Probiotic-Driven Metabolic Transformation in Dark Tea: Enhanced Bioactives and Gut Microbiota Modulation Underlie Its Alleviation Effect Against Constipation
Yufeng Xie, Xiaoyan Song, Zihao Zhang, Zitong Zhou, Hongyu Liu, Zhongfang Wang, Longgang Jia

TL;DR
Fermented dark tea with a probiotic helps relieve constipation better than regular dark tea by changing gut bacteria and boosting helpful compounds.
Contribution
The study reveals that probiotic fermentation enhances dark tea's anti-constipation effects through gut microbiota modulation and bioactive compound transformation.
Findings
Fermented dark tea (FDT) improved constipation symptoms more effectively than non-fermented dark tea (DT) in mice.
FDT increased beneficial gut bacteria like Lactobacillus and reduced harmful Prevotellaceae_UCG-001.
FDT's fermentation enriched bioactive compounds like cyclic dipeptides and hydroxylated linoleic acid derivatives.
Abstract
Constipation is a widespread gastrointestinal condition with complex causes and limited therapeutic efficacy of current treatments. Probiotics and natural-function foods like dark tea (DT) have emerged as promising alternatives. We investigated the protective effects of dark tea (DT) and Lactiplantibacillus paracasei K34-fermented dark tea (FDT) against constipation induced by loperamide (LOP) in mice. FDT exhibited a superior effect over DT in alleviating constipation, as evidenced by increased fecal water content, reduced defecation time, and accelerated small intestinal transit. FDT also restored serum gastrointestinal neurotransmitters and repaired intestinal barrier damage more effectively. FDT intervention significantly increased the richness and altered the composition of gut microbiota, notably elevating Lactobacillus, Ligilactobacillus, and Actinobacteria, while reducing…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
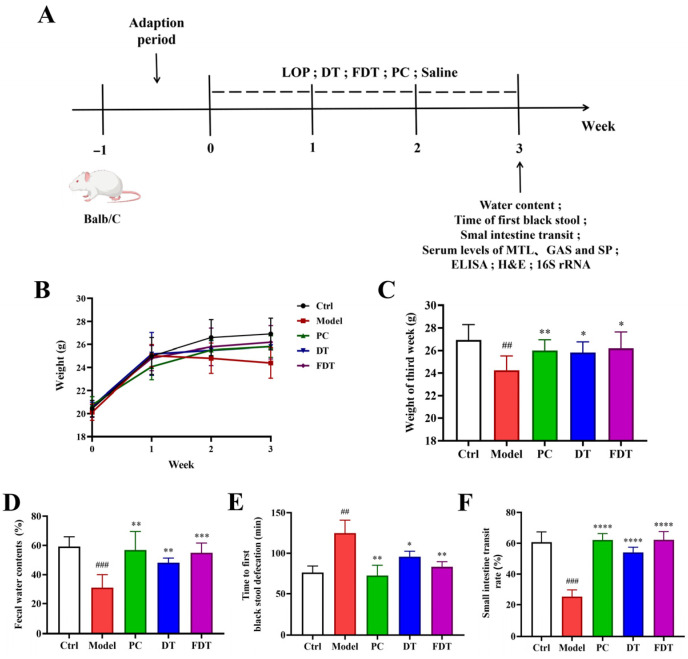 Figure 1
Figure 1- —Tianjin Science and Technology Plan Projects
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTea Polyphenols and Effects · Gut microbiota and health · Polysaccharides Composition and Applications
1. Introduction
As a common gastrointestinal disorder, constipation is clinically defined by infrequent bowel movements and hard stools, and has a global prevalence of approximately 15% [1]. Long-term constipation may lead to neurasthenia, toxemia, metabolic disorder and other pathological symptoms [2]. Substantial resources are devoted to constipation treatment globally each year; however, over half of patients are refractory to treatment [3], leading to a profoundly negative impact on their quality of life. A range of therapeutic options are currently available for constipation, encompassing lifestyle and dietary modifications, biofeedback therapy, and pharmacological interventions including laxatives, prokinetic agents, and prosecretory drugs [4]. Given their limited efficacy and potential for adverse effects, these conventional therapies are often insufficient, emphasizing the critical need for safer alternatives in constipation care.
Constipation is a multifactorial disorder whose pathogenesis is not fully understood. It is now widely accepted that constipation arises from a combination of factors, including dysregulation of the enteric and central nervous systems (ENS and CNS), gut microbial dysbiosis, and abnormal gastrointestinal motility [5]. A growing body of evidence has established a robust link between gut microbiota dysbiosis and constipation, positioning microbiota-targeted interventions as a promising therapeutic strategy [6,7]. The regulatory role of gut microbes on the ENS and gastrointestinal peristalsis is mediated by neurotransmitter production, as evidenced by altered levels of motilin (MTL), peptide YY (PYY), vasoactive intestinal peptide (VIP), and 5-hydroxytryptamine (5-HT) found in the serum and colons of constipated mice [8,9,10]. Leveraging the intricate interactions between probiotics, gut microbiota, and hosts, therapeutic manipulation of gut microbiota with probiotics is a cornerstone of biomedical strategy for constipation [11,12]. Accumulating evidence has substantiated the beneficial role of prominent genera, including lactobacillus and bifidobacterium, in relieving constipation symptoms [7]. For instance, L. casei Shirota [13], B. longum BB536 [14], L. reuteri DSM19738 [15], and B. lactis HN019 [16] have been demonstrated in clinical studies to alleviate constipation symptoms by modulating the intestinal microenvironment and ENS/CNS functions. Lacticaseibacillus paracasei is one of the most common types of lactobacilli found in the intestine, fermented foods, and dairy products. Studies have highlighted the health-promoting properties of L. paracasei strains isolated from various sources, underscoring their potential as functional foods or adjuvant therapies [17]. Studies have demonstrated that L. paracasei CNCM I-5220 contributes to overall gastrointestinal health by enhancing intestinal barrier integrity and reducing gut permeability [18]. Furthermore, L. paracasei exhibits significant immunomodulatory effects by promoting the production of the anti-inflammatory cytokine IL-10 while suppressing key pro-inflammatory cytokines, including TNF-α and IL-6 [19]. The potential of L. paracasei in constipation management is supported by human and animal studies showing that the strain and its metabolites can alleviate symptoms, further reinforcing its promising role in supporting immune health and managing inflammation [20,21,22,23].
Dark tea (DT), a distinctive variety of post-fermented tea, has had a longstanding role in traditional practices for addressing gastrointestinal discomfort, such as from dyspepsia, bloating, and constipation [24,25]. The fermentation process of DT fosters rich microbial diversity and promotes the transformation of tea-derived phytochemicals into bioactive metabolites [26]. Studies have demonstrated that the addition of probiotic microorganisms to food matrices can significantly influence both the viability of probiotics and the profile of volatile aromatic compounds, thereby enhancing the functional properties of fermented products [27]. Recent research has elucidated the dynamic changes in chemical constituents that occur during the microbial fermentation of dark tea. Consequently, numerous novel components contributing to tea quality have been identified, encompassing catechin derivatives, flavonols, flavones, and their glycosides, as well as phenolic acids, alkaloids, and terpenoids [28,29,30]. Importantly, the ability of DT to accelerate gastrointestinal transit makes it a promising candidate for relieving constipation and improving gastrointestinal motility. Here, using an LOP-induced constipation mouse model, this study investigated the anti-constipation effects of DT and FDT, employing 16S rRNA gene sequencing to analyze gut microbiota modulation, and untargeted metabolomics to identify differential bioactive components and pathways underlying their functional differences.
2. Materials and Methods
2.1. Probiotic Strain, Materials and Reagents
L. paracasei K34 was isolated from traditional Northeastern Chinese sour porridge and stocked in our laboratory, and it has been further identified and preserved by the China General Microbiological Culture Collection Center (CGMCC) under the deposit number CGMCC No. 36233.
Loperamide (LOP) was obtained from Xi’an Janssen Pharmaceutical Co., Ltd. (Xi’an, China). DT was purchased from the Hunan Xiangcha High-Tech Co., Ltd. (Yiyang, China). Combined Bacillus Subtilis and Enterococcus Faecium Granules with Multivitamins (Mommy Love) were sourced from the Beijing Hanmi Pharmaceutical Co., Ltd. (Beijing, China). Neutral formalin fixative solution (10%) was purchased from Tianjin Biyou Biotechnology Co., Ltd. (Tianjin, China). Trizol reagent, the ABScript III RT Master Mix, and the 2× Universal SYBR Green Fast qPCR Mix kit were purchased from Wuhan AB SciTech Biotechnology Co., Ltd. (Wuhan, China). Enzyme-linked immunosorbent assay (ELISA) kits of motilin (MTL), gastrin (GAS), P substance (SP), tumor necrosis factor-α (TNF-α), interleukin-1β (IL-β) and interleukin-10 (IL-10) were obtained from Nanjing Caobenyuan Biotechnology Co., Ltd. (Nanjing, China). All remaining chemicals, of the highest purity available, were sourced locally unless indicated otherwise.
For the preparation of FDT, dark tea powder was sterilized and inoculated with a 2% (v/w) suspension of L. paracasei K34 (approximately 10^8^ CFU/mL), followed by incubation at 37 °C for 48 h under static conditions. After fermentation, the samples were lyophilized and stored at −20 °C until further use.
2.2. Animals and Experimental Design
Male BALB/c mice (six-week-old, SPF grade) were purchased from Beijing Sibeifu Experimental Animal Co., Ltd. (Beijing, China). All animal experiments were conducted following protocols sanctioned by the Animal Research Ethics Committee of Tianjin University of Science and Technology, and in strict compliance with the university’s Guidelines for the Care and Use of Laboratory Animals. The mice were housed in a standard environment (23 ± 3 °C, 51 ± 13% humidity) under a 12 h light–dark cycle, with food and water provided ad libitum. After one week of acclimatization, the animals were randomly divided into five groups (n = 8 per group): Ctrl, Model, PC (Mommy Love), DT (dark tea), and FDT (dark tea fermented with L. paracasei K34), as depicted in Figure 1A. Constipation was induced in all groups except the Ctrl group by daily oral administration of LOP (10 mg/kg body weight). The Ctrl group received saline only. In the intervention stage, starting on the 17th day, the treatment group was administered DT (0.01 g/day) or FDT (0.01 g/day, containing approximately 10^9^ CFU K34) by gavage. On the 28th day, fecal water content, first black stool, and small intestine transit rate analyses were performed. After the animal experiment, the mice were fasted overnight and sacrificed, then the serum, small intestine tissues, and feces were collected for histological and microbiome analysis. Quantification of serum biomarkers, including MTL, GAS, SP, TNF-α, IL-1β, and IL-10, was performed via ELISA following the protocols provided by the manufacturer [31,32]. In parallel, body weight measurements were taken at a fixed time (09:00 a.m.) every week for the duration of the experiment.
2.3. Defecation Test of the Mice
Weekly body weight measurements were taken throughout the entirety of the study. At the endpoint, a final stool collection was performed for further analysis. After the treatment period, mice were placed in individual cages for 1 h. Fresh fecal samples were collected, immediately weighed to obtain the wet weight, and subsequently dried in an oven at a constant temperature for 24 h to determine the dry weight. The fecal water content was then calculated according to the following established formula [23,33,34]:
Intestinal transit capability was assessed at the experimental endpoint following a 12 h fasting period (water provided). Four mice per group were orally administered 0.2 mL ink to record the time to first black stool, while the remaining four were euthanized 30 min after ink administration to measure the distance of ink propulsion through the small intestine. These methodologies are standard for evaluating constipation-related parameters in rodent models [23,33,34]. The small intestinal transit rate was subsequently calculated as per the formula below:
2.4. Hematoxylin–Eosin (H&E) Staining
Following fixation in 4% neutral buffered paraformaldehyde, mouse small intestine tissues were rinsed overnight under running water and subsequently dehydrated through a graded ethanol series (70%, 80%, and 90% for 30 min each; 95% and 100% for 20 min each). The samples were then embedded in paraffin and sectioned at 5 μm thickness. The resulting sections were flattened and baked at 60 °C for 60 min in a drying oven, and then treated with xylene and rehydrated through a descending alcohol gradient. H&E staining was performed according to standard histological procedures [32] to assess morphological changes in the intestinal tissue. After staining, the sections were mounted with neutral gum. Once the mounting medium had fully solidified, the sections were scanned using a digital slide scanner (BX53, Olympus Corporation, Tokyo, Japan).
2.5. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
Using Trizol reagent, we extracted total RNA from small intestine tissue and then synthesized cDNA with the ABScript Neo RT Master Mix(ABclonal Biotechnology Co., Ltd., Wuhan, China), following the manufacturer’s instructions. We subsequently performed RT-qPCR to quantify the expression of target genes (TNF-α, IL-1β, and IL-10), employing β-actin as an internal control for normalization. Relative mRNA levels were calculated using the 2^−ΔΔCt^ method as established by Livak and Schmittgen [35]. Primer sequences are listed in Table S1.
2.6. Analysis of the Gut Microbiota
Fecal samples from 4 randomly selected mice per group were subjected to microbiota analysis. This sample size is consistent with previous microbiome studies [23,36,37,38,39,40] and was determined to be sufficient for detecting significant differences in microbial composition based on preliminary power analysis. Fecal microbial DNA was extracted, quantified, and subjected to 16S rDNA sequencing following a previously established protocol [32]. Sequencing data were analyzed on the BMK Cloud platform. For diversity analysis, α-diversity indices (Shannon, Simpson, Chao1, and Ace) were calculated, and β-diversity was examined via principal coordinates analysis (PCoA) based on OTU-level profiles. Taxonomic profiling at the phylum and genus levels was achieved via barplot analysis, while linear discriminant analysis effect size (LEfSe) was used to identify differentially abundant taxa between groups.
2.7. Untargeted Metabolomics Analysis of DT and FDT by Liquid Chromatography–Mass Spectrometry (LC–MS)
LC–MS-based untargeted metabolomics was carried out on DT and FDT samples following previously described methods [41,42,43]. Sample preparation entailed mixing 10 mg of powder (DT or lyophilized FDT) with 1 mL of pre-chilled methanol–acetonitrile (1:1, v/v). After centrifugation (12,000 rpm, 10 min), the supernatant was collected and evaporated to dryness. The extract was redissolved in 50% methanol (with internal standards) and centrifuged again under the same conditions. The resulting supernatant was filtered and transferred to a sample vial. A quality control sample was prepared by pooling 10–20 μL of each individual sample. Thereafter, each sample was injected into an ACQUITY UPLC HSS T3 column (100 Å, 1.8 μm, 2.1 mm × 100 mm; Waters, Milford, MA, USA) maintained at 40 °C. The mobile phase A was water with 0.1% formic acid, phase B was acetonitrile with 0.1% formic acid, and flow rate was 0.4 mL/min. The elution gradient was set as follows: 0–1 min, 5% phase B; 4.7–6 min, 95% B; 6.1–8.5 min, 5% B. Mass spectrometric analysis of all samples was performed on an Orbitrap Exploris 120 instrument (Thermo Fisher Scientific, Waltham, MA, USA) under the control of Xcalibur (v4.7). Data were acquired in data-dependent acquisition (DDA) mode using both positive and negative electrospray ionization (HESI), with key parameters including: spray voltage (±3.5/−3.0 kV), sheath gas (40 arb), auxiliary gas (10 arb), capillary temperature (320 °C), auxiliary gas temperature (300 °C), and scan range (70–1000 m/z). Acquired raw data were imported into MS-DIAL (v4.9.221218) for subsequent processing, which encompassed peak detection, alignment, filtering, and metabolite identification against the PSNGM database. To ensure data quality, features absent in >50% of QC samples were discarded, and missing values were imputed using the software’s gap-filling algorithm prior to normalization.
2.8. Statistical Analysis
Results are expressed as means ± SEM, with sample sizes ranging from 3 to 8 per group. GraphPad Prism 9.5 (GraphPad Software Inc., San Diego, CA, USA) was employed for all statistical evaluations. Differences between groups were assessed using either one-way ANOVA or Student’s t-test.
3. Results and Discussion
3.1. Intervention of DT and FDT Alleviated the Constipation Symptoms of Mice Induced by LOP
Constipation was induced in mice using LOP, an antidiarrheal drug that activates intestinal μ-opioid receptors, thereby delaying gastrointestinal transit, inhibiting smooth muscle contractions, and slowing peristalsis [44]. To evaluate the therapeutic effects of DT and FDT, a constipation mouse model was established using LOP, as illustrated in Figure 1A. Following one week of LOP administration, a progressive decline in body weight was observed in the model group compared to the Ctrl group, while the intervention of PC, DT and FDT changed the mice body weight decreasing trends (Figure 1B). During the last week of animal experiments, the body weight of LOP group mice was significantly lower than that of the Ctrl mice, and the body weights of all the three treatment group mice were significantly increased compared to the LOP group (Figure 1C, p < 0.05). Weight loss in a mouse constipation model is usually the result of multiple factors acting together, mainly including a reduction in the weight of gastrointestinal contents, decreased appetite, dehydration, and the side effects of the model-inducing agents themselves [34,45].
To confirm successful constipation induction, fecal water content was measured after 7 days of loperamide administration. As shown in Figure S1, mice in the model group exhibited significantly lower fecal water content compared to the Ctrl group (p < 0.0001), confirming the establishment of the constipation model. The measurements of fecal water content, first black stool time and small intestine transit rate of the mice across different groups were analyzed after the DT or FDT interventions. A significant decrease in fecal water content was observed in the LOP group relative to the Ctrl group (p < 0.001, Figure 1D). Similar to the PC group, oral administration of DT and FDT significantly elevated the fecal water content in the LOP-induced mice (p < 0.01). The prolongation of first black stool time induced by LOP was significantly reversed by treatment with either DT or FDT compared to the untreated model group (Figure 1E, p < 0.05). Furthermore, the trends in small intestine transmit rate were similar to that of fecal water content data, which showed that DT and FDT could remarkably increase down-regulated small intestine transit rate in LOP-induced mice (Figure 1F, p < 0.0001). These results demonstrated that DT or FDT supplementation significantly improved fecal humidity and enhanced intestinal motility in LOP-induced constipation mice. Of note, from the changes in the three gastrointestinal indicators, the effects of FDT intervention in the LOP-induced mice were better than for DT.
Supplementation of DT or FDT alleviated LOP-induced constipation symptoms. (A) Animal experimental outline of DT and FDT administration in LOP-induced constipation mice. (B) Body weight changes in mice during the animal experiments. (C) The body weight of mice in the last week. (D) Fecal water contents. (E) Time to first black stool defecation. (F) Small intestine transit rate. ## p < 0.01 and ### p < 0.001 compared to the Ctrl group. * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001 compared to the Model group.
3.2. Effects of DT and FDT on the Levels of Serum Neurotransmitters in the LOP-Induced Constipation Mice
It has been reported that excitatory gastrointestinal regulatory neurotransmitters, including MTL, GAS and SP, are positively correlated with the symptoms of constipation. This is because MTL stimulates gallbladder contraction, enhances insulin secretion, and increases gastrointestinal motility; GAS facilitates amino acid absorption and induces gastric distension; SP promotes smooth muscle relaxation and, concurrently, it augments the sensitivity of intestinal smooth muscle to acetylcholine, thereby stimulating smooth muscle contraction and ultimately enhancing intestinal peristalsis [46,47,48,49]. Therefore, as can be seen in Figure 2, the serum MTL, GAS and SP levels of the mice in the LOP group were significantly decreased compared to the Ctrl group (p < 0.001). Treatment with DT or FDT significantly increased serum MTL, GAS, and SP concentrations relative to the Model group, mirroring the effects of the positive control (Figure 2, p < 0.05). By restoring these suppressed excitatory neurotransmitters to levels approaching those of healthy controls, both DT and FDT promoted normal intestinal motility and ameliorated constipation via hormonal pathways. Our results corroborate previous findings that highlight neurotransmitter regulation as a key mechanism in constipation therapy [33,34,46].
3.3. Intervention of DT and FDT Alleviated the Intestinal Barrier Damage in the LOP-Induced Constipation Mice
Intestinal barrier dysfunction is a common pathological feature of constipation, underscoring that restoration of barrier integrity constitutes an effective approach to its alleviation [31,36]. The small intestinal epithelial cells in the Ctrl group exhibited an intact structure without atrophy or necrosis, and were neatly arranged with tight intercellular spaces, normal morphology, and a clear architecture (Figure 3A). However, the LOP-induced constipation mice group exhibited obvious atrophy of the small intestinal epithelial cells, which had shrunken appearance, reduced size, and disorganized, sparse arrangement (Figure 3A). The intestinal villi in the LOP group were significantly shortened and mostly lost (Figure 3A). The FDT group, in particular, exhibited marked amelioration of intestinal barrier damage following oral administration, an effect that paralleled the PC group and surpassed that observed in the DT mice, which exhibited intact, neatly arranged epithelial cells and a notable reduction in intercellular space (Figure 3A).
The intestinal barrier serves as both a physical and functional blockade, preventing luminal contents from entering systemic circulation [50]. Moreover, the integrity of this barrier is crucial not only for efficient nutrient absorption, but also for defending against pathogenic bacteria, maintaining intestinal mucosal immune function, and promoting recovery from inflammatory injury [51]. So, the serum contents of cytokines including TNF-α, IL-1β and IL-10 were analyzed to evaluate the effects of DT and FDT on intestinal barrier damage in the LOP-induced constipation mice. The LOP group mice showed significantly higher levels of pro-inflammatory cytokines TNF-α and IL-1β, and lower levels of anti-inflammatory cytokine IL-10 than the Ctrl group (Figure 3B–D, p < 0.01). After intervention with DT and FDT, the levels of TNF-α and IL-1β were significantly decreased, and IL-10 levels were increased compared to the LOP-induced constipation mice group (Figure 3B–D, p < 0.05). Relative levels of colon cytokines among the different groups showed similar results (Figure S2). Therefore, DT and FDT mitigated LOP-induced constipation by fortifying the intestinal barrier, reducing permeability, and suppressing inflammation. It should be noted that a comprehensive investigation of the enteric serotonergic system, including colonic tryptophan hydroxylase 1 (TPH1) and serotonin transporter (SERT) expression, is a critical next step to validate the mechanistic link between the gut microbiota, the enteric nervous system, and the systemic neurotransmitter changes observed with FDT treatment [52,53].
3.4. Intervention of DT and FDT Regulated the Diversity and Composition of Gut Microbiota in LOP-Induced Constipation Mice
Analysis of gut microbiota via 16S rRNA gene sequencing revealed distinct compositional shifts associated with DT and FDT treatment in LOP-induced constipated mice. While 245 OTUs were common to all groups (Figure S3A) [54], OTU richness, significantly reduced in the Model group versus the Ctrl group, was partially restored following PC, DT, or FDT treatment (Figure S3A). Among the three treatments, the FDT group had the highest increase in OTUs (Figure S3A). As shown in Table 1, LOP-induced constipation (Model group) resulted in a significant decrease in both the ACE and Chao1 indices relative to the Ctrl group, indicating reduced gut microbial richness (p < 0.001). These indices, along with the Shannon and Simpson indices (which reflect species diversity), are standard measures for assessing the α-diversity of microbiota [55]. After treatment with PC, DT or FDT, the ACE and Chao1 indexes were obviously elevated, especially for the FDT group (p < 0.001 compared to the Model group), whereas the Simpson and Shannon diversity indices remained comparable across all experimental groups (Table 1). The α-diversity results indicated that oral administration of LOP affected the species richness of gut microbiota, while intervention with DT or FDT could reverse the reduction. PCoA of gut microbiota β-diversity revealed that although the scatter regions of each group were closely distributed, they did not completely overlap, suggesting discernible differences in the composition and structure of gut microbiota among the groups. Compared with the Ctrl group, the Model group showed a significant shift in microbial composition, indicating that LOP modeling induced distinct gut microbiota profiles in constipated mice relative to normal mice (Figure S3B). Post-intervention analysis revealed a clear divergence in gut microbiota structure between the treatment groups (DT and FDT) and the Model group (Figure S3B). Of particular note, the microbial community of FDT-treated mice clustered more closely with the Ctrl group, exhibiting partial overlap and suggesting that FDT intervention effectively mitigated LOP-induced dysbiosis and partially restored the gut microbial community structure toward a normal state.
To further characterize the disparities in gut microbial composition across different groups, we analyzed the abundance of specific gut microbiota at different taxonomic levels. At the phylum level, Firmicutes and Bacteroidetes constituted the dominant microbiota across all groups, and a relatively stable Firmicutes/Bacteroidetes ratio is known to be vital for the health of the host [56]. Compared with the Ctrl group, Model mice exhibited a reduced abundance of Firmicutes, leading to a significantly lower Firmicutes/Bacteroidetes ratio (Figure 4A,B, p < 0.05). A comparable shift in this ratio has been documented in other disease states, paralleling findings in conditions like colitis and colon cancer [57]. After intervention with PC, DT and FDT, the relative abundances of Firmicutes and Bacteroidetes were altered, leading to a significant elevation in Firmicutes/Bacteroidetes ratio compared to the Model group (Figure 4A,B, p < 0.05). Alterations were also observed in the relative abundance of Actinobacteria across groups. Notably, a marked decrease was evident in the Model group compared to the Ctrl group (Figure 4A,C, p < 0.05). In contrast, intervention with FDT significantly restored the Actinobacteria level compared to the Model group (Figure 4A,C, p < 0.01). Compared to the Ctrl group, the Model group exhibited markedly reduced abundances of Lactobacillus and Ligilactobacillus at the genus level (Figure 4D and Figure S4A,B). However, following FDT treatment, these genera rebounded significantly, with their levels elevated above those observed in the Model group (Figure 4D and Figure S4A,B). The relative abundances of Lachnospiraceae_NK4A136 and Alloprevotella were also decreased in the Model group compared to the Ctrl group (Figure 4D and Figure S4C,D, p < 0.05). DT intervention markedly increased the abundance of Lachnospiraceae_NK4A136 (Figure 4D and Figure S4C, p < 0.001), while PC administration specifically and substantially elevated Alloprevotella levels (Figure 4D and Figure S4D, p < 0.001). Intervention with PC, DT or FDT mitigated the increase in Prevotellaceae_UCG_001 abundance triggered by LOP (Figure 4D and Figure S4E, p < 0.01). In short, DT and FDT elicited obvious shifts in gut microbiota structure and composition, especially for the relative abundances of Lactobacillus and Ligilactobacillus. Through the synthesis of key digestive enzymes, amino acids, and vitamins, Lactobacillus plays a crucial role in facilitating efficient nutrient digestion, absorption, and utilization within the host. Beyond its role in digestion, this genus contributes to immunomodulatory properties, including fortifying the intestinal mucosal barrier, suppressing harmful bacterial growth, and bolstering resistance to pathogens [58]. Moreover, previous studies have demonstrated that oral administration of Ligilactobacillus salivarius Li01 promotes intestinal barrier repair and restores gut microbial balance [59]. Consistent with these attributes, prior research has demonstrated that supplementation with Ligilactobacillus salivarius Li01 promotes the restoration of intestinal barrier integrity and rebalances the gut microbiota [60].
To identify differentially abundant microbial taxa, linear discriminant analysis effect size (LEfSe) analysis was conducted. As depicted in Figure 5A, the cladogram displays the taxonomic hierarchy from phylum to species via radiating circles (Figure 5A). Within this structure, each node corresponds to a specific taxon, with node size proportional to relative abundance (Figure 5A). Taxa showing no significant inter-group variation are denoted by yellow nodes, while the colored regions, including blue, red, purple, green, and orange, denote taxa that showed significant differences in the Ctrl, Model, PC, DT, and FDT groups (Figure 5A). In cross-group comparisons, the LDA score histogram revealed differentially abundant taxa across multiple taxonomic levels (Figure 5B, LDA score > 3, p < 0.05). The dominant microbial genus in the Ctrl group was Blautia, Aerococcus, and Barnesiella, while in the Model group, characteristic strains were Phascolarctobacterium, Acidaminococcaceae and Rikenella (Figure 5B). Signature taxa for the DT group included Lachnospiraceae_NK4A136, Negativibacillus, and Turicibacter, whereas the FDT group was distinguished by Atopostipes, Rodentibacter, and Actinobacteria (Figure 5B). By enhancing diversity and enriching beneficial microbes, FDT intervention effectively rebalanced the gut microbiota in constipated mice. This remodeling is particularly noteworthy, as Actinobacteria is a key phylum implicated in the preservation of gut homeostasis [37], and their altered abundance is associated with conditions including inflammatory bowel disease [38], ankylosing spondylitis [39], type 2 diabetes [40], and irritable bowel syndrome [61]. We acknowledge that the sample size for gut microbiota analysis (n = 4 per group) was smaller than that for physiological parameter assessments (n = 8 per group). While this sample size was adequate to detect significant differences in microbial composition in our study, future investigations with larger cohorts would further validate these findings and provide greater statistical power for detecting subtle changes in low-abundance taxa.
3.5. Analysis of Correlations Between Gut Microbiota and Physiological Parameters in Constipated Mice Using Spearman’s Correlation Methods
The correlation heatmap reveals significant associations between specific gut microbial taxa and key physiological, biochemical, and immunological parameters in constipated mice. Notably, the genera Lactobacillus and Ligilactobacillus exhibited strong positive correlations with intestinal motility markers, including fecal water content, small intestine transit rate and MTL levels. These taxa were also positively associated with IL-10, suggesting a potential role in enhancing gut motility and modulating immune responses (Figure 6). The genus Ligilactobacillus also showed positive correlations with GAS and SP, indicating a possible link between this microbial group and neuro-immune signaling in the gut (Figure 6). The phylum Actinobacteria and the genera Lactobacillus, Ligilactobacillus and Alloprevotella were negatively correlated with TNF-α and IL-1β. These inverse relationships suggested that higher abundances of these taxa might contribute to reduced intestinal inflammation (Figure 6). The genus Prevotellaceae_UCG-001 was positively associated with TNF-α, and negatively associated with IL-10, indicating that this genus might increase inflammatory response (Figure 6). The genus Alloprevotella demonstrated a negative correlation with time to first black stool defecation, implying a potential role in promoting faster colonic transit and alleviating constipation (Figure 6). Collectively, these data imply that the compositional remodeling of the gut microbiota is functionally linked to the regulation of host physiology, offering insights into potential therapeutic targets. The enrichment of Actinobacteria, Lactobacillus, and Ligilactobacillus appeared to support gut motility, stool hydration, and anti-inflammatory activity, whereas Actinobacteria and Prevotellaceae_UCG-001 might contribute to inflammation reduction and accelerated transit. These findings collectively suggested that modulating specific bacterial taxa could serve as a therapeutic strategy for improving constipation symptoms and associated inflammatory dysregulation.
It is important to note that the LOP-induced constipation model employed in this study is primarily a motility-based model, as LOP acts directly on intestinal μ-opioid receptors to inhibit peristalsis [44]. While this model effectively mimics the delayed intestinal transit characteristics of slow-transit constipation, it does not fully replicate the complex etiology of constipation in humans, which often involves interactions among dietary factors, stress, neurological dysfunction, and gut microbiota dysbiosis [5]. The gut microbiota alterations observed in LOP-treated mice (reduced microbial richness, decreased Firmicutes/Bacteroidetes ratio, and reduced abundances of beneficial strains such as Lactobacillus and Actinobacteria) likely represent secondary effects of slowed intestinal transit rather than primary drivers of constipation. This distinction is important because it suggests that microbiota modulation by FDT, particularly the enrichment of Lactobacillus, Ligilactobacillus, and Actinobacteria, may be a consequence of improved intestinal motility creating a more favorable environment for beneficial bacteria, in addition to any direct prebiotic or probiotic effects. The correlation analysis (Figure 6) supports this interpretation, showing positive associations between these beneficial taxa and motility markers (fecal water content, small intestinal transit rate, and MTL). Nevertheless, the remarkable increase in these beneficial taxa following FDT intervention, combined with the enrichment of anti-inflammatory bioactive metabolites (cyclo-(Gly-Pro) and hydroxylated linoleic acid derivatives), suggests a multi-targeted mechanism that addresses both motility impairment and its downstream consequences on the gut ecosystem. While the LOP model provides valuable insights into FDT’s effects on intestinal motility, future studies should evaluate its efficacy in complementary constipation models that better reflect the multifactorial nature of human constipation. Specifically, diet-induced constipation models (e.g., low-fiber or high-fat diets), stress-induced constipation models, and models of dysbiosis-associated constipation (e.g., antibiotic-induced) would help determine whether FDT’s benefits extend beyond motility enhancement to address other etiological factors. Such investigations would also clarify the relative contributions of direct motility stimulation versus microbiota modulation in FDT’s anti-constipation effects.
While our study demonstrates that FDT (containing both live L. paracasei K34 and its fermented metabolites) effectively alleviates constipation and modulates gut microbiota, the current experimental design does not allow us to distinguish the individual contributions of live bacterial cells versus fermentation-derived bioactive compounds. The marked increase in beneficial genera such as Lactobacillus and Ligilactobacillus observed in the FDT-treated group could potentially arise from several mechanisms, for example direct colonization and metabolic activity of the administered K34 strain, prebiotic-like effects of the fermented tea metabolites that selectively promote the growth of indigenous beneficial bacteria, or synergistic interactions between the live bacteria and the transformed phytochemicals. Furthermore, the specific role of live K34 bacteria in driving these microbial shifts remains to be elucidated, as our current analysis reflects the net effect of the combined intervention. Future studies incorporating comparative experimental groups, such as heat-killed FDT (postbiotic), K34 culture alone, and unfermented DT co-administered with K34, would be valuable to elucidate the relative importance of live bacterial colonization versus metabolite-mediated effects. Such investigations would not only clarify whether active colonization of K34 is necessary for marked changes in gut microbiota composition but also determine the extent to which postbiotic metabolites alone can reproduce the therapeutic benefits. These mechanistic dissections represent important next steps in understanding the full therapeutic potential of FDT and could inform the development of more targeted probiotic or postbiotic interventions for constipation.
3.6. Differential Bioactive Compounds Comparison Between DT and FDT Using Untargeted Metabolomics
According to the findings from the aforementioned animal experiments and the assessment of physiological and biochemical results, the FDT group demonstrated a slightly superior improvement effect compared to DT. To investigate the underlying mechanisms, we employed untargeted metabolomics to analyze changes in specific bioactive components of DT before and after probiotic fermentation. Unsupervised principal component analysis (PCA) revealed distinct clustering of metabolite profiles between the DT and FDT groups in both positive and negative ionization modes (Figure S5), indicating substantial metabolic alterations induced by fermentation. To further characterize these changes, differentially abundant metabolites were identified based on fold change and P-value thresholds and visualized using volcano plots. Compared to the DT group, FDT exhibited 733 upregulated and 426 downregulated metabolites in positive ion mode, and 504 upregulated alongside 396 downregulated metabolites in negative ion mode (Figure 7A,B). Notably, clustering analysis of differentially abundant metabolites with key bioactive properties in the FDT group compared to the DT group revealed substantial alterations in specific compounds. In positive ion mode, relative levels of cyclo-(Gly-Pro) and α-eleostearic acid were significantly elevated by 132.4-fold and 26.3-fold, respectively (Figure 7C,D, p < 0.0001). Correspondingly, the relative abundances of N-(17-hydroxy-9,12-octadecadienoyl)-glutamine, nigrolineaxanthone M, and Trp-Asp increased by 123.8-fold, 48.4-fold, and 15.8-fold, respectively, in the negative ion mode (Figure 7E,G, p < 0.0001). The key differential metabolites identified above are all recognized for their potent antioxidant and anti-inflammatory activities. A notable example is cyclo-(Gly-Pro), a cyclic dipeptide with documented therapeutic potential against pain and inflammatory conditions. Its antinociceptive and anti-inflammatory effects are mediated primarily through interaction with the opioid system, leading to reduced hypernociception and paw edema in experimental models [62]. It has also highlighted that exopolysaccharides produced by probiotic strains possess significant immunomodulatory capabilities, further supporting the role of probiotic-derived bioactive compounds in regulating immune responses [63]. α-Eleostearic acid, a representative conjugated trienoic fatty acid, demonstrates strong antioxidant activity by enhancing the activities of SOD, CAT, and GPx, while suppressing NOS activity in a sodium arsenite-induced rat model [64]. Hydroxylated linoleic acid derivatives and their amides (e.g., N-(17-hydroxy-9,12-octadecadienoyl)-glutamine) frequently act as ligands for the PPARγ receptor or modulators of inflammatory signaling pathways such as NF-κB, conferring anti-inflammatory and immunomodulatory potential [65]. Furthermore, nigrolineaxanthone M, a xanthone derivative, has shown promising activities in accelerating gastrointestinal transit, combating microbial infections, and exerting anticancer effects [66,67].
Moreover, KEGG pathway analysis showed a differential enrichment of genes in the FDT group, with significantly higher representation in key metabolic pathways including tryptophan metabolism, cyanoamino acid metabolism, glucosinolate biosynthesis, and branched-chain amino acid (BCAAs) biosynthesis (Figure 7H), and there were fewer genes involved in flavone and flavonol biosynthesis pathways in the FDT group than in the DT group (Figure 7H). Ye et al. [68] demonstrated that bacterial tryptophan catabolites could modulate intestinal motility and activate vagal neurons by engaging the TrpA1 receptor on enteroendocrine cells. Additionally, Bifidobacterium animalis subsp. lactis A6 has been shown to mitigate concurrent constipation and depression symptoms through the restoration of tryptophan metabolic balance [69]. Moreover, as one of the key metabolites of tryptophan and one indole derivatives, 5-HT also plays a key role in alleviating constipation and modulating intestinal microecology [70]. Many studies have revealed that prebiotics, probiotics and herbal medicines improved constipation by mediating 5-HT and gut microbiota [23,36,71,72]. While leucine, isoleucine, and valine (collectively known as BCAAs) are crucial for maintaining normal physiological function and metabolic balance, conversely, their chronic accumulation is associated with increased risks of metabolic disorders such as type 2 diabetes, obesity, and metabolic-associated fatty liver disease [73]. Supplementation with BCAAs in animal models has been reported to promote intestinal development and enterocyte proliferation, enhance nutrient absorption, and bolster immune defenses, while simultaneously shaping a favorable gut microbiota composition through the enrichment of beneficial bacteria [74]. All in all, the untargeted metabolomics analysis between DT and FDT suggested that probiotic fermentation elevated key bioactive metabolites (cyclo-(Gly-Pro), α-eleostearic acid, etc.), and up-regulated tryptophan metabolism and the BCAA biosynthesis process.
Our findings demonstrate strong associations between FDT-induced metabolic shifts and improved constipation symptoms, although we acknowledge that direct causal relationships remain to be established. The multitargeted nature of FDT’s therapeutic effects suggests that multiple bioactive compounds likely act synergistically. Future studies will focus on isolating key differentially abundant metabolites, particularly cyclo-(Gly-Pro), α-eleostearic acid, and hydroxylated linoleic acid derivatives, and evaluating their individual and combined effects on intestinal motility using both in vitro contractility assays and in vivo constipation models. Such approaches will help elucidate the specific molecular mechanisms and potential synergistic interactions underlying the enhanced efficacy of FDT. Furthermore, the stability of the substantial enrichment of bioactive compounds under typical storage conditions and during consumer brewing processes has not been evaluated in the present study. Factors such as temperature, humidity, light exposure, and extraction conditions could potentially affect the concentration and bioactivity of these compounds, thereby influencing the functional efficacy of FDT as a consumer product. Future studies should systematically investigate the stability of key FDT-derived bioactives (e.g., cyclo-(Gly-Pro) and hydroxylated linoleic acid derivatives) under various storage and preparation conditions in order to establish optimal handling guidelines and ensure consistent product performance. Collectively, our findings provide compelling evidence that L. paracasei K34 fermentation bio-transforms dark tea into a more potent functional food, with superior efficacy arising from the concerted action of newly generated bioactive metabolites and probiotic-mediated rebalancing of the gut microbiome. This work offers a promising microbiota-targeted strategy for managing functional constipation and highlights the potential of probiotic fermentation as a valuable approach for developing enhanced functional foods.
4. Conclusions
In this study, we have demonstrated that fermentation with L. paracasei K34 synergistically enhances the anti-constipation properties of dark tea through a multi-targeted mechanism. The FDT intervention effectively alleviated constipation symptoms, repaired intestinal epithelial damage, and restored gastrointestinal hormone balance in loperamide-induced constipated mice. Notably, FDT reversed gut microbiota dysbiosis by increasing microbial richness and selectively promoting beneficial bacterial genera, which were positively correlated with improved intestinal motility and anti-inflammatory status. Mechanistically, probiotic fermentation fundamentally alters the metabolic profile of dark tea, leading to significant enrichment of bioactive metabolites with established antioxidant and anti-inflammatory properties. Furthermore, fermentation shifts key metabolic pathways, including upregulation of tryptophan metabolism and branched-chain amino acid biosynthesis pathways intimately linked to gut motility, neural signaling, and microbial health.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bharucha A.E. Lacy B.E. Mechanisms, Evaluation, and Management of Chronic Constipation Gastroenterology 202015812321249.e 310.1053/j.gastro.2019.12.03431945360 PMC 7573977 · doi ↗ · pubmed ↗
- 2Sumida K. Molnar M.Z. Potukuchi P.K. Thomas F. Lu J.L. Yamagata K. Kalantar-Zadeh K. Kovesdy C.P. Constipation and risk of death and cardiovascular events Atherosclerosis 201928111412010.1016/j.atherosclerosis.2018.12.02130658186 PMC 6399019 · doi ↗ · pubmed ↗
- 3Nelson A.D. Camilleri M. Chirapongsathorn S. Vijayvargiya P. Valentin N. Shin A. Erwin P.J. Wang Z. Murad M.H. Comparison of efficacy of pharmacological treatments for chronic idiopathic constipation: A systematic review and network meta-analysis Gut 2017661611162210.1136/gutjnl-2016-31183527287486 · doi ↗ · pubmed ↗
- 4Sharma A. Rao S.S.C. Kearns K. Orleck K.D. Waldman S.A. Review article: Diagnosis, management and patient perspectives of the spectrum of constipation disorders Aliment. Pharmacol. Ther.2021531250126710.1111/apt.1636933909919 PMC 8252518 · doi ↗ · pubmed ↗
- 5Xu X. Wang Y. Long Y. Cheng Y. Chronic constipation and gut microbiota: Current research insights and therapeutic implications Postgrad. Med. J.202410089089710.1093/postmj/qgae 11239237119 · doi ↗ · pubmed ↗
- 6Zhang S. Wang R. Li D. Zhao L. Zhu L. Role of gut microbiota in functional constipation Gastroenterol. Rep.2021939240110.1093/gastro/goab 03534733524 PMC 8560038 · doi ↗ · pubmed ↗
- 7Pan R. Wang L. Xu X. Chen Y. Wang H. Wang G. Zhao J. Chen W. Crosstalk between the Gut Microbiome and Colonic Motility in Chronic Constipation: Potential Mechanisms and Microbiota Modulation Nutrients 202214370410.3390/nu 1418370436145079 PMC 9505360 · doi ↗ · pubmed ↗
- 8Cheng S. Li B. Ding Y. Hou B. Hung W. He J. Jiang Y. Zhang Y. Man C. The probiotic fermented milk of Lacticaseibacillus paracasei JY 062 and Lactobacillus gasseri JM 1 alleviates constipation via improving gastrointestinal motility and gut microbiota J. Dairy Sci.20241071857187610.3168/jds.2023-2415437923200 · doi ↗ · pubmed ↗