Phytochemical Profiling, Antioxidant and Antibacterial Activities of Persicaria odorata Ethanolic Leaf Extract and Evaluation of Growth Performance, Disease Resistance, and Gene Expression in Labeo chrysophekadion (Bleeker, 1849)
Sontaya Sookying, Dutrudi Panprommin, Grissana Pook-in, Pinhatai Pardubyew, Thanatip Kaeothep, Tunyatorn Nakwong, Paiboon Panase

TL;DR
This study explores the health benefits of Persicaria odorata leaf extract in fish, showing improved disease resistance and growth potential.
Contribution
The study demonstrates the potential of Persicaria odorata extract as a natural feed additive to enhance fish health and disease resilience.
Findings
POE extract contains phenolics and flavonoids with antioxidant and antibacterial properties.
Fish fed POE showed reduced mortality and upregulated immune and growth-related genes.
POE supplementation did not significantly improve growth indices but improved disease resistance.
Abstract
Persicaria odorata is a medicinal plant with various reported pharmacological properties. This study evaluated the chemical composition and biological activities of P. odorata leaf extract and assessed its effects on growth performance and disease resistance in Black Sharkminnow (Labeo chrysophekadion). Dietary supplementation with P. odorata extract showed favorable effects on fish growth and survival following Aeromonas hydrophila challenge. The extract was also associated with enhanced expression of genes related to immune function and growth regulation, suggesting its potential as a natural feed additive to support fish health and disease resilience in aquaculture. Persicaria odorata is a medicinal plant recognized for its diverse biological activities. This study investigated the phytochemical profile, antioxidant and antibacterial activities of P. odorata ethanolic leaf extract…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
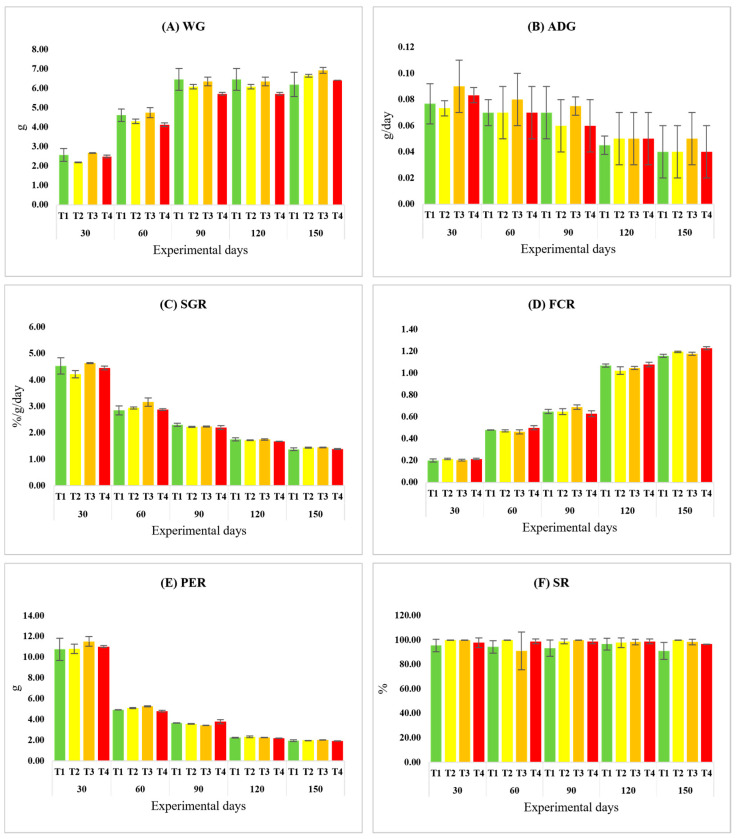 Figure 1
Figure 1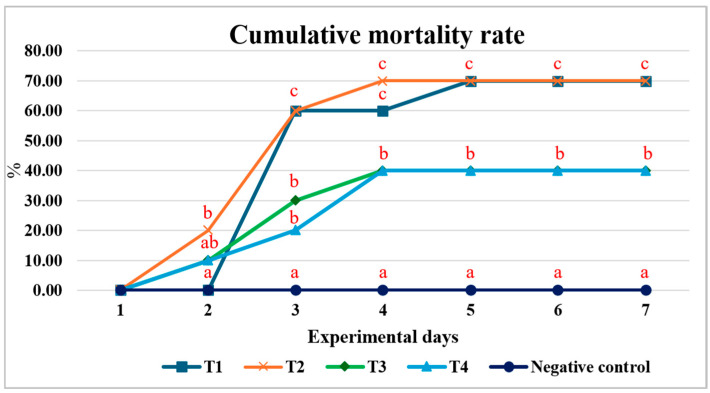 Figure 2
Figure 2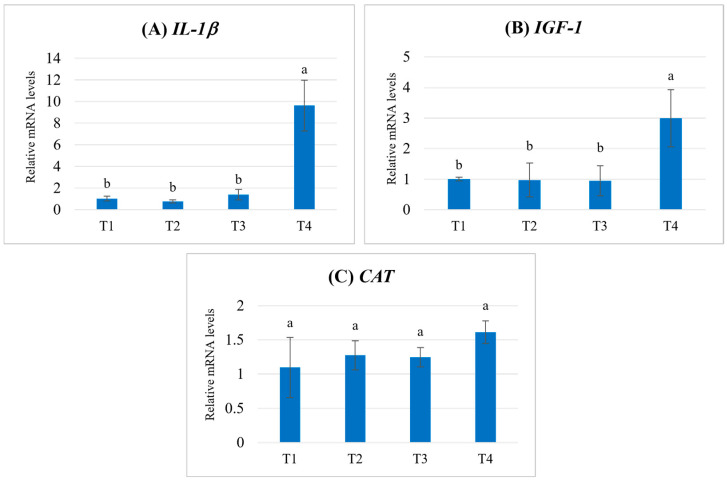 Figure 3
Figure 3- —Thailand Science Research and Innovation Fund and University of Phayao (Fundamental Fund 2025)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture disease management and microbiota · Aquaculture Nutrition and Growth · Invertebrate Immune Response Mechanisms
1. Introduction
The increasing global demand for sustainable and health-conscious aquaculture practices has prompted a shift toward the use of natural products as alternatives to synthetic antibiotics and chemical growth promoters. Among these, medicinal plants with proven pharmacological properties offer promising solutions for enhancing fish health, immunity, and growth performance without compromising environmental safety or food quality [1,2]. Persicaria odorata (Lour.) Soják, commonly known as Vietnamese coriander, is a traditional herb widely used in Southeast Asia for culinary and medicinal purposes. The bioactive constituents of P. odorata have been associated with several pharmacological properties relevant to animal health [3]. Previous phytochemical studies have reported a wide array of bioactive compounds in P. odorata leaves, including flavonoids (e.g., rutin, quercetin, luteolin, apigenin), phenolic acids (e.g., gallic acid, ferulic acid), terpenoids (e.g., β-caryophyllene, α-humulene), and sulfur-containing compounds such as 3-sulfanylhexanal [4,5,6], which have been associated with antioxidant [7,8], antimicrobial [7,9,10], anti-inflammatory, and immunostimulatory activities [9,11,12,13,14,15].
Consequently, scientific studies have demonstrated its ability to inhibit pathogenic bacteria [9,10], modulate immune responses [9], enhance growth performance without hepatotoxicity [15] and improve physiological parameters in various animal models [9,15]. These findings suggest that P. odorata may serve as a viable phytotherapeutic agent in aquaculture systems. To explore its potential, this study first investigated the phytochemical profile of P. odorata ethanol extract (POE), including preliminary screening, quantification of key compounds, and evaluation of antioxidant and antibacterial activities. Synergistic effects with conventional antibiotics were also assessed to determine its role in enhancing antimicrobial efficacy. These properties collectively position P. odorata as a promising candidate for phytogenic feed additives in livestock and aquaculture production. This potential has begun to be explored in terrestrial animals, particularly in poultry.
The application of P. odorata as a functional feed additive in animal production is an emerging area of research. A seminal study by Zhang et al. [16] investigated the effects of dietary supplementation with P. odorata fibrous root powder in broiler chickens, demonstrating significant improvements in growth performance parameters, enhanced serum antioxidant capacity, and favorable modulation of cecal microbiota by enriching beneficial butyrate-producing bacteria. In aquaculture, the use of medicinal plants as dietary supplements has gained considerable traction as a strategy to enhance disease resistance, improve feed utilization, and reduce antibiotic dependence [17,18]. Several phytogenic additives have been reported to improve growth performance, stimulate immune gene expression and confer protection against bacterial pathogens in farmed fish [19,20,21,22]. However, despite the well-documented bioactivities of P. odorata and its proven benefits in poultry, studies evaluating its application in aquatic animals remain remarkably scarce. To date, no comprehensive investigation has been conducted on the effects of dietary P. odorata extract on growth performance, immune-related gene expression, and disease resistance in Labeo chrysophekadion, a freshwater fish of significant economic value in Southeast Asia.
Therefore, this study was designed to address this knowledge gap through a two-fold approach. First, the objective was to characterize the phytochemical profile of an ethanolic leaf extract of P. odorata (POE), quantify its key bioactive compounds, and evaluate its in vitro antioxidant and antibacterial activities. Second, the study sought to assess the effects of dietary POE supplementation in juvenile Labeo chrysophekadion. Growth performance indices, resistance to Aeromonas hydrophila infection, and the expression of genes associated with growth (insulin-like growth factor 1; IGF-1), immunity (interleukin-1β; IL-1β), and antioxidant defense (catalase; CAT) were examined. This integrative approach provides the first evaluation of P. odorata as a potential functional feed additive in aquaculture.
2. Materials and Methods
2.1. Ethical Approval and Authorization for the Use of Animals
All experimental procedures received prior approval from the Institutional Animal Care and Use Committee of the University of Phayao, Thailand (Approval ID: 1-012-67; Approval Date: 1 August 2024) and were carried out in compliance with the university’s ethical regulations.
2.2. Chemicals and Reagents
HPLC-grade methanol and acetonitrile were obtained from RCI Labscan (Dublin, Ireland). Unless otherwise specified, all chemicals used in the experimental procedures were of analytical grade. Methanol (99.9%) and ethanol (99.9%) were supplied by RCI Labscan. Anhydrous sodium carbonate (99.5%) and aluminum chloride hexahydrate (95%) were purchased from KemAus (Sydney, Australia), whereas sodium hydroxide (99%) was also sourced from RCI Labscan. Gallic acid (98%) was obtained from AK Scientific (Union City, CA, USA). L-ascorbic acid (99.7%) and quercetin dihydrate (98%) were procured from Sisco Research Laboratories (Mumbai, India). Folin–Ciocalteu’s phenol reagent was supplied by Loba Chemie (Mumbai, India). In addition, 2,2-diphenyl-1-picrylhydrazyl (DPPH), catechin, and rutin were acquired from Sigma-Aldrich (Burlington, VT, USA).
Muller-Hilton agar (MHA) and broth (MHB), and Brain heart infusion agar (BHIA) were purchased from HiMedia Laboratories, Mumbai, India.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) was conducted utilizing reagents sourced from various suppliers. MS-222 solution was obtained from Sigma (St. Louis, MO, USA), while TRIzol reagent was acquired from the Molecular Research Center (Cincinnati, OH, USA). Complementary DNA synthesis was conducted utilizing the iScript™ Select cDNA Synthesis Kit (Bio-Rad, Hercules, CA, USA). The amplification reactions employed a 2X master mix derived from the THUNDERBIRD™ SYBR^®^ qPCR Mix (TOYOBO, Osaka, Japan).
2.3. Plant Material and Preparation of Persicaria odorata Ethanolic Leaf Extract
Fresh leaves of P. odorata were collected from a local market in Phayao Province, Thailand. Whole plants from the same source were additionally obtained for voucher specimen preparation. The plant materials were pressed and air-dried prior to taxonomic verification by a qualified botanist. The authenticated specimen was then deposited at the Queen Sirikit Botanic Garden (QBG) Herbarium under voucher number 149873.
After thorough washing, the leaves were dried in a hot-air oven at a controlled temperature below 40 °C. The dried samples were pulverized into powder, weighed, and macerated with 99.9% ethanol at a 1:5 (w/v) ratio. The extraction was conducted for 48 h with intermittent agitation every 12 h. The mixture was subsequently filtered through standard filter paper (pore size 20–25 µm) and concentrated to dryness using rotary evaporation (BÜCHI Labortechnik AG, Flawil, Switzerland) at temperatures below 45 °C. The resulting crude extract of P. odorata (POE) showed a yield of 20.18%. The extract was kept in a tightly sealed, light-protected container at temperatures below 8 °C until use [7].
2.4. Phytochemical Screening
The phytochemical profile of POE was analyzed using established standard methods as previously described [23,24,25]. Alkaloids were detected using Dragendorff’s, Wagner’s, and Mayer’s reagents. Terpenoid constituents and steroid compounds were analyzed by the Liebermann–Burchard reaction. Carbohydrates were assessed through Molisch’s, Fehling’s, and Benedict’s tests, whereas deoxy sugars were characterized using the Keller–Killiani method. The presence of α,β-unsaturated five-membered lactone rings was verified by Kedde’s test. Cyanogenic glycosides were evaluated via the sodium picrate assay, and saponins were examined using the frothing test. Phenolics and tannins were screened with ferric chloride reagent and gelatin solution, respectively. Coumarins were detected by the sodium hydroxide paper method, while anthraquinones were analyzed according to a modified Borntrager’s procedure.
2.5. Quantification of Bioactive Compounds
2.5.1. Total Phenolic Content
Total phenolic content (TPC) of the extract was quantified using the Folin–Ciocalteu colorimetric method according to the procedure described by Sookying et al. [26]. Gallic acid served as the calibration standard, and a standard curve was generated over a concentration range of 3.125–200 µg/mL. The calibration equation obtained from linear regression was y = 0.005x − 0.0569, with a coefficient of determination (R^2^) of 0.9973, indicating excellent linearity. The TPC was calculated and reported as mg GAE/g extract. All measurements were performed in triplicate.
2.5.2. Total Flavonoid Content
Total flavonoid content (TFC) was quantified using an assay following the procedure described by Sookying et al. [26]. Quercetin was applied as the reference compound. Standard quercetin and POE were prepared in methanol (80% v/v in water). A calibration curve was established with quercetin (3.125–200 µg/mL). The TFC values were calculated from the quercetin standard curve (y = 0.0007x − 0.0008, R^2^ = 0.9989) and expressed as mg QE/g extract. All assays were performed in triplicate.
2.5.3. Catechin, Quercetin and Rutin Contents
A high-performance liquid chromatography (HPLC) method was developed and validated for the quantitative determination of rutin, quercetin, and catechin. Chromatographic analysis was performed using a Shimadzu LC-2050C system (Shimadzu Corporation, Kyoto, Japan). Separation was achieved on a silica-based C18 analytical column fitted with a guard cartridge (Inertsil, 150 mm × 4.6 mm, 5 µm; GL Sciences Inc., Tokyo, Japan). The mobile phase comprised methanol, acetonitrile, and water containing 2% acetic acid and was delivered under gradient conditions at 1 mL/min. Sample injection volume was set at 10 µL, and total run time was 15 min. Detection wavelengths were 276 nm for catechin and 368 nm for quercetin and rutin. System control was carried out using LC Solutions software (version 1.25) (Shimadzu Corporation, Singapore), while data processing was conducted with PostRun Analysis software (version 2.1) (Shimadzu Corporation, Singapore). Detailed system parameters and method validation data are provided in the Supplementary Materials.
2.6. Antioxidant Capacity Assays
The antioxidant capacity of POE was determined using the DPPH radical scavenging assay according to the procedure previously reported by Sookying et al. [26], with minor modifications. Briefly, DPPH solution (0.1 mM) was prepared in methanol, and sample solutions were diluted in 80% methanol. Equal volumes (100 µL) of sample or standard and DPPH solution were combined in a 96-well microplate and incubated for 30 min in the dark. Absorbance was recorded at 517 nm. Radical scavenging activity was expressed as percentage inhibition relative to the control, and the IC_50_ value was calculated from the concentration–response curve. All assays were conducted in triplicate.
2.7. Antibacterial Activity Evaluation
The antibacterial activity of POE was assessed against six bacterial strains: Staphylococcus aureus ATCC25923, Staphylococcus epidermidis ATCC12228, Bacillus cereus DMST6229, Cutibacterium acnes ATCC14916, Escherichia coli ATCC25922, and Pseudomonas aeruginosa ATCC27853. Antibacterial efficacy was determined by measuring the minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) following the method described by Elshikh et al. [27] with minor modifications. Briefly, bacterial suspensions adjusted to McFarland Standard No. 0.5 were inoculated into 96-well microplates containing Mueller–Hinton broth (MHB) and serial two-fold dilutions of POE (starting from 250 mg/mL). After incubation at 37 °C for 24 h, MIC values were determined using a resazurin-based colorimetric assay. The lowest concentration that prevented the color change from blue to pink was recorded as the MIC. For MBC determination, aliquots from wells showing no visible growth were subcultured onto Mueller–Hinton agar (MHA) plates and incubated at 37 °C for 24 h. The lowest concentration showing no colony formation was defined as the MBC. For C. acnes, brain heart infusion agar (BHIA) was used, and plates were incubated under anaerobic conditions for 48–72 h. All experiments were performed in triplicate.
2.8. Synergistic Effects Evaluation
The synergistic activity of POE in combination with tetracycline or gentamicin was evaluated by co-treatment at various concentration ratios. MIC and MBC values for the combinations were determined as described in Section 2.7. Drug interactions were interpreted using the fractional inhibitory concentration index (FICI). The effects were categorized as synergistic (≤0.5), additive (>0.5 and ≤1.0), indifferent (>1.0 and <4.0), or antagonistic (≥4.0) according to established criteria [28].
2.9. Growth Performance Study in Labeo chrysophekadion
2.9.1. Experimental Fish and Acclimatization Conditions
Juvenile Labeo chrysophekadion were purchased from a commercial fish farm in Phayao Province, Thailand. They were held in a net cage (5 m × 3 m × 2 m) placed in earthen ponds and allowed to acclimate for four weeks under natural light–dark cycles. Throughout the acclimation period, essential water quality variables were regularly checked and kept within specified optimal ranges: temperature 28.6 ± 3.18 °C, dissolved oxygen 6.50 ± 1.8 mg/L, and pH 7.8 ± 1.52, measured using a multiparameter probe (HORIBA U50 series, Kyoto, Japan). During acclimation, the fish were fed a commercial feed containing 40% crude protein twice a day (at 08:00 a.m. and 05:00 p.m.) at a daily ration of 4% of their body weight, and no POE was added to the diet during this period.
2.9.2. Preparation for a POE-Supplemented Diet
Commercial fish feed pellets (2 mm diameter; Hi-grade 9961, CPF Co., Ltd., Bangkok, Thailand) served as the basal diet. The feed composition comprised 40% crude protein, 4% crude fat, 12% moisture, and 4% crude fiber, and was formulated from fish meal, soybean meal, broken rice, maize, vitamins, and minerals. POE was initially dissolved in 100 mL of deionized water before incorporation into the basal diet. Four dietary treatments were prepared: a control diet without POE (0.00 g/kg; T1) and three experimental diets supplemented with 0.25 g/kg (T2), 0.50 g/kg (T3), and 0.75 g/kg (T4). The extract solution was uniformly applied to the pellets using a pan coating machine equipped with an air blower (CM/Thai, CMCA-10 model; Pharmaceutical and Medical Supply Co., Ltd., Samut Sakhon, Thailand). Subsequently, the coated pellets were treated with 4% agar solution (10 mL/kg feed) and air-dried prior to use [20]. Prepared diets were portioned into sterile containers and stored at room temperature. Diet preparation was conducted weekly to ensure feed quality and palatability.
2.9.3. Experimental Design
After the acclimation period, healthy fish (0.84 ± 0.43 g mean body weight) were allocated to twelve net cages (1 m × 2 m × 0.8 m; mesh size 2.5 mm). The experiment consisted of four dietary treatments, each conducted in triplicate. Stocking density was standardized at 30 fish per cage (approximately 15 fish/m^2^), and all cages were installed within the same earthen pond. Throughout the 150-day feeding trial, fish received their respective experimental diets at 4% of body weight per day, administered in two equal meals. Water quality parameters were recorded regularly, with temperature ranging from 29.5 to 31.2 °C, dissolved oxygen from 6.3 to 7.3 mg/L, pH between 7.12 and 8.43, and total dissolved solids from 0.24 to 0.55 g/L.
2.9.4. Growth Performance Study
All fish in each cage were batch-weighed every 15 days to recalibrate feeding rates. Growth performance was assessed periodically, and the resulting data were compiled at 30-day intervals. The evaluated growth metrics included weight gain (WG), average daily gain (ADG), specific growth rate (SGR), feed conversion ratio (FCR), protein efficiency ratio (PER), and survival rate (SR). These parameters were determined according to the formulae presented below [29].
2.10. Pathogenic Challenge Test
The Aeromonas hydrophila strain DMST 21250 was sourced from the Department of Medical Sciences under the Ministry of Public Health, Nonthaburi, Thailand. The bacterium was propagated aerobically in Tryptic Soy Broth at 37 °C for 18–24 h to obtain log-phase cultures. Cell density was adjusted with sterile physiological saline to 0.5 McFarland standard (approximately 1.0 × 10^8^ CFU/mL), verified by serial dilution and plate counting. Fresh bacterial suspensions were prepared daily and kept on ice prior to administration. The median lethal dose (LD_50_) was determined in advance to establish the challenge dose (10^8^ CFU/mL) for Labeo chrysophekadion. For the resistance assay, 60 fish per treatment (20 fish per replicate) were randomly selected using a portable hand net. After 150 days of dietary feeding, fish in the treatment groups were intraperitoneally injected with 0.1 mL of bacterial suspension (10^8^ CFU/mL), whereas control fish received 0.1 mL of sterile physiological saline. Post-challenge mortality was monitored daily for 7 days, and cumulative mortality (%) was calculated and graphically presented.
2.11. Gene Expression Analysis
2.11.1. Total RNA Extraction and cDNA Synthesis
For gene expression analysis, six fish per treatment were randomly sampled. Fish were anesthetized with MS-222 (0.2 g/L) prior to tissue collection. Liver samples were aseptically excised and immediately processed for total RNA isolation using TRIzol reagent (Molecular Research Center Inc., Cincinnati, OH, USA) in accordance with the manufacturer’s protocol. First-strand cDNA was synthesized from 1 µg of total RNA using the iScript™ Select cDNA Synthesis Kit (Bio-Rad Laboratories, Hercules, CA, USA) following the supplier’s instructions.
2.11.2. RT-qPCR
Transcript levels of IL-1β, IGF-1, and CAT were quantified by real-time PCR using gene-specific primers obtained from the previous studies (see Table 1 for sequences and amplicon sizes). β-actin was selected as the reference gene for normalization. Each 20 µL reaction contained SYBR Green master mix (THUNDERBIRD™ SYBR^®^ qPCR Mix, TOYOBO, Osaka, Japan), forward and reverse primers (10 µM), first-strand cDNA, and nuclease-free water. Amplification was conducted under the following program: enzyme activation at 95 °C for 1 min, followed by 40 amplification cycles consisting of denaturation (95 °C, 15 s), annealing (60 °C, 15 s for IL-1β and IGF-1; 57 °C, 15 s for CAT), and extension (72 °C, 10 s). Product specificity was confirmed by dissociation curve analysis over a temperature gradient from 55 °C to 95 °C with a temperature increment of 0.3 °C/s. All samples were analyzed in triplicate. Relative expression was determined by the comparative Ct (2^−ΔΔCt^) approach [30], with target gene expression normalized to β-actin and expressed relative to the control group.
2.12. Statistical Analysis
All statistical analyses were performed using SPSS (version 25.0; IBM Corp., Armonk, NY, USA). Differences among treatment groups were evaluated by one-way analysis of variance (ANOVA), and when significant effects were detected, Tukey’s post hoc test was applied for pairwise comparisons. Statistical significance was established at p < 0.05. Assumptions of normal distribution and homogeneity of variance were verified using the Shapiro–Wilk and Levene’s tests, respectively. Data are presented as mean ± standard deviation (SD).
3. Results
3.1. Phytochemical Constituents of the Extract
Phytochemical screening indicated the presence of alkaloids, anthraquinones, steroids, triterpenoids, volatile and nonvolatile coumarins, phenolics, flavonoids, tannins, and carbohydrates in POE (Table 2). Saponins, cyanogenic glycosides, and cardiac glycosides were not detected.
3.2. Total Phenolic and Total Flavonoid Contents
The total phenolic content (TPC) and total flavonoid content (TFC) of POE are presented in Table 3. The TPC was 140.43 ± 31.82 mg GAE/g extract, whereas the TFC was 122.86 ± 0.71 mg QE/g extract.
3.3. Catechin, Quercetin and Rutin Contents
HPLC analysis confirmed the presence of catechin and quercetin in POE, while rutin was not detected under the analytical conditions employed. The concentrations of catechin and quercetin were 0.08 ± 0.02% and 1.91 ± 0.16% (w/w), respectively (Table 3).
3.4. Antioxidant Capacity
The antioxidant capacity of POE, as evaluated by the DPPH assay, yielded an IC_50_ value of 86.30 ± 0.03 µg/mL (Table 4). In comparison, the reference standard ascorbic acid showed an IC_50_ of 13.00 ± 0.03 µg/mL.
3.5. Antibacterial Activities Against Multiple Bacterial Species and Synergistic Interaction in Combination with Antibiotics
The MIC values of POE against S. aureus, S. epidermidis, B. cereus, C. acnes, and P. aeruginosa were 7.81 mg/mL, whereas a higher MIC of 15.63 mg/mL was recorded for E. coli (Table 5). The MBC values ranged from 7.81 to 125 mg/mL depending on the bacterial strain. Specifically, MBC values of 7.81, 15.63, and 31.25 mg/mL were obtained for C. acnes, S. epidermidis, and S. aureus, respectively, while the remaining strains exhibited an MBC of 125 mg/mL.
For the combination study, MIC values of POE in the presence of antibiotics were determined and the fractional inhibitory concentration index (FICI) was calculated (Table 5). The POE–tetracycline combination yielded a synergistic interaction against S. aureus and an additive effect against B. cereus. An indifferent interaction was observed for C. acnes. In contrast, antagonistic interactions were recorded for S. epidermidis, E. coli, and P. aeruginosa.
3.6. Growth Performance in Labeo chrysophekadion
Over the 150-day feeding trial involving four dietary POE levels, including the control, no statistically significant differences were observed among treatments for any growth performance indices, including WG, ADG, SGR, FCR, PER, and SR (p > 0.05) (Figure 1A–F).
3.7. Cumulative Mortality Rate Following Pathogenic Exposure
After the 150-day feeding trial, fish in each treatment group were intraperitoneally injected with an A. hydrophila suspension (10^8^ CFU/mL), whereas the negative control group received an equal volume of 0.85% physiological saline. Observations for clinical signs and mortality were recorded continuously for 7 days post-injection. The results showed that groups T1 and T2 exhibited the highest cumulative mortality (70%). Both T1 and T2 reached their highest cumulative mortality on day 2 post-injection, and T1 experienced continued mortality through day 5 post-injection. While groups T3 and T4 had equal cumulative mortality rates (40%), which were significantly lower than those of T1 and T2. Additionally, mortalities in T3 and T4 occurred only during the first 4 days post-injection (Figure 2).
3.8. Expression of Growth, Immune, and Antioxidant-Related Genes
The relative mRNA expression levels of IL-1β, IGF-1, and CAT in fish fed diets supplemented with different concentrations of POE are shown in Figure 3. For IL-1β, expression levels remained low and not significantly different among the T1–T3 groups, whereas a marked upregulation was observed in the T4 group (p < 0.05) (Figure 3A). Similarly, IGF-1 expression was comparable among T1–T3, but fish receiving the highest supplementation level (T4) exhibited a significant increase in transcript abundance (p < 0.05) (Figure 3B). For CAT, no significant differences in relative mRNA expression levels were detected among fish fed diets supplemented with POE at different concentrations (T1–T4) (p > 0.05). Nevertheless, the T4 group showed a tendency toward higher expression compared with the other treatments (Figure 3C).
4. Discussion
4.1. Phytochemical Composition of POE
The phytochemical constituents detected in Persicaria odorata extract (POE) in the present study were generally consistent with the previous summary reported by Azmi et al. [34]. The presence of alkaloids and saponins in P. odorata leaves has been reported inconsistently across earlier studies, suggesting that their occurrence may vary depending on extraction conditions, plant origin, and analytical methods. In contrast, cyanogenic glycosides and cardiac glycosides were not detected in the POE prepared in this study, which is in agreement with previous phytochemical screening reports [34]. P. odorata is known to be an aromatic plant rich in essential oils, with many bioactive constituents occurring in volatile forms, particularly terpenoids. Nevertheless, the results of the phytochemical screening tests indicated that POE contained multiple classes of secondary metabolites, each potentially contributing to distinct biological activities. Among these, phenolic compounds and flavonoids are of particular interest due to their well-documented antioxidant properties. Accordingly, TPC and TFC were quantitatively analyzed in this study. The results revealed that POE obtained in the present work possessed substantially higher TPC and TFC compared with ethanolic and aqueous P. odorata leaf extracts previously reported by Nguyen et al. [35], with increases of approximately 2.39- and 1.73-fold, respectively (140.43 vs. 58.56 mg GAE/g extract and 122.86 vs. 70.65 mg QE/g extract, respectively). This enhanced phytochemical content was consistent with the antioxidant capacity assessed by the DPPH radical scavenging assay. The IC_50_ value obtained in this study corresponded to 6.63-fold that of ascorbic acid, whereas Nguyen et al. [35] reported a markedly weaker activity, with an IC_50_ equivalent to 64.84-fold that of ascorbic acid.
In this study, rutin was not detected in POE, which is consistent with the findings reported by Pawłowska et al. [36]. Quercetin was also identified at a relatively high level (1.91% w/w), further supporting the richness of flavonoid constituents in the extract. Collectively, these findings suggest that POE contains phenolic and flavonoid constituents, as indicated by total phenolic and total flavonoid contents and the quantified presence of quercetin and catechin. These phytochemicals may underlie its antioxidant capacity and contribute, at least in part, to its antibacterial and health-promoting effects observed in subsequent biological assays. The present study also demonstrated that POE exhibited antibacterial activity against both Gram-negative and Gram-positive bacteria, with MIC values ranging from 7.81 to 15.63 mg/mL. These findings are consistent with previous reports indicating that plant-derived extracts rich in flavonoids and phenolic compounds can inhibit bacterial growth through disruption of cell wall integrity and membrane permeability [37,38]. These compounds also exert antibacterial activity through ROS-mediated oxidative stress, DNA interaction, and inhibition [39].
4.2. Antibacterial Activity and Synergistic Effects
The interaction between POE and antibiotics varied depending on bacterial species, as reflected by the FICI. A synergistic interaction (FICI = 0.50) was observed against S. aureus, suggesting that POE may enhance antibiotic efficacy, possibly by increasing bacterial membrane permeability or interfering with cellular defense mechanisms. In contrast, antagonistic interactions were observed in Gram-negative bacteria, including E. coli and P. aeruginosa. This phenomenon may be attributed to the presence of an outer membrane in Gram-negative bacteria, which serves as an effective permeability barrier that restricts the penetration of phytochemicals and antibiotics. Although POE did not exhibit synergistic antibacterial effects against Gram-negative bacteria in vitro, the extract still demonstrated bactericidal activity at higher concentrations, as indicated by its MBC values. However, it is important to note that the role of herbal extracts in aquaculture extends beyond direct antimicrobial activity.
4.3. Effects of POE on Growth Performance
Growth performance did not significantly improve after POE supplementation, according to the current study. However, the effects of dietary supplements may vary depending on the specific bioactive constituents, inclusion levels, fish species, and rearing conditions. Herbal-derived bioactive compounds—such as flavonoids, phenolics, terpenoids, and alkaloids—primarily exert immunomodulatory and antioxidant functions rather than directly promoting growth. These substances do not directly improve nutritional digestion, absorption, or anabolic metabolism [40,41]. Moreover, the supplementation level used in this study may not have reached the threshold necessary to elicit significant growth responses. Effective dosages for enhancing immune activity or antioxidant defense are frequently lower than those required to promote growth. Accordingly, dosage selection in numerous studies has been guided by immunological indicators or pathogen challenge outcomes rather than growth optimization [42]. Conversely, elevated dosages aimed at promoting growth may occasionally exhibit immunosuppressive effects or induce palatability concerns, therefore undermining any advantages [43]. Healthy fish typically reach their natural development potential under carefully regulated rearing settings with nutrient-dense meals and no pathogenic pressure. Consequently, the inclusion of immunostimulatory agents under optimal conditions may not lead to marked improvements in growth performance. The benefits of such additives are more likely to become evident under challenging conditions, such as bacterial-related illnesses or stressful environment, where enhanced immune readiness plays a protective role [44]. Additional study may be necessary to investigate the possible advantages of POE on growth efficiency under various stressful circumstances or in conjunction with other nutritional supplements.
4.4. Effects of POE on Disease Resistance
The improved survival observed in fish fed diets containing 0.50 and 0.75 g/kg POE suggests a potential immunostimulatory and/or antibacterial role of POE in increasing resistance to A. hydrophila challenge. These findings align with those of Sookying et al. [26], which demonstrated that dietary inclusion of Phlogacanthus pulcherrimus leaf extract at 0.25–0.75 g/kg did not affect the growth performance of L. chrysophekadion, but conferred protection against A. hydrophila (10^8^ CFU/mL) infection in fish fed 0.75 g/kg, reducing cumulative mortality by up to 20% relative to the control. Previous studies have documented the prophylactic efficacy of various plant extracts against bacterial diseases in fish, demonstrating the potential of natural substances as antibiotic substitutes in aquaculture [45]. Furthermore, combinations of herbal extracts and probiotics have been shown to improve hematological indices, immune responses, and antioxidant status in fish [46]. All of these results show that herbal extracts can significantly improve fish health and lessen the need for artificial chemotherapeutics in aquaculture systems [47].
4.5. Modulation of Immune, Growth, and Antioxidant-Related Gene Expression
The upregulation of IL-1β and IGF-1 transcripts was observed in fish fed the highest level of POE. In teleosts, IL-1β serves as a pivotal pro-inflammatory cytokine, playing an essential role in initiating innate immune responses [48]. The increased expression observed in this study suggests that POE can stimulate immune activation. Moreover, this upregulation of IL-1β was consistent with the reduced mortality observed in experimental fish challenged with the pathogenic bacterium A. hydrophila. Similarly, Honghirun et al. [49] demonstrated that dietary supplementation with P. odorata leaf powder at 30 g/kg enhanced immune responses, including the upregulation of IL-1β, in Nile tilapia (Oreochromis niloticus). Comparable effects were also reported with other phytogenic additives, such as P. pulcherrimus, which upregulated IL-1β and improved resistance to A. hydrophila in L. chrysophekadion [26], and with Ocimum sanctum (holy basil), which also enhanced IL-1β expression and improved tolerance against Streptococcus agalactiae in Nile tilapia (O. niloticus) [50]. Similarly, IGF-1 was also stimulated at the highest level of POE. IGFs play a fundamental role in mediating growth in vertebrate species, including fish [51]. This finding is consistent with the weight gains observed in all experimental groups fed POE, although no statistically significant differences were detected, which agrees with the results obtained by Sookying et al. [26]. Several phytogenic supplements have also been reported to upregulate IGF-1 expression, including myrtle (Myrtus communis) powder in zebrafish (Danio rerio) [52], and turmeric (Curcuma longa) powder in juvenile sevruga (Acipenser stellatus) [53].
Although no statistically significant differences in CAT gene expression were detected among treatments (p > 0.05), POE-supplemented groups exhibited numerically higher expression levels compared with the control. Comparable a finding has been reported in common carp (Cyprinus carpio), where curcumin administration did not produce significant differences among treatment groups, although CAT mRNA expression was slightly elevated at dietary levels of 10 and 15 g/kg [54]. Therefore, further investigation incorporating additional oxidative stress-related genes, such as superoxide dismutase (SOD) and glutathione peroxidase (GPx), would be necessary to provide a more comprehensive evaluation of antioxidant status.
In the in vivo challenge experiment, fish fed POE-supplemented diets showed improved survival and health status following infection with A. hydrophila, a Gram-negative rod-shaped microbe frequently linked to severe disease outbreaks in aquaculture. This protective effect is likely mediated through indirect mechanisms rather than direct bactericidal action. Dietary supplementation with POE may enhance host resistance by modulating the immune response, as evidenced by the regulation of pro-inflammatory cytokines including IL-1β, which are pivotal in the first activation of innate immunity. In addition, the upregulation of antioxidant-related genes, including CAT, suggests that POE contributes to the mitigation of oxidative stress induced by bacterial infection. Reducing oxidative damage may help maintain cellular integrity and physiological homeostasis during pathogenic challenges. Furthermore, the elevated expression of IGF-1 in POE-fed fish indicates improved growth performance and tissue recovery, which are essential for resilience and post-infection recovery. Together, these findings suggest that POE functions as a functional feed additive that enhances disease resistance through immunomodulatory and antioxidant pathways rather than acting solely as a direct antibacterial agent. These findings indicate that POE supplementation at 0.75 g/kg resulted in numerically higher CAT expression; however, the differences were not statistically significant. Further studies should therefore explore optimal dosage levels and experimental durations to maximize its antioxidant potential.
Overall, the present findings indicated that dietary POE supplementation enhanced resistance against A. hydrophila, particularly at 0.75 g/kg, and this effect was associated with a significant upregulation of the immune-related gene IL-1β. In contrast, growth performance and CAT expression were not significantly affected, suggesting that POE primarily exerted immunomodulatory effects rather than broadly influencing growth or antioxidant gene transcription. These results were consistent with the findings of Sookying et al. [26], who reported similar immunostimulatory effects of phytogenic additives in the same fish species. Further investigations incorporating additional oxidative stress–related genes and long-term feeding trials to evaluate growth performance would provide deeper insight into the mechanisms underlying the protective effects of POE.
5. Conclusions
This study demonstrated that ethanolic leaf extract of P. odorata (POE) contains diverse bioactive compounds, including phenolics and flavonoids (e.g., quercetin, catechin), which contribute to its pronounced in vitro antioxidant and antibacterial activities. When supplemented in the diet of L. chrysophekadion at 0.50 and 0.75 g/kg for 150 days, POE did not significantly enhance growth performance indices but markedly improved survival following A. hydrophila challenge. This protective effect was associated with the upregulation of immune-related (IL-1β) and growth-related (IGF-1) genes, suggesting immunomodulatory and physiological regulatory roles for POE. Although CAT expression was not significantly increased, the numerical trend at the highest supplementation level suggests a potential antioxidant contribution that warrants further investigation.
Application prospects: The findings highlight the potential of POE as a natural functional feed additive in aquaculture, particularly for enhancing disease resistance and reducing reliance on antibiotics. The effective dietary inclusion level of 0.50–0.75 g/kg identified in this study provides a practical reference for future applications in fish farming, especially under conditions of bacterial disease outbreaks.
Limitations and future directions: Several limitations should be acknowledged. The lack of statistically significant improvements in growth and CAT expression may be attributed to the dosage levels, experimental duration, or the absence of pathogenic stress during the growth trial. Additionally, the precise molecular mechanisms underlying the immunomodulatory effects of POE, particularly the involvement of the Nrf2 pathway, were not fully elucidated. Future studies should explore a wider range of POE inclusion levels, longer feeding periods, and challenges under various stress conditions. Investigations at the protein level and using pathway-specific inhibitors would help clarify the mechanistic basis of POE’s effects. Furthermore, extending the antibacterial screening to include A. hydrophila and other fish pathogens would complement the in vivo findings.
In conclusion, this study provides the first evidence supporting the use of P. odorata extract as a promising phytogenic additive to improve disease resistance and health status in aquaculture species, contributing to the advancement of sustainable and environmentally friendly aquatic animal production.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tadese D.A. Song C. Sun C. Liu B. Liu B. Zhou Q. Kevin N.T. The role of currently used medicinal plants in aquaculture and their action mechanisms: A review Rev. Aquac.20221481684710.1111/raq.12626 · doi ↗
- 2Sumana S.L. Xue T. Hu H. Abdullateef M.M. Shui Y. Ayana G.U. Kayiira J.C. Zhang C. Samwel B.J. Zhu J. Medicinal plants as ecological solutions for fish growth and immunostimulatory effects in aquaculture Aquac. Res.20251977862310.1155/are/9778623 · doi ↗
- 3Bi J. Fang H. Zhang J. Lu L. Gu X. Zheng Y. A review on the application, phytochemistry and pharmacology of Polygonatum odoratum, an edible medicinal plant J. Future Foods 2023324025110.1016/j.jfutfo.2023.02.006 · doi ↗
- 4Starkenmann C. Luca L. Niclass Y. Eric P. Roguet D. Comparison of volatile constituents of Persicaria odorata (Lour.) Soják and Persicaria hydropiper J. Agric. Food Chem.2006543067307110.1021/jf 053161116608232 · doi ↗ · pubmed ↗
- 5Khuayjarernpanish T. Sookying S. Duangjai A. Saokaew S. Sanbua A. Bunteong O. Rungruangsri N. Suepsai W. Sodsai P. Soylaiad J. Anticancer activities of Polygonum odoratum Lour.: A systematic review Front. Pharmacol.20221387501610.3389/fphar.2022.87501635571080 PMC 9092523 · doi ↗ · pubmed ↗
- 6ŘebíčkováK. Bajer T. Šilha D. HoudkováM. Ventura K. BajerováP. Chemical composition and determination of the antibacterial activity of essential oils extracted from Houttuynia cordata and Persicaria odorata Molecules 202025243210.3390/molecules 2510243232456033 PMC 7287994 · doi ↗ · pubmed ↗
- 7Ahongshangbam S.K. Shantibala Devi G.A. Chattopadhyay S. Bioactive compounds and antioxidant activity of Polygonum odoratum Lour Int. J. Basic Appl. Biol.201429497
- 8Kawvised S. Prabsattroo T. Munkong W. Pattum P. Iamsaard S. Boonsirichai K. Uttayarat P. Maikaeo L. Sudchai W. Kirisattayakul W. Polygonum odoratum leaf extract attenuates oxidative stress and cell death of RAW 264.7 cells exposed to low-dose ionizing radiation J. Food Biochem.202145 e 1390910.1111/jfbc.1390934423456 · doi ↗ · pubmed ↗