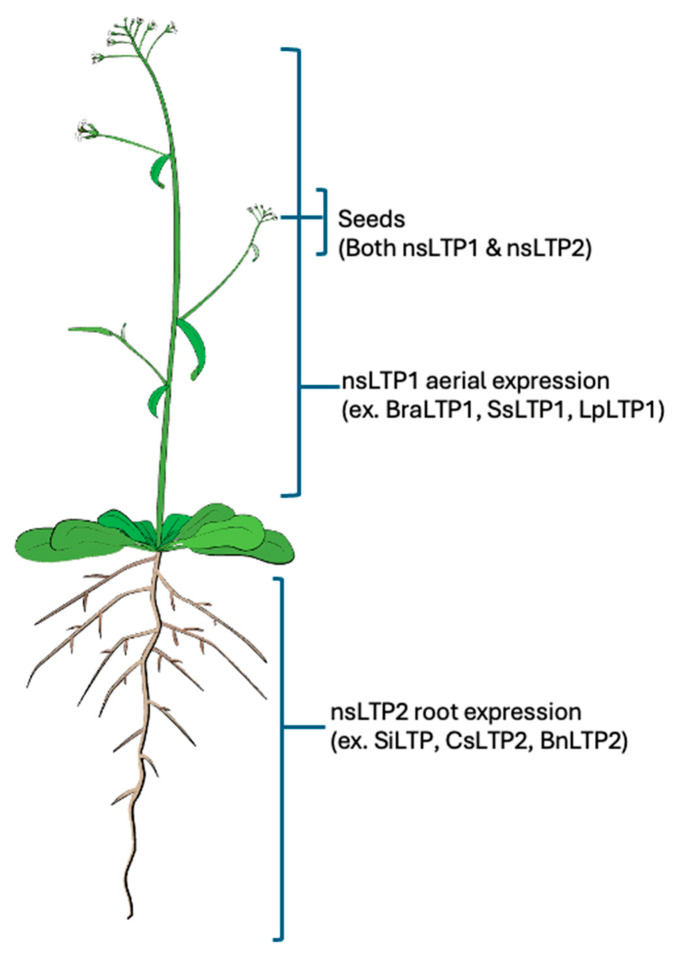 Figure 3
Figure 3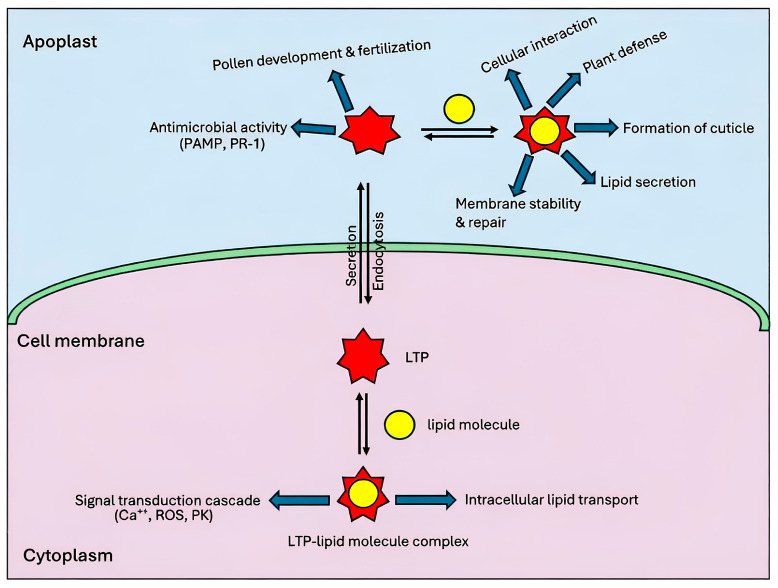 Figure 4
Figure 4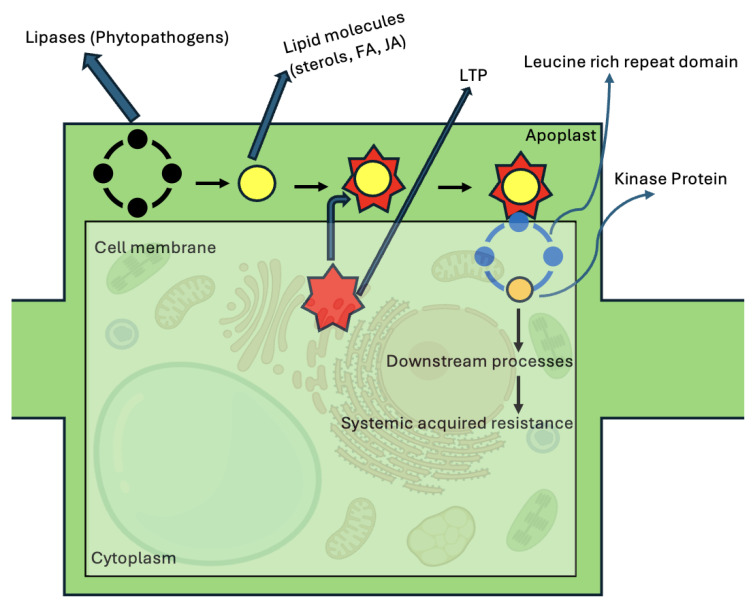 Figure 5
Figure 5- —National Science Foundation (NSF)
- —East Tennessee State University (ETSU)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLipid metabolism and biosynthesis · Lipid Membrane Structure and Behavior · Cellular transport and secretion
1. Introduction
Non-specific lipid transfer proteins (nsLTPs) are a unique family of proteins that transfer lipid molecules such as fatty acids, acyl-CoA, phospholipids, and glycolipids [1]. In addition to their basic function in lipid translocation, nsLTPs support important biological processes by linking structural configurations, evolutionary patterns, and subcellular localization. The primary research focus has been on the localization of nsLTPs and their roles in plant growth, development, and defense. Recent advances in gene-editing and breeding techniques across multiple traits and varieties have expanded opportunities to explore how nsLTP gene expression is regulated [2].
Most nsLTPs belong to multigene families. Members of these families exhibit functional diversity, as evidenced by expression across various tissues and at multiple stages of growth and development, and respond to stress [3]. All nsLTPs share several conserved traits, including small molecular size, stable domain structure, and high resistance to denaturing agents. Additionally, the presence of an N-terminal signal peptide facilitates their secretion and subcellular localization [4]. Despite our understanding of nsLTPs in plants, their precise biological roles, structural relationships, and lipid-transfer mechanisms remain poorly studied. Further research is needed to elucidate their specific roles in plant development, stress responses, and metabolism. This review highlights recent advances in nsLTP research and examines their functional significance across diverse biological processes.
2. Evolution and Structural Confirmation of nsLTPs
Various reports suggest that the evolution of plant nsLTPs is linked to stress adaptation and defense [5,6]. As described [7], the first lipid transfer protein was isolated from potatoes. Later, spinach leaves were used to purify and characterize nsLTPs, which were named for their ability to mediate in vitro phospholipid transfer between membranes [6]. Recent genome-wide characterization of nsLTP gene families has revealed 79 members in Arabidopsis, 156 in wheat, 64 in tomato, 58 in sorghum, 63 in maize, 77 in rice, and 189 across three Gossypium species [8,9]. Most nsLTPs isolated and characterized so far belong to flowering and non-flowering plants (Figure 1) [5]. The conservation of nsLTP gene families is largely attributable to gene duplication in angiosperms [10]. The nsLTPs are encoded by multi-gene families with more than 40 members in flowering plants and 50 members in bryophytes and ferns. Further, phylogenetic analysis has significantly advanced our understanding of the evolution and functionality of the nsLTP family [8]. However, in non-flowering plants, low sequence homology has challenged the classification of nsLTPs [5].
Despite variations in amino acid sequence, identities among nsLTPs range from 50 to 90% across plant species. Most nsLTPs share common characteristics, including small protein size (6–15 kDa) and presence of an eight-cysteine motif (8CM) arranged as C-Xn-C-Xn-CC-Xn-CXC-Xn-C-Xn-C, where C represents ‘Cysteine’ and ‘Xn’ is an amino acid variable number (Figure 2A) [11]. They possess an N-terminal signal peptide that directs the protein to specific subcellular locations, and a hydrophobic tunnel-like cavity, formed by four disulfide bridges, facilitates the lipid transfer (Figure 2B) [5,12].
Although computational modeling has been instrumental, the pivotal advances in understanding the structural biology of nsLTPs have stemmed from experimental structural elucidation. As of January 2026, the Protein Data Bank (PDB) contains 25 nsLTP structures, determined by techniques such as X-ray diffraction and Nuclear Magnetic Resonance (NMR) spectroscopy. A distinct characteristic of nsLTPs is a hydrophobic tunnel, noted for its adaptability, which accommodates single- and double-chain lipids [12]. In most cases, following ligand binding, such as phosphatidylglycerol and 1,2-dimyristoyl phosphatidylglycerol, the helix position of the N-terminal and C-terminal in 8CM expands the hydrophobic cavity by two to four times, reflecting the intrinsic mechanism of lipid binding and transfers [11,13]. The binding of lipid molecules occurs through hydrophobic interactions within the cavity. Primarily, non-polar residues, such as leucine, isoleucine, phenylalanine, and valine, engage with lipid tails. The ligand binding of phosphatidylcholine to LTP12 is shown in Figure 2B, where a hydrogen bond forms between the ligand carboxyl group and the hydroxyl group of tyrosine. The confirmational flexibility studied using molecular dynamics (MD) simulations and crystallographic studies highlights cavity expansion and binding of fatty acids like oleic acid and myristic acid [13]. This observation is further supported by studies involving maize (ZmLTP1.6), rice (OsLTP1.18), and tobacco (NtLTP1.1) nsLTPs, where the cavity orientation changes upon binding of the ligand to the protein driven by differences in sequence loop regions on H1-H2 [14,15,16,17].
The exchange of lipids occurs between organelles where nsLTPs are found at membrane contact sites (MCS). Transport simulations suggest a non-vesicular shuttle mechanism, where nsLTPs do not require metabolic energy to transfer lipids as lipid contact hydrophobic cavity rather than aqueous environments, which are less favorable [18,19]. Although some of the lipids are transported against a concentration gradient requiring energy, as in ATP-binding cassette transporters [20,21,22,23]. While information on in vivo transport lacks direct observation, there has been in vitro evidence suggesting involvement of MCS in efficient lipid trafficking [24]. MCS also shortens the diffusion distance for nsLTPs, accelerating lipid transfer [25]. As discussed earlier, disulfide bonds stabilize the nsLTPs to maintain a rigid extracellular alpha-helical structure for stable binding of hydrophobic lipid signals, such as oxylipins, within the internal cavity. The MCS helps in transporting lipid cargo at the plasma membrane by leucine-rich repeat receptor-like kinases (LRR-RLKs). The ligand recognition complexes promote the receptor conformational change, triggering kinase activation and downstream signaling, involving activation of mitogen-activated protein kinase (MAPK) cascade and the activation of systemic acquired resistance (SAR) [26,27]. Nevertheless, whether nsLTPs act as lipid shuttles or multifunctional scaffolds for signaling remains unclear, and resolving this requires integrating studies using in vivo imaging, functional assays, and advanced structural dynamics (EM/NMR/MD simulations).
3. Classification of Plant nsLTPs
In plants, nsLTPs are small, basic proteins and account for up to 4% of total soluble proteins [7]. Initial classification of nsLTPs was based on molecular mass. The nsLTP1 (Type I) family members were around 10 kDa, and the nsLTP2 (Type 2) were 7 kDa proteins [14]. The classification of nsLTPs was regularly updated based on structural features, sequence similarity, hydrophobic cavities, and other broad classifications [4,8,28,29,30,31,32]. Boutrot has developed a classification based on sequence similarity and cysteine residue spacing, with roles in SAR supported by antimicrobial and stress regulatory motifs. Based on this, the nsLTPs from rice (Oryza sativa L.), Arabidopsis thaliana, and wheat (Triticum aestivum L.) were classified into nine distinct types (Types I–IX) (Table 1) [28]. This was later expanded to include Types X and XI. Type X nsLTPs represented a new group that included nsLTPs from Solanaceae [31].
The classification was further refined by Edstam et al. [5], incorporating the presence of the GPI anchor (Types 1, 2, C-K). Subsequently, Wang et al. [4] proposed a revised system based on the pattern of the 8CM and categorized the nsLTPs into Types I–V. More recently, Fleury et al. [8] combined sequence alignment, phylogeny, and structural biochemistry to classify nsLTPs into two types, Type I and Type II, as suggested by Boutrot et al. [28]. A 2023 study by Huang et al. [33] described a new class of nsLTP from algae distinguished by an extended 8CM spacing, larger molecular mass (10.36–50.28 kDa), and lower pI (<8). Further analysis is needed to determine if this new group of nsLTPs falls into either the updated group of Type I or II described by Fleury et al. [8].
Compared to early classification model systems, the newer approaches are based on the spacing of eight cysteine motifs, the presence of glycosylphosphatidylinositol (GPI) anchors, and the use of advanced bioinformatics tools such as AlphaFold, molecular docking, and PDB’s pairwise structural alignment. Using the computational approach will help to characterize novel subfamilies of nsLTPs and understand diverse roles in defense and plant physiology. The nsLTP classification systems are summarized in Table 1.
4. Spatial Expression and Cellular Localization of Plant nsLTPs
The cellular expression of nsLTPs varies among plant species, providing diverse biological functions. Different plant organs are known to express nsLTPs, including seeds, leaves, stems, roots, flowers, and fruits [21]. The expression profile and localization differ among nsLTP types, with nsLTP1s present in cuticle-covered epidermal cells and embryonic and vascular tissues [22]. The most widely recognized and abundant type, nsLTP1, is found in the aerial organs of plants, while nsLTP2s are expressed in roots (Figure 3). Seeds contain both nsLTP types [23]. In seeds, diverse physiological functions are supported by multiple localizations within the cell wall, plasma membrane, and extracellular space [24].
Data from diverse species have provided crucial information on the localization of nsLTPs and their role in plants. The extracellular localization of some nsLTPs has been reported, including in barley [25], carrot [26], Arabidopsis [27], tobacco [28], and soybeans, having a potential role in forming the cuticle barrier and antimicrobial property [29]. Additionally, research has challenged the apoplastic protein tag, revealing that nsLTPs are expressed at the plasma membrane, exhibit intracellular localization, and associate with the intracellular matrix, providing intracellular lipid transport and defense signaling. The barley, Arabidopsis, grapevine, tobacco, and rice nsLTPs are localized outside the cell [30]. While some nsLTPs from castor bean seeds (RcLTP) are intracellularly localized (glyoxysomes), facilitating intracellular lipid mobilization, the cowpea seed nsLTP (VuLTP) is in vacuoles, having a role in lipid homeostasis, and the pepper seed nsLTP (CaLTP1) is in vesicles and the cell wall involved in lipid trafficking [31].
In some cases, nsLTP localization is dynamic. Based on cell shape and cell wall curvature during cell growth and differentiation, the A. thaliana AtLTPg localizes to the apoplast, plasma membrane, and cell wall, assisting in cuticular wax deposition and cell wall fortification [34]. Similarly, HaAP10, an nsLTP from Helianthus annuus seeds, localizes to the apoplast and plasma membrane in dry seeds and upon imbibition, relocates to the intracellular matrix, highlighting the versatility of nsLTPs in adapting lipids and regulating subcellular trafficking [35]. Additionally, the cellular localization of type G nsLTPs (LTPGs) is regulated by alternative splicing, resulting in two distinct transcripts. One contains the GPI anchor signal for cuticle integrity and lipid deposition, whereas the non-anchored isoform potentially supports antimicrobial defense and lipid shuttling. Splicing events enable differential localization of LTPGs in response to varying tissue types and environmental conditions [5,36]. Various approaches and methods have been used to investigate the subcellular localization of nsLTPs, as detailed in Table 2. Proteins identified as extracellularly localized were obtained from cell culture or from proteomic analysis of apoplastic fluid. Similarly, immunochemical studies highlighted the association of nsLTPs with cell wall locations. Furthermore, localization varies with developmental stage and environmental conditions. Therefore, a combination of experimental methods under different conditions is required to enhance the robustness of detecting the subcellular localization of nsLTPs [37]. Furthermore, future studies could use mass spectrometry-based lipidomics to understand organelle trafficking of lipids during stress, providing clues on their mobilization and interaction with nsLTPs.
5. Biological Significances of nsLTPs
In recent years, extensive research has elucidated the diverse functions of nsLTPs, revealing their involvement in numerous physiological responses (Figure 4). These proteins are implicated in growth and reproduction, defense against pathogenic microbes, symbiosis, stress adaptation, and antimicrobial activity [55]. However, the precise mechanisms by which nsLTPs operate across these processes remain poorly understood.
5.1. Defensive Functions of nsLTPs Under Biotic Stress
Numerous studies have reported the expression of nsLTPs in response to attacks by bacterial and fungal pathogens, such as wheat, potato, and rice [56,57,58]. nsLTPs exhibit antimicrobial and antifungal properties, inhibiting the growth of pathogenic bacteria and fungi, including Pseudomonas solanacearum, P. syringae, Fusarium solani, Alternaria brassicola, and Botrytis cinerea [59,60]. The antimicrobial activity of nsLTPs is effective against a specific range of microorganisms. For instance, nsLTPs from Arabidopsis, radish, and onion have demonstrated antimicrobial activity at micromolar concentrations. McLaughlin et al. reported that overexpressing AtLTP4.4 in transgenic wheat significantly reduced the growth of the pathogen Fusarium graminearum, a pathogen that causes diseases in small grain cereals [59]. Additionally, Ace-AMP1 from transgenic wheat exhibits an antifungal response to Blumeria graminis [61], while rice and onion have demonstrated both fungal and pathogen resistance [62]. Further findings revealed that Durum wheat nsLTP1 (TdLTP4) exhibits anti-inflammatory, antimicrobial, and antifungal properties and plays a crucial role against human and foodborne pathogenic bacteria [63]. Interestingly, nsLTPs also synergize with other antimicrobial peptides, such as defensins and thionins, forming a broader network of immunological proteins in plant physiology [64]. Furthermore, BrLTP2.1 from Brassica rapa found in nectar displayed both antifungal and antimicrobial activities [65].
Plant immunity often involves recognition of pathogen attacks via pathogen-associated molecular pattern (PAMP) signaling, triggered by bacteria and fungi, which in turn elicits a defense response mediated by hydrophobic ligands (Figure 4) [66]. Additionally, nsLTPs in plants are categorized as pathogenesis-related proteins within the PR14 family, which are specialized for plant defense [36]. Specifically, type IV nsLTPs in Arabidopsis form complexes with glycerol-3-phosphate, facilitating their translocation and signaling induced SAR, providing broad-spectrum protection against pathogens [67]. This function highlights the essential role of nsLTPs in plant immune response pathways, contributing to effective resistance to pathogenic attacks. Similarly, overexpression of nsLTP genes such as CaLTP and CaLTP2 in pepper, LjAMP1 and LjAMP2 in motherwort, and nsLTP in barley has shown significant resistance to bacterial and fungal pathogens [68,69].
Furthermore, signaling molecules such as abscisic acid, salicylic acid, ethylene, and methyl jasmonate trigger a signaling cascade that regulates nsLTP gene expression. Recent studies have identified the involvement of jasmonates with various saturated and unsaturated fatty acids in LcLTP2 (Lens culinaris) and BrLTP2 (Brassica rapa) to stimulate defense response [65]. Interestingly, research on wheat LTP1 has shown that it interacts with elicitin, an elicitor protein from oomycetes, potentially competing for binding sites on tobacco cells for plant defense [64]. The interaction, enhanced by the presence of jasmonic acid (JA), has been linked to increased resistance against the pathogen Phytophthora parasitica [70]. Nonetheless, many such findings are based on in vitro experiments and may behave differently in vivo.
5.2. Defensive Functions of nsLTPs Under Abiotic Stress
nsLTPs enable plants to adapt to diverse environmental conditions, such as high salinity [71,72], drought [45,73], and freezing stress [43,74]. In bread wheat (Triticum aestivum), TaLTP3 is induced by cold, drought, ABA, and oxidative stress [75]. The transcription factor MYB96 regulates the expression of various nsLTP genes, including LTP3, coordinating plant responses to freezing and drought stress in Arabidopsis [45]. Stress-responsive motifs (TGA, TCA, and ABRE) and signaling cascades with ROS homeostasis in HcnsLTP111 are reported to regulate drought and stress tolerance in Kenaf (Hibiscus cannabinus) [76]. Further, overexpression of nsLTPs, such as OsDIL in rice, LTP3 and AZI1 in Arabidopsis, and CaLTP1 in pepper, significantly enhances tolerance to abiotic stress.
Across multiple monocot and dicot species, such as rice, wheat (TaLTP40, TaLTP75, TdLTP4, TdLTP2), Arabidopsis (AtGBF3), cabbage, tobacco (NtLTP4), foxtail millet (SiLTP), nsLTP overexpression has been shown to enhance tolerance to various abiotic stresses [77,78,79,80,81]. The cabbage-derived nsLTP WAX9 also provides cryoprotection to isolated spinach thylakoids [82]. In addition, plants overexpressing nsLTPs altered sensitivity to key stress-related hormones, including SA, JA, and ABA [64,83]. Recent findings by Zhou et al. showed that three members of wheat nsLTP, located on chromosomes 1B, 5D, and 7B, enhance protein abundance and mediate salt tolerance in the wheat overexpression line TaNRX1-2D [84].
5.3. Role of nsLTPs in Cuticular Wax Deposition
Long-chain fatty acids (C20–34) and their derivatives make up waxes, whereas a polymer network of C16 and C18 fatty acids makes up plant cutin layers [85]. Insulation, stomatal water loss prevention, radiation damage reduction, and defense against pathogen attacks, including herbivores, are all provided by these structures. Notably, lipid metabolism in epidermal cells primarily targets cuticular lipids [86]. Plant colonization success relies on molecular barriers formed by lipid polyesters [12]. Research has shown that cuticular and wax polymer synthesis occurs in the apoplast, potentially facilitated by nsLTPs trafficking monomers [87,88]. Interestingly, nsLTP expression is notably higher in young, developing tissues synthesizing surface wax in species such as Brassica oleracea and Arabidopsis [39]. Within the extracellular matrix, cell walls, or plasma membrane of epidermal cells and secretory tissues, nsLTPs contribute to cutin monomer deposition [89,90]. LTPGs, which contain GPI anchors, are implicated in the biosynthesis and accumulation of suberin and cutin monomers, and disruption of GPI-anchored gene LTPG1 in Arabidopsis resulted in altered cuticular lipid composition, increasing susceptibility to fungal infection [47,91].
Furthermore, nsLTP expression increases in response to environmental stressors, including drought, heavy metal exposure, and heat stress, and correlates with wax deposition [92,93]. Under stress conditions, broad classes of nsLTP1 and nsLTP2 enhance cuticular layer deposition and thickening, preventing pathogen attack with antimicrobial properties [94]. In Solanum lycopersicum (Sltpg3), nsLTPs delay water loss by thickening the cuticle, reducing permeability, and softening tomato fruits for a longer shelf life [95]. Another study suggests that Brassica napus BraLTP1 plays a role in enriching epicuticular wax deposition [96]. The Arabidopsis AtLTP2 and AtLTP4 were found to link nsLTPs to cuticular wax deposition and to control water permeability [48]. Conversely, knocking out nsLTP expression alters the cuticle’s lipid composition and the density of the cuticle layer [89,97]. Despite these insights, the precise mechanisms by which nsLTPs transport lipid components to form the cuticle remain unclear.
5.4. Involvement of nsLTPs in Seed Development
Expression of nsLTPs increases significantly during seed germination and development. In rice, OsLTPL23 and OsLTPL18 are key regulators of seed germination [98]. In Coffea arabica and C. canephora, nsLTP expression is observed in the endosperm and embryo during seed germination [99]. The biochemical and physiological functions of nsLTPs in seed germination include mobilizing stored lipids through the regulation of fatty acid beta-oxidation [42]. Sesame LTP, SiLTP1.23, and SiLTP1.28 (Sesamum indicum) play a crucial role in oil accumulation in seeds [9]. Additionally, research from [100] showed that ElLTP1 and ElLTP2 in Euphorbia lagascae act as protease inhibitors, protecting cotyledons from proteolysis during programmed cell death. In sunflower (Helianthus annuus), HaAP10 relocates to glyoxysomes, the organelles involved in lipid metabolism, after seed imbibition [35]. Five lipid transfer and storage proteins were expressed (ACP, ACBP, DIR1, FPKM), regulating the wax ester synthesis pathway for seed development in Jojoba [101]. Furthermore, nsLTPs are involved in pollen development, as demonstrated by recent studies in chili pepper, Arabidopsis, maize, wheat, and rice [44,102,103]. However, the precise pathways and mechanisms through which nsLTPs mobilize lipids during seed development remain not fully understood.
5.5. nsLTPs as Modulators of Plant Signal Transduction
By forming complexes with lipid molecules, nsLTPs play a crucial role in initiating and regulating a variety of signaling pathways in plants. After binding to nsLTPs, these lipid molecules interact with receptors such as serine/threonine protein kinases, which have a transmembrane region, extracellular leucine-rich repeat (LRR) domains, a transmembrane region, and the kinase (PK) domain towards the cytosol. This interaction initiates a signal transduction pathway with adaptable second messengers. Transcription factors, protective agents, pathogenesis-related (PR) proteins, and other antimicrobial peptides (AMPs) are stimulated by the activation of the MAPK cascade [104]. SAR is induced by this chain of events (Figure 5) [30].
One signaling molecule, the oxylipin, is produced from unsaturated fatty acids in response to reactive oxygen species (ROS) and plays a crucial role in enhancing plant growth and development under stress conditions [105,106]. ROS scavenging activity protects plants from oxidative stresses induced by pathogen infections. nsLTPs are directly or indirectly involved in ROS scavenging as shown by studies involving NtLTP1.38, NtLTP2, NtLTP25, and the overexpression of Type I nsLTP in Nicotiana benthamiana (NbLTP1), conferring resistance to tobacco mosaic virus (TMV) [77,107,108,109,110]. In barley, the combined action of LTPs and oxylipins is known to protect plant cells under stress by neutralizing toxic components [111]. Similarly, in Cucumis sativus, CsnsLTP6 overexpression confers resistance against Corynespora cassiicola [112]. This is achieved first through activation of defence signaling pathways and then by rapidly activating the ROS-scavenging enzymes. The LTP2 (DIR-1) in Arabidopsis and glycerol-3-phosphate (G3P) are involved in SAR development and long-distance signaling [113]. These nsLTPs bind to lipid molecules, for example, oxylipins, fatty acids, produced by lipases during pathogen infection and trigger a signaling cascade that leads to the SAR response (Figure 5) [114,115]. Furthermore, the xylogen culture of Zinnia, which has a GPI anchor, binds to plant sterols and is characteristic of Type II nsLTPs [116]. This protein participates in signaling pathways and intercellular interactions. It is suggested that similar proteins in other plants, which have nsLTP domains resembling those of nsLTP2s, may also be involved in intercellular interactions and signal transduction, functioning within a complex that includes a lipid molecule [116]. The MAPK cascade is involved in response to pathogens mediated by nsLTPs, such as StLTP10 [117], and in Arabidopsis, where both camalexin and antimicrobial phytoalexin production are triggered by MAK3/MPK6 signaling [72,118]. As described earlier, nsLTPs contribute to the lipid exchange between organelles at MCS, highlighting the dynamic shuttling of lipids to coordinate cellular resilience and signal immune activation.
6. Precise Gene Editing Technology: Future Directions
Despite our understanding of the types and subtypes of nsLTPs, the precise role of nsLTPs under different stress conditions remains poorly elucidated. Researchers still face challenges posed by the absence of information on tissue-specific expression, structural differences in nsLTPs, and the functional diversity of nsLTPs during development. Finally, advances in precise gene-editing technologies have started to circumvent these limitations [119]. The use of CRISPR/Cas9 has changed the traditional approach to plant editing, offering greater precision and efficiency [120]. CRISPR/Cas9 can be used to multistack gene knockout of the nsLTP family to understand their precise role. For instance, gain- and loss-of-function mutants in sesame (SiLTP1) have provided critical insights into seed oil accumulation [121], while CRISPR-mediated lysine malonylation in Dendranthema grandiflorum DgnsLTP1 resulted in resistance to cold [122]. Furthermore, the targeted modification of nsLTP genes through CRISPR could enhance plant resilience to biotic and abiotic stresses [75,112,123].
7. Conclusions
In summary, research on nsLTPs has increased over the years; however, their specific functions and localization-based roles remain incompletely characterized. Studies using overexpression and knockout lines have demonstrated the role of nsLTPs in mediating stress responses, highlighting their involvement in lipid binding and transfer, and orchestrating complex signaling networks. As we continue to decode the nsLTPs, advanced genetic engineering techniques and computational modeling are likely to provide an improved approach for the functional characterization of different nsLTPs across plant species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Carvalho Ade O. Gomes V.M. Role of plant lipid transfer proteins in plant cell physiology—A concise review Peptides 2007281144115310.1016/j.peptides.2007.03.00417418913 · doi ↗ · pubmed ↗
- 2Liang Y. Huang Y. Chen K. Kong X. Li M. Characterization of non-specific lipid transfer protein (ns Ltp) gene families in the Brassica napus pangenome reveals abundance variation BMC Plant Biol.2022222110.1186/s 12870-021-03408-534996379 PMC 8740461 · doi ↗ · pubmed ↗
- 3Dos Santos-Silva C.A. Ferreira-Neto J.R.C. Amador V.C. Bezerra-Neto J.P. Vilela L.M.B. Binneck E. de Santana Rêgo M. da Silva M.D. de Melo A.L.T.M. da Silva R.H. From Gene to Transcript and Peptide: A Deep Overview on Non-Specific Lipid Transfer Proteins (ns LT Ps)Antibiotics 20231293910.3390/antibiotics 1205093937237842 PMC 10215178 · doi ↗ · pubmed ↗
- 4Wang N.-J. Lee C.-C. Cheng C.-S. Lo W.-C. Yang Y.-F. Chen M.-N. Lyu P.-C. Construction and analysis of a plant non-specific lipid transfer protein database (ns LTPDB)BMC Genom.201213 S 910.1186/1471-2164-13-S 1-S 9PMC 330372122369214 · doi ↗ · pubmed ↗
- 5Edstam M.M. Viitanen L. Salminen T.A. Edqvist J. Evolutionary History of the Non-Specific Lipid Transfer Proteins Mol. Plant 2011494796410.1093/mp/ssr 01921486996 · doi ↗ · pubmed ↗
- 6Liu F. Zhang X. Lu C. Zeng X. Li Y. Fu D. Wu G. Non-specific lipid transfer proteins in plants: Presenting new advances and an integrated functional analysis J. Exp. Bot.2015665663568110.1093/jxb/erv 31326139823 · doi ↗ · pubmed ↗
- 7Kader J.-C. Lipid-transfer proteins in plants Annu. Rev. Plant Physiol. Plant Mol. Biol.19964762765410.1146/annurev.arplant.47.1.62715012303 · doi ↗ · pubmed ↗
- 8Fleury C. Gracy J. Gautier M.-F. Pons J.-L. Dufayard J.-F. Labesse G. Ruiz M. de Lamotte F. Comprehensive classification of the plant non-specific lipid transfer protein superfamily towards its sequence–structure–function analysis Peer J 20197 e 750410.7717/peerj.750431428542 PMC 6698131 · doi ↗ · pubmed ↗