Short-Term Feeding Disruption Effects and Efficacy of Six Biopesticides Against Empoasca onukii (Hemiptera: Cicadellidae)
Zhifei Jia, Chunling Yang, Yilan Liu, Yilin Yang, Rui Zhou, Zhenzhen Cheng, Shubao Geng, Yongyu Xu, Zhenzhen Chen, Li Qiao

TL;DR
This study examines how six biopesticides affect the feeding behavior of tea leafhoppers and confirms their effectiveness in controlling the pest over 21 days.
Contribution
The study reveals distinct short-term feeding disruption patterns caused by six biopesticides against tea leafhoppers.
Findings
Plant-based and fungal-based biopesticides increased non-feeding time and reduced active feeding.
Bacteria-based pesticides extended resting periods and decreased passive feeding, while a virus combined both effects.
All tested biopesticides achieved 100% pest control in field trials within 21 days.
Abstract
The tea leafhopper is a major pest that damages tea plants by sucking sap, prompting the use of biopesticides as an eco-friendly alternative to chemical pesticides. However, past studies focused mostly on how these agents kill pests, leaving their short-term effects on feeding behavior unclear. This study tested six biopesticides, using a precise method to monitor feeding over 6 h, and conducted field trials to evaluate their effectiveness. Results showed that different biopesticides disrupted feeding in distinct ways: plant-based and fungal-based biopesticides spent increased time not feeding and reduced active feeding. Bacteria-based pesticides extended resting periods and decreased passive feeding, while a virus combined both effects. Field trials confirmed that all treatments fully controlled the pest within 21 days, performing consistently across various concentrations. These…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
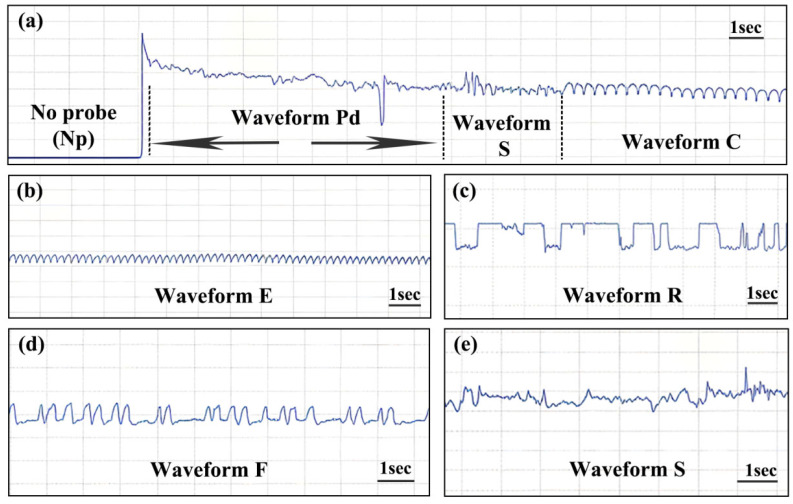 Figure 1
Figure 1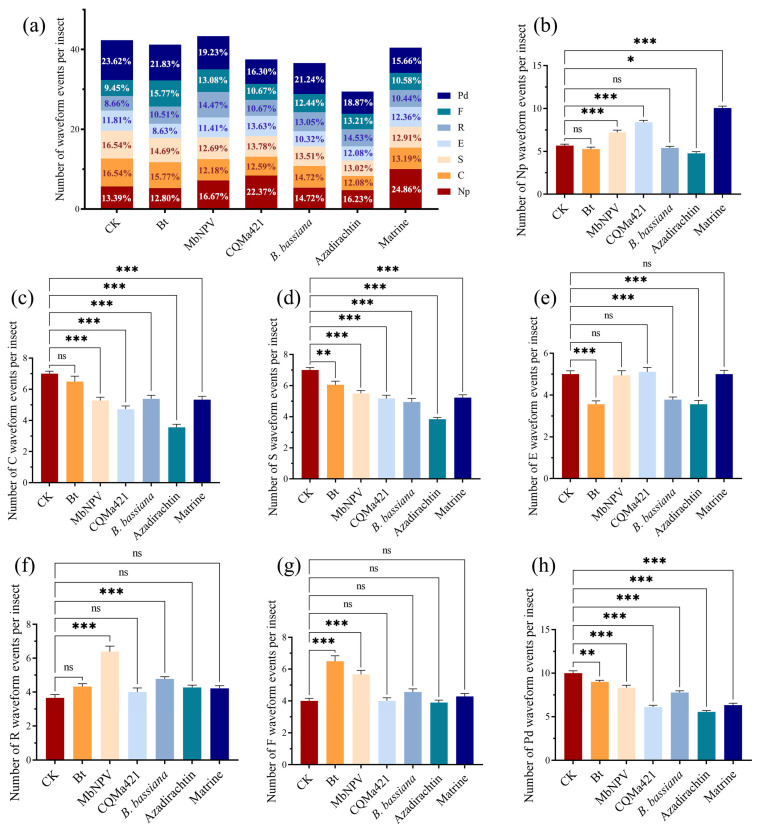 Figure 2
Figure 2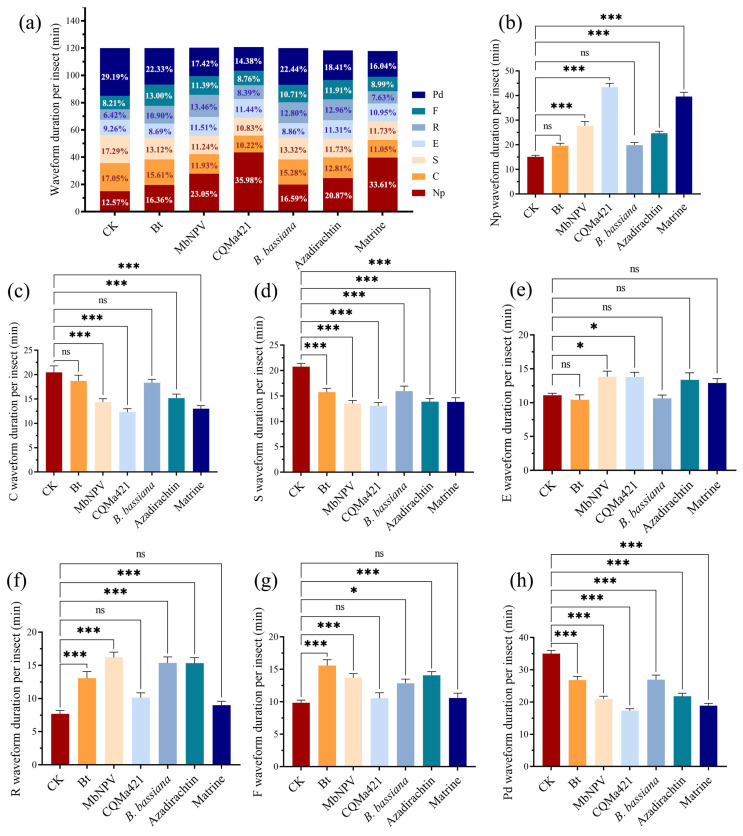 Figure 3
Figure 3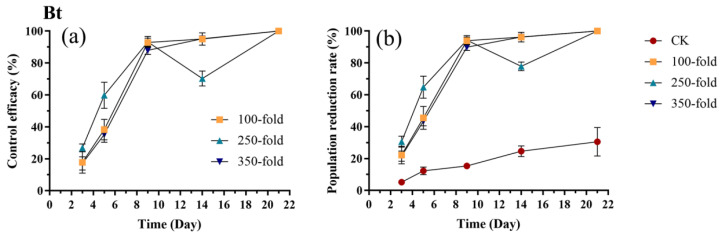 Figure 4
Figure 4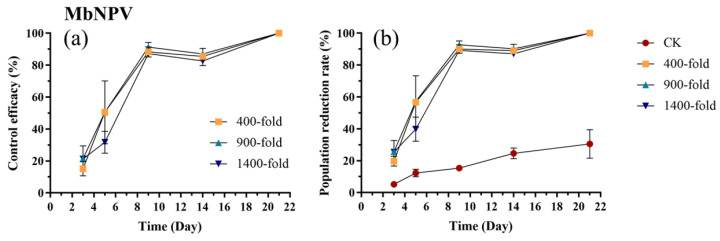 Figure 5
Figure 5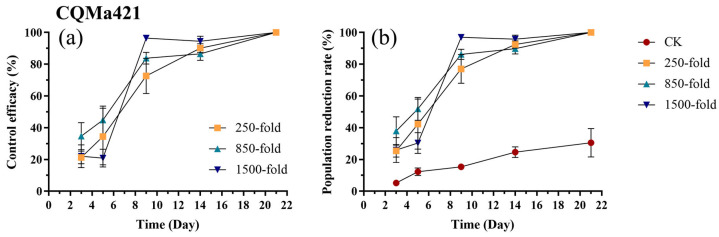 Figure 6
Figure 6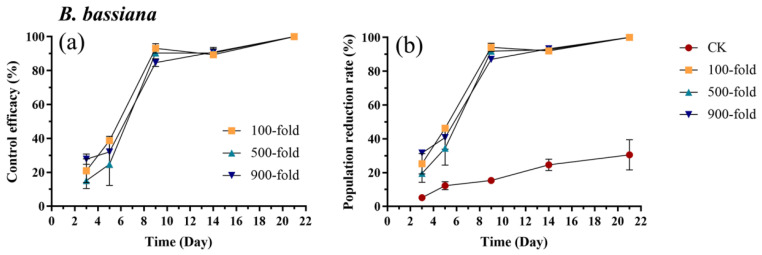 Figure 7
Figure 7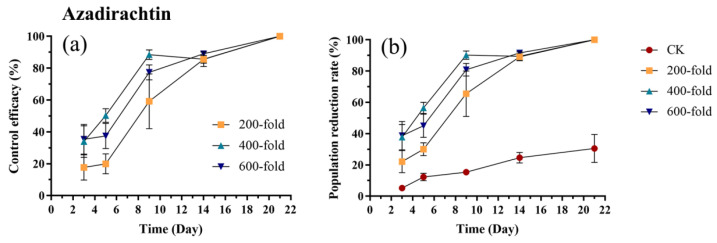 Figure 8
Figure 8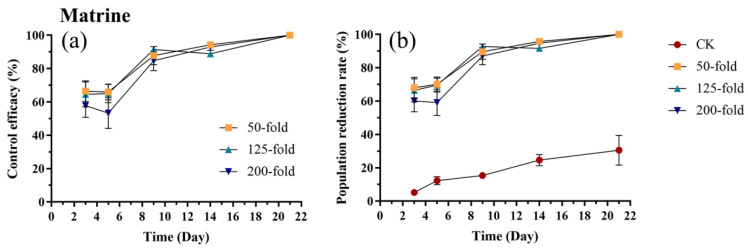 Figure 9
Figure 9- —Key Research and Development Program of Henan Province
- —University-Enterprise Collaborative Innovation Project of Henan Province
- —Modern Tea Industry Technology System of Shandong Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhytoplasmas and Hemiptera pathogens · Viral Infectious Diseases and Gene Expression in Insects · Insect-Plant Interactions and Control
1. Introduction
Tea plant [Camellia sinensis (L.) O. Kuntze] is a perennial plantation crop that requires a warm and humid climate for optimal growth and production. Such climatic conditions also favor pests and diseases that infest tea plants [1,2]. Empoasca onukii Matsuda is recognized as one of the four most damaging piercing-sucking pests, alongside Toxoptera aurantii, Apolygus lucorum, and Aleurocanthus spiniferus, among the major insect pests affecting tea plantations worldwide, and it occurs in tea-producing countries across Asia such as China, Vietnam, Japan, and Indonesia [3,4,5]. Female adults lay eggs by inserting their ovipositors into the tender shoots of tea plants, impeding the transport of nutrients. Both nymphs and adults cause damage by sucking sap from young buds, leaves, and stems. Lightly infested tea plants show chlorosis and premature aging of buds and leaves, severely affecting both yield and quality [6,7,8]. Currently, controlling E. onukii in production relies primarily on chemical pesticides such as bifenthrin, chlorfenapyr, and thiamethoxam, which readily lead to the development of pesticide resistance in E. onukii and even raise concerns over pesticide residues [9,10]. Non-chemical and sustainable pest management strategies are increasingly viewed as transformative for plant protection in the tea industry [10,11]. They have been incorporated into integrated pest management (IPM) programs for tea pests, including entomopathogenic bacteria, fungi, nematodes, and certain viruses, which are highly efficient, target-specific, and safe for soil, plants, and the environment [12,13,14,15]. Furthermore, pest management using biological control agents can also promote the proliferation of beneficial microbial communities in the soil, reduce soil-borne pathogens, and protect microbial diversity [16].
In addition to synthetic chemical pesticides, a wide range of biologically based alternatives have been developed and registered for pest management in tea plantations. Among these, botanical pesticides represent a major category of commercial biopesticides. Neem-based products are representative examples, and other active substances such as nicotine, rotenone, matrine, azadirachtin, veratrine, toosendanin, veratridine, limonin, eucalyptol, and citronella oil are all registered for use [10]. Entomopathogens are promising biocontrol agents against tea pests [10]. Bacillus thuringiensis (Bt), Metarhizium anisopliae, and Beauveria bassiana have shown significant efficacy in controlling tea pests [17]. Bt is the predominant bacterial species isolated from the cadavers of tea pests and the tea garden ecosystem. Various Bt strains have been applied in China, Japan, India, Sri Lanka, Bangladesh, and Vietnam to control lepidopteran pests in tea plantations [18]. Nucleopolyhedroviruses (NPVs) have reached commercial scale as biocontrol agents in tea plantations due to their safety and high efficacy. For instance, a Biston suppressaria NPV variant can infect the geometrid pests Hyposidra infixaria and Hyposidra talaca, which are newly recorded in India [19]. The Ectropis obliqua nucleopolyhedrovirus QF4 strain isolated in China effectively infects E. obliqua and Ectropis grisescens [20]. The Mamestra brassicae nucleopolyhedrovirus developed in China demonstrates good control efficacy against various lepidopteran pests, including E. grisescens and Spodoptera frugiperda [21,22,23]. M. anisopliae sensu lato (s. l.) has shown potential efficacy against a wide range of insects on various crops, including tea [1,24]. For example, under field conditions, a formulation of M. anisopliae s. l. demonstrated high activity against several pests on tea plants in Kenya [25]. The M. anisopliae strain CQMa421, developed in China, can infect over 80 species (or types) of agricultural, forestry, and public health pests across seven orders, including Lepidoptera, Coleoptera, Orthoptera, Hemiptera, Diptera, Hymenoptera, and Thysanoptera, at various developmental stages [26].
Biopesticides often function by interfering with and inhibiting insect feeding behavior, a critical lethal or sublethal mechanism of action [27,28]. Specifically, they can directly reduce nutrient intake and lead to starvation by suppressing feeding, damaging feeding-related physiological structures, or blocking the neuromuscular functions required for feeding. This effect synergizes with their primary toxicological actions, thereby increasing insect mortality, reducing crop damage, and potentially slowing the development of pest resistance [29,30]. It has been reported that azadirachtin acts as an antifeedant against tea pests such as Helopeltis theivora, Scirtothrips dorsalis, and Empoasca flavescens, while matrine functions primarily as a toxicant [31,32,33,34,35]. The core mechanism of Bt is stomach action [36,37]. In contrast, M. anisopliae and B. bassiana infect pests through contact, a mode of action which requires successful penetration of the cuticle and suppression of the host’s immune system [38,39]. A unique additional mechanism of B. bassiana involves secreting antimicrobial peptides that specifically inhibit Gram-positive bacteria on the insect’s body surface. This reduces interference from beneficial surface microbiota during fungal infection and significantly enhances infection efficiency [40]. However, few studies have reported the effects of infection by the aforementioned biopesticides on short-term (6–8 h) pest feeding behavior. Investigating short-term behavioral responses holds significant scientific value for the development and evaluation of biopesticides. Compared to traditional toxicity tests that rely on mortality rates, monitoring short-term behavior can reveal the sublethal effects and modes of action of pesticides earlier and more sensitively, while the observations are also directly relevant to field control efficacy [41]. The EPG technique, owing to its high temporal resolution and capacity to discriminate specific probing phases, constitutes an ideal tool for observing short-term feeding behavior [7]. By linking early behavioral disruption quantified via EPG with subsequent field efficacy data, a direct bridge can be established between laboratory-based mechanistic insights and practical pest management outcomes under field conditions.
This study utilized the electrical penetration graph (EPG) technique to analyze differences in the feeding behavior of E. onukii after treatment with different biopesticides (two entomopathogenic fungi, one virus, one bacterium, and two botanical pesticides) and integrated these behavioral findings with field efficacy trials to comprehensively evaluate control effects. It aims to investigate the early effects of biopesticides on pest feeding behavior, thereby providing a theoretical basis for the targeted design and optimization of pesticides.
2. Materials and Methods
2.1. Test Insects
The tea green leafhoppers and tea plants used in this study were collected from a tea plantation in Xiajiachong Village, Shihegang Town, Xinyang, China (32°18′ N, 114°35′ E). The insects were maintained in rearing cages (60 × 60 × 60 cm) within a controlled-environment insectary (24 ± 2 °C, 70–80% relative humidity, 16:8 h light:dark cycle) and fed with fresh tea shoots. Adults 5–7 days post-emergence were selected as experimental subjects. Preliminary analyses indicated no consistent sex-based differences in feeding behavior under the tested conditions. Therefore, test insects were used in random sex combinations. Each individual was used only once. The insects used in the EPG experiments and field trials were completely separate populations. No individuals were reused across the two types of experiments.
2.2. Test Biopesticides
The biopesticides used in this study are as follows: 0.3% azadirachtin EC (Chengdu Green Gold Biotechnology Co., Ltd., Chengdu, China), 2.0% matrine OD (Hebei Ruibaode Biochemical Co., Ltd., Shijiazhuang, China), 300 billion spores/g Beauveria bassiana WP (Shanxi Lühai Pesticide Technology Co., Ltd., Yuncheng, China), 80 billion spores/mL Metarhizium anisopliae CQMa421 OD (Chongqing Julixin Bioengineering Co., Ltd., Chongqing, China), Mamestra brassicae nucleopolyhedrovirus (MbNPV, Jiangxi Xinlong Biotechnology Co., Ltd., Yichun, China), and Bacillus thuringiensis (Shandong Yitai Biotechnology Co., Ltd., Tai’an, China).
2.3. EPG Test
To investigate the feeding behavior of adult E. onukii on tea plants treated with six different biopesticides, the following treatments were established: (1) Bt was diluted 250-fold in distilled water; (2) MbNPV was diluted 900-fold in distilled water; (3) CQMa421 was diluted 850-fold in distilled water; (4) B. bassiana was diluted 500-fold in distilled water; (5) azadirachtin was diluted 400-fold in distilled water; (6) matrine was diluted 125-fold in distilled water. Distilled water served as the blank control. The concentration of each biopesticide was chosen based on the manufacturer’s recommended dosage for field application against piercing-sucking pests in tea plantations. Healthy, uniformly growing tea shoots approximately 30 cm in length were selected, immersed in the prepared pesticide solutions for 30 s, and then inserted into nutrient soil in small pots. The treated plants were placed in a cool, ventilated area for approximately 5–10 min to air-dry. Eighteen plants were used for recording per treatment. Each tea shoot was infested with a single E. onukii adult, constituting one biological replicate. A total of 18 replicates were measured for each treatment. Tea plants of uniform growth stage were randomly assigned to different treatment groups to minimize plant variability.
Prior to the experiment, the EPG equipment was zeroed and preheated for 15 min. Test insects were starved for 1 h before the assay. A gold wire (20 mm in length, 18.5 μm in diameter; EPG Systems, Wageningen, The Netherlands) was attached to the pronotum of a leafhopper using a hand-mixed, water-based silver conductive paint glue (EPG Systems). The other end of the gold wire was connected to the top of a copper electrode (20 mm in length). The copper electrode with the attached insect was inserted into a foam board, and the insect was observed for 2–3 min to ensure secure attachment and good condition. The prepared insect was then placed on the abaxial surface of a fresh tea leaf with sufficient space for movement. The copper electrode was inserted into the input of the EPG head-stage amplifier. Another copper electrode (100 mm in length, 2 mm in diameter) was inserted into the soil of the plant container to complete the circuit. The Faraday cage was closed, and the recording was initiated using the EPG Stylet+ 01.20 software to monitor the probing behavior of E. onukii on the tea plant for a 6 h period.
A Giga-8 DC-EPG device (EPG Systems, Wageningen, The Netherlands) was used to monitor the probing and feeding activities of adult E. onukii on tea plants. Signals were digitized using a USB analog/digital converter card (DI-158U; DATAQ Instruments, Akron, OH, USA) and transmitted to a laptop computer for recording.
2.4. Field Trial
A field trial was conducted from 8 July to 29 July 2025 in Xiajiachong Village, Shihegang Town, Xinyang, China (32°18′ N, 114°35′ E). The tea plantation was in the peak occurrence period of E. onukii, received no prior pesticide applications, and the tea plants were in good growth condition. The tested biopesticides were diluted to their respective concentrations (Table 1), with distilled water serving as the control. The intermediate concentration for each biopesticide was chosen based on the manufacturer’s recommended dosage for field application against piercing-sucking pests in tea plantations. To evaluate the robustness and flexibility of each biopesticide under practical conditions, we additionally tested one lower and one higher concentration relative to the recommended rate. A completely randomized block design was employed, in which each treatment had three replicates, resulting in a total of 57 plots, each measuring 60 m^2^ and randomly allocated across treatments. To prevent interference between different treatments, plots for different agents were separated by a row of tea bushes approximately 5 m wide. Applications were performed using a motorized knapsack sprayer (Shijiazhuang Funong Machinery Co., Ltd., Shijiazhuang, China). During spraying, emphasis was placed on achieving uniform coverage on the tea canopy surface and the abaxial sides of the tender shoots. The application was conducted under suitable weather conditions, without precipitation or excessive wind. The pre-treatment pest density was determined by random sampling within each treatment area before 9:00 a.m. on the day of application. For each plot, five points were randomly selected. Using the sweep net sampling method, we employed a standard sweep net (38 cm in diameter) with a sturdy handle and a fine mesh collection bag to conduct a uniform sweep within a 5 m^2^ area at each point to count the number of E. onukii. Post-treatment pest populations were surveyed using the same method before 9:00 a.m. on days 3, 5, 9, 14, and 21 after treatment.
The control efficacy and population reduction rate were calculated using the following formulas:
2.5. Data Analysis
All data were analyzed in GraphPad Prism 9 software (GraphPad Software, San Diego, CA, USA). All data were tested for normality (Shapiro–Wilk test) and homogeneity of variance (Levene’s test) prior to analysis. Data on EPG were analyzed by one-way analysis of variance using Tukey’s HSD post hoc test. The field trial data were analyzed using two-way analysis of variance with Tukey’s HSD.
3. Results
3.1. Effects of Six Biopesticides on the Frequency and Duration of EPG Waveforms in E. onukii
The feeding of adult E. onukii on tea plants produced seven distinct waveforms: Np, Pd, C, E, R, F, and S (Figure 1). The count and total duration of waveform Np were significantly higher following treatment with MbNPV, CQMa421, and matrine compared to the control. Azadirachtin increased the total duration of waveform Np but decreased its frequency (Figure 2b and Figure 3b). All six biopesticides significantly reduced the count of S and Pd waveforms (Figure 2a,d,h) and significantly shortened their total durations (Figure 3a,d,h). With the exception of Bt, the other five biopesticides significantly reduced the number of waveform C (Figure 2c). However, among these five, only four (MbNPV, CQMa421, azadirachtin, and matrine) significantly shortened the total duration of waveform C (Figure 3c). Treatments with Bt, azadirachtin, and B. bassiana significantly reduced the number of waveform E (Figure 2e), while MbNPV and CQMa421 significantly increased the total duration of waveform E (Figure 3e). Bt, MbNPV, B. bassiana, and azadirachtin significantly prolonged the total durations of R and F waveforms (Figure 3f,g). Only Bt and MbNPV significantly increased the count of waveform F (Figure 2g), and only MbNPV and B. bassiana significantly increased the count of waveform R (Figure 2f).
3.2. Field Trial of Bt at Different Concentrations Against E. onukii
The control efficacy of Bt against E. onukii increased to approximately 90% by day 9 and peaked at 100% by day 21 for all three dilutions (Figure 4a). No significant differences in efficacy were observed among the different concentrations on days 3, 9, and 21 (p > 0.05). On day 5, the efficacy of the 250-fold dilution was significantly higher than that of both the 100-fold (p = 0.003, t = 3.614) and the 350-fold dilution (p < 0.001, t = 4.134), reaching 59.78% (Figure 4a, Table S1). Conversely, on day 14, the efficacy of the 250-fold dilution was significantly lower than that of the 100-fold (p < 0.001, t = 4.154) and the 350-fold dilution (p < 0.001, t = 4.186), measuring 70.27% (Figure 4a, Table S1). The population reduction rates of E. onukii in all Bt-treated groups were significantly higher than in the control (p < 0.05) (Table S1). By day 9, the population reduction rates for all three concentrations reached approximately 90% (Figure 4b). However, on day 14, the reduction rate for the 250-fold dilution decreased to 77.88%. By day 21, the population reduction rates for all three concentrations reached 100% (Figure 4b).
3.3. Field Trial of MbNPV at Different Concentrations Against E. onukii
The efficacy of MbNPV against E. onukii increased over time following application, with no significant differences observed among the three concentrations (p > 0.05). Control efficacy reached approximately 90% by day 9 and 100% by day 21 for all concentrations (Figure 5a, Table S2). The population reduction rate followed a similar trend. On day 3, the reduction rates for the 900-fold (p = 0.04, t = 2.88) and 1400-fold (p = 0.04, t = 0.84) dilutions were significantly higher than the control, whereas the 400-fold dilution showed no significant difference from the control (p = 0.26, t = 2.03) (Figure 5b, Table S2). From day 5 to day 21, the population reduction rates for all three concentrations were significantly higher than the control (p < 0.001) (Figure 5b, Table S2).
3.4. Field Trial of CQMa421 at Different Concentrations Against E. onukii
Application of CQMa421, the control efficacy against E. onukii gradually increased, with no significant differences observed among different concentrations (p > 0.05). By day 9, all three concentrations exceeded 86%, and by day 21, all three reached 100% (Figure 6a, Table S3). The trend in the population reduction rate was similar to that of the control efficacy. On day 3, the population reduction rates for the 250-fold dilution (p = 0.12, t = 2.39) and the 1500-fold dilution (p = 0.11, t = 2.45) showed no significant differences compared to the control, while the 850-fold dilution resulted in a significantly higher population reduction rate than the control (p = 0.002, t = 3.867) (Figure 6b, Table S3). On day 5, the population reduction rates for the 250-fold dilution (p = 0.006, t = 3.54) and the 850-fold dilution (p < 0.001, t = 4.68) were significantly higher than the control, whereas the 1500-fold dilution showed no significant differences compared to the control (p = 0.21, t = 2.14) (Figure 6b, Table S3). From day 9 to day 21, the population reduction rates for all three concentrations were significantly higher than the control (p < 0.001) (Figure 6b, Table S3).
3.5. Field Trial of B. bassiana at Different Concentrations Against E. onukii
Application of B. bassiana, the control efficacy against E. onukii gradually increased, with no significant differences observed among different concentrations (p > 0.05). By day 9, all three concentrations exceeded 84%, and by day 21, all three reached 100% (Figure 7a, Table S4). The trend in the population reduction rate was similar to that of the control efficacy. On day 3, the population reduction rates for the 100-fold dilution (p = 0.008, t = 3.45) and the 900-fold dilution (p < 0.001, t = 4.55) were significantly higher than the control, while the 500-fold dilution showed no significant differences compared to the control (p = 0.10, t = 2.46). From day 5 to day 21, the population reduction rates for all three concentrations were significantly higher than the control (p < 0.001) (Figure 7b, Table S4).
3.6. Field Trial of Azadirachtin at Different Concentrations Against E. onukii
After the application of azadirachtin, the control efficacy against the E. onukii gradually increased. No significant differences were observed among different concentrations on days 3, 14, and 21 (p > 0.05). On days 5 and 9, the efficacy of the 200-fold dilution was significantly lower than that of the 400-fold dilution (Day 5: p = 0.01, t = 3.095; Day 9: p = 0.02, t = 2.976) (Figure 8a, Table S5). The trend in the population reduction rate was similar to that of the control efficacy. On day 3, the population reduction rates for the 400-fold dilution (p = 0.001, t = 4.08) and the 600-fold dilution (p < 0.001, t = 4.209) were significantly higher than the control, while the 200-fold dilution showed no significant differences compared to the control (p = 0.22, t = 2.12). On day 3, the population reduction rates for the 400-fold dilution (p < 0.001, t = 5.55) and the 600-fold dilution (p = 0.001, t = 4.11) were significantly higher than the control, while the 200-fold dilution showed no significant differences compared to the control (p = 0.18, t = 2.23). From day 9 to day 21, the population reduction rates for all three concentrations were significantly higher than the control (Figure 8b, Table S5).
3.7. Field Trial of Matrine at Different Concentrations Against E. onukii
Within three days of matrine application, control efficacy exceeded 50%, with the 50-fold dilution reaching 66.44% (Figure 9a). Efficacy for all three concentrations increased to 100% by day 21. From day 3 to day 21, no significant differences in efficacy were observed among different concentrations (p > 0.05) (Figure 9a, Table S6). The trend in the population reduction rate was similar to that of the control efficacy. The population reduction rates for all three concentrations were significantly higher than the control from day 3 to day 21 (p < 0.001) (Figure 9b, Table S6).
4. Discussion
This study identified seven feeding waveforms for E. onukii: Np, Pd, S, E, C, R, and F. These findings are consistent with previous reports on the closely related species Empoasca vitis by Miao et al. (2007) [42] and Zheng et al. (2017) [43]. Different nomenclature systems have been proposed for the feeding waveforms of the two tea leafhoppers, E. onukii and E. vitis. For example, Yorozuya (2016) [8] categorized the feeding waveforms of E. onukii into Np (non-probing phase, stylet not yet inserted into plant tissue); Eo1 (presumably a pathway exploration and channel construction phase, stylet inserted but no sap ingestion); Eo2 (presumably a phloem feeding phase, likely ingesting sap from the phloem); and Eo3 (presumably a non-phloem feeding phase, likely ingesting sap from mesophyll tissue). Jin et al. (2012) [7] classified the feeding waveforms of E. vitis into Np, E1, E2, E3, E4, E5, and E6. Among these, four waveforms (Np, E1, E2, and E3) were associated with probing and non-probing activities. E1 and all three of its waveform subtypes (E1-A to E1-C) were linked to the formation of a salivary sheath, stylet tearing, and channel cutting in viscous artificial diets. Both waveforms E2 and E3 were associated with active ingestion from liquid artificial diets. Most waveforms observed in this study can be mapped to those described in the aforementioned studies (Table 2). For instance, waveform Eo1 of E. onukii corresponds to waveform Pd in this study, Eo2 corresponds to waveform E, and Eo3 corresponds to waveform C.
The initiation of stylet probing is observed as waveform Np. This flat waveform indicates that the insect is not probing or that the stylet has not yet penetrated the leaf tissue. An increase in either the duration or frequency of waveform Np is positively correlated with the level of disruption to normal feeding behavior induced by the treatment [41]. In this study, treatments with MbNPV, CQMa421, and matrine significantly increased both the frequency and duration of waveform Np produced by adult E. onukii during probing (Figure 2b and Figure 3b). In contrast, azadirachtin significantly reduced the frequency of waveform Np but prolonged their duration (Figure 2b), indicating that all four aforementioned biopesticides possess feeding-interfering activity. All six biopesticides significantly reduced the frequency of waveforms S and Pd (Figure 2a,d,h) and shortened their duration (Figure 3a,d,h). Intracellular probing (waveform Pd) is the process by which the tea leafhopper penetrates leaf tissue, establishes channels, and damages multiple cells. It is primarily used to assess plant chemical composition and its palatability to the insect [41,43]. Waveform S is associated with saliva secretion and salivary sheath formation. E. vitis secretes gel-like saliva only during the initial probing phase, forming an incomplete salivary sheath that envelops the base of the stylet [7]. However, it is unclear whether E. onukii also forms a salivary sheath. In this study, waveform S often followed waveform pd, suggesting that the six biopesticides may interfere with E. onukii feeding by reducing the duration and frequency of channel cutting and salivary sheath formation.
Waveforms C and E represent the two primary feeding phases of E. onukii, corresponding to active ingestion when the stylet has not fully penetrated the phloem and passive ingestion when the stylet enters the phloem, respectively. Treatments with azadirachtin and B. bassiana significantly reduced the frequency of waveforms C and E (Figure 2c,e), indicating that these two biopesticides hinder sap-ingestion behavior of E. onukii. Previous studies have shown that the duration of waveform C is substantially shorter than that of waveform E in both duration and proportion, suggesting that sap-sucking insects primarily acquire nutrients via pressure-driven flow from the phloem into the stylets [8,43]. The results of this study appear to contradict this conclusion, as the duration and frequency of waveform C were roughly equivalent to, or even slightly higher than, those of waveform E. However, we observed that in the control group, E. onukii feeding was accompanied by mechanical obstruction (waveform F), which accounted for 9.45% of the total waveform events and 8.21% of the total duration (Figure 2a and Figure 3a). Waveform F frequently occurred before, between, or after waveforms C and E, suggesting that upon encountering a mechanical barrier, E. onukii is likely forced to abort ingestion and search for a new probing site. This conclusion is supported by Zheng et al. (2017) [43], who reported that the resistant tea cultivar ‘Huangguanyin’ exhibited the longest average duration of waveform F, whereas the susceptible cultivar ‘Tieguanyin’ showed the longest average duration of waveform E.
At the onset of waveform R, the stylets of E. onukii remain located in the phloem, which may represent either a buffering rest period during feeding obstruction or a slow withdrawal process of the stylets from the leaf tissue following prolonged hindered feeding. Consequently, waveforms R and F consistently increase or decrease simultaneously [43]. In this study, Bt, MbNPV, B. bassiana, and azadirachtin prolonged the duration of both R and F waveforms (Figure 3f,g), whereas MbNPV increased their frequency (Figure 2f,g). These findings align with the results reported by Hamouche et al. (2024) [41], who observed significantly longer total durations of waveform F in plants treated with Bt and B. bassiana. The disruption observed during the feeding process may be associated with pathogen-induced systemic resistance. The control function of Bt exhibits diversity, as both its spores and crystal proteins can induce broad-spectrum systemic immunity in tomatoes against multiple fungal diseases and pests [44]. B. bassiana induces resistance by colonizing plant tissues. For example, seed treatment with B. bassiana in the maize systemically enhances the activity of defense enzymes such as peroxidase (POD) and polyphenol oxidase (PPO) in leaves, thereby inhibiting the growth and development of pests like Ostrinia furnacalis [45].
Studies have found that aphid nitrogen acquisition primarily depends on a specific sustained feeding phase within the phloem (waveform E), whose successful establishment and maintenance are significantly influenced by host nitrogen levels. Specifically, aphids exhibit notably longer feeding durations in the high-nitrogen plants [46,47,48,49]. Furthermore, arbuscular mycorrhizal fungi (AMF) colonization is associated with reduced nitrogen content in leaves [46], suggesting that microbes may indirectly affect insects by modulating host nitrogen nutrition. Hence, we speculate that Bt and B. bassiana may similarly affect host plant nitrogen metabolism, potentially disrupting the establishment and maintenance of the insects’ passive phloem-feeding phase (waveform E). This disruption could, in turn, contribute to population decline, though further research is needed to confirm this hypothesis. In addition, piercing-sucking pests such as aphids primarily assess host plant nutritional quality by sensing amino acid concentrations in phloem sap, which determines whether they continue feeding [48]. Fungi can enhance proline content and the activity of multiple defense enzymes in plants, thereby improving plant resistance against pests [11]. Thus, fungal priming of plants may interfere with or influence early insect feeding behavior by altering the nutritional signals that insects rely on for host recognition. This sheds light on how entomopathogens may affect early host recognition and feeding in insect pests.
In this study, the control efficacy of fungi (B. bassiana, CQMa421), bacteria (Bt), and virus (MbNPV) increased rapidly to 84–90% during days 5–9 after application. This observation aligns with previous findings that within 5–7 days after fungal infection, the rapid proliferation of the fungus within host tissues and hemocoel (producing toxic compounds such as beauvericin that inhibit metabolic enzyme activity in insects) leads to complete paralysis of the infected insects [11]. Notably, matrine achieved 66.44% control efficacy as early as day 3, indicating its relatively rapid initial action. Studies of insect detoxification mechanisms further indicate a rise in glutathione-S-transferase activity during the initial three-day period after matrine application. This enzyme plays a major role in the detoxification of insecticides or pathogens [50,51]. Antioxidant enzyme activity rose initially but declined afterwards, weakening the insect’s capacity to scavenge reactive oxygen species, resulting in denaturation of various biomolecules and ultimately death [52,53].
In the present field trial, where the initial pest density was low to moderate (approximately 1–7 individuals/m^2^, Table S7), all six biopesticides demonstrated satisfactory control efficacy. During the early stage (3–9 days) following application, slight variations in efficacy were observed across different concentrations; however, these differences gradually narrowed over the treatment period. By day 21 post-application, all treatments (covering a wide range of concentrations, e.g., 100- to 350-fold for Bt; 50- to 200-fold for matrine) achieved 100% efficacy, with no significant differences among concentrations for most agents at the final time point. This suggests that lower concentrations within the tested ranges may be sufficient to achieve adequate control, particularly under low-to-moderate pest pressure. Therefore, this study supports the potential for dose optimization in practice, which could reduce application costs and environmental load while maintaining efficacy. Growers may consider using lower recommended concentrations, especially during early pest outbreaks, to enhance cost-effectiveness and sustainability.
It is important to acknowledge that field efficacy is influenced by multiple environmental variables, including weather conditions (temperature, humidity, rainfall) and the activity of natural enemies [54,55]. In field trials, applications were conducted under suitable weather conditions, and the observed pest population decline likely resulted from a combination of biopesticide action and natural mortality factors. Natural enemies, such as predatory spiders and parasitoid wasps, are known to play significant roles in regulating E. onukii populations in tea ecosystems [54,56], and it cannot be discounted that they contributed to the observed population decline, especially during the later stages of the trial [10]. Future studies should aim to quantify the relative contributions of biopesticides versus biotic and abiotic factors to better understand the mechanisms underlying field efficacy and to optimize application strategies under varying environmental conditions. Additionally, the present study did not directly measure tea yield parameters, which represents a limitation. Although the observed complete pest control and significant reduction in feeding activity suggest that treated plots likely experienced reduced damage and potentially higher yields compared to untreated controls, direct yield measurements would provide more concrete evidence of the agronomic and economic benefits of biopesticide application [57]. Therefore, future studies should incorporate comprehensive yield assessments alongside efficacy and behavioral evaluations to fully elucidate the practical value of biopesticides in tea production systems.
The six biopesticides represent four mechanistic classes with divergent target sites and behavioral disruption patterns: botanicals (azadirachtin and matrine) act primarily as rapid antifeedants [31,32,33,34,35], interfering with initial host recognition and active feeding; Bt functions as a stomach toxin that disrupts midgut physiology [36,37], thereby reducing passive phloem feeding; entomopathogenic fungi (B. bassiana and CQMa421) infect via cuticle penetration and proliferate within the hemocoel [38,39], causing systemic infection and prolonged resting behavior; and MbNPV induces viral pathogenesis with combined effects on both probing initiation and sustained feeding. Rotating these classes, rather than repeatedly using a single agent, reduces the likelihood of resistance evolution [10]. A practical rotation schedule could involve matrine or azadirachtin for rapid early-season knockdown when pests first appear, followed by Bt or B. bassiana during mid-season to disrupt feeding and maintain suppression; and MbNPV later in the season or during subsequent generations to introduce a composite mode of action. This approach not only preserves the efficacy of individual biopesticides but also aligns with IPM principles by integrating multiple control tactics. Future research should focus on validating optimal rotation sequences under field conditions and assessing their long-term impact on resistance dynamics, natural enemy communities, and overall tea ecosystem health.
While this study focused on evaluating the efficacy of six biopesticides against E. onukii, it is instructive to consider how these biopesticides compare with conventional chemical pesticides commonly used in tea plantations, such as bifenthrin, chlorfenapyr, and thiamethoxam [9,10]. Chemical pesticides typically exhibit rapid knock-down effects, often achieving > 90% control within 24–72 h post-application. However, they are associated with significant drawbacks, including the development of pesticide resistance, resurgence of secondary pests, negative impacts on natural enemy populations, and concerns over chemical residues in harvested tea [9,10]. In contrast, the biopesticides evaluated in this study demonstrated a slower but progressive mode of action, with control efficacy reaching 84–90% by days 5–9 and 100% by day 21 post-application. Although their initial speed of kill is slower than synthetic chemicals, biopesticides offer distinct advantages in terms of environmental safety, compatibility with biological control agents, and suitability for organic and residue-free tea production [10]. Furthermore, their ability to disrupt feeding behavior within hours (as revealed by EPG analysis) means that feeding damage is reduced almost immediately, even before mortality occurs. Regarding residual activity, biopesticides generally have shorter environmental persistence compared to synthetic chemicals, which can be both an advantage (reduced environmental contamination) and a limitation (potentially requiring more precise timing or repeated applications) [10]. Future research should directly compare biopesticides and conventional products under identical field conditions to quantify trade-offs in speed of action, persistence, cost-effectiveness, and overall compatibility with sustainable tea production goals.
5. Conclusions
In summary, all six tested biopesticides were able to disrupt the feeding behavior of E. onukii within a short period. Two botanical pesticides (azadirachtin and matrine) interfered primarily by prolonging the duration or increasing the frequency of non-feeding periods (waveform Np) while reducing both the duration and frequency of the active non-phloem feeding phase (waveform C). Bt extended the duration of the resting phase (waveform R) and decreased the frequency of passive phloem feeding (waveform E). The two entomopathogenic fungi, CQMa421 and B. bassiana, acted through slightly different mechanisms: CQMa421 prolonged waveform Np but reduced waveform C in both duration and frequency, resembling the botanical pesticides; B. bassiana increased the duration and frequency of waveform R and reduced waveform E frequency, similar to Bt. MbNPV not only extended the duration and frequency of both waveform Np and waveform R but also decreased the duration and frequency of waveform C. Beyond causing early feeding disruption, all six biopesticides also demonstrated effective field control of E. onukii under low-density infestation conditions. This study reveals the specific mechanisms by which different types of biopesticides interfere with pest feeding behavior, provides promising candidates for the green control of E. onukii in tea plantations, and highlights their significance for promoting safe tea production and ecologically friendly pest management.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Deka B. Babu A. Pandey A.K. Kumhar K.C. Rajbongshi H. Dey P. Peter A.J. Amalraj E.L.D. Talluri V.R. Potential of the entomopathogenic fungus, Metarhizium anisopliae s. l. for control of red spider mite, Oligonychus coffeae Nietner on tea crop Int. J. Acarol.20224812112910.1080/01647954.2022.2041089 · doi ↗
- 2Pokharel S.S. Yu H. Fang W. Parajulee M.N. Chen F. Intercropping cover crops for a vital ecosystem service: A review of the biocontrol of insect pests in tea agroecosystems Plants 202312236110.3390/plants 1212236137375986 PMC 10304037 · doi ↗ · pubmed ↗
- 3Hazarika L.K. Bhuyan M. Hazarika B.N. Insect pests of tea and their management Annu. Rev. Entomol.20095426728410.1146/annurev.ento.53.103106.09335919067632 · doi ↗ · pubmed ↗
- 4Kumar J. Shakil N.A. Khan M.A. Malik K. Walia S. Development of controlled release formulations of carbofuran and imidacloprid and their bioefficacy evaluation against aphid, Aphis gossypii and leafhopper, Amrasca biguttula biguttula Ishida on potato crop J. Environ. Sci. Health Part B 201146678682 Available online: https://pubmed.ncbi.nlm.nih.gov/21806462/(accessed on 20 January 2026)10.1080/03601234.2012.59206621806462 · doi ↗ · pubmed ↗
- 5Chen K. Huang M.X. Shi Q.C. Xie X. Jin L.H. Xu W.M. Li X.Y. Screening of a potential leafhopper attractants and their applications in tea plantations J. Environ. Sci. Health Part B 20195485886510.1080/03601234.2019.163385631264923 · doi ↗ · pubmed ↗
- 6Backus E.A. Serrano M.S. Ranger C.M. Mechanisms of hopperburn: An overview of insect taxonomy, behavior and physiology Annu. Rev. Entomol.20055012515110.1146/annurev.ento.49.061802.12331015471532 · doi ↗ · pubmed ↗
- 7Jin S. Chen Z.M. Backus E.A. Sun X.L. Xiao B. Characterization of EPG waveforms for the tea green leafhopper, Empoasca vitis Göthe (Hemiptera: Cicadellidae), on tea plants and their correlation with stylet activities J. Insect Physiol.2012581235124410.1016/j.jinsphys.2012.06.00822750027 · doi ↗ · pubmed ↗
- 8Yorozuya H. Monitoring and characterization of DC electrical penetration graph waveforms of tea green leafhopper, Empoasca onukii, on tea plants Entomol. Sci.20161940140910.1111/ens.12224 · doi ↗