Noncoding RNAs in Pediatric Solid Tumors: Advances in Understanding and Critical Knowledge Gaps
Graham Duff, Christine Mella, Alexa Amato-Loudon, Meredith Farrell, Rachael Aldridge, Hope C. Ball

TL;DR
This paper reviews the role of noncoding RNAs in pediatric solid tumors, emphasizing their potential as biomarkers and therapies while highlighting key research gaps.
Contribution
The paper provides a comprehensive overview of ncRNAs in pediatric cancers, identifying novel research directions and challenges.
Findings
Noncoding RNAs regulate gene expression and may serve as biomarkers in pediatric solid tumors.
Current research on miRNAs, lncRNAs, and circRNAs reveals their roles in tumor progression and treatment response.
Significant knowledge gaps remain in understanding ncRNA functions and their therapeutic potential.
Abstract
The etiology of pediatric cancers is unique, stemming from developmental dysregulation rather than acquired mutations from carcinogenic exposure. These diseases demonstrate vastly different underlying genetic and epigenetic alterations and unique tissue microenvironments which are only now beginning to be explored. While many pediatric cancers have seen improved overall and event-free survival rates thanks to innovations in diagnosis and treatment, many have seen little to no improvement in patient outcomes. This highlights a critical need for additional research into the underlying genetic and epigenetic alterations in these pathologies. Non-coding RNAs (ncRNAs) are functional RNA molecules known to regulate gene expression at epigenetic, transcriptional, and translational levels and can serve as biomarkers of disease. Here, we examine current knowledge of the roles of microRNAs…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
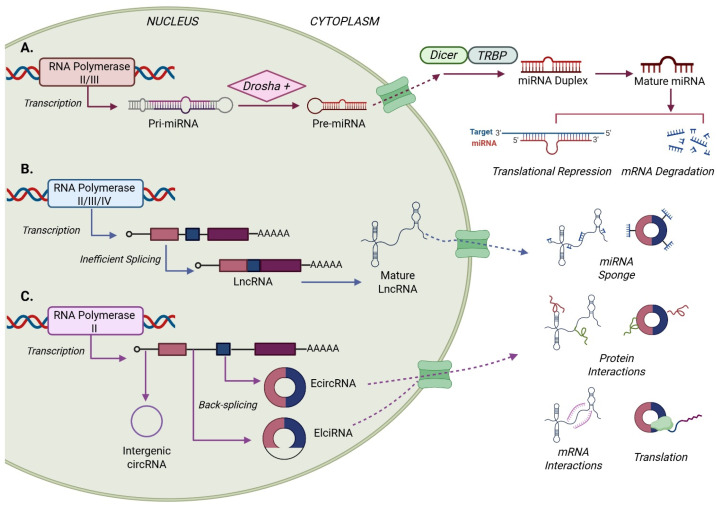 Figure 1
Figure 1 Figure 2
Figure 2| Disease/ | Expression | Non-Coding RNA | Validated Target Gene(s) | Functional Role | Reference(s) |
|---|---|---|---|---|---|
| MB (WNT) | Downregulated | miRNAs-206, -183, -133b, -128a/b, -148a, -383 | N/A (Oncogenic targets) | Increases survival, proliferation, and migration | [ |
| MB (SHH) | Downregulated | miRNA-466-3p | NRP2 and VEGFA | Enhances EMT via unchecked modulation | [ |
| MB (SHH) | Upregulated | miRNA-17-92 cluster (18a, 19a, 20a, 21, 25, 106) | N/A | Promotes proliferation and tumor progression | [ |
| MB (Group 3) | Upregulated | miRNA cluster 183-96-182 | Cell cycle regulation | Promotes migration; alters cell cycle | [ |
| MB (Group 3) | Downregulated | miRNA-30 family | Autophagy (process) | Reduces cancer cell autophagy | [ |
| MB (Group 4) | Downregulated | miRNA-4521 | FOXM1 | Increases proliferation | [ |
| MB | Upregulated | lncRNA HOTAIR/TP73-AS1 | miRNA Sponges | Promotes tumorigenicity, migration, and viability | [ |
| MB (Group 3) | Upregulated | lncMB1, lncMB2, lncMB3 | MYC | Associated with aggressive MYC subgroup | [ |
| MB (SHH) | Upregulated | lncRNA LOXL1-AS1 | N/A | Linked to poor prognosis | [ |
| MB | Upregulated | lncRNA NEAT1 | N/A | Linked to chemo- and apoptotic resistance | [ |
| MB | Downregulated | lncRNA Nkx2-2as | N/A | Suppresses proliferation and invasion | [ |
| MB | Upregulated | circ_SKA3 | CDK6 | Promotes tumor invasion | [ |
| MB (SHH) | Upregulated | circ_DTL, _KDHRBS2, -103128, _63706 | Lipid metabolism | Alters metabolic regulation | [ |
| Disease/ | Expression | Non-Coding RNA | Validated Target Gene(s) | Functional Role | Reference(s) |
|---|---|---|---|---|---|
| RB | Upregulated | miRNA-320a | TUSC3 | Promotes proliferation and apoptotic resistance | [ |
| RB | Upregulated | miRNA-889-3p | JNK/MAPK/ERK pathway | Promotes tumor growth; inhibits apoptosis | [ |
| RB | Upregulated | miRNA-25-3p | PTEN | Enhances EMT and cell migration | [ |
| RB | Upregulated | miRNA-141-3p | SUSD2 | Promotes angiogenesis and proliferation | [ |
| RB | Upregulated | miRNA-222 | VHL | Promotes chemotherapy resistance | [ |
| RB | Upregulated | miRNA-224-3p/miRNA-492 | LATS2 | Increases cancer invasion | [ |
| RB | Upregulated | miRNA-92a-3p | KLF2 | Enhances tumor-associated angiogenesis | [ |
| RB | Downregulated | miRNA-34a/34b-5p | NOTCH signaling | Increases chemosensitivity; suppresses invasion | [ |
| RB | Downregulated | miRNA-153-3p | IGFR1/Raf/MEK & PI3K/AKT pathways | Leads to unchecked proliferation/survival | [ |
| RB | Downregulated | miRNA-361-3p | GLI1/GLI3 | Increases expression of Hedgehog factors | [ |
| RB | Upregulated | lncRNA-MALAT1 | miRNA-20b-5p (STAT3)/miRNA-124 & 598-3p | Upregulates STAT3; chemoresistance | [ |
| RB | Upregulated | lncRNA-NEAT1 | miRNA-124, -3619-5p, -24-3p, -106a, -148-3p | Promotes tumor growth (sponging) | [ |
| RB | Upregulated | lncRNA-XIST | miRNA-101/miRNA-140-5p & 191-5p | Contributes to EMT, proliferation, and invasion | [ |
| RB (Hypoxic) | Upregulated | lncRNA-ANRIL | miRNA-328 | Contributes to chemotherapeutic resistance | [ |
| RB | Downregulated | lncRNAs-MT1JP, -MBLN1, -MEG3 | WNT/β-catenin pathway | Tumor suppression | [ |
| RB | Upregulated | Circ-DHDDS | miRNA-361-3p | Promotes RB progression | [ |
| RB | Upregulated | circ-FAM158A | XIAP, SMAD2, and LRP6 | Promotes metastasis and optic nerve invasion | [ |
| RB | Upregulated | circ-E2F3 | miRNA-204-5p (ROCK1)/LASP1 | Enhances motility and invasion | [ |
| RB | Upregulated | circ_RNF20 | miRNA-132-3p (PAX6) | Promotes invasion | [ |
| RB | Downregulated | circ-TET1 | miRNA-492/miRNA-484-3p (WNT/β-catenin) | Targets WNT signaling | [ |
| RB | Downregulated | circ-SHPRH | AKT and mTOR | Promotes cell apoptosis | [ |
| RB | Downregulated | circ-MKLN1 | miRNA-425-5p | Curbs cell invasion | [ |
| RB | Downregulated | circ-CUL2 | E2F2 | Reduces proliferation and migration | [ |
| RB | Upregulated | miRNA-320a | TUSC3 | Promotes proliferation and apoptotic resistance | [ |
| RB | Upregulated | miRNA-889-3p | JNK/MAPK/ERK pathway | Promotes tumor growth; inhibits apoptosis | [ |
| RB | Upregulated | miRNA-25-3p | PTEN | Enhances EMT and cell migration | [ |
| Disease/ | Expression | Non-Coding RNA | Validated Target Gene(s) | Functional Role | Reference(s) |
|---|---|---|---|---|---|
| ARMS | Upregulated | miRNA-486-5p/-9-5p | PAX3-FOXO1 induced | Increases proliferation and invasion | [ |
| ARMS | Downregulated | miRNA-27a/-221/-222 | PAX3-FOXO1 repressed | Tumor suppression (loss of) | [ |
| ERMS | Upregulated | miRNA-130a/b | PPARG | Increases proliferation | [ |
| ERMS | Downregulated | miRNA-181a/-212 | N/A | Reduced myogenic differentiation; increased invasion | [ |
| RMS (Both) | Downregulated | miRNA-7/-324-5p | ITGA9 | Increases metastasis | [ |
| RMS (Both) | Upregulated | lncMYCNOS | N/A | Promotes tumor growth | [ |
| RMS (Both) | Upregulated | lncRNA-H19 | miRNA-675-3p/-375-5p | Promotes myogenic differentiation | [ |
| RMS (Both) | Upregulated | lncRNA-SYISL/ | N/A | Reduces differentiation; accelerates proliferation | [ |
| RMS (Both) | Upregulated | linc-MD1 | miRNA-206/-133b | Decreases differentiation; enhances proliferation | [ |
| RMS (Both) | Upregulated | circ-AFF1 | Cellular adhesion | Increases cell migration | [ |
| ARMS | Upregulated | circ-VAMP3/circ-ZNF609 | AKT pathway | Disrupts cell cycle regulation | [ |
| Disease/ | Expression | Non-Coding RNA | Validated Target Gene(s) | Functional Role | Reference(s) |
|---|---|---|---|---|---|
| HB | Upregulated | miRNA-193b, -760, -23a-5p | WNT pathway | Promotes disease progression | [ |
| HB | Upregulated | miRNA-193a-5p, -495, -206 | PI3K/AKT/mTOR pathway | Promotes oncogenesis | [ |
| HB | Upregulated | lncRNA MIR205HG, NBR2, TUG1, HOXA-AS2, ZFAS1, lnc01124 | MAPK and PI3K/AKT pathways | Modifies stemness; enhances invasion and proliferation | [ |
| HB | Upregulated | lncRNA OIP5-AS1 | WNT/β-catenin pathway | Promotes invasion | [ |
| HB | Upregulated | lncRNA HOXA-AS2, ZFAS1, CRNDE | EMT (process) | Increases malignancy and EMT | [ |
| HB | Upregulated | circ_0015756 | N/A | Enhances proliferation and viability | [ |
| HB | Upregulated | circ_0000594 | miRNA-217 | Promotes malignancy | [ |
| HB | Upregulated | circHMGCS1 (circ_0072391) | miR-503-5p (IGF/PI3K/Akt) | Activates IGF/PI3K/Akt; alters glutamine metabolism | [ |
| HB | Upregulated | circRNA_CCT2 | WNT/β-catenin pathway | Pro-tumor properties | [ |
| HB | Downregulated/Suppressive | circMTO1, circEPB41L2, circDLC1 | N/A | Halts HB progression | [ |
| HB | Upregulated | circRNA_SORE | N/A | Linked to increased chemoresistance | [ |
| HB | Upregulated | miRNA-193b, -760, -23a-5p | WNT pathway | Promotes disease progression | [ |
| Disease/ | Expression | Non-Coding RNA | Validated Target | Functional Role | Reference(s) |
|---|---|---|---|---|---|
| OS | Down-regulated | lncRNA-HIF2PUT | HIF2 | Correlates with poor prognosis | [ |
| OS | Upregulated | lncRNA-MALAT1 | PI3K/Akt/HMGB1 | Increases proliferation and metastasis | [ |
| OS | Upregulated | lncRNA-HOTAIR | Akt/mTOR/ZEB1/ALP | Promotes pathogenesis; inhibits mineralization | [ |
| OS | Upregulated | lncRNA-DANCR | AXL/MSI2 | Enhances stemness and immune evasion | [ |
| OS | Upregulated | lncRNA-CRNDE | WNT/β-catenin/ABCC12/NOTCH1 | Promotes metastasis, EMT, and proliferation | [ |
| OS | Upregulated | lncRNA-SNHG12 | N/A | Promotes metastasis; doxorubicin resistance | [ |
| OS | Upregulated | lncRNA-UCA1 | N/A | Increases invasion and tumor growth | [ |
| OS | Upregulated | lncRNA-THOR | Akt/ERK/SOX9 | Promotes tumor growth; stabilizes mRNA | [ |
| OS | Upregulated | circ-FOXM1 (hsa_circ_0025033) | miRNA-320a/b/WNT pathway | Contributes to proliferation and migration | [ |
| OS | Upregulated | circRAB3IP (hsa_circ_0000419) | miRNA-580-3p/TWIST1 | Promotes invasion | [ |
| OS | Upregulated | circ_03955 | miRNA-3662/MTDH | Enhances metadherin expression | [ |
| OS | Upregulated | circ-LRP6 | HDAC4/HMGB1 | Promotes invasion | [ |
| OS | Upregulated | circDOCK1 (hsa_circ_0020378) | IGF-1R/DNMT3A/LEF1 | Apoptotic and chemotherapy resistance | [ |
| OS | Upregulated | circ_001422 | FGF2/PI3K/Akt/E2F3 | Promotes activation of growth pathways | [ |
| OS | Upregulated | circPVT1 (hsa_circ_0001821) | Multiple oncogenic pathways | Diagnostic potential | [ |
| OS | Down-regulated | lncRNA-HIF2PUT | HIF2 | Correlates with poor prognosis | [ |
| Disease/ | Expression | Non-Coding RNA | Validated Target Gene(s) | Functional Role | Reference(s) |
|---|---|---|---|---|---|
| ES | Upregulated | miRNAs-30d, -125b, -21 | BAX | Prevents cancer cell apoptosis | [ |
| ES | Upregulated | miRNA-181c | TNFRSF6 | Promotes tumor growth | [ |
| ES | Upregulated | miRNA-130b | CDC42 | Cell cycle dysregulation | [ |
| ES | Downregulated | miRNA-683 | VEGFA | Inhibits angiogenesis | [ |
| ES | Downregulated | Let-7a | EWSR1 | Inhibits protein production (RISC) | [ |
| ES | Downregulated | Let-7b | RAS pathway/HIF1α | Inhibits survival; reduces TME hypoxia | [ |
| ES | Downregulated | miRNA-708-5p | MMP12 | Inhibits pathogenic ECM remodeling | [ |
| ES | Downregulated | miRNA-27a | IGF-1 | Reduces proliferation and tumor growth | [ |
| ES | Upregulated | lncRNA PncCCND1_B | CCND1 (Cyclin D1) | Promotes progression (sponging) | [ |
| ES | Upregulated | lncRNA FOXP4-ASI | TMPO (Thymopoietin) | Promotes progression (sponging) | [ |
| ES | Upregulated | lncRNA METTL3 | miRNA-124-3p/CDK4 | Promotes cell migration | [ |
| ES | Polymorphism | lncRNA SENCR | FLI1/IGF2BP3 | Upregulates IGF2BP3 | [ |
| ES | Upregulated | circZNF609 | miRNA-145-5p/EWS-FLI1 | Promotes metastasis; inhibits apoptosis | [ |
- —Akron Children’s Hospital Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCircular RNAs in diseases · Cancer-related molecular mechanisms research · MicroRNA in disease regulation
1. Introduction
Cancer affects both adult and pediatric patient populations and is a major global socioeconomic health problem [1,2]. Adult cancers often arise from accumulated genetic and/or epigenetic mutations affecting epithelial cell transcription/translation, cycle regulation, or DNA damage repair mechanisms [3,4]. The etiology of pediatric cancers, however, lies in the developmental dysregulation of mesoderm-, ectoderm-, or endoderm-originating cells, resulting in fusion genes or oncogenic changes in stem cell populations or the tissue microenvironment (TME) [5,6,7]. Furthermore, the pediatric cancer-associated gene mutations are significantly different from adult-onset counterparts, and the mechanisms of disease onset, metastasis, and therapeutic response remain poorly understood.
In the pediatric population, cancer stands as the second leading cause of death, second only to injury-related mortalities [8,9]. Solid tumors comprise around 60% of all diagnosed pediatric cancers and include intracranial (central nervous system CNS, neuroblastoma) and extracranial (Wilms, hepatoblastoma, retinoblastoma, soft tissue sarcomas, rhabdomyosarcomas, osteosarcoma, Ewings sarcoma) tumors [10,11]. Improvements in chemotherapy/radiotherapy regimens, surgical approaches, and targeted gene therapies have improved childhood cancer survival overall, but treatment options remain limited for children with metastatic, relapsed, or therapy-resistant disease [12,13]. Recent years have seen a significant rise in the use of next-generation sequencing (NGS) assessment of patient samples, leading to improved understanding of the underlying mutations and pathways in many pediatric cancer types. In cancers that lack common risk factors, demonstrate high genetic heterogeneity, or arise spontaneously, studies have focused on elucidating alternative mechanisms of developmental genetic/epigenetic regulation such as non-coding RNAs (ncRNAs).
ncRNAs have known roles in cancer pathogenesis where they regulate oncogenic pathways, serve as potential therapeutic targets, and as biomarkers for disease sub-type and/or stage [14,15,16]. Here, we provide an updated review on the role of ncRNAs in the pediatric solid tumors medulloblastoma, retinoblastoma, rhabdomyosarcoma, hepatoblastoma, osteosarcoma, and Ewing sarcoma. These malignancies were selected based on diagnostic prevalence and scientific advancement in the roles on ncRNAs for clinical and therapeutic applications. We provide in-depth insight into the roles of miRNAs, lncRNAs, and circRNAs in tumor biogenesis, progression, metastasis, and therapy resistance, and identify areas of interest that could benefit from additional scientific study.
2. Materials and Methods
A PUBMED literature review was completed to search for original research articles and recent in-depth reviews detailing the role of various non-coding RNAs on solid tumor onset, progression, metastasis, clinical correlation, and therapeutics in the pediatric population. Database searches utilized the specific disease name, pediatric, and the key words “pathogenesis,” “prevalence,” “therapeutic treatment,” “outcome,” “survival,” “miRNA,” “long non-coding RNA,” circRNA,” “non-coding RNA,” “non-coding RNA and clinical correlate,” and “RNA pediatric cancer biomarker.” Figures were created using BioRender.com (Toronto, ON, Canada)
3. Biogenesis of miRNAs, lncRNAs, and circRNAs and Their Roles in Health and Disease
ncRNAs are RNA molecules that do not have protein-coding functions, but regulate gene expression at epigenetic, transcriptional, and translational levels [17]. There are many subtypes of ncRNAs classified according to size and three-dimensional structure. These include microRNAs (miRNAs), small nucleolar RNAs (snoRNAs), circular RNAs (circRNAs), Piwi-interacting RNAs (piRNAs), small interfering RNAs (siRNAs), small cytoplasmic RNAs (scRNAs), and long non-coding RNAs (lncRNAs) [18]. All ncRNA subtypes demonstrate critical roles in cell and tissue homeostasis, inter- and intra-cellular signaling, gene expression, and pathway activation, and their dysregulation contributes to the pathogenesis of human disease [19].
Micro-RNAs (miRNAs) are small ncRNAs measuring approximately 22 nucleotides (nt) in length that are capable of promoting mRNA degradation via the formation of RNA-induced silencing complex (RISC) or causing translational repression through imperfect binding to the 3′ untranslated regions (UTR) of mRNA. miRNA biogenesis begins with RNA polymerase II/III transcription of pri-miRNAs which are cleaved to form pre-miRNAs before being exported to the cytoplasm for Dicer-associated processing (Figure 1) [20]. Once within a RISC, miRNAs can bind mRNA targets to induce endonuclease degradation or 3′UTRs for translational repression or interact with other ncRNAs to regulate expression profiles and/or signaling pathways [21]. Their role in health and disease is influenced by the tight regulation of expression in cell-type, developmental, spatial, and temporal manners [22]. In pediatric cancer, miRNAs are of increasing interest due to their roles as regulators of oncogenic pathways, apoptosis, and cell metabolism, as well as their potential as biomarkers and precision therapeutic targets [23,24]. Next generation sequencing studies of cell lines and patient tumors are continuing to identify novel miRNAs and better elucidate how alterations in miRNA expression correlate with clinical diagnosis, therapeutic response, and overall survival (Figure 2) [25].
Long non-coding RNAs (lncRNAs) are modular RNAs greater than 200 nt in length that often comprise repeating units and demonstrate tissue- and cell lineage-specific spatiotemporal expression patterns. lncRNAs are transcribed by RNA polymerases I/II/III and undergo inefficient splicing to form mature lncRNAs (Figure 1) [26,27,28]. Mature lncRNAs influence normal cell differentiation, development, and play a role in the pathology of disease [29]. In cancer, lncRNAs modulate cell metabolism, intra- and intercellular signaling, transcription, and oncogenic or tumor-suppressive processes through genetic/epigenetic mechanisms such as miRNA sponging, RNA-RNA, RNA-protein, and/or RNA-DNA interactions or via chromatin or histone associations (Figure 2) [30,31].
Circular RNAs (circRNAs) are formed from single-stranded, covalently bonded circular RNA created from alternative splicing of PolII-transcribed pre-mRNAs (Figure 1) [32]. The sequence types that are included during splicing lead to the generation of three distinct circRNA sub-types: EIciRNA (containing exon–intron sequences), ciRNAs (containing intron-only sequences), or ecircRNAs (containing exon sequences only) [33]. circRNAs function as miRNA sponges to modulate gene transcription, or interact with RNA-binding proteins, proteases, and/or transcription factors to regulate gene/protein function and stability [34,35]. circRNAs play a role in normal cell differentiation, embryonic development, and organogenesis and have been implicated in the onset and progression of diseases including Alzheimer’s, Parkinson’s, diabetes, and cancer, where they contribute to tumorigenesis by enhancing cell proliferation and invasiveness and are being examined as potential cancer biomarkers and as potential therapeutic targets (Figure 2) [36,37].
4. ncRNAs in Pediatric Solid Tumors
4.1. Medulloblastoma
Medulloblastoma (MB) is the most common undifferentiated central nervous system (CNS) embryonal neuroectodermal tumor that represents between 15 and 20% of all childhood CNS tumors [38,39]. Commonly arising in the cerebellum, these cancers are aggressive with metastatic lesions disseminating via cerebral spinal fluid (CSF) to the spine and leptomeningeal and subarachnoid spaces [40,41]. MB is divided into four sub-groups based on histological and molecular characteristics: Sonic-Hedgehog-amplified (SHH), Wingless amplified (WNT), and non-WNT/non-SHH groups 3 and 4 [42,43]. Of these groups, SHH-amplified and non-WNT/non-SHH are the most common. Common symptoms at presentation include persistent morning headaches, nausea, ataxia, and unexplained changes in coordination and/or behavior [44,45]. Standard-of-care therapy is interdisciplinary and includes adjuvant/neo-adjuvant chemotherapy, gross surgical resection, and craniospinal irradiation [46,47]. Overall survival rates and risk of recurrence vary greatly between subgroups. Diagnoses of WNT- and SHH-amplified MB are considered more favorable than non-WNT/non-SHH amplified group 3 and group 4, which demonstrate the highest rate of metastasis at diagnosis (40–45%), the highest risk of recurrence, and worst survival rates of all subtypes [48,49,50]. Recent research has greatly advanced our understanding of the biological and pathological roles of ncRNAs in MB (Table 1).
Of all MB-associated ncRNAs, miRNAs are the best known, and their roles in apoptosis, tumor suppression, metastasis, and associations with specific sub-groups are beginning to be elucidated. The four sub-groups of MB are known to demonstrate unique miRNA profiles that correlate to prognosis and clinical outcome. Within the WNT-amplified subgroup, miRNAs-206, -183, -133b, -128a/b, -148a, and -383 are significantly reduced, resulting in overexpression of their oncogenic targets and increased cell survival, proliferation, and migration [51,52,53,54,55]. In SHH amplified MB, let-7 has been identified as a potential biomarker, and miRNA-466-3p has been shown to be downregulated, leading to enhanced epithelial–mesenchymal cell transition via unchecked modulation of target genes neuropilin 2 (Nrp2) and vascular endothelial growth factor alpha (Vegfa) [56,57]. Also, inhibition of miRNAs-18a, -19a, -20a, -21, -25, and -106 (together, the miRNA-17-92 cluster) has been shown to significantly inhibit cancer proliferation and tumor progression [58]. Group 3 and 4 non-WNT/non-SHH amplified are the most aggressive MB sub-groups and share alterations in chromosome 17 structure and/or number [59]. In Group 3, elevated expression of the miRNA cluster 183-96-182 promotes cell migration and alters cell cycle regulation in both mouse MB models and in human cell lines and tissues [60,61]. Furthermore, studies demonstrate downregulation of the miRNA-30 family in Group 3 MB results in reduced cancer cell autophagy, which can be restored with lentiviral miRNA-30a treatment [62,63]. Group 4 MB cases demonstrate loss of miRNA-4521, leading to increased cell proliferation via overexpression of oncogenic transcription factor forkhead box M1 (FOXM1) [64,65]. Also, Group 4 MBs commonly overexpress miRNA-187-3p and miRNA-660-5p, which have been linked to poor survival rates [66].
Long non-coding RNAs (lncRNAs) are another MB-associated ncRNA with oncogenic and tumor suppressive functions [67,68]. lncRNAs HOTAIR (HOX transcript antisense RNA) and TP73-AS1 (P73 antisense RNA 1T) have both been found to be significantly enhanced in MB tumors and cell lines, where they function as miRNA sponges to promote in vivo tumorgenicity and in vitro cancer cell migration, viability, and proliferation [69,70,71]. Several other lncRNAs have been identified as drivers of MB progression and metastasis. These include SPRY4-IT1 (SPRIGHTLY), LOXL1-AS1 (LOXL1 antisense RNA1), Linc-NeD125 (neuronal differentiation long intergenic non-coding RNA NeD125), CCAT1 (colon cancer associated transcript 1), CRNDE (colorectal neoplasia differentially expressed), HHIP-AS1 (Hedgehog interacting protein antisense 1), and UCA1 (Urothelial carcinoma associated 1) [72,73,74,75,76,77,78,79,80]. Other lncRNAs have been identified with links to specific MB sub-groups. For instance, lncRNAs lncMB1, lncMB2, and lncMB3 have been linked to the aggressive Group 3 MYC proto-oncogen (MYC) subgroup, and LOXL1-AS1 is associated with poor prognosis in SHH MB. Finally, lncRNA NEAT1 (nuclear enriched abundant transcript 1) is associated with MB chemo- and apoptotic resistance, and Nkx2-2as (Nkx2-2 antisense RNA1) has been shown to suppress MB cell proliferation and invasion in vitro [81,82].
While circRNAs are less well-known in the pathophysiology of MB, several have been identified and associated with the disease. One of the most studied is Circ_SKA3, which is upregulated in MB tissues compared to normal cerebellum, and has been shown to promote tumor invasion by maintaining cyclin-dependent kinase 6 activity [83,84]. Other upregulated circRNAs include circ_DTL, circ_KDHRBS2, circ-103128, and circ_63706, which have been associated with altered lipid metabolism in SHH subgroup MB [85,86,87]. Other circRNAs such as circ_CRTAM, circ_MAP3K5, circ_RIMS1-1, and circ_FLT3-1 are significantly downregulated in MB, but their roles and mechanisms remain largely unknown [88].
4.2. Retinoblastoma
Retinoblastoma (RB) is the most common type of ocular malignancy affecting children between two and four years of age [89]. While RB does not demonstrate gender predisposition, incidence does show geographical and socioeconomic correlations. Geographically, higher incidence rates occur in the Andean regions of Latin America, Asia, Western Europe, and in the eastern regions of sub-Saharan Africa [90,91]. Socioeconomically, incidences rates and age at diagnosis are both higher in lower-income nations [92]. Global prevalence is on the rise with a 34% increase in diagnoses since 1990 [93]. RB is caused by de novo inactivation of the retinoblastoma 1 gene (RB1) on chromosome 13q14, resulting in dysregulated DNA replication, cell differentiation, and cell senescence, and impairments in chromatin remodeling [94,95]. Inactivation can occur through RB1 mutations or exon deletion and can be inherited (autosomal dominant, ~40% of cases) or somatic (~60% of cases) in nature [96,97]. Mode of inheritance influences clinical presentation, with bilateral disease associated with inherited mutations and unilateral with somatic [98]. Along with RB1, activating mutations in MYCN proto-oncogene (MYCN) are also associated with RB onset and progression [94]. Common clinical presentation of primary RB is an abnormal whitish retinal reflection (leukocoria), but other symptoms may include bilateral/unilateral strabismus or vision changes [99,100]. If metastasized at time of diagnosis, alternative presentations may include orbital swelling, intraocular pressure changes or hemorrhage, lymph node enlargement, or bone pain [101,102]. Treatment strategies vary widely based on accessibility of care and include enucleation as well as intravitreal, intracameral, periocular, or intra-arterial delivery of chemotherapy agents such as Melphalan, Topotecan and/or Carboplatin [103,104]. Additional systemic chemotherapy can also be applied, commonly employing Vincristine, Etoposide, and/or Carboplatin agents [105,106]. Overall survival rates for patients with primary RB are ~90% for heritable RB1 mutations and ~95% for RB of somatic origin [96]. The most common causes of RB-related mortality are metastatic disease or recurrent primary RB [107,108]. Medical innovations to uncover innovative treatment options continue to be explored, including at the role of ncRNAs and their respective mechanisms of action (Table 2).
Expression profiles and pathway analyses have yielded tremendous information regarding the pro- and anti-tumor roles of miRNAs in RB proliferation, therapeutic resistance, and invasion. For instance, miRNA-320a has been found to promote RB cell proliferation and apoptotic resistance through the inhibition of tumor suppressor candidate 3 (TUSC3) [109]. miRNA-889-3p also promotes RB tumor growth and inhibits apoptosis through activation of the c-Jun N-terminal kinase (JNK)/mitogen-activated protein kinase (MAPK)/extracellular signal-related kinase (ERK) pathway [110]. Studies have shown miRNA-25-3p directly targets PTEN, enhancing epithelial-to-mesenchymal transition (EMT) and increasing cell migration [111]. miRNA-141-3p is also upregulated in RB, and promotes angiogenesis and cell proliferation by inhibiting sushi domain-containing protein 2 (SUSD2) [112]. miRNA-222 is also upregulated and promotes chemotherapy resistance via the inhibition of tumor suppressor Von Hippel–Lindau (VHL) [113]. Similarly, miRNAs-224-3p and-492 are upregulated and function to increase cancer invasion by suppressing the functions of enzyme large tumor suppressor kinase 2 (LATS2) [114,115]. Finally, RB exosome-derived miRNA-92a-3p enhances tumor-associated angiogenesis through suppression of Krüppel-like factor 2 (KLF2) [116]. Conversely, tumor suppressor miRNAs-34a and -34b-5p both inhibit neurogenic locus notch homology protein (NOTCH) signaling, which increases chemosensitivity and suppresses RB invasion, respectively [117,118]. Tumor suppressor miRNA-153-3p is downregulated in RB, resulting in unchecked expression of the insulin-like growth factor 1 receptor (IGFR1)/rapidly accelerated fibrosarcoma (Raf)/mitogen-activated protein kinase kinase 1 (MEK) and phosphoinositide 3-kinase (PI3K)/AKT serine/threonine kinase 1 (AKT) pathways [119]. Similarly, suppression of miRNA-361-3p results in the increased expression of Sonic Hedgehog transcription factors GLI family zinc fingers 1 and 3 (GLI1/GLI3) [120].
Studies into the tumorigenic or tumor-suppressive functions of lncRNAs in RB pathogenesis have greatly expanded in the past few years. For example, lncRNAs-AFAP1-AS1 and -BANCR are elevated in RB and correlated with increased optic nerve invasion and tumor size [121,122]. lncRNA-MALAT1 upregulates STAT3 expression via sponging inhibition of miRNA-20b-5p and also contributes to chemotherapy resistance by sponging miRNAs-124 and -598-3p [123,124]. lncRNA-NEAT1 also promotes RB tumor growth by sponging tumor suppressive miRNAs miRNA-124, -3619-5p, 24-3p, -106a, and -148-3p [125,126]. The lncRNA-XIST contributes to RB EMT via repression of miRNA-101 and increased proliferation and invasion through the inhibition of miRNAs-140-5p and -191-5p [127,128]. In RB, the expression of lncRNAs-TMPO-AS1 and -MIR17HG is increased in the hypoxic TME, where they promote invasion and disrupt cell cycle regulation, respectively [129,130]. Similarly, lncRNA-ANRIL is upregulated by HIF1α and contributes to chemotherapeutic resistance by sponging miRNA-328 [131]. Other significantly upregulated lncRNAs associated with RB include lncRNA-SNHG20, -HOTAIR, -PVT1, -LINC00202, and MIMT1 [132,133,134,135,136]. Tumor suppressor lncRNAs have also been identified in RB cell lines and tissues. lncRNAs-MT1JP, -MBLN1, and -MEG3 all target various aspects of the WNT/β-catenin pathway [137,138,139]. Two additional lncRNAs demonstrate clinical correlations in RB: lncRNAs -BDNF-AS and -NKILA are correlated with poor overall survival and tumor grade/size, respectively [140,141].
Recent studies have added to the body of literature regarding the role of circRNAs in RB onset, progression, and therapeutic response. Circ-DHDDS (has_circ_0000034) promotes RB progression via sponging tumor suppressor miRNA-361-3p [142,143]. circ-FAM158A (has_circ_0000527) promotes RB onset, metastasis, and optic nerve invasion through miRNA sponging, which increases XIAP and smad family member 2 (SMAD2) expression and regulates the expression of low-density lipoprotein receptor-related protein 6 (LRP6), [144,145]. Oncogenic circ-E2F3 (has-circ_0075804) sponges miRNA-204-5p to regulate rho-associated protein kinase 1 (ROCK1), and enhances RB motility and invasion via the regulation of LIM and SH3 protein 1 (LASP1) expression [146,147]. circ_RNF20 (has_circ_0087784) is also upregulated and promotes RB invasion by sponging miRNA-132-3p and increases PAX6 expression [148]. Tumor suppressor circRNAs have also been identified in RB. lncRNA circ-TET1 (has_circ_0093996) targets the WNT/β-catenin pathway by inhibiting miRNAs-492 and -484-3p [149]. Similarly, circ-SHPRH (has_circ_0001649) promotes cell apoptosis by promoting phosphorylyation and activation of AKT and mTOR [150]. circ-MKLN1 (has_circ_0082415) curbs RB-associated cell invasion by sponging tumorigenic miRNA-425-5p, and circ-CUL2 (hsa_circ_0000234) inhibits the expression of E2F transcription factor 2 (E2F2) to reduce RB-associated cellular proliferation and migration [151,152].
4.3. Rhabdomyosarcoma
Rhabdomyosarcoma (RMS) is a malignant soft tissue cancer that arises from cells of the myogenic lineage [153]. According to the World Health Organization (WHO), there are four main subtypes of RMS based on histological and molecular characteristics [154,155]. These include two commonly diagnosed types, alveolar (ARMS) and embryonal (ERMS), as well as the two rarer subtypes pleomorphic (usually diagnosed in adult patients) and spindle cell/sclerosing RMS, which carries a poor prognosis [156]. Between the two main subtypes, ARMS is less differentiated with a tightly packed alveolar histology and a higher probability of metastasis [157,158]. ARMS-associated fusion genes PAX3-Forkhead box 01 (FOXO1) and PAX7-FOXO1, identified in more than half of ARMS patients, drive increased proliferation rates and reduced differentiation [159]. In contrast, ERMS is more differentiated with a round skeletal muscle-like appearance and lacks associated fusion genes, although these have been identified in some rare cell lines [160]. ERMS-associated mutations affect the RAS pathway and have also been identified in tumor protein p53 (TP53), BCL6 corepressor (BCOR), and/or neurofibromin (NF1) [161,162]. In general, most RMS patients are asymptomatic, but localized pain, swelling, and/or impaired movement can present depending on tumor location [163]. Treatment involves a combination of chemotherapy, surgical resection, and radiation depending on tumor type and the extent/site of disease [164,165]. While there have been significant improvements in RMS imaging, diagnostics, and treatment strategies (low to moderate risk survival is 80–90%), patients without complete localized control or with high-risk, metastatic, or recurrent RMS still face survival rates of less than 30% [166]. This highlights a critical need for additional research to improve scientific understanding of RMS and identify novel therapeutic targets (Table 3).
The function and mechanisms of miRNAs in RMS onset and progression are the most studied of all ncRNAs. Compared with other pediatric solid tumor types, RMS-associated miRNA research has slowed in recent years with a focus shift to those involved in muscle growth and development as opposed to pathology. Studies conducted in tumors and cell lines have identified subtype-specific miRNAs. For instance, the ARMS-associated fusion of PAX3-FOXO1 leads to an increase in the expression of miRNAs-486-5p and -9-5p, which increase cell proliferation and invasion [167,168]. The same fusion gene also inhibits the expression of tumor suppressor miRNAs-27a, -221, and -222 [169,170]. miRNA-335-5p is also overexpressed in ARMS, and it may function as a disease biomarker for this subtype [171]. Oncogenic miRNA-130a/b expression is increased in ERMS, leading to increased cell proliferation and decreased levels of peroxisome proliferator-activated receptor gamma (PPARG) [172]. Tumor suppressor miRNAs-181a and -212 are both significantly reduced in ERMS leading to reduced myogenic differentiation and increased cell invasion [173]. Other miRNAs have no subtype specification. Oncogenic miRNA-223 is increased pediatric RMS where it promotes inflammation and EMT [174]. Decreased expression of tumor suppressor miRNAs-7 and -324-5p results in elevated alpha-9-integrin (ITGA9) expression and increasing RMS-associated metastasis [175]. Two other tumor suppressor miRNAs, miRNAs-28-3p and -193-5p, are also reduced in both RMS subtypes [176]. miRNAs-223 and -29a/b/c inhibit cancer cell aggressiveness in both ARMS and ERMS [177,178]. Finally, miRNA-26a demonstrates a reduced expression in RMS tissues and is under evaluation as a potential disease biomarker [179].
The role of lncRNAs in RMS remains largely unknown, with only a few RMS-associated molecules identified. lncMYCNOS was studied previously in the pathogenesis of neuroblastoma and in ARMS where tumor growth was inhibited following knockdown [180]. lncRNA-H19 promotes myogenic differentiation through the upregulation of miRNAs-675-3p and -375-5p [181,182]. lncRNAs-SYISL and -NEAT1 both reduce myogenic differentiation and drive accelerated proliferation rates of RMS cells [183,184]. Inhibition of tumor suppressor lncRNA-GAS5 leads to increased proliferation and decreased apoptosis [185]. Finally, linc-MD1 decreases myogenic differentiation and enhances proliferation via binding inhibition miRNAs-206 and -133b [186].
Similar to lncRNAs, the expression and mechanisms underlying circRNAs in RMS pathophysiology are poorly understood. There are only three RMS-associated circRNAs that have been studied for disease relevance and mechanism. circ-AFF1 is increased in both ARMS and ERMS, where it increases cell migration via altered cellular adhesion [187]. Overexpression of circ-VAMP3 and circ-ZNF609 in RMS disrupts normal cell cycle regulation in ARMS and alters AKT pathway signaling [188,189]. The lack of knowledge regarding circRNAs in RMS pathology highlights a critical area that could benefit from further study to better understand the roles of these molecules in the disease and identify potential target for therapy development.
4.4. Hepatoblastoma
Hepatoblastoma (HB) is a rare, malignant liver tumor affecting children three years of age and younger [190]. Contributing risk factors include overgrowth syndromes or congenital syndromes such as Beckwith–Wiedemann syndrome (BWS) and familial adenomatous polyposis (FAP) due to chromosomal alterations and inherited mutations in adenomatous polyposis colic (APC) gene structure [191,192]. HB diagnoses also demonstrate significantly higher rates in Asian countries than Western counterparts [193]. Also, premature children with low birth weight (<1500 g) demonstrate predisposition for the disease [194]. Genetic studies have identified oncogenic mutations in the beta-catenin (CTNNB1) and NFE2 like BZIP transcription factor 2 (NFE2L2) genes, as well as the promoter regions of telomerase reverse transcriptase (TERT) that result in increased tumor growth [195,196]. Symptoms at clinical presentation include a painless abdominal mass (often distended), jaundice, unexplained weight loss, and/or hepatomegaly [190]. Official diagnosis is confirmed with a combination of imaging (CT or MRI), serum levels of HB biomarker alpha fetoprotein (AFP), and/or histology following biopsy [197]. Treatment is multimodal and includes a combination of surgical resection and adjuvant/neoadjuvant chemotherapy, with liver transplant utilized in a large portion of higher risk, therapy responsive cases [198,199]. HB patients treated with the standard of care see overall survival rates around 70%, but patients with small cell undifferentiated, metastatic, or relapsed disease have an unfavorable prognosis [200]. Next-generation sequencing of HB cell lines and patient-derived tissues have improved our understanding of the role of ncRNAs in HB pathogenesis, progression, and relapse (Table 4).
The role of miRNAs in the pathogenesis of HB is the most well-characterized of all the ncRNAs, although knowledge remains moderately sparse. Due to the importance of the WNT pathway in HB disease progression, several WNT pathway-associated miRNAs have been identified, including miRNAs-193b, -760, and -23a-5p [201,202]. The PI3K/AKT/mTOR pathway is also dysregulated in HB, with miRNAs-193a-5p, -495, and -206 showing oncogenic dysregulation in HB cells [203,204]. miRNAs have also shown promise as potential biomarkers with clinical importance, including: miRNAs-17, -21, -19b, -146a, -492, and -186 [205,206,207,208]. More recently, studies of miRNA variance between HB tumors and normal liver have begun to unravel some of the geographical differences observed in HB prevalence. Polymorphisms in miR-34b/c (rs4938723) have been described that alter HB susceptibility in an Eastern Chinese population of patients, adding geographical data to biomarker and prognostic knowledge [209].
Oncogenic alterations in lncRNA also contribute to HB risk and malignancy. For instance, upregulation of lncRNAs MIR205HG, neighbor of BRCA1 lncRNA2 (NBR2), taurine upregulated 1 (TUG1), HOXA-AS2, ZFAS1, and lnc01124 have been detected in HB samples, where they modify cancer cell stemness to enhance invasion and proliferation via MAPK and PI3K/AKT signaling pathways [210,211,212,213]. Similarly, upregulation of lncRNA OIP5-AS1 promotes HB invasion via the WNT/β-catenin pathway [214]. Other lncRNAs that have shown associations with increased HB malignancy and EMT are HOXA-AS2, ZFAS1, and CRNDE [214,215]. Finally, geographic polymorphisms in lncRNA-H19 that correlate to an increased risk of disease have also been identified [216].
circRNAs have shown clinical relevance in adult liver cancers, and recent studies have begun to shed light on their roles in pediatric HB [217]. circ_0015756 is upregulated in HB patient samples and cell lines, where it enhances HB proliferation and viability [218]. Similarly, circ_0000594 promotes HB malignancy via binding suppression of miRNA-217 [219]. circHMGCS1 (circ_0072391) functions as a sponge for tumor suppressor miR-503-5p, resulting in abnormal activation of the IGF/PI3K/Akt pathway and oncogenic alterations in glutamine metabolism [220]. Similarly, circRNA_CCT2 has pro-tumor properties via activation of the WNT/β-catenin pathway [221]. Several circRNAs have shown potential for halting HB progression, such as circMTO1, circEPB41L2, and circDLC1 [222,223,224]. Finally, circRNA_SORE has been linked to increased chemoresistance in HB [225].
4.5. Osteosarcoma
Osteosarcoma (OS) is the most common primary bone tumor in children and adolescents. The disease demonstrates a bimodal age distribution, occurring between 10 and 14 years and in those over 65 years of age [226,227]. OS can arise in any bone, but it most commonly affects the metaphyses of lower limb bones [228]. Patients often present with persistent localized pain and/or swelling that cannot be attributed to accident or injury [229]. Treatment is multimodal, involving surgical excision and neoadjuvant/adjuvant chemotherapy with methotrexate, adriamycin (doxorubicin), and platinum (cisplatin; MAP therapy) [230,231]. MAP implementation and surgical advances have improved overall survival to over 80%, but patients with metastatic OS, relapsed OS, or with poor chemotherapeutic response still face survival rates of 30% or below [232,233]. Therapeutic development has been significantly hampered by the fact that OS demonstrates a high degree of genetic heterogeneity, and only a few OS-associated germline mutations have been identified [234]. These include mutations in TP53, MYC proto-oncogene, RB1, isocitrate dehydrogenase 1 (IDH1), and cyclin-dependent kinase inhibitor 2A (CDKN2A) [235,236,237,238,239]. Because of this, OS-related ncRNA research has focused on identifying molecules associated with progression, therapy resistance, and for use as potential biomarkers (Table 5).
As with many of the other pediatric cancers discussed in this review, miRNAs are the most thoroughly studied OS-associated ncRNA. Studies have characterized miRNA biomarkers, oncogenic and tumor suppressive miRNAs, and elucidated many of the target genes, biological functions, and underlying molecular signaling pathways. These miRNAs have been the focus of several excellent recent reviews and will not be discussed in depth here [240,241].
lncRNAs play critical roles in the modulation of OS transcription patterns, post-transcriptional modifications, mRNA stabilization, and epigenetic regulation [242]. Downregulation of tumor suppressor lncRNA-HIF2PUT correlates with prognosis in OS cell lines and tumors via regulation of hypoxia-inducible factor 2 (HIF2) [243,244]. lncRNA-MALAT1 increases OS cell proliferation and metastasis through PI3K/Akt overactivation and the inhibition of high mobility group box 1 (HMGB1) [245]. Similarly, lncRNA-HOTAIR promotes OS pathogenesis through Akt/mTOR pathway activation, increased zinc finger E-box bonding homeobox 1 (ZEB1), and the inhibition of mineralization via bone-specific alkaline phosphatase (ALP) activity [246,247]. In OS, lncRNA-DANCR expression is correlated with disease stage, enhances stemness and improves immune evasion through the upregulation of the tyrosine kinase AXL and enhanced Musashi RNA binding protein 2 (MSI2) expression [248,249]. The lncRNA-CRNDE demonstrates diagnostic and prognostic expression patterns, and promotes metastasis, EMT, and enhances proliferation through overactivation of the WNT/β-catenin, ATP binding cassette subfamily C member 12 (ABCC12/MRP9), and NOTCH1 signaling pathways [250,251,252,253]. lncRNA-SNHG12 promotes OS metastasis, contributes to doxorubicin resistance, and demonstrates clinical trends that suggest its potential use as a diagnostic tool [254,255]. Enhanced expression of lncRNA-UCA1 correlates with prognosis and with increased invasion and tumor growth [256,257]. Finally, lncRNA-THOR promotes OS tumor growth via the overactivation of the Akt/ERK pathway and by the increased stability of SRY-box transcription factor 9 (SOX9) mRNA [258].
circRNAs have demonstrated biological functions in both adult and pediatric OS, where they alter cell–cell communication, gene and protein expression, therapy response, and the physical and biochemical composition of the TME [259]. circ-FOXM1 (hsa_circ_0025033) sponges the tumor suppressor miRNAs-320a and -320b which contributes to the proliferation and cell migration via WNT pathway activation [260]. Similarly, circRAB3IP (hsa_circRNA_0000419) promotes invasion by sponging the tumor suppressor miRNA-580-3p to enhance the expression of twist family BHLH transcription factor (TWIST1) [261]. Upregulation of circ_03955 enhances metadherin (MTDH) expression through binding inhibition of miRNA-3662 [262]. circ-LRP6 promotes invasion through the regulation of histone deacetylase 4 (HDAC4) and HMGB1 [263,264]. OS-associated upregulation of circDOCK1 (hsa_circ_0020378) enhances apoptotic resistance and chemotherapy resistance by enhancing IGF-1R, DNA methyltransferases 3A (DNMT3A), and lymphoid enhancer binding factor 1 (LEF1) expression [265,266,267]. The expression of circ_001422 correlates with stage and extent of disease (primary versus metastatic), and promotes activation of the fibroblast growth factor 2 (FGF2)/PI3K/Akt pathway and downregulation of cell cycle regulator E2F transcription factor 3 (E2F3) [268,269]. More recently, oncogenic circPVT1 (hsa_circ_0001821) has shown diagnostic promise via roles in multiple oncogenic pathways [270,271].
4.6. Ewing Sarcoma
Ewing sarcoma (ES) is a rare, aggressive bone and soft tissue cancer that arises in cells of the bone-derived mesenchymal stem cell (MSCs) lineage in children and adolescents [272,273]. Genetic analyses have uncovered ES-associated chromosomal translocations such as EWS RNA binding protein 1 (EWSR1) with friend leukemia integration 1 (FLI1; ~85% of cases) and ETS-related gene (ERG), ETS translocation variant 1 (ETV1), ETS variant gene 4 (E1AF) or FEV transcription factor (FEV), which together account for 10–15% of cases [274,275,276]. Common symptoms include fatigue, unexpected weight loss, bone pain or fracture, localized pain, or detection of a palpable mass [277]. Diagnosis of localized disease occurs in approximately 75% of cases, but 25% of cases are diagnosed after the development of bone marrow or pulmonary metastases [272]. Multimodal treatment is utilized depending on the disease stage and location, combining neoadjuvant/adjuvant chemotherapy, radiotherapy, and surgical resection [278,279]. Survival rates for patients diagnosed with localized disease are around 70%, but the survival rates significantly drop (~30%) in those diagnosed with metastatic and/or relapsed disease [280,281]. ncRNAs are being explored for their potential roles as biomarkers and for therapeutic development (Table 6).
Biological functions, mechanisms, and clinical relevance of ES-associated miRNAs have been examined in both patient tumors and cell lines. miRNAs-20b, and -34b both demonstrate correlations with ES disease progression [282]. miRNAs-30d, -125b, and -21, both overexpressed in ES, enhance the expression of Bcl-2-like protein 4 (BAX) to prevent cancer cell apoptosis [283,284,285]. miRNA-181c promotes tumor growth via suppression of tumor necrosis factor receptor super family member-6 (TNFRSF6) [286]. Enhanced expression of miRNA-130b contributes to cell cycle dysregulation by through activation of cell division cycle 42 (CDC42) [287]. Conversely, miRNA-683 has been shown to inhibit angiogenesis by targeting VEGFA mRNA for degradation [288]. The let7 family of tumor-suppressor miRNAs also plays a role in ES [289]. Let-7a forms RISCs withEWSR1 mRNA, which inhibits protein production [290]. Let-7b targets the RAS pathway to inhibit tumor cell survival and also reduces the expression of HIF1α within the TME [291]. miRNA-708-5p has been shown to inhibit pathogenic ECM remodeling by targeting matrix metallopeptidase 12 (MMP12), and miRNA-27a reduces cell proliferation and tumor growth by targeting IGF-1) [292,293].
The studies of lncRNAs in ES pathogenesis are still fairly limited and confined primarily to in vitro studies of ES cell models. However, a recent study examining RNA-seq data from patient samples detected significant upregulation in expression patterns of several lncRNAs (SNHG17, LINC00623, WAC-AS1, SSBP3-AS1, and TDRG1) that are indicative of poor overall survival rates [294]. Similarly, microarray analyses have identified ES-associated increases in lncRNAs PncCCND1_B and FOXP4-ASI expression, and have demonstrated these target cyclin D1 (CCND1) and thymopoietin (TMPO), respectively via miRNA sponging [295,296]. HOTAIR and TUG1 are upregulated in ES (as in other pediatric solid tumors discussed here) and contribute to disease progression [297,298]. lncRNA METTL3 promotes cell migration through competitive binding interactions with miRNA-124-3p, which results in elevated cyclin-dependent kinase 4 (CDK4) expression in tumor-derived cells [299]. Finally, studies of genetic variations in patient samples uncovered polymorphisms in lncRNA SENCR, associated with FLI1, and their role in the regulation of ES-associated upregulation of insulin-like growth factor 2 mRNA-binding protein 3 (IGF2BP3) [300].
The role of circRNAs in ES remains mostly unexplored. To date, only one circRNA has been studied in ES pathogenesis. circZNF609 has been shown to be associated with the EWS-FLI1 gene fusion and to promote metastasis and inhibit apoptosis in ES cell lines through sponging interaction with miRNA-145-5p [301]. Future studies into the role of circRNAs in ES will be beneficial in identifying novel targets and biomarkers to improve diagnosis and treatment.
5. Discussion
Cancer is a multifactorial disease that affects both adult and pediatric patients. Improvements in diagnostics, imaging, and therapeutic techniques have greatly improved overall survival and event free survival rates for many cancers, but recurrence and therapy resistance still comprise a significant number of cases. Towards the goal of finding novel therapeutic strategies, research has turned to determining the role of epigenetic mechanisms such as aberrant DNA methylation, histone modifications, and non-coding RNAs [302].
The term non-coding RNAs encompasses a wide range of biological molecules that have been shown to play vital roles in health and disease [303]. In the field of pediatric medicine, genome and tumor RNA-sequencing have greatly improved our knowledge of prognostic biomarkers and the mechanistic roles of miRNAs, lncRNAs, and circRNAs in the biology of many solid cancers. These findings have led to the development and incorporation of targeted therapies currently approved or in clinical trials, such as RAF and/or MEK inhibitors (dabrafenib, trametinib, and selumetinib) in the treatment of low-grade gliomas (LGGs) and some gene-mutant solid tumors to augment traditional systemic treatment [304,305]. However, while knowledge of cancer-associated ncRNA variances has greatly improved, the implementation of non-coding RNA-targeted therapies is still hampered by off-target effects, drug toxicity, and the existing knowledge gaps regarding the functions and disease/stage specificities of ncRNAs.
6. Conclusions
This review highlights current knowledge regarding the roles and underlying mechanisms of ncRNAs in many pediatric solid cancers in addition to critical knowledge gaps where the role of these molecules remains poorly understood. Also, differences in ncRNA profiles between disease subtypes, identification of alterations upon relapse, and primary versus metastatic disease remain unknown. Further research into these areas would greatly improve biomarker and therapeutic target identification to improve survival rates and patient quality of life.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bizuayehu H.M. Ahmed K.Y. Kibret G.D. Dadi A.F. Belachew S.A. Bagade T. Tegegne T.K. Venchiarutti R.L. Kibret K.T. Hailegebireal A.H. Global disparities of cancer and its projected burden in 2050 JAMA Netw. Open 20247 e 244319810.1001/jamanetworkopen.2024.4319839499513 PMC 11539015 · doi ↗ · pubmed ↗
- 2Miller K.D. Nogueira L. Devasia T. Mariotto A.B. Yabroff K.R. Jemal A. Kramer J. Siegel R.L. Cancer treatment and survivorship statistics, 2022 CA Cancer J. Clin.2022724094363573663110.3322/caac.21731 · doi ↗ · pubmed ↗
- 3Byrne S. Boyle T. Ahmed M. Lee S.H. Benyamin B. Hyppönen E. Lifestyle, genetic risk and incidence of cancer: A prospective cohort study of 13 cancer types Int. J. Epidemiol.20235281782610.1093/ije/dyac 23836651198 PMC 10244040 · doi ↗ · pubmed ↗
- 4Novikov N.M. Zolotaryova S.Y. Gautreau A.M. Denisov E.V. Mutational drivers of cancer cell migration and invasion Br. J. Cancer 202112410211410.1038/s 41416-020-01149-033204027 PMC 7784720 · doi ↗ · pubmed ↗
- 5Derks L.L. van Boxtel R. Stem cell mutations, associated cancer risk, and consequences for regenerative medicine Cell Stem Cell 2023301421143310.1016/j.stem.2023.09.00837832550 PMC 10624213 · doi ↗ · pubmed ↗
- 6La Haye S. Fitch J.R. Voytovich K.J. Herman A.C. Kelly B.J. Lammi G.E. Arbesfeld J.A. Wijeratne S. Franklin S.J. Schieffer K.M. Discovery of clinically relevant fusions in pediatric cancer BMC Genom.20212287210.1186/s 12864-021-08094-z 34863095 PMC 8642973 · doi ↗ · pubmed ↗
- 7Mella C. Tsarouhas P. Brockwell M. Ball H.C. The Role of Chronic Inflammation in Pediatric Cancer Cancers 20251715410.3390/cancers 1701015439796780 PMC 11719864 · doi ↗ · pubmed ↗
- 8Siegel D.A. King J.B. Lupo P.J. Durbin E.B. Tai E. Mills K. Van Dyne E. Buchanan Lunsford N. Henley S.J. Wilson R.J. Counts, incidence rates, and trends of pediatric cancer in the United States, 2003–2019 JNCI J. Natl. Cancer Inst.2023115133713543743307810.1093/jnci/djad 115PMC 11018256 · doi ↗ · pubmed ↗