Dietary Effects of Carotenoid-Biofortified Wheat on Feed Conversion and Tissue Antioxidant Concentrations in Broiler Chickens
Jan Szmek, Michaela Englmaierová, Miloš Skřivan, Věra Skřivanová, Pavel Horčička, Eva Pěchoučková

TL;DR
A study found that feeding chickens a special type of wheat improved their feed efficiency and increased antioxidants in their meat.
Contribution
The study demonstrates that carotenoid-biofortified wheat improves feed conversion and tissue antioxidant concentrations in broiler chickens.
Findings
Pexeso wheat improved feed conversion ratio and crude protein retention in broiler chickens.
Pexeso wheat increased γ-tocopherol, lutein, and zeaxanthin concentrations in chicken breast meat and liver.
The improved antioxidant profile suggests enhanced nutritional quality of poultry meat.
Abstract
This study aimed to investigate the effects of carotenoid-biofortified Pexeso wheat compared with those of common Tercie wheat on performance characteristics, nutrient retention, and tissue antioxidant concentrations in broiler chickens. A total of 180 one-day-old Ross 308 broiler chicks were randomly allocated to 2 dietary treatments (i.e., Tercie vs. Pexeso), with 6 replicate pens and 15 chicks per pen. Pexeso wheat, characterized by increased lutein and zeaxanthin concentrations, in combination with rapeseed oil as the primary dietary fat source, significantly improved the feed conversion ratio (FCR; p < 0.001), despite not affecting the body weight of the chickens at 35 days of age or feed intake. This improved efficiency was consistent with the significantly increased retention of crude protein (p = 0.004). Specifically, the concentrations of γ-tocopherol (p = 0.006) and lutein (p…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
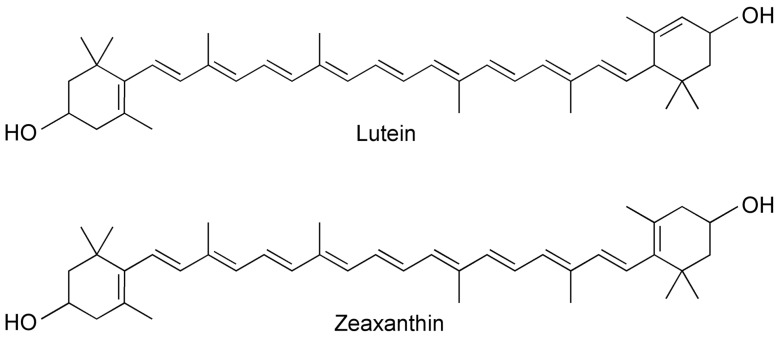 Figure 1
Figure 1- —Ministry of Agriculture of the Czech Republic
- —METROFOOD-CZ Research Infrastructure Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Nutrition and Physiology · Antioxidant Activity and Oxidative Stress · Meat and Animal Product Quality
1. Introduction
As the projected global population is expected to reach 9–10 billion by 2050, the global demand for food, particularly protein, is predicted to rise dramatically in the following decades [1,2]. In terms of both absolute and relative growth rates, poultry meat production has surpassed that of other major meat types worldwide over the past 50 years [2]. Consequently, the increasing global demand for broiler chicken meat, driven by the increasing need for high-quality animal protein [3], is compelling poultry producers to enhance bird health, growth performance, feed efficiency, and, especially, product quality [4]. At the same time, the intensification of poultry production is associated with increased environmental pressures, highlighting the need for more sustainable and resource-efficient feeding strategies. In this context, the use of feeds that meet multiple animal nutritional requirements is promising because it can reduce the need to produce additional ingredients (such as lipophilic vitamins and carotenoid pigments) and can reduce the environmental footprint of animal feed production [5]. Specifically, carotenoid-biofortified wheat has the potential to meet the energy requirements of poultry while also enhancing the antioxidant profile and thereby improving the quality of the produced poultry meat.
Carotenoids, best known for their wide range of bioactive and health-promoting properties, represent one of the most widespread groups of natural lipophilic pigments. These pigments are responsible for a broad spectrum of colors and have been identified in a variety of organisms, including plants, microalgae, bacteria, yeasts, molds, and fungi, in which they are synthesized de novo [6,7,8]. Carotenoids are categorized into two primary groups on the basis of their chemical structure, particularly the presence or absence of oxygen in the molecule, and their functional properties. The two groups include carotenes (i.e., oxygen-free carotenoids), such as α-carotene, β-carotene, or lycopene, and xanthophylls (i.e., carotenoids containing at least one oxygen atom in the molecule), such as lutein and zeaxanthin (Figure 1) [6,9].
Carotenoid pigments can be further categorized according to their provenance, distinguishing between naturally occurring carotenoids and synthetically produced carotenoids. Animals and birds are incapable of synthesizing carotenoids in vivo; thus, these pigments must be supplied through the diet [11]. The supplementation of poultry feed mixtures with carotenoids is applicable across all poultry categories; however, the reasons differ partly, as carotenoids serve primarily for meat and skin pigmentation in broiler chickens and for achieving optimal yolk coloration in laying hens [6]. Moreover, with the increasing number of poultry product consumers and their growing concerns about the use of synthetically produced additives, including carotenoids, in animal nutrition and welfare [12], a rising trend has emerged toward the incorporation of natural carotenoid sources, particularly xanthophylls. In this context, natural carotenoids have recently attracted considerable attention in animal nutrition because of their rich bioactive and health-promoting properties, including antioxidant [13,14], anti-inflammatory [15,16], and immunomodulatory [17,18] effects. Moreover, their growing use in broiler chicken diets as alternatives to synthetic pigments is motivated not only by concerns related to safety and bioavailability [19] but also by increasing evidence supporting their potential to improve poultry health and performance [7]. For example, grazing broiler chickens obtain natural xanthophylls directly from pasture vegetation [20]. Additionally, increased carotenoid concentrations in both feed mixtures and poultry products have been reported following the inclusion of various natural ingredients in poultry diets, such as tomato powder [21], colored carrot [22], dried common nettle [23], marigold flower extract [24], alfalfa concentrate [25], spray-dried Chlorella vulgaris [26], dried calendula flowers [12], or dried stevia leaves [27].
In addition to various natural sources, carotenoid-biofortified cereal grains represent an innovative way to increase the dietary carotenoid supply and, consequently, the carotenoid content of poultry products. Biofortification refers to strategies aimed at enhancing the concentration of essential nutrients in staple crops through agronomic practices or genetic improvement [28]. Examples of successful biofortification include golden rice, which produces high levels of β-carotene [29]; transgenic multivitamin maize, which is capable of simultaneously accumulating elevated levels of β-carotene, lycopene, lutein, zeaxanthin, folate, and ascorbic acid [30]; provitamin A carotenoid-biofortified maize, which produces increased levels of β-cryptoxanthin [31]; and micronutrient-biofortified wheat, which is enriched in zinc, iodine, selenium, and iron [32]. Consistent with these approaches, the present study evaluated a newly developed carotenoid-biofortified spring wheat variety, Pexeso, in which the breeding process resulted in increased lutein and zeaxanthin concentrations. Such crops not only have the potential to improve human health directly in regions predominantly reliant on cereals for nutrition [33,34] but can also enhance animal health when included in feed [35] and may subsequently pass these benefits on to humans consuming meat and other products derived from these animals [36]. However, most biofortification research has focused on nutrient accumulation in planta rather than on the fate of these nutrients after consumption, although bioaccessibility and bioavailability provide more accurate indicators of nutritional quality than nutrient content alone does [37,38]. Therefore, the present study aimed to investigate the dietary effects of the new carotenoid-biofortified wheat variety, Pexeso, with elevated carotenoid concentrations, compared with those of the common wheat variety, Tercie, with standard carotenoid levels, on performance characteristics, nutrient retention, and tissue antioxidant concentrations in broiler chickens.
2. Materials and Methods
All the experimental procedures in this study complied with the guidelines of the Ethics Committee of the Central Commission for Animal Welfare at the Ministry of Agriculture of the Czech Republic (Prague, Czech Republic) and with Directive 2010/63/EU of the European Parliament and the Council on the protection of animals used for scientific purposes. The research protocol (code 06/2023) was reviewed and approved by the Ethical Committee of the Institute of Animal Science, v.v.i. (Prague-Uhříněves, Czech Republic).
2.1. Experimental Design, Management of Broiler Chickens, and Dietary Treatments
This experiment was performed with a total of 180 one-day-old Ross 308 broiler chicks that were randomly allocated to 2 dietary treatments (i.e., groups), each with 6 replicate pens and 15 chicks per pen. The experimental design and allocation of broiler chickens to dietary treatments are presented in Table 1. The dietary treatments differed by the wheat variety used, namely, Tercie and Pexeso. Specifically, a new carotenoid-biofortified spring wheat variety, Pexeso, with increased lutein and zeaxanthin concentrations, was compared with the common wheat variety Tercie. Pexeso wheat was biofortified through a breeding process (variety code: 5095202; year of registration: 2018) and contained 1.115 mg/kg lutein and 0.755 mg/kg zeaxanthin. By contrast, the lutein and zeaxanthin concentrations in Tercie wheat were 0.439 mg/kg and 0.214 mg/kg, respectively. Both wheat varieties were bred in the Czech Republic by the company Plant Breeding Station Stupice, Selgen, a.s. (Sibřina—Stupice, Czech Republic). The ingredients (Table 2) and chemical compositions (Table 3 and Table 4) of the experimental broiler chicken diets and wheat varieties are presented in the following tables. The dietary treatments used (i.e., diet with Tercie wheat and diet with Pexeso wheat) were designed to provide similar levels of metabolizable energy (~11.95 MJ/kg) and crude protein (~221 g/kg). A balanced metabolizable energy concentration in the experimental broiler chicken diets was achieved by adjusting the amount of added rapeseed oil (30 g/kg in both diets). The use of rapeseed oil as the primary dietary fat source was based on findings from our previous study [39], which highlighted its favorable nutritional effects in poultry diets. The experimental broiler chicken diets were pelleted. Only a single type of feed mixture was used throughout the experimental period to avoid potential confounding effects associated with diet changes during different growth phases. Both experimental broiler chicken diets, differing only in the wheat variety used (i.e., Tercie or Pexeso), most closely corresponded to a BR1-type feed mixture based on their formulation and nutrient composition. Broiler chickens had ad libitum access to pelleted feed and fresh water that complied with high hygiene standards.
The broiler chickens were housed in floor pens with wood shavings used as litter material. The stocking density, pen equipment (i.e., pan feeders, nipple drinkers, and gas heaters), lighting, ventilation, inspection frequency, and all related management practices complied with the requirements of the European Council Directive 2007/43/EC [40]. The technological and microclimatic housing conditions were maintained according to the standard management guidelines for the Ross 308 hybrid. The housing room was heated by a gas heating system, and ventilation was provided by a temperature-controlled fan. During the first 3 days after hatching, the room temperature was maintained at 30–32 °C and then gradually decreased to 20–22 °C by 28 days of age. The relative humidity was maintained between 50% and 60%. The light cycle during the first week (i.e., 1–7 days of age) consisted of 23 h of light and 1 h of darkness. From 8 to 35 days of age, the photoperiod was reduced to 16 h of light and 8 h of darkness per day. The experiment was conducted in full compliance with the highest animal welfare standards. Throughout the experiment, the broiler chickens were not subjected to any form of restraint or distress. Their health status was assessed twice daily by evaluating general activity, normal behavioral patterns (e.g., active feed and water intake, walking, wing stretching, alert responses, and calm respiration), vocalization, plumage condition, skin integrity, posture, and foot and limb formation.
2.2. Performance Characteristics and Nutrient Retention in the Broiler Chickens
To determine the performance characteristics, the broiler chickens were weighed (g) at 0, 14, 28, and 35 days of age, and the feed intake (g/chicken/day) was monitored daily. The feed conversion ratio (FCR; kg/kg) was calculated as the total feed intake divided by the overall body weight gain during the experimental period (i.e., 35 days). At 28 days of age, 6 broiler chickens (one per replicate pen; n = 6) with an average body weight (i.e., chickens with a weight closest to the average weight per pen) were selected from each dietary treatment (i.e., group) and individually housed in cages to determine the retention (%) of crude protein, crude fat, tocopherols (i.e., α-tocopherol and γ-tocopherol), and carotenoids (i.e., lutein and zeaxanthin). During the balance period, daily feed intake (g/chicken/day) and feces output (g/chicken/day) were recorded for 7 days (including a 5-day adaptation period and a 2-day feces collection period). The collected feces were lyophilized and stored for further chemical analyses. The retention of the selected nutrients was calculated on the basis of the total tract apparent digestibility coefficient (ADC; %) using the indicator method. As an indicator for determining digestibility, 5 g/kg of celite (i.e., diatomaceous earth) was added to the experimental broiler chicken diets, and the acid-insoluble ash in the diets was subsequently measured using 3 M hydrochloric acid (HCl). The formula for calculating the total tract apparent digestibility coefficient (ADC) was as follows:
where I_D_ represents the content of the indicator in dry matter (DM; %) of the diet; N_F_ represents the content of the nutrient in DM (%) of the feces; I_F_ represents the content of the indicator in DM (%) of the feces; and N_D_ represents the content of the nutrient in DM (%) of the diet.
At 35 days of age, 1 broiler chicken with an average body weight was selected from each replicate pen (n = 6) and humanely slaughtered in a certified commercial slaughterhouse. The broiler chickens were slaughtered by exsanguination following prior electrical stunning, without analgesia or anesthesia. After slaughter, the carcasses were chilled for 24 h, after which the breast muscle (pectoralis major) and liver were excised for subsequent laboratory analyses.
2.3. Laboratory Analyses of the Experimental Diets, Wheat Varieties, Breast Meat, and Livers
Nutrient analyses of the experimental broiler chicken diets, wheat varieties (i.e., Tercie and Pexeso), and feces output were carried out in accordance with the official methods of the Association of Official Analytical Chemists [41]. The dry matter content (g/kg) of the diets, wheat varieties, and feces was established by heating the sample in an oven at 105 °C to constant weight (Memmert ULM 500; Memmert, Schwabach, Germany). The crude protein content (g/kg) of the diets, wheat varieties, and feces was measured with a Kjeltec Auto 1030 Analyzer (Tecator, Högänas, Sweden). The crude fat content (g/kg) of the diets, wheat varieties, and feces was analyzed using a Soxtec 1045 Extraction Unit (Tecator, Högänas, Sweden) with petroleum ether as the extraction solvent. The crude fiber content (g/kg) of the diets, wheat varieties, and feces was determined by acid–alkaline hydrolysis using a Fibertec 2010 System (FOSS, Hillerød, Denmark). The dried and homogenized samples of the diets and wheat varieties were incinerated at 550 °C in a muffle furnace (LMH 11/12 with Ht40 AL temperature controller; LAC Asia Limited, Hong Kong, China) to obtain the mineral ash. The ash was then dissolved in 3 M HCl for subsequent analysis. The calcium (Ca) concentration (g/kg) in the resulting solution was quantified by atomic absorption spectrometry with a Solar M6 instrument (TJA Solutions, Cambridge, UK). The total phosphorus (P) concentration (g/kg) was assessed in the HCl extract employing the vanadate-molybdate reagent (method No. 965.17) [41].
The concentrations of α-tocopherol and γ-tocopherol (mg/kg) in the experimental broiler chicken diets, wheat varieties (i.e., Tercie and Pexeso), breast meat (i.e., muscle; pectoralis major), livers, and feces output (n = 6) were determined with a Shimadzu high-performance liquid chromatography (HPLC) system (VP series; Shimadzu, Kyoto, Japan) equipped with a diode array detector (HPLC-DAD). Prior to analysis, the samples were subjected to alkaline saponification with 60% aqueous potassium hydroxide (KOH) and subsequently extracted with diethyl ether, in accordance with the EN 12822-1 [42] European standard. Chromatographic separation was achieved on a Phenomenex Synergi 4 μm Fusion-RP 80 Å column (150 × 4.6 mm, 4 μm; P/No. 00F-4424-E0, Torrance, CA, USA) using methanol as the mobile phase. A gradient flow program was applied, where the solvent composition remained constant while the flow rate was varied from 0.6 mL/min to 1.5 mL/min and returned to 0.6 mL/min. Sample injection volume was 50 μL. Quantification was conducted using individual calibration curves for each analyte. The detection wavelengths (λ) were set as 292 nm for α-tocopherol and 296 nm for γ-tocopherol. The concentrations of tocopherols in the experimental broiler chicken diets, wheat varieties, breast meat, and livers were expressed as mg/kg fresh weight.
The concentrations of lutein and zeaxanthin (mg/kg) in the experimental broiler chicken diets, wheat varieties (i.e., Tercie and Pexeso), breast meat (i.e., muscle; pectoralis major), livers, and feces output (n = 6) were determined by a modified method described by Froescheis et al. [43]. Chromatographic analyses were performed with a Phenomenex C18 Kinetex column (100 × 4.6 mm, 2.6 µm; P/No. 00D-4462-E0, Torrance, CA, USA) coupled with a Shimadzu HPLC system (VP series; Shimadzu, Kyoto, Japan) and a diode array detector (HPLC-DAD). The mobile phase consisted of solvent A (acetonitrile-water-ethyl acetate, 88:10:2) and solvent B (acetonitrile-water-ethyl acetate, 88:0:15), and a gradient elution program starting from 100% A/0% B at 0.01 min with a flow rate of 0.8 mL/min was used. The flow rate was increased to 2.0 mL/min at 3.01 min. The composition was changed to 0% A/100% B at 6.00 min and maintained until 6.01 min, when the flow rate was returned to 0.8 mL/min. At 11.00 min, the initial composition (i.e., 100% A/0% B) was restored for column re-equilibration. The total run time was 18 min. The detection of both lutein and zeaxanthin was performed at λ = 450 nm. The concentrations of carotenoids in the experimental broiler chicken diets, wheat varieties, breast meat, and livers were expressed as mg/kg fresh weight.
2.4. Statistical Analysis
The experimental data obtained were analyzed with the general linear model (GLM) procedure in the Statistical Analysis System [44]. Differences between the dietary treatments (i.e., groups) were tested with Student’s t-test. The experimental unit was a pen (n = 6). The results in the tables are presented as the means (^a,b^), the standard error of the mean (SEM), and the probability (p) values. All differences were considered significant at p < 0.05.
3. Results
3.1. Compositional Analysis
All the results of the experimental data evaluated are summarized and presented in the following tables. The ingredient compositions of the experimental broiler chicken diets are shown in Table 2. As demonstrated in Table 3, the use of two distinct wheat varieties (i.e., Tercie and Pexeso), characterized by different concentrations of lutein and zeaxanthin, resulted in varying levels of these carotenoids within the experimental broiler chicken diets (0.84 mg/kg lutein and 0.58 mg/kg zeaxanthin in the Tercie wheat diet and 1.22 mg/kg lutein and 0.77 mg/kg zeaxanthin in the Pexeso wheat diet). In the carotenoid-biofortified Pexeso wheat, the concentrations of lutein and zeaxanthin were approximately 2.5- and 3.5-fold higher, respectively, than those found in the common Tercie wheat (Table 4). This increase was reflected in an almost 1.5-fold increase in the concentrations of these two xanthophylls in the experimental broiler chicken diet containing Pexeso wheat. Moreover, compared with common Tercie wheat, the new carotenoid-biofortified Pexeso wheat also presented higher contents of crude fat, ash, Ca, P, α-tocopherol, and γ-tocopherol.
3.2. Performance Characteristics of the Broiler Chickens
The performance characteristics of the broiler chickens are presented in Table 5. The experimental broiler chicken diet containing carotenoid-biofortified Pexeso wheat did not affect the body weight of the chickens at 35 days of age or feed intake, but it significantly improved the FCR (p < 0.001). Broiler chickens fed the Pexeso wheat diet showed a lower FCR (1.40 kg/kg) than those fed the Tercie wheat diet (1.45 kg/kg). No significant differences in mortality rates were detected between the dietary treatments, indicating that the inclusion of the newly bred wheat variety Pexeso had no detrimental effect on bird viability.
3.3. Retention of the Selected Nutrients in the Broiler Chickens
As shown in Table 6, the dietary treatment with Pexeso wheat significantly increased crude protein (p = 0.004) and γ-tocopherol (p = 0.008) retention in the broiler chickens. The improvement in crude protein retention corresponded with the reduced FCR (Table 5), indicating more efficient utilization of dietary nutrients for tissue accretion. The experiment demonstrated that the dietary xanthophylls, lutein and zeaxanthin, were not fully metabolized by the broiler chickens but were largely excreted in the feces. Nevertheless, feeding broiler chickens with Pexeso wheat, which is rich in carotenoids, significantly increased the retention of both lutein (p < 0.001) and zeaxanthin (p < 0.001). Among all the evaluated bioactive compounds (i.e., tocopherols and carotenoids), γ-tocopherol exhibited the highest retention (84.69%), particularly in broiler chickens fed the Pexeso wheat diet. No significant effect of the dietary treatment with Pexeso wheat was observed on the retention of crude fat or α-tocopherol.
3.4. Tissue Antioxidant Concentrations in the Broiler Chickens
The tissue (i.e., breast meat and liver) antioxidant (i.e., tocopherols and carotenoids) concentrations in the broiler chickens are reported in Table 7. Compared with the experimental diet containing Tercie wheat, the experimental diet containing Pexeso wheat significantly increased γ-tocopherol (p = 0.006) and lutein (p = 0.004) concentrations in the breast meat and γ-tocopherol (p = 0.047), lutein (p < 0.001), and zeaxanthin (p < 0.001) concentrations in the liver. This accumulation was supported by the significantly greater retention of these antioxidants observed in broiler chickens receiving the experimental diet with carotenoid-biofortified Pexeso wheat (Table 6). The concentrations of α-tocopherol in both the breast meat and liver, however, remained unaffected.
4. Discussion
Poultry meat is among the most popular sources of animal protein worldwide, offering high nutritional value and bioactive compounds that are important for cell regeneration and the maintenance of human health [45,46,47]. Thanks to its rich nutritional composition, poultry meat serves as an excellent medium for supplying additional health-promoting bioactive compounds, the levels of which can be substantially influenced by the diet of broiler chickens. Meat enriched with essential nutrients, such as tocopherols and carotenoids, is regarded as a designed product [46]. If such enrichment contributes to increasing the daily intake of these nutrients and, consequently, exerts beneficial effects on human health, the product meets the criteria for being labeled as functional food [48]. However, the benefits of foods containing essential nutrients depend not only on their nutritional content but also on how they are delivered (e.g., as supplements, additives, or as parts of the food matrix), as this can affect their accessibility, solubility, and potential absorption in the gastrointestinal tract [36]. In the present study, the new carotenoid-biofortified spring wheat variety Pexeso was evaluated as a feed ingredient in broiler chicken diets and compared with the common wheat variety Tercie.
As shown in Table 4, the lutein and zeaxanthin concentrations were approximately 2.5- and 3.5-fold higher, respectively, in Pexeso wheat (1.115 mg/kg and 0.755 mg/kg, respectively) than in Tercie wheat (0.439 mg/kg and 0.214 mg/kg, respectively). Similar effects of biofortification have been observed in transgenic multivitamin maize, where Naqvi et al. [30] reported that lutein levels increased by 30%, from 8.74 mg/kg to 11.36 mg/kg, and that zeaxanthin levels increased by up to 300%, from 2.05 mg/kg to 8.19 mg/kg, with initial concentrations already far exceeding those in Pexeso wheat. However, comparing the carotenoid content of experimental broiler chicken diets with that of other published studies is challenging, as the present work focuses uniquely on the use of newly developed carotenoid-biofortified wheat. Carotenoids, owing to the system of conjugated double bonds between individual carbon atoms in the core of the polyene chain structure (Figure 1), can effectively quench singlet oxygen (^1^O_2_) and act as scavengers of highly reactive oxygen species (ROS), i.e., as antioxidants [49]. Among all carotenoids, including α-carotene, β-carotene, lutein, and zeaxanthin, lycopene has been identified as one of the most potent antioxidants for protecting cells against ROS-induced damage [14]. Furthermore, carotenoids and several plant-derived compounds have excellent therapeutic and health-promoting potential for enhancing the production performance of poultry [7]. Although, in the present study, the experimental broiler chicken diet containing carotenoid-biofortified Pexeso wheat did not affect the body weight of the chickens at 35 days of age or feed intake, it significantly improved the FCR compared with the experimental broiler chicken diet containing common Tercie wheat (Table 5). These results are in line with those of Díaz-Gómez et al. [50], who reported that compared with a commercial diet supplemented with synthetic color additives, a diet supplemented with carotenoid-biofortified maize, which accumulates higher β-carotene, β-cryptoxanthin, lutein, and zeaxanthin concentrations, resulted in similar pigmentation and growth performance of broiler chickens. The beneficial effects of carotenoids on performance characteristics have also been reported by Sun et al. [51], Mezbani et al. [52], and Fathi et al. [53], who tested different levels of lycopene in broiler chicken diets. However, it should be noted that the present study was designed to investigate the effects of a single, practically relevant inclusion level of carotenoid-biofortified Pexeso wheat; therefore, dose–response relationships cannot be inferred from the current experimental data. In agreement with previous reports, Ševčíková et al. [54] reported that dietary supplementation with lycopene (100 mg/kg) increased the final live weight of broiler chickens. Furthermore, astaxanthin, a xanthophyll pigment obtained from Phaffia rhodozyma [55] or produced as a powder from Haematococcus pluvialis [56], has been shown to improve FCR, total weight gain, and final body weight, but it also significantly reduces total mortality. In the present study, however, this health indicator, mortality, was not influenced by the inclusion of Pexeso wheat in the experimental broiler chicken diet (Table 5). In addition to the antioxidant and performance-enhancing effects mentioned above, carotenoids exhibit anti-inflammatory properties [15,16] and modulate immune responses [17,18] by boosting immune activity and inducing lymphocyte proliferation [57], which may further contribute to the improved overall performance of broiler chickens. For example, Nogareda et al. [35] reported that feeding broiler chickens carotenoid-biofortified maize, which contains high levels of β-carotene, lycopene, lutein, and zeaxanthin, enhanced protective immunity against infectious bursal disease, further supporting the immunomodulatory potential of dietary carotenoids. Moreover, a link between specific carotenoids, oxidative stress, and the immune system has previously been described in birds [58], highlighting the integrative role of these bioactive compounds in modulating both antioxidant defenses and immune responses.
Dietary carotenoids are absorbed by birds, as in other animals, through the intestinal mucosa, where they are incorporated into lipoprotein particles and transported via the blood to peripheral organs and target tissues [59]. The efficiency of this process is shaped by several digestive steps that determine how effectively carotenoids are released from the feed matrix, incorporated into mixed micelles, and delivered to enterocytes [60,61,62]. Carotenoid polarity, dietary fatty acid (FA) composition, and competition among different carotenoids can markedly influence micellarization and uptake [62,63,64]. Once inside enterocytes, carotenoids enter circulation via chylomicrons (i.e., portomicrons), with different classes of carotenoids preferentially associated with distinct lipoproteins and subsequently delivered to specific tissues [36,62,65]. In general, serum carotenoids represent the mobile pool of pigments available for delivery to peripheral tissues, whereas the liver serves as the main storage and metabolic hub, redistributing carotenoids and their metabolites through high-density lipoprotein (HDL), low-density lipoprotein (LDL), and very low-density lipoprotein (VLDL) fractions [17,66,67]. These physiological constraints and selective transport pathways are crucial for understanding the variation in carotenoid bioavailability in poultry.
Our experiment demonstrated that the dietary xanthophylls, lutein and zeaxanthin, were not fully metabolized by the broiler chickens but were largely excreted in the feces. Quantitative data on the absorption and retention of these carotenoids in poultry are lacking in the literature. For example, Reboul [68] reported substantial variability in the absorption of xanthophylls. If the storage tissues (i.e., liver) or the transport routes leading to them become saturated, any carotenoids not utilized directly in peripheral tissues are cleared from the body and excreted in the bird’s feces [36], as also observed in humans [69]. In the present study, however, feeding broiler chickens with the experimental diet containing Pexeso wheat significantly increased γ-tocopherol (0.375 mg/kg) and lutein (0.082 mg/kg) concentrations in the breast meat as well as γ-tocopherol (1.22 mg/kg), lutein (0.397 mg/kg), and zeaxanthin (0.241 mg/kg) concentrations in the liver, compared with the experimental diet containing Tercie wheat (0.306 mg/kg and 0.051 mg/kg in the breast meat and 0.89 mg/kg, 0.197 mg/kg, and 0.092 mg/kg in the liver, respectively; Table 7). Moreover, this accumulation was supported by the significantly higher retention of these antioxidants observed in broiler chickens receiving a diet with carotenoid-biofortified Pexeso wheat (Table 6). Similarly, in a study by Nogareda et al. [35], an analysis of carotenoid levels in breast meat revealed that β-carotene, lutein, and zeaxanthin levels were significantly greater in the breast meat of broiler chickens fed a carotenoid-biofortified maize diet than in that of the control group. In addition, these birds, reared on the high-carotenoid diet, accumulated the highest levels of liver retinol (i.e., vitamin A; 814 µg/g) compared with the control group (471 µg/g) and with commercial diets with and without color additives (573 µg/g and 531 µg/g, respectively), potentially reflecting the greater supply of β-carotene and β-cryptoxanthin in the carotenoid-biofortified maize [35]. These natural carotenoids, together with α-carotene, are principally considered precursors of vitamin A [7], an essential component of the visual pigment rhodopsin, which also plays a role in maintaining epithelial and immune cells [18]. Vitamin A and carotenoid metabolism is similar in broiler chickens and humans [70]. However, broiler chickens convert provitamin A carotenoids into retinol more efficiently via the action of β-carotene 15,15’-monooxygenase [50], making them suitable animal models for studying the impact of carotenoid-enhanced diets in humans [36]. Furthermore, van het Hof et al. [71] reported that lutein is five times more bioavailable than β-carotene is in humans. Although the Pexeso wheat used in our study was enriched solely with carotenoids lacking provitamin A activity (i.e., lutein and zeaxanthin), and retinol content in tissues was therefore not measured, it is worth noting that Nogareda et al. [35] reported that the increased level of retinol in the liver, the primary site of retinol storage, appears to be one of the key benefits of a carotenoid-biofortified maize diet and is likely directly linked to the improved protective immunity observed in challenge studies. Moreover, Nogareda et al. [35] reported that high levels of β-carotene and zeaxanthin oxidation products were present in the blood, breast, thigh, and shank skin and fat. These results indicate that the carotenoids were actively involved in antioxidant defense in these tissues, protecting the broiler chickens from oxidative stress.
Among all the evaluated bioactive compounds (i.e., tocopherols and carotenoids), γ-tocopherol exhibited the highest retention (84.69%), particularly in broiler chickens fed the Pexeso wheat diet (Table 6). This enhanced retention was reflected in the elevated γ-tocopherol levels in both the breast meat (0.375 mg/kg) and liver (1.22 mg/kg), as determined by HPLC-DAD analysis, compared with those in the Tercie wheat diet (0.306 mg/kg and 0.89 mg/kg, respectively; Table 7). The main factor affecting the tocopherol (i.e., vitamin E isomers) content of poultry products is nutrition, followed by age and animal variety or genotype [72]. Tocopherols in poultry diets originate exclusively from dietary ingredients, primarily from dietary fat sources such as rapeseed oil [73]. Vegetable oils, particularly rapeseed oil, contain substantial amounts of α-linolenic acid (ALA, C18:3, n-3), which acts as a precursor for the synthesis of its long-chain derivatives, eicosapentaenoic acid (EPA, C20:5, n-3) and docosahexaenoic acid (DHA, C22:6, n-3), in broiler chicken meat [74,75]. These bioactive forms of essential FAs are necessary for many physiological processes [76]. In addition, rapeseed oil is characterized by a high content of bioactive compounds, such as tocopherols, phytosterols, and phenolic compounds [77]. Although both experimental broiler chicken diets used in our study contained an identical amount of rapeseed oil (30 g/kg; Table 2), the markedly higher γ-tocopherol concentrations observed in the tissues of the broiler chickens fed the Pexeso wheat diet were most likely attributable to the carotenoid-biofortified Pexeso wheat itself, in which the γ-tocopherol content also increased through the biofortification process (Table 4). These findings may be further supported by the fact that supplementation of broiler chicken diets with fats rich in unsaturated fatty acids (UFAs), such as rapeseed oil, has been shown to increase vitamin E bioavailability compared with supplementation with fats rich in saturated fatty acids (SFAs) [78]. In contrast, compared with fats high in monounsaturated fatty acids (MUFAs) and polyunsaturated fatty acids (PUFAs), dietary fats rich in SFAs have been reported to increase the bioavailability of lutein and zeaxanthin [39,79]. Moreover, MUFAs improve the absorption of polar carotenoids (i.e., lutein and zeaxanthin), particularly when associated with a high metabolizable energy density (11.4 MJ/kg) [80]. This finding may be consistent with our findings, as the experimental broiler chicken diet containing Pexeso wheat had a metabolizable energy content of 11.9 MJ/kg, potentially further supporting carotenoid absorption. Tocopherols are recognized for their potent antioxidant activity, which can enhance the antioxidant potential and, consequently, the oxidative stability of poultry products [81]. Among all tocopherol isomers, γ-tocopherol is considered the most active antioxidant in lipophilic matrices [82], whereas α-tocopherol is regarded as the only tocopherol isomer with vitamin E activity in the human body [83]. As summarized in detail by Szmek et al. [81], however, the bioactive properties of tocopherol isomers depend not only on dietary concentrations but also on how they are absorbed and utilized by the organism. Besides its potent antioxidant effects, γ-tocopherol exhibits anti-inflammatory activity, notably by inhibiting prostaglandin-endoperoxide synthase (i.e., cyclooxygenase) activity [84,85,86]. The increased γ-tocopherol concentrations observed in the tissues of broiler chickens fed the Pexeso wheat diet may be partly related to the antioxidant properties of naturally occurring carotenoids, which protect this lipophilic compound against oxidative degradation in the upper gastrointestinal tract [87]. Conversely, carotenoids may also compete with γ-tocopherol for incorporation into micelles or for uptake by putative transporters, potentially impairing its absorption [88]. Moreover, the specific polyene chain structure of xanthophylls (Figure 1) confers lipophilic properties, enabling these pigments, together with tocopherols, to slow lipid peroxidation and stabilize lipid–protein structures, such as cell membranes or lipoproteins present in broiler chicken meat [89]. The synergistic effect of lycopene (75 mg/kg) and vitamin E (50 mg/kg or 100 mg/kg) has been demonstrated by Englmaierová et al. [90], who reported that the combined addition of these antioxidants improved both the growth performance and oxidative stability of fresh leg meat as well as that of meat stored for 3 days.
In light of the aforementioned findings, it can be concluded that dietary supplementation with natural xanthophylls, such as lutein and zeaxanthin provided by carotenoid-biofortified Pexeso wheat, has the potential not only to support bird health and production performance but also to enhance the nutritional quality and oxidative stability of poultry meat.
5. Conclusions
The findings of this study indicate that carotenoid-biofortified Pexeso wheat represents a promising functional feed ingredient, particularly because of its beneficial effects on the performance characteristics of broiler chickens and its enriching influence on the nutritional quality of poultry meat. Specifically, the inclusion of Pexeso wheat, characterized by increased lutein and zeaxanthin concentrations, in combination with rapeseed oil as the primary dietary fat source, significantly improved the FCR and enhanced the antioxidant profile of broiler chicken tissues (i.e., breast meat and liver). This effect was primarily associated with greater retention and tissue accumulation of bioactive antioxidants, especially naturally occurring xanthophylls and γ-tocopherol. Consequently, carotenoid-biofortified Pexeso wheat may be considered a viable functional alternative to conventional wheat varieties in broiler chicken diets under commercial production conditions. In addition, its inclusion in broiler chicken diets may contribute to increased deposition of bioactive compounds with potential health-promoting benefits for human consumers. Further research should investigate the long-term effects of carotenoid-biofortified cereal grains, such as Pexeso wheat, on poultry performance, meat nutritional quality, and antioxidant stability, as well as the potential health-promoting effects of enriched poultry products (i.e., functional foods) in humans.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Van Dijk M. Morley T. Rau M.L. Saghai Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050 Nat. Food 2021249450110.1038/s 43016-021-00322-937117684 · doi ↗ · pubmed ↗
- 2Ullah H. Feed Sustainability and Efficiency Poultry Farming—New Perspectives and Applications Téllez-Isaías G. Intech Open London, UK 202310.5772/intechopen.110332 · doi ↗
- 3Devi P.C. Samanta A.K. Das B. Kalita G. Behera P.S. Barman S. Effect of Plant Extracts and Essential Oil Blend as Alternatives to Antibiotic Growth Promoters on Growth Performance, Nutrient Utilization and Carcass Characteristics of Broiler Chicken Indian J. Anim. Nutr.20183542110.5958/2231-6744.2018.00064.6 · doi ↗
- 4Belhadj Slimen I. Val Boa M. Lengliz S. Bejaoui B. Samti H. Abbassi M.S. M’Hamdi N. Chaveriat L. Martin P. Abderrabba M. Effects of Carvacrol on Growth Performance, Carcass Traits, Serum Biochemistry, and Intestinal Gut Microflora of Broiler Chickens Front. Anim. Sci.20256156391510.3389/fanim.2025.1563915 · doi ↗
- 5Kljak K. Duvnjak M. BedekovićD. KišG. JanječićZ. Grbeša D. Commercial Corn Hybrids as a Single Source of Dietary Carotenoids: Effect on Egg Yolk Carotenoid Profile and Pigmentation Sustainability 2021131228710.3390/su 132112287 · doi ↗
- 6Marounek M. Pebriansyah A. Use of carotenoids in feed mixtures for poultry: A review Agric. Trop. Subtrop.20185110711110.2478/ats-2018-0011 · doi ↗
- 7Nabi F. Arain M.A. Rajput N. Alagawany M. Soomro J. Umer M. Soomro F. Wang Z. Ye R. Liu J. Health benefits of carotenoids and potential application in poultry industry: A review J. Anim. Physiol. Anim. Nutr.20201041809181810.1111/jpn.1337532333620 · doi ↗ · pubmed ↗
- 8Sun T. Rao S. Zhou X. Li L. Plant carotenoids: Recent advances and future perspectives Mol. Hortic.20222310.1186/s 43897-022-00023-237789426 PMC 10515021 · doi ↗ · pubmed ↗