DNA Barcoding for Herbarium Specimens of the Red Alga Meristotheca pilulaora and Molecular Marker Development for Species Identification
Soon Jeong Lee, Eun-Young Lee, Bo Yeon Kim, Sang-Rae Lee

TL;DR
This study used DNA barcoding to correct misidentifications in algal herbarium specimens and developed a molecular marker to distinguish between two red algal species in Korea.
Contribution
The study developed a cox1 molecular marker for accurate species identification and revealed inter-ordinal misidentifications in herbarium specimens.
Findings
Six out of twelve M. papulosa specimens were misidentified as M. pilulaora or G. textorii.
A cox1 marker was developed to effectively distinguish between M. pilulaora and G. textorii.
DNA barcoding provided insights into the geographic distribution of M. pilulaora in Korea.
Abstract
The red alga Meristotheca pilulaora (Gigartinales, Solieriaceae) has recently been described and newly recorded from Korea based on analyses of a species previously reported as Meristotheca papulosa. We re-examined M. papulosa herbarium specimens deposited in the National Institute of Biological Resources algal herbarium (Korea) to clarify the taxonomic relationship between M. papulosa and M. pilulaora. Using DNA barcoding, we examined M. papulosa specimens and unexpectedly identified Meristotheca pilulaora and Gracilaria textorii (Gracilariales, Gracilariaceae), but not M. papulosa. Thus, we revealed a misidentification at the inter-ordinal level (6/12 specimens). Moreover, we developed a cox1 marker as an effective molecular tool to discriminate between M. pilulaora and G. textorii. The molecular taxonomic results obtained in this study could contribute to gaining useful genetic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1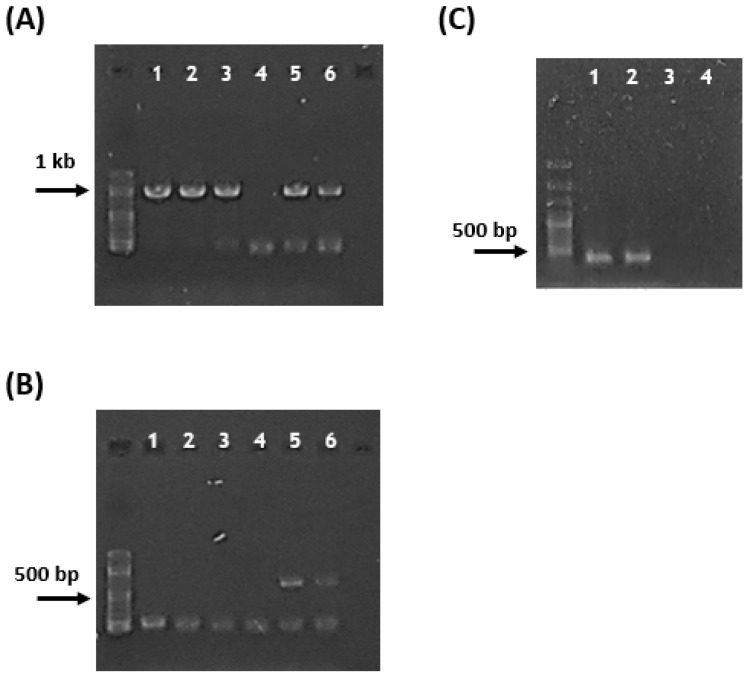 Figure 2
Figure 2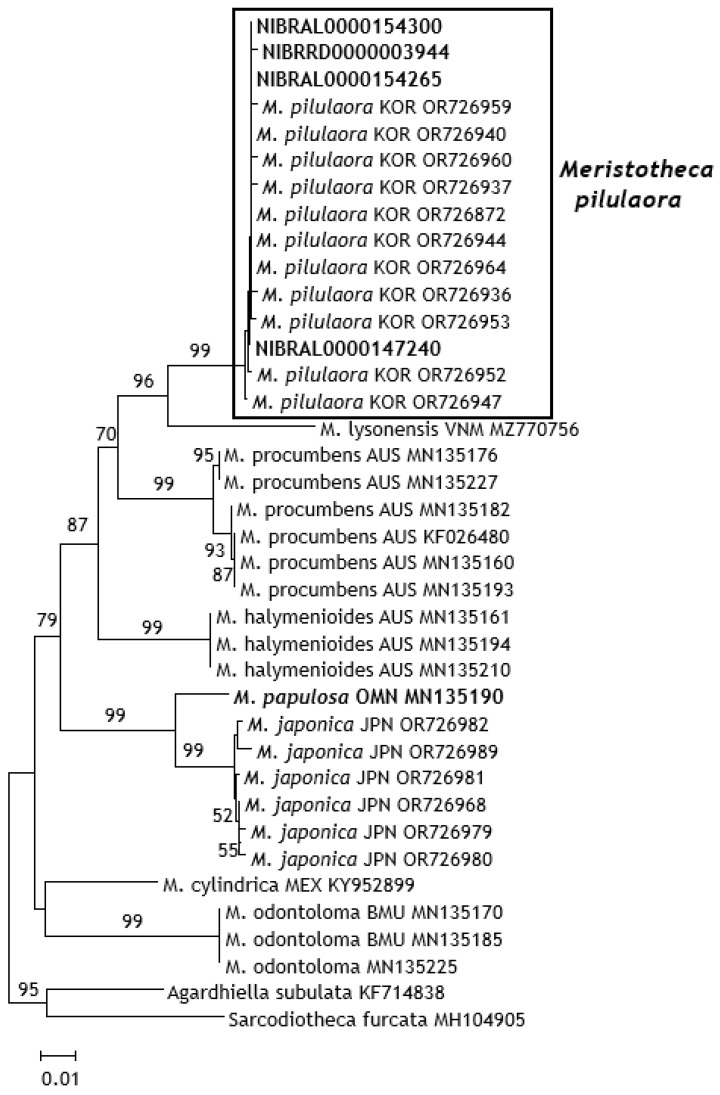 Figure 3
Figure 3- —National Fishery Products Quality Management Service (Development and Management of Disease Control Program for Aquatic Life)
- —National Institute of Biological Resources (NIBR)
- —Korea Ministry of Environment (MOE)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSeaweed-derived Bioactive Compounds · Marine and coastal plant biology · Agriculture, Soil, Plant Science
1. Introduction
Meristotheca (Solieriaceae, Rhodophyta) species are edible red algae with economic value in Korea, Japan, and China [1,2]. The genus comprises species distributed worldwide [3]. To date, three Meristotheca species (M. coacta, M. papulosa, and M. pilulaora) have been reported from Korea [4]. Meristotheca papulosa has been widely reported in the Indo-Pacific region [3], whereas M. coacta has only rarely been included in the floristic list [4,5]. Meristotheca pilulaora was described as a new species, which was previously considered as M. papulosa on Jeju Island (Korea) [6].
The type locality of M. papulosa (Montagne) J. Agardh 1872 is Yemen, in the southern Arabian Peninsula and it has been reported from Africa, Asia, Australia, New Zealand, and the Pacific Islands [3]. Ecological and physiological studies on M. papulosa have been performed in Korea and Japan [2,7]. In Korea, M. papulosa has been reported only from Jeju Island [2,4,5,6].
DNA analyses of herbarium specimens could be used to explore the species diversity and identify new taxonomic entities, including novel and unrecorded species [8,9,10,11,12,13,14]. Taxonomists spend considerable time collecting specimens from various regions. Even though herbarium specimen analyses are likely to provide a diverse and unbiased approach to species classification, several biases could be introduced [10]. Therefore, morphological species identification without genetic information would likely establish the misidentification of related species with morphological similarities [11].
Numerous specimens are stored in herbaria worldwide. Therefore, DNA barcoding of herbarium specimens could be a useful taxonomic tool for identifying species. Moreover, the genetic diversity of the collected samples could provide new taxonomic information, including the identification of entirely new species and species previously unrecorded in local regions [8,9,10,11,12,13,14,15].
A recent rapid reduction in natural M. papulosa populations on Jeju Island in Korea indicates the necessity to monitor fluctuations in Meristotheca populations [2]. The National Institute of Biological Resources (NIBR, Korea; https://species.nibr.go.kr/index.do; accessed on 31 January 2026) possesses M. papulosa herbarium specimens, including those from a recent study on an algal herbarium, indicating broad distribution of the species around the Korean peninsula [5].
The verification of herbarium specimens would provide not only accurate taxonomic information based on morphological and genetic data, but also important basic information for future industrial use. The new genetic resources obtained from algal herbarium specimens could present important information about seaweeds that exist in natural ecosystems, as the specimens contain data such as the collection site and date of collection from natural seaweed beds [2,7].
In this study, we re-examined M. papulosa herbarium specimens collected from the Korean Peninsula and developed a molecular marker to discriminate species with similar morphologies. Identification of a given species is the first step in estimating the economic and ecological value of seaweeds. In particular, accurate identification of the taxonomic entities of M. papulosa specimens and the development of new molecular markers could provide useful information for further studies in taxonomy and aquaculture.
2. Materials and Methods
In this study, herbarium specimens of Meristotheca papulosa from the NIBR (Korea) were analyzed (Table 1 and Table 2; Figure 1). We focused on the molecular analyses of specimens identified as M. papulosa from previous field surveys based on morphological examination.
Considering the collection sites and dates, we selected 22 sheets of M. papulosa herbarium specimens collected around the Korean Peninsula and successfully identified 12 specimens (Table 1 and Table 2; Figure 1). We obtained a small amount (<0.5 cm^2^) of sample from a single herbarium specimen for DNA extraction; the extracted DNA was subsequently subjected to polymerase chain reaction (PCR) analysis and sequencing as described previously [11]. Total genomic DNA was extracted using the DNeasy Plant Mini Kit (Qiagen, Germantown, MD, USA) following previously described protocols [11].
We used multiple molecular markers based on reference sequences deposited in GenBank (National Center for Biotechnology Information, NCBI). Two organelle-coding genes (rbcL, a plastid gene; cox1, a mitochondrial gene) were selected as molecular markers. New primers for the rbcL and cox1 regions were designed. Forward (rbcL-Red-8F; 5′-AATCTGTAGAAGAACGGACA-3′) and reverse (rbcL-Gig-978R; 5′-TACACCWGCCATACGCATCC-3′) primers were designed to amplify a 971-bp fragment in the rbcL region of the herbarium specimens (Figure 2A).
For the amplification of the cox1 region, two sets of cox1 primers were designed. For Meristotheca species, forward (cox1-Mer-84F; 5′-TGGTGCYTTTTCAGGTTTAATWGGA-3′) and reverse (cox1-Gig-749R; 5′-TCAGGGTGGCCRAARAAYCA-3′) primers were designed (Figure 2B) with 666-bp amplicons. For G. textorii, a different cox1-GraT-378F/cox1-Gig-749R primer set was used (cox1-GraT-378F (5′- CGAAGTTGGTGTAGGGACAGTA 3′)) with 372-bp amplicons (Figure 2C).
PCR was performed with an initial denaturation step at 94 °C for 3 min, followed by 40 cycles at 94 °C for 30 s, 50 °C for 30 s, and 72 °C for 1 min; and a final extension step at 72 °C for 7 min using Gene Amp-PCR-System-9700 (Applied Biosystems, Foster City, CA, USA). AmfiXpand PCR Master Mix (GenDEPOT, Katy, TX, USA) was used for PCR, and the products were sequenced using a commercial sequencing service provider (GenoTech, Daejeon, Republic of Korea). Chromatograms of sequencing were assembled in both directions using the Sequencher 5.4.6 software (Gene Codes, Ann Arbor, MI, USA). The sequencing quality was checked using the chromatograms, and we used the high-quality region in the sequencing data.
We successfully isolated the rbcL and cox1 sequences from 12 herbarium specimens of M. papulosa. The DNA sequences have been deposited in GenBank (the accession numbers are presented in Table 1 and Table 2). DNA sequences (rbcL and cox1) isolated from Korean herbarium specimens were compared with reference sequences deposited in GenBank at the NCBI (Meristotheca pilulaora (rbcL and cox1) and Gracilaria textorii (ptDNA NC_046043 and mtDNA NC_037892) (Figure 3).
For molecular phylogenetic analyses, reference sequences for cox1 were obtained from GenBank (NCBI) (Figure 3). Molecular phylogenetic analyses were conducted using MEGA ver. 6 [15] using the neighbor-joining and maximum likelihood methods with 2000 bootstrap replicates. The pairwise distances were calculated using Kimura’s two-parameter method. Outgroups were selected based on a previous phylogenetic study of Solieriaceae [16].
3. Results
We obtained four rbcL gene sequences [OP554369–OP554371 (878 bp) and OP554372 (836 bp)] and four cox1 gene sequences (622 bp of OQ594746–OQ594749 (622 bp)] from six specimens (Meristotheca pilulaora; Table 1, Figure 2). We isolated six rbcL [PX613529–PX613534 (931 bp)] and six cox1 [PX613535—PX613540 (330 bp)] DNA sequences from six specimens and used them for species identification (Gracilaria textorii; Table 2, Figure 2). The primer pair of cox1 marker (cox1-Mer-84F/cox1-Gig-751R) produced 668-bp amplicons from M. pilulaora DNA templates, but not from G. textorii DNA samples (Figure 2B). The primer combination cox1-GraT-378F/cox1-Gig-751R of cox1 marker for Gracilaria species produced 374-bp amplicons from G. textorii DNA templates (Figure 2C).
Using BLAST search (GenBank, NCBI), we identified six “M. papulosa” specimens as M. pilulaora, recently established in Korea [6] (Table 1, Figure 3). Moreover, the other six “M. papulosa” specimens exhibited 100% identity with G. textorii (ptDNA NC_046043 and mtDNA NC_037892) (Table 2).
The rbcL sequences of five “M. papulosa” specimens contained the same DNA sequence and showed 100% identity with those of M. pilulaora populations. The cox1 gene of four “M. papulosa” specimens showed a difference of just one base among the specimens (NIBRRD0000003944 differed by one base from the sequences of the other specimens), demonstrating 99.8–100% identity with that of the M. pilulaora population in GenBank. Moreover, the “M. papulosa” specimens showed 93.7–89.8% cox1 sequence variation at the interspecific level. Therefore, the “M. papulosa” specimens examined in this study could be re-identified as M. pilulaora, as reported previously [6] (Figure 3).
In this study, we revealed a high level of misidentification (i.e., 50%, corresponding to 6 out of 12 specimens) of G. textorii, which was originally identified as M. papulosa. Moreover, we developed molecular markers for effective identification of M. pilulaora and G. textorii herbarium specimens. Moreover, M. pilulaora and G. textorii were successfully distinguished by the electrophoresis experiments using cox1 molecular markers (Figure 2B,C).
4. Discussion
The herbarium DNA barcoding has enabled to identify the biodiversity [8,9,10,11,12,13,14,15]. During the taxonomic re-examination of herbarium specimens of M. papulosa in the NIBR, we identified new genetic and species diversity in the algal herbarium (e.g., [11]).
Six of the 12 “M. papulosa” specimens examined were identified as M. pilulaora (Gigartinales; Solieriaceae) and the other six as G. textorii (Gracilariales; Gracilariaceae). Although M. papulosa and M. coacta have been reported by previous studies on Korean algal flora [4,5], they were not identified in this study. In a previous study [6], M. pilulaora was found only in the coastal regions of Jeju Island (Korea). In contrast, the herbarium DNA barcoding method used in the present study revealed the distribution of M. pilulaora, for the first time, on Ulleung Island, Korea (Table 1).
Regarding the Korean species of the genus Gracilaria, molecular phylogenetic studies were conducted with morphological observations [17,18]. We uncovered a misidentification at the inter-ordinal level between M. pilulaora and G. textorii (6/12 samples) (Table 2). Thus, most herbarium specimens lacking evidence of DNA sequences could have been taxonomically misidentified (e.g., [11]). Therefore, taxonomic re-examination is required to verify herbarium specimens. Our approach can provide a case study for the DNA barcoding method for herbarium specimens, and the molecular markers developed in this study represented an effective molecular tool for the species-level identification between M. pilulaora and G. textorii.
5. Conclusions
Many taxonomists worldwide have spent considerable time searching for new and unrecorded species. Moreover, numerous herbarium specimens have been deposited without DNA evidence to confirm species identification. Our work represents a case study for establishing misidentification in herbarium specimens and finding a new geographic distribution. Therefore, the DNA barcoding analysis for herbarium specimens can provide the useful information for algal taxonomic studies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Faye E.J. Shimada S. Kawaguchi S. Masuda M. Characterization of the edible red alga Meristotheca papulosa (Gigartinales, Solieriaceae) from Japan Phycol. Res.20055323424510.1111/j.1440-1835.2005.tb 00376.x · doi ↗
- 2Kim B.Y. Ko J.C. Shan T. Choi H.G. Reproductive capacity of Meristotheca papulosa (Solieriaceae, Rhodophyta) and effects of temperature on carpospore release and sporeling growth Phycol. Res.202573707810.1111/pre.12575 · doi ↗
- 3Guiry M.D. Guiry G.M. Algae Base; World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2025 Available online: www.algaebase.org(accessed on 4 December 2025)
- 4Lee Y.P. Kang S.Y. A Catalogue of the Seaweeds in Korea Jeju National University Press Jeju, Republic of Korea 2001425
- 5National Institute of Biological Resources 2024 Biodiversity Statistics of Korea National Institute of Biological Resources Incheon, Republic of Korea 2024 Available online: https://kbr.go.kr/content/view.do?menu Key=799&content Key=174(accessed on 11 December 2025)
- 6Yang M.Y. Kang J.C. Fujita D. Kim M.S. Molecular phylogeny and genetic diversity of the economic seaweed Meristotheca (Gigartinales, Rhodophyta) in the Northwest Pacific, with a description of M. pilulaora sp. nov J. Appl. Phycol.20243648549910.1007/s 10811-023-03135-4 · doi ↗
- 7Nishihara G.N. Noro T. Terada R. Effect of temperature and light on the photosynthetic performance of two edible sea-weeds: Meristotheca coacta Okamura and Meristotheca papulosa J. Agardh (Solieriaceae, Rhodophyta)Aquacult. Sci.201260377388
- 8Bebber D.P. Carine M.A. Wood J.R. Wortley A.H. Harris D.J. Prance G.T. Davidse G. Paige J. Pennington T.D. Robson N.K.B. Herbaria are a major frontier for species discovery Proc. Natl. Acad. Sci. USA 2010107221692217110.1073/pnas.101184110821135225 PMC 3009773 · doi ↗ · pubmed ↗