The Extracellular Matrix Regulates Invasion in Fusion-Negative Rhabdomyosarcoma via YAP–PIEZO1 Signaling Axis
Yuanzhong Pan, Juha Kim, Brian M. Wong, Esteban Cobo Espuny, JinSeok Park

TL;DR
This study shows how the density of the extracellular matrix affects the spread of fusion-negative rhabdomyosarcoma through a signaling pathway involving YAP and PIEZO1, offering a new treatment target.
Contribution
The study identifies a novel YAP–PIEZO1 signaling axis linking extracellular matrix density to cancer invasion in fusion-negative rhabdomyosarcoma.
Findings
High extracellular matrix density activates YAP, which increases PIEZO1 expression and calcium influx to promote invasion.
Pharmacological inhibition of the YAP–PIEZO1 axis reduces invasive potential in fusion-negative rhabdomyosarcoma cells.
Elevated PIEZO1 expression correlates with poorer survival in patients with fusion-negative rhabdomyosarcoma.
Abstract
Fusion-negative rhabdomyosarcoma (FNRMS) is the prevalent subtype of rhabdomyosarcoma, the most common pediatric soft-tissue sarcoma. Its invasion often leads to recurrence and poor prognosis. This study investigates how the density of the extracellular matrix, surrounding cancer cells in the tissue, influences FNRMS invasion. We found that high extracellular matrix density activates the protein YAP, which directly triggers the expression of the mechanical sensor PIEZO1. This sensor allows calcium to enter the cells, providing a signal that facilitates invasion. Pharmacological inhibition of this axis successfully reduced the invasive potential, highlighting a novel therapeutic vulnerability for FNRMS patients. Background: Fusion-negative rhabdomyosarcoma (FNRMS) represents the most prevalent subtype of rhabdomyosarcoma, the most common pediatric soft-tissue sarcoma. Although its…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
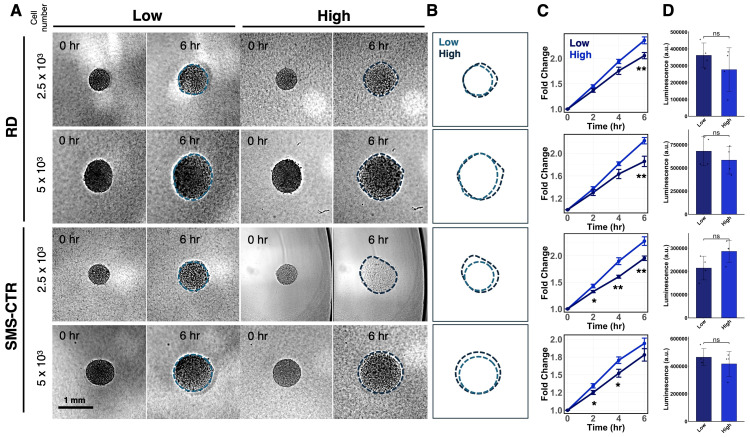 Figure 1
Figure 1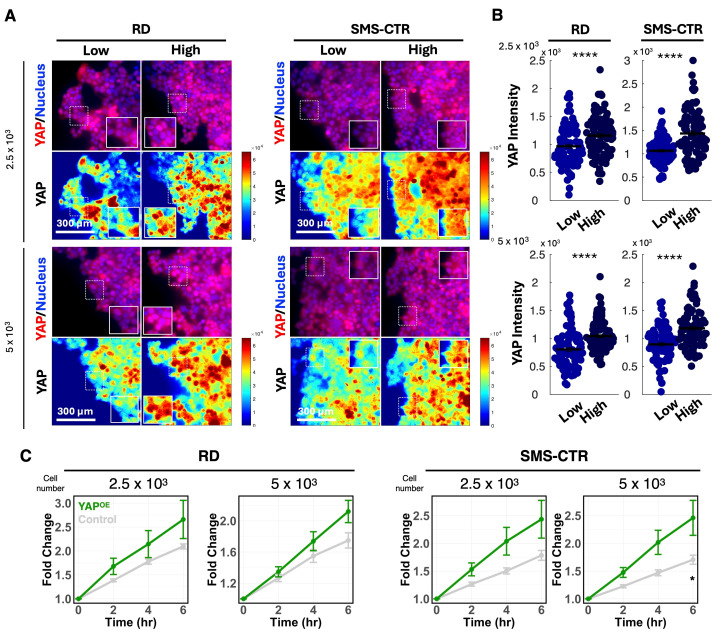 Figure 2
Figure 2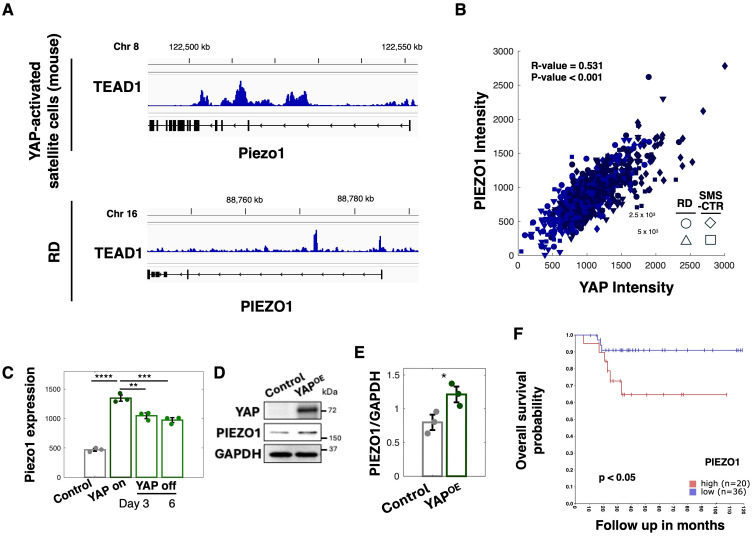 Figure 3
Figure 3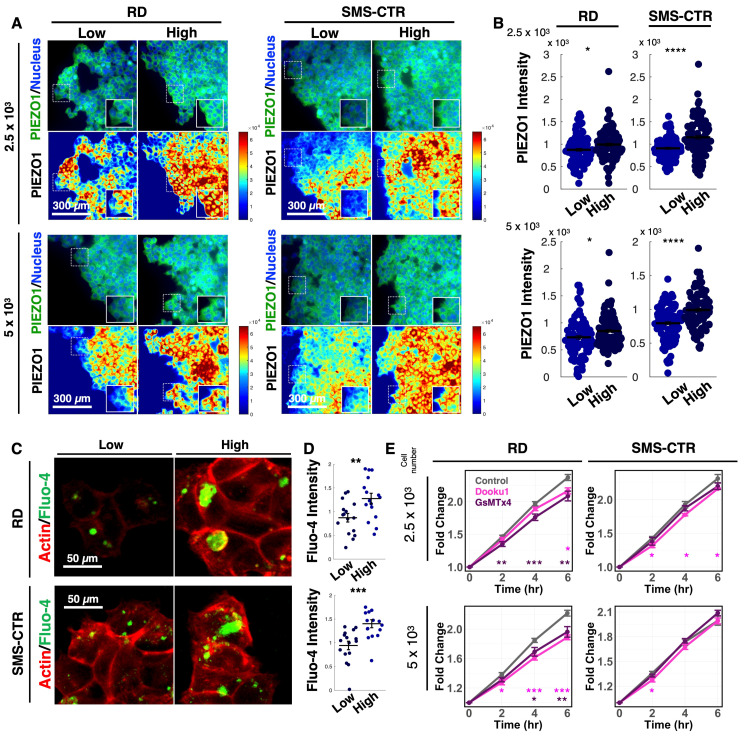 Figure 4
Figure 4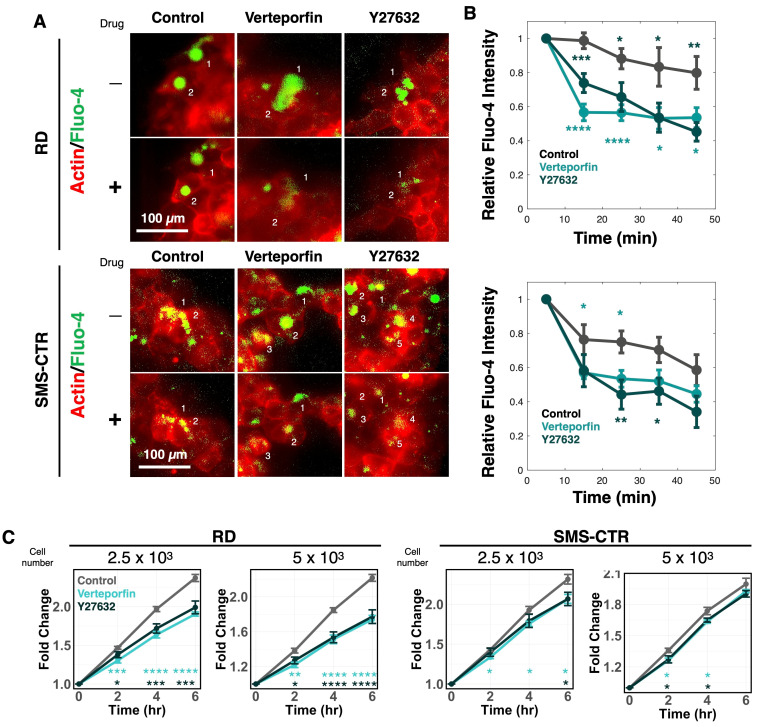 Figure 5
Figure 5- —Tower Cancer Research Foundation
- —Rally Foundation
- —Infinite Love for Kids Fighting Cancer Foundation
- —Alex’s Lemonade Stand Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHippo pathway signaling and YAP/TAZ · Cellular Mechanics and Interactions · Sarcoma Diagnosis and Treatment
1. Introduction
Rhabdomyosarcoma (RMS) is the most common soft-tissue sarcoma of childhood and adolescence, arising from skeletal muscle progenitor cells that undergo malignant transformation [1]. Despite advances in multimodal therapy, clinical outcomes remain suboptimal, particularly for patients with high-risk disease or recurrent tumors. RMS comprises two major molecular subtypes with distinct clinical behaviors: fusion-positive (FPRMS), characterized by chromosomal translocations such as PAX3–FOXO1 or PAX7–FOXO1, and fusion-negative (FNRMS), which lacks these fusions. Although FPRMS has historically been associated with higher aggressive disease, FNRMS accounts for approximately 60% of RMS cases and remains incompletely understood at the mechanistic level [2]. Consequently, a deeper understanding of the biological underpinnings of FNRMS is essential for improving therapeutic strategies for this clinically challenging subtype.
Tumor progression is governed not only by cell-intrinsic oncogenic programs but also by the biophysical and biochemical properties of the tumor microenvironment (TME) [3,4]. Among these, the extracellular matrix (ECM) plays a critical role in regulating cancer cell migration, invasion, and survival [5,6,7]. Changes in ECM composition, architecture, and stiffness are hallmarks of many solid tumors and can profoundly influence cellular behaviors through mechano-transduction pathways. While increasing evidence suggests that RMS cells respond to the ECM [8,9,10,11], how RMS senses and adapts to the ECM cues remains poorly defined. Notably, FNRMS shows greater dependence on and interaction with the ECM compared with FPRMS [8]. Given that invasion is a major clinical concern in FNRMS, and the ECM-mediated mechanobiology regulates invasion, elucidating the mechanobiology of FNRMS may uncover previously unrecognized vulnerabilities.
A key effector of mechanical signaling is the transcriptional co-activator Yes-associated protein (YAP), a central component of the Hippo pathway [12,13,14,15]. YAP shuttles between the cytoplasm and nucleus in response to increased ECM density, where it regulates gene expression programs that control proliferation, survival, and migration. YAP has been implicated in the progression of multiple cancer types. While YAP activity is highly activated in FNRMS and upregulates pro-proliferative and oncogenic genes [16], its mechanistic role in FNRMS, particularly in the context of the interaction with the TME, remains unclear. Understanding whether YAP functions as a mediator that bridges environmental ECM cues to FNRMS cell-invasive behaviors associated with clinical outcomes could provide critical insight into potential therapeutic intervention points.
In this study, we investigate how ECM density modulates the invasive behavior of FNRMS cells and test the hypothesis that YAP serves as a mechano-transduction regulator of this process. We identify PIEZO1, a mechanosensitive ion channel that facilitates calcium influx [17,18,19], as a direct downstream target of YAP. Our findings demonstrate that the ECM-YAP–PIEZO1 axis regulates calcium signaling to promote FNRMS invasion. Furthermore, we determine that repression of this regulatory axis reduces the invasive potential of FNRMS cells, identifying potential therapeutic targets for FNRMS.
2. Materials and Methods
2.1. Cell Culture
FNRMS cell lines SMS-CTR and RD were obtained from the Childhood Cancer Repository (Lubbock, TX, USA). Cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM, Gibco, Waltham, MA, USA, cat. 11965-092) supplemented with 20% Fetal Bovine Serum (FBS, Gibco, Waltham, MA, USA, cat. A5670701) and 1% penicillin/streptomycin (Gibco, Waltham, MA, USA, cat. 15140-122) in a 37 °C, 5% CO_2_ incubator.
2.2. Spheroid Formation and Collagen Invasion Assay
To generate spheroids, the liquid overlay method was employed. Adherent cell cultures were detached using Accutase (Millipore, St. Louis, MO, USA, cat. SCR005), followed by dilution with culture media to allow for the seeding of 5000 and 2500 cells per 100 µL cell culture media, which were deposited in a 96-well spheroid plate (Corning, Corning, NY, USA, cat. 4515). The cells were maintained in a 37 °C, 5% CO_2_ incubator for 24 h until compact, aggregated spheroids were formed.
For the collagen invasion assay, a stock solution of type I collagen from rat tail (Gibco cat. A10483-01) was used to prepare the collagen gel at a final concentration of 0.5% and 1.0%. In a sterile tube, for each 400 µL of final gel solution, 253 µL (187 µL for 1.0% gel) of H_2_O, 40 µL of HEPES (Sigma, St. Louis, MO, USA, cat. H0887), 40 µL of DMEM, 6.8 µL of NaOH (Thermo Scientific, Waltham, MA, USA, cat. 047124.K2), and 67 µL (134 µL for 1% gel) of collagen stock solution were combined to make the gel mixture, and 40 µL of the mixture was deposited into each well of a 96-well plate or an 8-chamber cover slide (Thermo Scientific cat. 155409). The plate was incubated at 37 °C for 20 min to allow the gel to solidify. The spheroids were then collected using a standard pipette tip with 3 mm of the end cut off and deposited onto the collagen gel.
Live-cell imaging was conducted to visualize the invasion of spheroids on a Nikon Eclipse C2 inverted microscope (Nikon Instruments, Melville, NY, USA) that is equipped with an OKO atmosphere controlling system to maintain humidity, 37 °C temperature, and 5% CO_2_. The spreading area was quantified using ImageJ (version 1.54g).
2.3. Quantification of Proliferation
CellTiter-Glo assay was used to quantify the proliferation of spheroids under different conditions. Spheroids were prepared by seeding on top of the collagen gel, as described in the previous section. Six hours after seeding, 100 µL of CellTiter-Glo 3D (Promega, Madison, WI, USA, cat. G9681) reagent was added to each well and incubated at room temperature for 25 min. Then, luminescence was measured with a Thermo Varioskan plate reader (Waltham, MA, USA).
2.4. Immunofluorescent (IF) Staining and Imaging
For IF staining, media were removed from cultured spheroids or cell monolayers, which were washed with PBS twice. Cells were fixed with 4% formaldehyde at room temperature for 20 min, then washed twice. PBS containing 0.1% Triton X-100 (Thermo Scientific, Waltham, MA, USA, cat. J66624-AE) was added to permeabilize the membrane for 45 min at room temperature. Blocking was performed with PBS containing 3% BSA (Sigma cat. A9647) for 45 min, followed by primary antibody (YAP Rabbit antibody, CST, Danver, MA, USA, cat. 14074; PIEZO1 mouse antibody, Invitrogen, Waltham, MA, USA, cat. MA5-32876; 1:200 dilution) staining for 1 h at room temperature or overnight at 4 °C. After two washes, secondary antibodies (AF594 goat anti-rabbit IgG, Invitrogen, Waltham, MA, USA, cat. A-11012; AF488 goat anti-mouse IgG, Invitrogen, Waltham, MA, USA, cat. A-10680; 1:1000 dilution) and nucleus dye (Hoechst 33342, Invitrogen, Waltham, MA, USA, cat. 62249, 1:10,000 dilution) diluted in PBS containing 1% BSA was added and incubated at room temperature in the dark for 45 min. The cells were washed three times with PBS and imaged with a Nikon fluorescent microscope.
Quantification of immunofluorescence (IF) intensities was performed using a custom MATLAB script ( version R2024a, MathWorks, Natick, MA, USA) and ImageJ. Nuclear regions were identified based on Hoechst staining and segmented using the imbinarize function in MATLAB. After selecting each nucleus in the segmented image using the bwselect function, the average YAP fluorescence intensity within each segmented nuclear region was quantified on a per-cell basis.
2.5. Sequencing Data Reanalysis
To investigate the transcriptional regulation of PIEZO1 by YAP1 and TEAD, we analyzed publicly available ChIP-seq and RNA-seq datasets. ChIP-seq data from YAP-activated satellite and RD cells were obtained from GSE55186. Additionally, RNA-seq data from doxycycline-inducible YAP1 S127A-driven FNRMS model tumors and tumors regressed following doxycycline withdrawal were downloaded from GSE47198. Processed data and genomic tracks were visualized using the Integrative Genomics Viewer (IGV).
ChIP-seq raw data from YAP-activated satellite cells and RD cells were obtained from the Gene Expression Omnibus (GEO) under accession number GSE55186. Sequencing reads were aligned to the reference genome (mm10 for mouse satellite cells and hg19 for human RD cells). Peak calling was performed using MACS2 with default parameters. The resulting processed genomic tracks, represented as fold-enrichment over input, were visualized at the Piezo1/PIEZO1 locus using the Integrative Genomics Viewer (IGV) to identify TEAD1 binding sites.
Transcriptomic changes associated with YAP1 activity were analyzed using RNA-seq data from a doxycycline (Dox)-inducible YAP1 S127A-driven FNRMS mouse model (GSE47198). Normalized expression values for Piezo1 were extracted from the following conditions: control (Dox−), YAP1-on (Dox+), and YAP1-off (following Dox withdrawal for 3 or 6 days to induce tumor regression).
2.6. YAP Overexpression
YAP overexpression was performed by expressing the YAP-expressing plasmid (mEGFP-N1-YAP, addgene, Watertown, MA, USA, cat. 166457) using the Lipofectamine 3000 Transfection kit (Invitrogen, Waltham, MA, USA, cat. L3000-015) following the manufacturer’s protocol. Briefly, for each transfection reaction in one well of a 6-well plate, 7 µL was added to 125 µL OptiMEM (Gibco, Waltham, MA, USA, cat. 51985034) followed by vortexing for 3 s. In a separate 1.7 mL tube, 5 µg of plasmid DNA was added to 125 µL OptiMEM, then 10 µL of P3000 to the mixture. The Lipofectamine solution was then added to the DNA solution, mixed by pipetting, and incubated at room temperature for 10 min. A total of 250 µL of the transfection reaction mixture was then added dropwise to cultured cells that were over 90% confluent. For spheroid experiments, cells were harvested 24 h after transfection, and spheroids were made as described in the previous section. For the Western blot, cells were harvested 36 h after transfection.
2.7. Western Blotting
For the Western blot, cells were harvested with Accutase (Millipore, St. Louis, MO, USA, cat. SCR005) after 36 h of transfection, washed twice with PBS, and lysed with RIPA buffer (Thermo Scientific, Waltham, MA, USA, cat. 89901) containing 1× protease and phosphate inhibitor cocktail (Thermo Scientific, Waltham, MA, USA, cat. 78440). Cell lysates were centrifuged at 20,000× g for 15 min at 4 °C to pellet the insoluble portion of the lysate, and only the supernatant was kept. Protein concentration was quantified via BCA assay using the Pierce BCA kit (Thermo Scientific cat. 23227) and a Varioskan plate reader, and all samples were normalized to the same concentration.
In total, 5 µg of protein was loaded on a 15-well gel (BioRad, Hercules, CA, USA, cat. 4561096), and electrophoresis was run in 1× Tris/Glycine/SDS buffer (diluted from 10× stock, BioRad cat. 1610772) at 50 V for 3 h at room temperature. Semidry transfer was performed on a BioRad Trans-Blot Turbo transfer system at 25 V for 10 min. The membrane was then cut and incubated in 3% BAS in Tris-buffered saline containing 1% Tween-20 (TBST) solution at room temperature for 1 h to block unspecific binding. Primary antibodies (YAP Rabbit antibody, CST, Danvers, MA, USA, cat. 14074; PIEZO1 mouse antibody, Invitrogen, Waltham, MA, USA, cat. MA5-32876; GAPDH mouse antibody, CST cat. 97116S) were added to the membrane and incubated on a rocker at 4 °C overnight. After three washes with TBST, the secondary antibody (HRP-Goat pAb to Rabbit IgG, abcam, Waltham, MA, USA, cat. AB205718; HRP-Goat anti-mouse IgG, Invitrogen, Waltham, MA, USA, cat. 62-6520) was added and incubated at room temperature for 1 hour. The Western blot substrate (BioRad, Hercules, CA, USA, cat. 170-5061) was added, and membranes were imaged on an Invitrogen iBright imaging system (Waltham, MA, USA).
2.8. Survival Analysis
Survival analysis was performed using the R2: Genomics Analysis and Visualization Platform (https://r2.amc.nl, accessed on 20 December 2025) with the Tumor Rhabdomyosarcoma—Williamson dataset. The analysis was restricted to the chromosomal translocation-negative subgroup. Gene expression values were obtained from the microarray platform ps_avgpres_emtab1202ebi101_u133p2.
Patients were stratified into high and low PIEZO1 expression groups using the auto-selected cut-off option in R2, which determines the optimal expression threshold that maximizes separation between survival curves. Overall survival was used as the clinical endpoint. Kaplan–Meier survival curves were generated for the two groups, and differences in survival distributions were assessed using the two-sided log-rank (Mantel–Cox) test.
2.9. Calcium Influx Imaging
For calcium influx imaging, cells were seeded on 8-chamber cover slides to achieve 30–70% confluency on the day of assay. For each chamber, 500 µL culture media containing 2 µL Fluo-4 AM (abcam, Waltham, MA, USA, cat. Ab241082) and 1 µL SPY555-Actin (Cytoskeleton, Denver, CO, USA, cat. CY-SC202) was added to cultured cells and incubated for 1 h. If drug treatment was performed at this step, drugs were added and diluted to working concentration in the media prior to the addition of dyes. After incubation, the staining solution was discarded, cells were washed twice with PBS, and 500 uL of culture media without pheno-red (DMEM containing 10% FBS and 1% penicillin/streptomycin) was added to each well and imaged on a Nikon inverted fluorescent microscope. Calcium influx intensities were assessed using the MATLAB custom-made algorithm.
2.10. Drug Treatment
All small-molecule drugs used in this study were purchased from Selleckchem.com (Houston, TX, USA). For the spheroid invasion assay, 5 µM Dooku1 (cat. 2953) and 5 µM GsMTx4 (cat. P1205) were added to the culture media when the spheroids were transferred onto collagen gel. For calcium influx imaging, 100 nM verteporfin (cat. S1786) and 5 µM Y-27632 (cat. S6390) were added and diluted to working concentration in the culture media before actin and calcium dye were added.
2.11. AI Disclosure
During the preparation of this manuscript, Gemini (Google) was used for proofreading to improve language and readability. After using this tool, the authors reviewed and edited the content as needed, and they take full responsibility for the scientific integrity and final version of the manuscript.
3. Results
3.1. FNRMS Cells Exhibit Enhanced Invasive Behavior on High ECM Density
Multiple cancer types exhibit increased ECM density in high-risk or metastatic subgroups compared with low-risk or local counterparts [20,21,22]. Furthermore, elevated ECM density has been shown to promote tumor cell invasion [23,24]. To determine whether ECM density similarly influences the migratory behavior of FNRMS cells, we performed a three-dimensional (3D) spheroid invasion assay using collagen type I matrices at varying concentrations. Spheroids of uniform sizes were generated from two FNRMS cell lines, RD and SMS-CTR, and seeded onto collagen gels at low (0.5%) or high (1.0%) concentrations, hereafter referred to as low- and high-density matrices, respectively.
Live-cell imaging revealed that spheroids from both cell lines exhibited greater radial expansion, reflecting invasion into the surrounding matrix, when cultured on high-density collagen gels compared with low-density conditions (Figure 1A,B). This enhanced outgrowth was consistent across spheroids of different initial sizes, indicating that the effect was independent of initial spheroid size. Quantitative analysis of spheroid spreading area over multiple time points demonstrated a consistently larger invaded area on high-density matrices for all conditions tested (Figure 1C). We also performed the same assay on RH41, an FPRMS cell line that is less dependent on the ECM cues and, thus, serves as a control. The results showed that the outgrowth of RH41 spheroids did not differ across different concentrations (Supplementary Figure S1A). To exclude the potential contribution of cell proliferation to the outgrowth of spheroids, we quantified proliferation 6 h after seeding and observed no significant difference in cell proliferation (Figure 1D).
Collectively, these results indicate that increased collagen matrix density enhances the invasive potential of FNRMS cells in a 3D context.
3.2. YAP Mediates Matrix-Dependent Invasion of FNRMS Cells
Cancer cell invasion is regulated by mechano-transduction signaling pathways that enable cells to sense and respond to the physical properties of the ECM [25,26,27]. The transcriptional co-activator YAP is a central effector of mechano-transduction and has been implicated in ECM density-dependent regulation of cell invasion in multiple cancer types. We therefore investigated whether YAP mediates the enhanced invasive behavior of FNRMS cells observed on high-density collagen matrices. To assess YAP activity in response to matrix properties, we performed immunofluorescence staining for YAP in FNRMS spheroids cultured on low- or high-density collagen gels.
YAP activity is regulated by its subcellular localization: upon activation, YAP translocates to the nucleus, where it functions as a transcriptional co-activator. Thus, we quantified nuclear YAP signal intensity in cells located near the invasive front of spheroids. Both RD and SMS-CTR spheroids, across all sizes examined, exhibited significantly increased nuclear YAP accumulation when cultured on high-density collagen matrices compared with low-density conditions (Figure 2A,B). These data indicate that increased matrix density elevates YAP activity in invading FNRMS cells.
To determine whether YAP activation is sufficient to promote invasion, we generated YAP-overexpressing (YAP-OE) FNRMS cells and assembled spheroids for invasion assays. Quantitative analysis of spheroid spreading area over time revealed that YAP overexpression significantly increased the invasive spreading relative to control spheroids (Figure 2C), and the contribution of proliferation was excluded (Supplementary Figure S1B). This effect phenocopied the enhanced invasion observed on high-density collagen matrices, supporting a functional role for YAP in driving ECM-dependent invasion. Together, these results demonstrate that YAP activity is elevated in response to increased matrix density and that YAP is sufficient to promote invasive behavior in FNRMS spheroids, identifying YAP as a key mediator of ECM-dependent invasion in this disease context.
3.3. PIEZO1 Functions as a Downstream Effector of YAP Signaling in FNRMS Cells
Because YAP primarily functions as a transcriptional co-activator, we next sought to identify downstream effectors through which YAP regulates ECM density-dependent invasion in FNRMS. In particular, we focused on genes involved in mechano-transduction that could link ECM density to intracellular signaling programs.
PIEZO1 is a mechanosensitive calcium-permeable ion channel that transduces mechanical cues from the extracellular environment into intracellular calcium signaling [17,18,19]. Given that calcium signaling promotes cancer cell invasion [28,29,30,31], PIEZO1-mediated calcium signaling has been implicated in cancer cell invasion. Tremblay et al. demonstrated that YAP activation drives oncogenic transformation of satellite cells, which exhibit hallmark features of FNRMS [16]. To determine whether PIEZO1 is a transcriptional target of YAP-TEAD1 (the primary DNA-binding partner of YAP [16]) in FNRMS, we reanalyzed publicly available datasets of chromatin immunoprecipitation sequencing (ChIP-seq) for TEAD1 of mouse satellite cells transformed via YAP activation and RD cells, both of which serve as robust models for FNRMS. Our analysis revealed prominent YAP-TEAD1 occupancy within the proximal promoter region of the PIEZO1 gene in both mouse and human FNRMS cells (Figure 3A). These findings suggest that YAP directly regulates PIEZO1 transcription in FNRMS.
Consistent with this genomic observation, quantitative analysis of immunofluorescence imaging data demonstrated a strong positive linear correlation between nuclear YAP intensity and PIEZO1 expression at the single-cell level (Figure 3B). To further determine whether YAP activity regulates PIEZO1 expression, we analyzed RNA-seq data from FNRMS cells using a doxycycline-inducible system to express a constitutively active YAP mutant (YAP-ON). Induction of YAP signaling resulted in a significant upregulation of PIEZO1 transcript levels compared with non-induced control cells (Figure 3C). Notably, this upregulation was reversible, as PIEZO1 expression returned to baseline levels following the withdrawal of the induction (YAP-OFF).
We next validated these findings at the protein level. Western blot analysis revealed elevated PIEZO1 expression in YAP-OE FNRMS cells relative to controls (Figure 3D). Quantification across multiple independent experiments confirmed a statistically significant increase in PIEZO1 abundance upon YAP overexpression (Figure 3E). Together, these data establish that PIEZO1 is a downstream target of YAP-mediated transcriptional regulation in FNRMS cells.
Finally, analysis of clinical FNRMS patient datasets revealed that elevated PIEZO1 expression is significantly associated with poorer overall survival (Figure 3F), underscoring the clinical relevance of PIEZO1 in FNRMS progression. These findings suggest that the ECM-YAP–PIEZO1 axis contributes to the invasive and aggressive behavior of FNRMS tumors and warrants further investigation.
3.4. PIEZO1-Dependent Calcium Signaling Promotes Invasion of FNRMS Cells on High-Density Matrices
To determine whether PIEZO1 functionally contributes to ECM density-dependent invasion, we first examined its expression in invading FNRMS spheroids. Immunofluorescence co-staining of PIEZO1 along with YAP revealed significantly elevated PIEZO1 signal intensity in cells within spheroids cultured on high-density collagen I matrices compared with low-density conditions (Figure 4A,B), consistent with increased YAP activity under these conditions. Because PIEZO1 functions as a mechanosensitive, calcium-permeable ion channel, we next assessed whether ECM density modulates intracellular calcium signaling. Live-cell calcium flux imaging using the Fluo-4 AM dye was performed on FNRMS spheroids cultured on low- or high-density collagen I gels (Figure 4C). Quantitative analysis demonstrated a significantly higher calcium influx in spheroids cultured on high-density matrices (Figure 4D), indicating that increased matrix concentration enhances calcium signaling in FNRMS cells.
To evaluate the functional contribution of PIEZO1-mediated calcium signaling to spheroid invasion, we pharmacologically inhibited PIEZO1 by the treatment of Dooku1 (PIEZO1 antagonist) [32] and GsMTx4 (a mechanical channel blocker) [33]. PIEZO1 inhibition resulted in a significant reduction in spheroid spreading area compared with vehicle-treated controls (Figure 4E), and the contribution of proliferation was excluded (Supplementary Figure S1C). Collectively, these data indicate that increased ECM density correlates with elevated PIEZO1 expression and enhanced calcium signaling, which in turn facilitates the invasive behavior of FNRMS cells. These findings support a model in which PIEZO1-mediated calcium signaling acts as a key effector downstream of YAP to drive ECM density-dependent invasion.
3.5. Pharmacological Inhibition of YAP Signaling Attenuates Calcium Influx and Spheroid
To evaluate the potential therapeutic relevance of the mechano-transduction pathways identified in this study—specifically the YAP–PIEZO1 signaling axis—we tested whether pharmacological inhibition of YAP-associated signaling could suppress the elevated calcium influx and invasive behavior observed in FNRMS spheroids cultured in high-density collagen matrices. Verteporfin has been utilized to disrupt the interaction between YAP and TEAD transcription factors, thereby inhibiting YAP-dependent transcriptional activity [34]. Additionally, we employed Y-27632, a potent inhibitor of Rho-associated protein kinase (ROCK), which reduces YAP nuclear localization by modulating cytoskeletal tension in response to the ECM density [35,36].
Live-cell imaging revealed that treatment with either verteporfin or Y-27632 significantly reduced intracellular calcium influx in FNRMS cells compared with vehicle-treated controls (Figure 5A,B). Consistent with these results, quantitative analysis of spheroid invasion assays demonstrated that both inhibitors significantly attenuated the spheroid spreading area over time (Figure 5C), and the contribution of proliferation was excluded (Supplementary Figure S1D). Although invasion was not completely abolished, pharmacological inhibition of the YAP signaling axis markedly suppressed the ECM density-enhanced invasive phenotype. Collectively, these results demonstrate that targeting the YAP–PIEZO1–calcium axis can effectively reduce the invasive capacity of FNRMS cells, highlighting its potential as a therapeutic vulnerability in the context of high ECM density.
4. Discussion
In this study, we demonstrate that ECM density enhances the invasive behavior of FNRMS cells and identify a mechano-transduction pathway linking matrix density to tumor cell invasion, the ECM-YAP-PIEZO1 signaling axis. Using 3D spheroid invasion models, we show that high-density collagen matrices promote increased spheroid spreading, indicating elevated invasive potential. The observed cellular outgrowth in our model reflects a biologically relevant phenotype where tumor spheroids coordinate their movement along the ECM surface to initiate invasive spreading and matrix remodeling [2,27]. This localized migratory behavior at the tumor–ECM interface serves as a necessary precursor to 3D invasion, providing insights into the early-stage interactions between cancer cell clusters and their surrounding microenvironment. These findings highlight the critical role of ECM properties in regulating FNRMS cell behavior and underscore the necessity of considering microenvironmental cues to fully understand disease progression.
Notably, the enhanced cellular outgrowth on high-density ECM is driven by invasive migration, rather than cell proliferation, which remained consistent across the densities during the 6 h observation period. Furthermore, collagen concentrations employed (0.5% and 1.0% (w/v)) are expected to exhibit bulk elastic moduli in the ranges of ~0.5–5 kPa [37], which partially overlap with reported stiffness ranges of soft tissue where FNRMS arises [38].
Mechanistically, we identify YAP as a key mediator of ECM density-dependent invasion in FNRMS. Increased matrix density was associated with elevated nuclear YAP localization in invading cells, and YAP overexpression was sufficient to enhance spheroid invasion. These observations are consistent with YAP’s established role as a central effector of mechano-transduction pathway and extend its functional relevance to FNRMS, a molecular subtype that is more prevalent than fusion-positive disease yet remains mechanistically understudied. Our data suggest that YAP integrates mechanical signals from the ECM to drive transcriptional programs that promote invasive behavior in this rhabdomyosarcoma subtype.
We further identify the mechanosensitive calcium channel PIEZO1 as a downstream effector of YAP signaling in FNRMS cells. ChIP-seq reanalysis, transcriptomic profiling, and protein-level validation collectively establish PIEZO1 as a YAP-regulated gene. Elevated PIEZO1 expression correlated with increased YAP activity and was significantly associated with poorer clinical outcomes in FNRMS patients, underscoring potential relevance in FNRMS. Functional analyses demonstrated that high-density matrices increase PIEZO1 expression and calcium influx in spheroids, directly linking mechanical cues to intracellular calcium signaling. Furthermore, pharmacological inhibition of PIEZO1 caused significant reductions in invasion. These data indicate that PIEZO1 positively contributes to ECM density-dependent invasive behavior, consistent with the involvement of parallel or compensatory mechano-transduction pathways.
Importantly, pharmacological inhibition of YAP-associated signaling attenuated both calcium influx and spheroid invasion, supporting the central role of YAP in coordinating mechanical signaling outputs in FNRMS. Although the inhibitors used in this study are not clinically optimized for sarcoma treatment, these experiments provide proof-of-concept that targeting YAP-centered mechano-transduction pathways can suppress FNRMS invasion.
5. Conclusions
Together, our findings support a model in which ECM density promotes YAP activation, leading to transcriptional upregulation of mechanosensitive effector PIEZO1 and enhanced calcium signaling, ultimately facilitating FNRMS cell invasion. Overall, this work reveals a previously unappreciated YAP–PIEZO1 signaling axis that links ECM mechanics to invasive behavior in FNRMS. These findings advance our understanding of FNRMS mechanobiology and suggest that targeting mechano-transduction pathways may represent a complementary therapeutic strategy to limit invasion in this aggressive pediatric cancer.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Skapek S.X. Ferrari A. Gupta A.A. Lupo P.J. Butler E. Shipley J. Barr F.G. Hawkins D.S. Rhabdomyosarcoma Nat. Rev. Dis. Primers 20195110.1038/s 41572-018-0051-230617281 PMC 7456566 · doi ↗ · pubmed ↗
- 2Breneman J.C. Lyden E. Pappo A.S. Link M.P. Anderson J.R. Parham D.M. Qualman S.J. Wharam M.D. Donaldson S.S. Maurer H.M. Prognostic factors and clinical outcomes in children and adolescents with metastatic rhabdomyosarcoma —A report from the Intergroup Rhabdomyosarcoma Study IVJ. Clin. Oncol.200321788410.1200/JCO.2003.06.12912506174 · doi ↗ · pubmed ↗
- 3Du Fort C.C. Paszek M.J. Weaver V.M. Balancing forces: Architectural control of mechanotransduction Nat. Rev. Mol. Cell Biol.20111230831910.1038/nrm 311221508987 PMC 3564968 · doi ↗ · pubmed ↗
- 4Swartz M.A. Iida N. Roberts E.W. Sangaletti S. Wong M.H. Yull F.E. Coussens L.M. De Clerck Y.A. Tumor microenvironment complexity: Emerging roles in cancer therapy Cancer Res.2012722473248010.1158/0008-5472.CAN-12-012222414581 PMC 3653596 · doi ↗ · pubmed ↗
- 5Jiang Y. Zhang H. Wang J. Liu Y. Luo T. Hua H. Targeting extracellular matrix stiffness and mechanotransducers to improve cancer therapy J. Hematol. Oncol.2022153410.1186/s 13045-022-01252-035331296 PMC 8943941 · doi ↗ · pubmed ↗
- 6Huang J. Zhang L. Wan D. Zhou L. Zheng S. Lin S. Qiao Y. Extracellular matrix and its therapeutic potential for cancer treatment Signal Transduct. Target. Ther.2021615310.1038/s 41392-021-00544-033888679 PMC 8062524 · doi ↗ · pubmed ↗
- 7Henke E. Nandigama R. Ergun S. Extracellular matrix in the tumor microenvironment and its impact on cancer therapy Front. Mol. Biosci.2019616010.3389/fmolb.2019.0016032118030 PMC 7025524 · doi ↗ · pubmed ↗
- 8Chronopoulos A. Chavez I. Vemula C.K. Mittal N. Zamloot V. Pan Y. Han S.J. Park J. The PAX 3-FOXO 1 fusion gene reduces cell-ECM interactions and TG Fbeta signaling in rhabdomyosarcoma J. Cell Biol.2025224 e 20240815510.1083/jcb.20240815540586712 PMC 12207912 · doi ↗ · pubmed ↗