Citric Acid Cycle Genes and Nutrigenetics
Anna Vesnina, Oksana Kozlova, Svetlana Ivanova, Alexander Prosekov

TL;DR
This review explores how genes in the citric acid cycle are linked to chronic diseases and how diet and nutrients may help prevent or treat these conditions.
Contribution
The paper compiles 45 years of research on TCA cycle genes, their mutations, and the restorative potential of specific nutrients.
Findings
Mutations in TCA cycle genes like ACO2 and IDH are linked to neurodegenerative and cancer diseases.
Nutrients like niacin and resveratrol can regulate TCA cycle activity at the genetic level.
Epigenetic modifications and diet influence mitochondrial function and TCA cycle activity.
Abstract
The citric acid cycle disruptions are implicated in the pathogenesis of chronic diseases, including diabetes, obesity, cancer, and cardiovascular conditions. Numerous publications link TCA cycle disorders to oncological, neurodegenerative, and osteoporotic diseases, and specific single-nucleotide polymorphisms have been proposed as potential markers. Nevertheless, lifestyle and diet have been strongly linked to risk factors for mitochondrial dysfunction; thus, preventive measures that minimize these risks are a relevant field of research. This review summarizes 45 years of relevant publications on the TCA cycle, its genetics and epigenetics, and the restorative potential of certain nutrients. The review includes articles in English and Russian, registered in PubMed, Elsevier, eLIBRARY.RU. The genes encoding the TCA cycle enzymes have been collected and presented. Information is provided…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
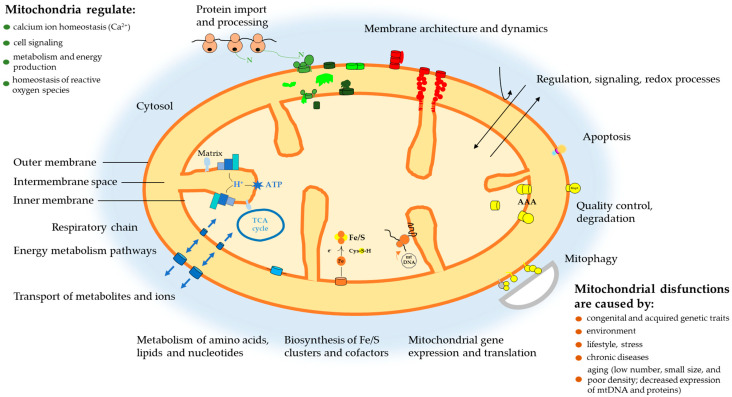 Figure 1
Figure 1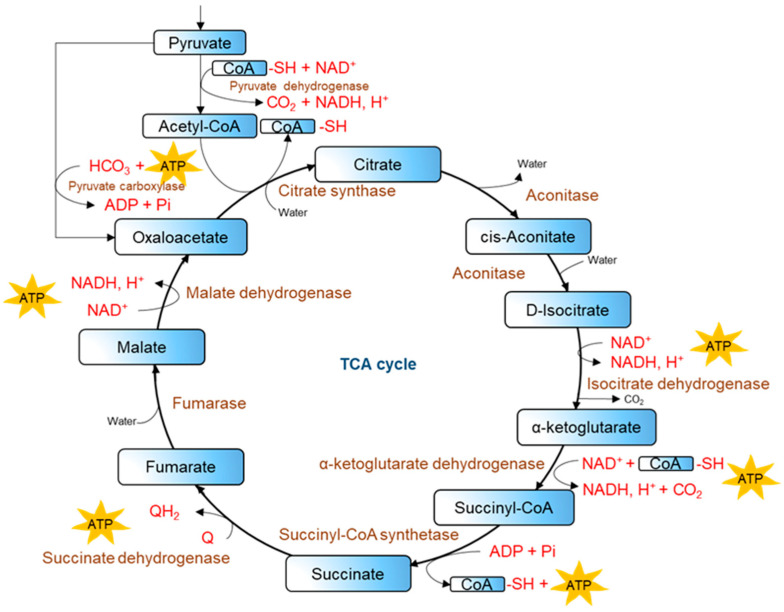 Figure 2
Figure 2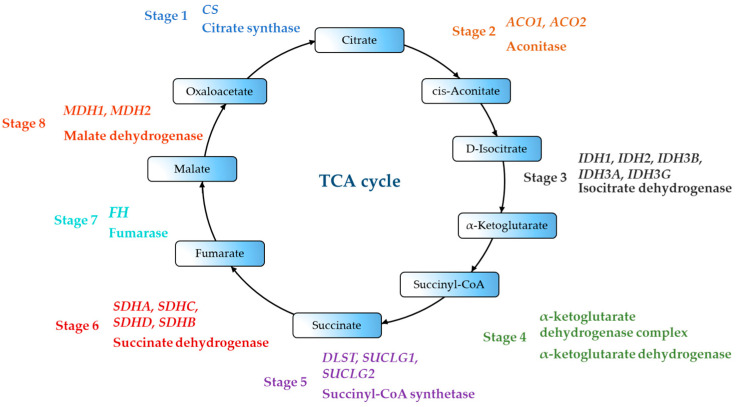 Figure 3
Figure 3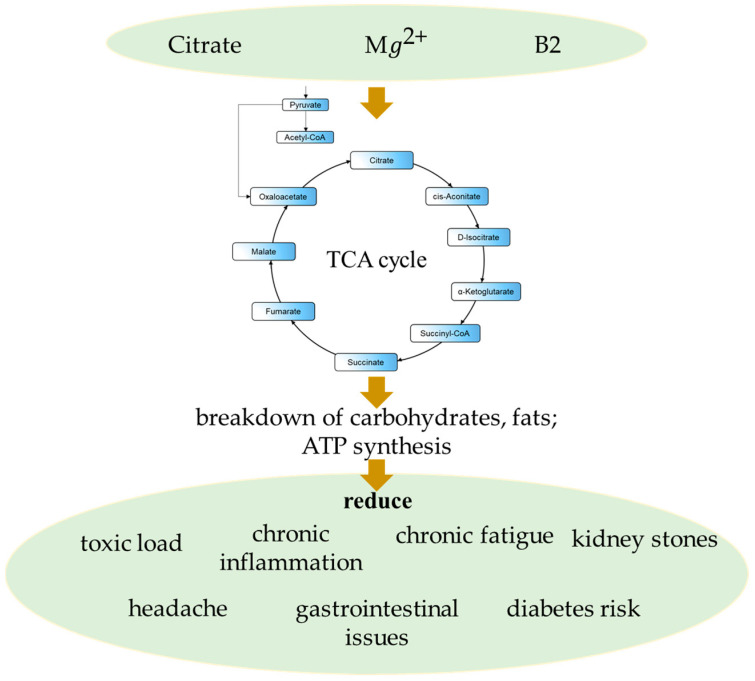 Figure 4
Figure 4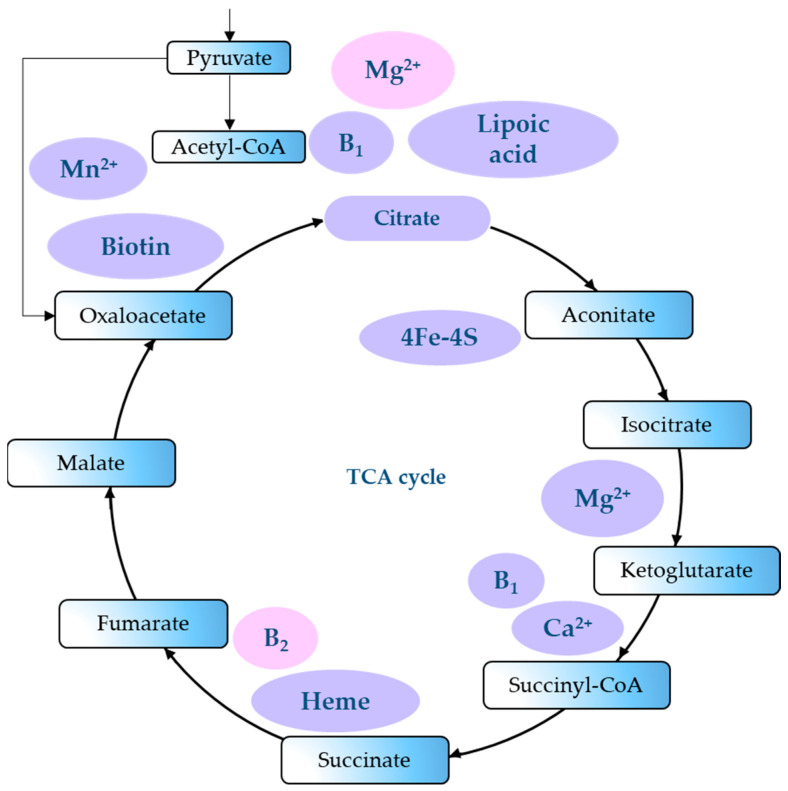 Figure 5
Figure 5- —Genomics and DNA Sequencing Laboratory, Kemerovo State University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBiochemical Acid Research Studies · Sirtuins and Resveratrol in Medicine · GDF15 and Related Biomarkers
1. Introduction
Mitochondria are crucial for cellular processes. They host two mechanisms that underlie the metabolic pathways of most living organisms. Oxidative phosphorylation (OXPHOS) takes place in mitochondria. The citric acid cycle, also known as the tricarboxylic acid (TCA) cycle or Krebs cycle, occurs in the mitochondrial matrix. Discovered in 1937 by H. A. Krebs and W. A. Johnson [1,2,3,4], the TCA cycle is a fundamental respiration stage for all living cells: it provides the body with energy and maintains metabolism (Figure 1) [1,5,6].
Disturbances in the TCA cycle cause mitochondrial dysfunction that may trigger obesity, diabetes, cardiovascular diseases, cancer, and premature aging. Moreover, it may be closely related to neurodegenerative conditions such as Alzheimer’s and Parkinson’s diseases [9,10,11,12].
A proper diet may prevent mitochondrial dysfunction and, consequently, a number of chronic diseases [13,14]. The ketogenic and Mediterranean diets positively impact mitochondrial function [15,16,17]. The Western diet, on the contrary, contributes to mitochondrial dysfunction due to plant fats, sugars, and monosodium glutamate [18].
This article is a review of the scientific publications on various issues related to the TCA cycle, e.g., its intermediates that regulate the epigenetic landscape of gene expression, nutrients and diets that affect the TCA cycle and cellular metabolism.
Generalization of the available data can contribute to a deeper understanding of the potential directions of dietary and metabolic correction that prevent violations of the tricarboxylic acid cycle. However, the practical implementation of preventive strategies requires further interdisciplinary research and accumulation of clinical evidence.
2. TCA Cycle: Concept and Role in Healthy Metabolism
The TCA cycle is a core part of cellular respiration: it is a cyclical pathway of enzymatic reactions inside mitochondria (Figure 2). As organic molecules of carbohydrates, proteins, and fats oxidize, they release energy in the form of adenosine triphosphate (ATP) [19].
As a TCA cycle component, pyruvate (pyruvic acid) appears during glycolysis. Glycolysis is a central metabolic pathway used by all cells during glucose oxidation to produce energy in the form of adenosine triphosphate and intermediates to serve other metabolic pathways [2].
Arnold et al. [2] described the processes of cataplerosis and anaplerosis. Cataplerosis is responsible for removing TCA cycle intermediates to support other metabolic pathways. Anaplerosis replaces these intermediates to maintain continuous oxaloacetate formation (Table 1).
Anaerobic glycolysis occurs without oxygen. Aerobic glycolysis involves oxidative decarboxylation of pyruvate to water and carbon dioxide in the presence of oxygen; i.e., decarboxylated pyruvate combines with coenzyme A and enters the TCA cycle [20,21].
The TCA cycle consists of eight stages, each with its own intermediate, namely citrate, isocitrate, oxoglutarate, succinyl-CoA, succinate, fumarate, malate, and oxaloacetate. The final products include carbon dioxide (CO_2_), nicotinamide adenine dinucleotide (NADH), flavin adenine dinucleotide (FADH_2_), and adenosine triphosphate (ATP) [19,22].
The TCA cycle reactions are carried out by such enzymes as citrate synthase (CS), aconitase (ACO2), α-ketoglutarate dehydrogenase (OGDH), succinate dehydrogenase (SDH), malate dehydrogenase (MDH), fumarase (FH), isocitrate dehydrogenase (IDH), and succinyl-CoA synthetase. Almost all enzymes of the tricarboxylic acid cycle are localized in the mitochondrial matrix. The exception is succinate dehydrogenase (SDH), which is associated with the inner mitochondrial membrane and simultaneously functions as part of the respiratory chain [19,23].
As part of its functions, the TCA cycle releases energy as adenosine triphosphate (ATP), produces electron carriers NADH and FADH_2_, participates in redox homeostasis, and synthesizes intermediates for metabolic pathways. Disturbances in the TCA cycle can alter cellular energy homeostasis and redox balance, potentially contributing to mitochondrial dysfunction and oxidative stress. Such metabolic alterations have been reported in association with neurodegenerative disorders, including Alzheimer’s disease [24]; however, current evidence mainly points to a correlation rather than a direct causal relationship. Furthermore, impaired TCA cycle progression can lead to the accumulation of intermediate metabolites such as 2-hydroxyglutarate (2-HG), succinate, and fumarate, which are known to exert epigenetic and signaling effects and are therefore termed oncometabolites [2,25,26,27]. Metabolic reprogramming involving TCA-related pathways has also been observed in osteoporosis, cardiovascular disease [28,29,30], and thyroid disease [3], although the underlying mechanisms remain poorly understood and are likely multifactorial.
De Castro Fonseca et al. [28] reported that succinate acts as an extracellular ligand that binds to GPR91, a G protein-coupled receptor associated with kidneys, liver, heart, retinal cells, and possibly many other tissues. The process may result in a wide range of physiological and pathological effects, which means that the TCA cycle is a crucial factor in healthy metabolism.
A disrupted TCA cycle and the resulting mitochondrial dysfunction are associated with poor nutrition. Excessive calorie consumption disrupts the balance of adenosine triphosphate, leading to oxidative stress, inflammation, and impaired cell apoptosis. Malnutrition or a diet low in vitamins B and minerals may disrupt the function of enzymes, leading to obesity, diabetes, cardiovascular diseases, joint and bone health issues, and autoimmune conditions [22].
A number of genetic traits pose a risk for the TCA cycle; a tailored diet may prevent mitochondrial dysfunction, which makes nutrigenomics a highly relevant field of medical research [7].
3. Genes That Encode TCA Cycle Enzymes
Figure 3 shows the genes encoding the enzymes of the TCA cycle.
The genes shown in Figure 3 are shown in Table 2. Table 2 shows the genes directly or indirectly involved in the TCA cycle.
Scientific evidence has been found that certain changes in the expression of TCA cycle genes influence the development of neurodegenerative diseases and oncology. In the work of J. Zhu [65], clinical studies have shown that variants Arg18Trp, Ser87Leu, Ala252Thr and Leu357Val of the ACO2 gene can be used as a biomarker in Parkinson’s disease. The work of R. Spiegel [66,67] has shown that the Ser112Arg mutation in the ACO2 gene leads to a neurodegenerative disease called cerebellar atrophy. Mitochondrial dysfunction is strongly associated with cancer [68]. Nölting et al. [69] linked TCA cycle gene mutations to pheochromocytoma or paraganglioma. Cho et al. [70] explained individual differences in energy balance by genetic polymorphisms in mitochondria, which affect energy metabolism at the cellular level. The team identified single nucleotide polymorphisms (SNPs) of the TCA cycle associated with colorectal cancer risk in the UK. Mutations in the IDH1 and IDH2 genes (rs121913499, rs121913500) lead to the formation of an oncometabolite, 2-hydroxyglutarate. This metabolite causes hypermethylation of histones and DNA, leading to tumor cell formation in vitro [71,72]. This oncometabolite also accumulated with the mutation c.1121G > A (p.Gly374Glu, rs1270341616) of the DLST gene [59]. Mutations in the FH gene lead to high levels of intracellular fumarate, which contributes to cancer development. Mutations in the TCA gene cause the accumulation of oncometabolites, triggering carcinogenic processes. Understanding these genetic characteristics can aid in studying the mechanisms of cancer development and in developing diagnostic methods [73].
Epigenetic modifications play a special role in understanding the development of various diseases [74].
3.1. Epigenetics of the TCA Cycle
Epigenetic modifications alter gene expression without changing their sequence [75]. They are dynamic, flexible, and reversible under external stimuli [76]. Epigenetic mechanisms include nucleosome remodeling and regulatory non-coding (small and micro) RNAs. However, the main key epigenetic modifications are DNA methylation and reversible histone modifications, e.g., acetylation, methylation, and ubiquitination [77].
DNA and histone methylation are the key epigenetic tools that regulate gene expression. DNA methylation is a covalent modification of DNA: a methyl group (-CH3) binds with cytosine groups in DNA as part of a CpG dinucleotide at C5 of the cytosine ring. DNA methylation is carried out by DNA methyltransferase (DNMT) enzymes, such as DNMT1, DNMT3A, and DNMT3B [78]. They catalyze the transfer of a methyl group from S-adenosyl-L-methionine (SAM), the ubiquitous methyl group donor, to the fifth position of the cytosine pyrimidine ring [79,80,81,82].
2-Ketoglutarate-dependent dioxygenase (2-OGDO) enzymes are DNA and histone demethylation enzymes, i.e., they remove methyl groups from DNA and histones. Such TCA cycle intermediates such as 2-oxoglutarate (ketoglutarate) are substrates for 2-OGDO enzymes. A 2-OGDO complex consists of three enzymes [81,83]:
- -2-oxoglutarate decarboxylase/2-oxoglutarate decarboxylase (E1);
- -dihydrolipoamide succinyl transferase (E2);
- -dihydrolipoamide dehydrogenase (E3).
The TCA cycle activity depends on the expression of 2-oxoglutarate dehydrogenase (2-OGDH) enzymes. The 2-OGDH occupies a central position at the interface of energy and epigenetic metabolism, since it controls the rate of conversion of 2-oxoglutarate to succinyl-CoA and thereby determines the intracellular pool of 2-oxoglutarate available for 2-OGDO [83]. It should be noted that 2-OGDH is not the only regulator of the tricarboxylic acid cycle, but is one of its key rate-limiting enzyme units along with citrate synthase and isocitrate dehydrogenase.
Low succinate dehydrogenase (SDH) leads to succinate accumulation. Succinate and fumarate are strong inhibitors of 2-OGDO enzymes. As Table 3 shows, this inhibition enhances DNA and histone methylation regulated by DNA methyltransferases (TET1–3) and histone methyltransferases (JmjC KDM2–7) [81].
According to Arnold et al. [2], metabolites formed in the TCA cycle are responsible for the regulation of chromatin. For instance, histone acetyltransferases change the accessibility of chromatin by transferring acetyl groups from acetyl-CoA to histones. Histone deacetylases remove acetyl groups from histones and produce acetate. α-Ketoglutarate-dependent dioxygenases regulate the demethylation of histones and nucleic acids, e.g., in FTO [98] and ALKBH5.
Other inhibitors include reactive oxygen species, metals (Co^2+^, Ni^2+^), and Fe^2+^ chelators. The list of cofactors includes NAD^+^, acetyl CoA, Ca^2+^, lipoic acid, thiamine, and ferrous iron (Fe^2+^). They can regulate the functional yield of 2-OGDH [81,82,83]. Therefore, the balance of TCA cycle reactions may affect DNA and histone methylation.
The work of J. Małecki [99] has demonstrated, using in vitro and in vivo approaches, that lysine methylation occurs in the CS gene at the Lys-395 residue. Methylation reduced the activity of the gene and was inhibited by the substrate oxaloacetate and adenosine. The authors hypothesized that methylation may regulate CS function in response to changes in metabolite levels. However, additional research is needed to confirm this hypothesis.
In the work of Z. Mao [100], the authors found that the activity of MDH2 is associated with an anti-aging mechanism. In vitro and in vivo studies showed that increased MDH2 expression led to increased cellular senescence. Inhibition of MDH2 expression increased histone methylation, which in turn slowed down cellular aging.
Lifestyle factors, in particular nutrition, affect the functioning of cells and the body through epigenetics. Therefore, the search for nutrients capable of epigenetic modifications is relevant [74,101].
Nutrients That Regulate the TCA Cycle
Nutraceuticals are biologically active substances included in beneficial added dietary supplements (BADS) and functional foods as part of preventive or therapeutic diets [101]. Such nutrients as magnesium, riboflavin, nicotinamide, and biotin affect the expression of mitochondrial enzyme genes through systems of transcriptional coactivators and sensors of cell energy status [102].
Magnesium ion, citrate anion, and riboflavin are part of the TCA cycle (Figure 4 and Figure 5).
Magnesium is an essential cofactor for isocitrate dehydrogenase and pyruvate dehydrogenase phosphatase in the TCA cycle [103]. Riboflavin maintains the activity of the enzyme succinate dehydrogenase, which oxidizes succinate anion into fumarate anion [103]. Magnesium is a reliable and universal regulator since it provides the structural stability of the pyruvate dehydrogenase complex, isocitrate dehydrogenase, and α-ketoglutarate dehydrogenase. In addition, it affects the transcription of energy metabolism genes by activating AMP-activated protein kinase (AMPK), which is sensitive to intramitochondrial Mg^2+^ levels [103].
Other nutrients exercise a targeted effect on the mitochondrial transcriptome. As a precursor of NAD^+^, nicotinamide activates SIRT3/5 deacetylases, thus changing the expression and post-translational modifications of such enzymes such as SDHA, IDH2, and MDH2 [36].
Riboflavin (B_2_) forms flavin mononucleotide (FMN) and flavin adenine dinucleotide (FAD). It supports succinate dehydrogenase and acyl dehydrogenases. In addition, it controls the expression of mitochondrial genes under conditions of energy substrate deficiency [104].
Omega-3 polyunsaturated fatty acids stimulate the expression of PGC-1α, which enhances the mitochondrial biogenesis and increases the flow of acetyl-CoA through the TCA cycle [105]. In their study, A. I. Borja-Magno and colleagues [106] studied the effect of omega-3 fatty acids on mitochondrial bioenergetics. A case-control study was conducted involving 20 women with a normal body mass index and 19 women with grade 2 obesity, who took 5.25 g of omega-3 fatty acids for 1 month. The study found that taking omega-3 fatty acids improved mitochondrial dysfunction in peripheral mononuclear blood cells in obese individuals. There was a decrease in triglycerides, insulin, and IL-1b, IL-6 levels.
Resveratrol and curcumin activate SIRT1 and AMPK to increase oxidative metabolism [107].
Lipoic acid (ALA) is a cofactor for mitochondrial metabolism, specifically the enzymes PDH, OGDH, branched-chain ketoacid dehydrogenase, and 2-oxoadipate dehydrogenase play a key role in carbon entry into the Krebs cycle [108]. A similar role is played by coenzyme Q (CoQ, ubiquinone), which is a cofactor of mitochondrial dehydrogenases—dihydroorotate dehydrogenase, involved in the biosynthesis of pyrimidines, and flavoprotein dehydrogenase, involved in the beta-oxidation of fatty acids [109].
Therefore, dietary supplements containing ALA, CoQ, vitamin B2, magnesium, and omega-3 are important to support healthy body function.
Table 4 summarizes the nutrients that affect the expression of TCA cycle genes.
Nutrients and biologically active substances act as cofactors for TCA cycle enzymes while affecting its regulation at the genetic level, thus shaping the adaptive metabolic response.
Nutrients use a variety of molecular mechanisms to modulate TCA cycle genes, from direct coenzyme support to activating energy sensors and transcriptional coactivators. Magnesium, riboflavin, nicotinamide, and biotin primarily support the catalytic activity and stability of TCA cycle enzymes, attracting substrates and improving the oxidative metabolism. ALA, omega-3 PUFAs, resveratrol, curcumin, arginine, leucine, and quercetin activate the AMPK–SIRT–PGC-1α signaling pathway, which boosts mitochondrial biogenesis, improves energy output, and reduces oxidative stress. Nutrients are important for mitochondrial metabolism at the level of genetic transcription and post-translational modifications.
The exact impact of nutrients on TCA cycle genetics requires more research as they are connected with metabolic pathways and susceptibility to a number of socially significant diseases. New data make it possible to develop new preventive dietary recommendations, BADS, and functional foods [121].
Despite the bioactivity of nutrients and their metabolic significance, their application is rather limited. For example, a number of biologically active substances (resveratrol, curcumin, quercetin, ursolic acid) demonstrate low solubility or lipophilicity, which means poor bioavailability. Hydrophilic compounds cannot pass through the cell membrane; lipophilic compounds do not dissolve in the gastrointestinal tract; compounds are adsorbed only after being fermented by metabolites of gastrointestinal microbiota. In this regard, food science and medicine need more effective measures to increase the bioavailability of nutrients [101,122], e.g., by derivatization, exosomal and liposomal nanocarriers, and bioavailable derivatives [123].
4. Conclusions
Mitochondrial metabolism and the citric acid cycle provide cells with energy and maintain homeostasis. Mitochondrial dysfunction is a universal pathogenetic mechanism involved in a wide range of diseases, from neurodegenerative and metabolic to inflammatory [13]. Although all the biochemical stages of the TCA cycle were described back in 1937, we still have no clear understanding of how to control it. However, some nutrients (niacin, α-lipoic acid, succinic acid, resveratrol, curcumin, arginine, leucine, quercetin, ursolic acid, alternol) can modulate gene expression, thereby improving the efficiency of the TCA cycle, which makes this research direction extremely relevant.
This review left out publications on mitochondrial diseases caused by mtDNA defects and attempts to regulate the TCA cycle with hormones and signal transduction systems.
5. Strategies/Methods
This review covered 45 years of articles published in Russian and English and indexed in the PubMed (US National Library of Medicine), Scopus (Elsevier), and the Russian Scientific Electronic Library (eLIBRARY.RU). The final pool of the most relevant publications consisted of 123 articles.
The list of keywords included citric acid cycle, TCA cycle, Krebs cycle, mitochondrial dysfunction, TCA epigenetics, TCA genes, and mitochondrial health diet.
We focused on the papers describing the results of clinical and preclinical studies (regardless of the model objects), as well as review articles. We excluded the articles describing the results obtained in silico, conference proceedings, and monographs.
All the publications returned by the search were screened for relevance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1El-Taravi Y.A. Erikenova D.K. Odarchenko A.S. Magerramova S.T. Shestakova M.V. Nobel prize winners in metabolism history and diabetology Diabetes Mellit.202326375381(In Russia)10.14341/DM 13032 · doi ↗
- 2Arnold P.K. Finley L.W.S. Regulation and function of the mammalian tricarboxylic acid cycle J. Biol. Chem.202329910283810.1016/j.jbc.2022.10283836581208 PMC 9871338 · doi ↗ · pubmed ↗
- 3Krebs H.A. Johnson W.A. Metabolism of ketonic acids in animal tissues Biochem. J.19373164566010.1042/bj 031064516746382 PMC 1266984 · doi ↗ · pubmed ↗
- 4Krebs H.A. Johnson W.A. The role of citric acid in intermediate metabolism in animal tissues FEBS Lett.198011738339010.1016/0014-5793(80)80564-36998725 · doi ↗ · pubmed ↗
- 5Chen T.-H. Koh K.-Y. Lin K.M.-C. Chou C.-K. Mitochondrial Dysfunction as an Underlying Cause of Skeletal Muscle Disorders Int. J. Mol. Sci.2022231292610.3390/ijms 23211292636361713 PMC 9653750 · doi ↗ · pubmed ↗
- 6Arneth B.M. The Impact of Krebs Cycle Intermediates on the Endocrine System and Immune System: A Comparison Endocrines 2023417919310.3390/endocrines 4010016 · doi ↗
- 7Zong Y. Li H. Liao P. Chen L. Pan Y. Zheng Y. Zhang C. Liu D. Zheng M. Gao J. Mitochondrial dysfunction: Mechanisms and advances in therapy Signal Transduct. Target Ther.2024912410.1038/s 41392-024-01839-838744846 PMC 11094169 · doi ↗ · pubmed ↗
- 8Du J. Zhu M. Bao H. Li B. Dong Y. Xiao C. Zhang G.Y. Henter I. Rudorfer M. Vitiello B. The Role of Nutrients in Protecting Mitochondrial Function and Neurotransmitter Signaling: Implications for the Treatment of Depression, PTSD, and Suicidal Behaviors Crit. Rev. Food Sci. Nutr.2016562560257810.1080/10408398.2013.87696025365455 PMC 4417658 · doi ↗ · pubmed ↗