Linking Seasonal Dietary Strategies and Selectivity to Inform Forage Restoration for Przewalski’s Gazelle on the Qinghai–Tibet Plateau
Lili Hou, Ming Xu

TL;DR
Przewalski’s gazelles rely on specific plants during food-scarce spring and switch to a more varied diet in summer, offering guidance for habitat restoration in Qinghai Lake National Park.
Contribution
The study identifies season-specific forage preferences and key plant species for gazelle survival in fragmented, seasonal habitats.
Findings
Gazelles depend on a few grass species in spring but diversify their diet in summer.
Electivity analyses reveal consistent core plant preferences and seasonal priority forage species.
Findings guide habitat restoration and zoned management in Qinghai Lake National Park.
Abstract
The Przewalski’s gazelle is an endangered antelope found only in the Qinghai Lake Basin of China. These animals live in fragmented habitats and face serious challenges in finding enough food, especially during the food-scarce spring. To support effective conservation, it is important to understand not only what plants gazelles eat, but which plants they actively prefer. In this study, we examined the diets of nine gazelle subpopulations across different seasons by comparing plant remains in feces with the vegetation available in their habitats. We found that gazelles depend heavily on a small number of grass species to survive the difficult spring period, but shift to a more diverse diet that includes legumes and high-quality plants during summer. Importantly, gazelles do not simply consume the most abundant plants; they consistently select certain preferred species depending on the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1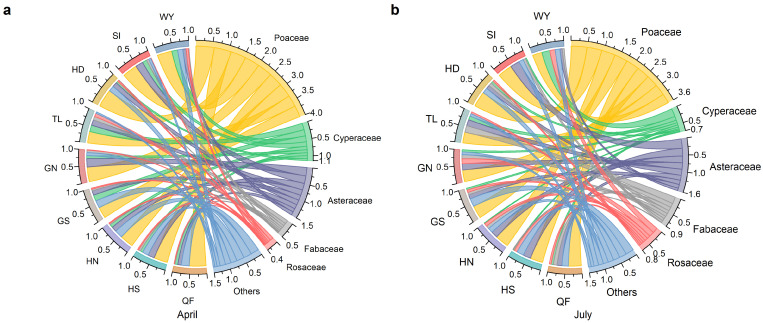 Figure 2
Figure 2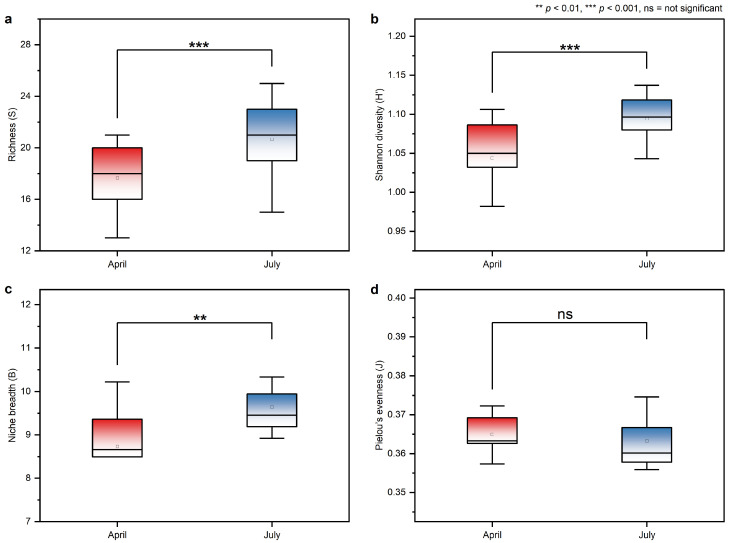 Figure 3
Figure 3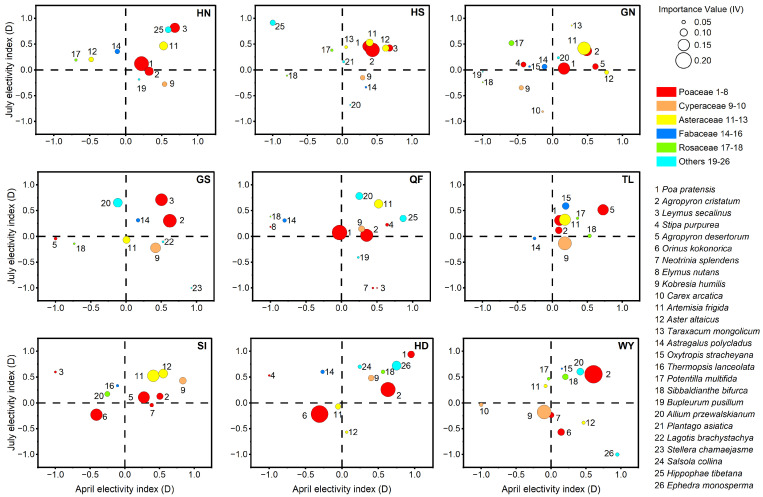 Figure 4
Figure 4- —Guangdong–Hong Kong Joint Laboratory for Carbon Neutrality
- —Guangdong Talent Program
- —Guangdong Science and Technology Program
- —Special Fund for Science and Technology Innovation Strategy of Guangdong Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsWildlife Ecology and Conservation · Rangeland Management and Livestock Ecology · Evolution and Paleontology Studies
1. Introduction
Herbivores are experiencing widespread global declines [1], with large-bodied ungulates particularly vulnerable to habitat fragmentation, climate change [2,3] and human persecution [4]. These pressures are amplified in high-elevation grasslands characterized by strong seasonality, such as the Qinghai–Tibet Plateau [5]. Przewalski’s gazelle (Procapra przewalskii), an endemic and endangered ungulate restricted to the Qinghai Lake Basin, now persists in several geographically isolated subpopulations and plays an important role in maintaining alpine grassland ecosystem structure [6,7]. Despite recent conservation progress, the species continues to face severe threats from ongoing habitat fragmentation and human disturbance [8]. Understanding the ecological basis of its foraging strategies is therefore essential for developing effective management measures [9,10], particularly in the context of the newly established Qinghai Lake National Park [11].
Food resources form the primary link between herbivores and their environment, directly governing survival and reproduction [12]. Dietary ecology thus provides critical insights into how ungulates meet nutritional demands under fluctuating environmental conditions [13,14]. However, many studies remain descriptive, reporting diet composition without simultaneously quantifying plant availability [13,15,16]. This limitation makes it difficult to distinguish active selection from passive consumption based on local abundance [14]. Assessing dietary choices relative to environmental resource availability provides a robust conceptual framework to quantify true forage selectivity and its seasonal shifts [17]. Such selectivity-based approaches are widely used across ungulate systems to identify priority forage taxa, offering valuable guidance for habitat restoration and conservation management [18,19].
The application of availability-corrected selectivity frameworks to understand the foraging preferences of Przewalski’s gazelle remains limited [20,21]. Existing studies have largely concentrated on diet composition or habitat use during the summer growing season and are often restricted to single local subpopulations [21]. Although several dominant forage species, such as Stipa and Leymus, have been identified, these findings rarely integrate dietary intake with quantitative measures of plant availability [20,21]. Consequently, it remains unclear which forage taxa are consistently preferred across the fragmented landscape and which plant resources are most critical during seasonal bottlenecks. This knowledge gap limits the ability to translate dietary information into science-based forage restoration priorities for regional conservation programs.
Here, we investigated the seasonal dietary strategies and forage selectivity of Przewalski’s gazelle across nine geographically isolated subpopulations in the Qinghai Lake Basin. By linking diet composition with environmental resource availability, we quantified true forage selectivity during both the spring bottleneck (April) and the summer growing season (July). Specifically, we aimed to: (i) characterize seasonal shifts in diet composition, diversity, and niche breadth; (ii) identify forage taxa that are actively selected disproportionately to their availability; and (iii) derive season-specific and core priority plant lists to inform forage restoration and zoned management within Qinghai Lake National Park.
2. Materials and Methods
2.1. Study Area
Our study was conducted in the Qinghai Lake Basin on the northeastern Qinghai–Tibet Plateau (97°05′–101°02′ E, 35°05′–37°05′ N). The region is characterized by a continental plateau climate, with annual mean temperatures ranging from −0.8 °C to 1.1 °C and annual precipitation between 327 and 423 mm [22]. Vegetation is dominated by alpine grasslands and meadows, with common species including Kobresia humilis, Achnatherum splendens, and Stipa purpurea [23]. We selected nine geographically isolated subpopulations of Przewalski’s gazelle (QF, TL, GN, GS, HN, HS, HD, SI, WY) from the core groups consistently covered by our long-term monitoring program in the Qinghai Lake Basin. Together, they broadly represent the species’ remaining distribution and habitat variation, while avoiding areas where taxonomic identification may be complicated by hybridization with sympatric gazelle species (Figure 1).
2.2. Vegetation Monitoring and Fecal Sample Collection
Field surveys were conducted in April (spring nutritional bottleneck) and July (summer growing season) of 2023 across all nine gazelle subpopulations. Plant availability was quantified during the peak growing season (July) using quadrat sampling. At each site, at least 10 quadrats (1 × 1 m) were randomly placed along transects spaced approximately 50 m apart within representative habitat patches. Within each quadrat, all vascular plant species were recorded, and their percent cover, height, frequency, and density were measured following standard alpine grassland survey protocols [24,25]. On the Qinghai–Tibet Plateau, the effective plant growing season is short (approximately 3–4 months), during which most annual aboveground biomass and plant taxonomic expression occurs [26]. Moreover, local alpine grasslands are dominated by perennial species (e.g., Stipa spp., Kobresia spp.) that contribute a stable standing vegetation structure across the year. We therefore used July surveys as a basin-wide baseline of the annual standing vegetation assemblage for availability-corrected analyses. In April, vegetation consisted largely of standing dead biomass and early regreening was limited, which precluded a complete quadrat-based assessment of spring availability; accordingly, spring selectivity patterns are interpreted conservatively as bottleneck-stage resource reliance relative to the annual vegetation background [27].
Fecal samples were collected concurrently in both seasons. At each subpopulation, fresh dry pellets were gathered from multiple locations to ensure representative sampling. Pellets were distinguished from sympatric livestock dung based on morphology and associated field signs [14]. All samples were geo-referenced and stored dry prior to laboratory analysis.
2.3. Fecal Microhistological Analysis
Diet composition was determined using fecal microhistological analysis following a modified sodium hypochlorite digestion method [28]. Reference slides were prepared from locally collected plant specimens to facilitate fragment identification (Table S1). For each subpopulation and season, fecal pellets were pooled into a composite sample to characterize subpopulation-level dietary patterns. Composite samples were examined under a Leica DM2500 microscope (Leica Microsystems, Wetzlar, Germany) at 100× magnification. Plant epidermal fragments were identified by comparison with the reference collection, and the relative density (RD) of each plant species was calculated to represent its proportional contribution to the diet [14]. To ensure adequate sampling coverage, 300 microscope fields were analyzed per composite sample in April and 400 fields per composite sample in July, reflecting higher plant diversity during the growing season [29]. In fecal microhistological analysis, observer bias often arises from the subjective identification of partially digested plant fragments and the morphological similarity of epidermal structures among closely related plant taxa (e.g., grasses and sedges). Each field was reviewed systematically, and all identifications were conducted by the same trained observer to minimize inter-observer variability and ensure consistency in taxonomic assignment across all samples.
2.4. Calculation of Vegetation Importance Values (IV)
Plant importance values (IV) were calculated for July quadrats as the mean of relative coverage, relative density, and relative frequency for each species [30]. Specifically, IV was computed as
where RC, RD, and RF represent the relative coverage, relative density, and relative frequency of each species, respectively. These metrics were obtained from field surveys, and IV were used to assess the relative availability of each plant species in the gazelle’s foraging habitat.
2.5. Diversity and Niche Breadth Indices
Dietary diversity and niche breadth were assessed using the Shannon–Wiener diversity index (H), Pielou’s evenness index (J), species richness, and Levins’ niche breadth index (B), all based on the relative proportion of each plant species in the diet [14,21].
where pi is the relative proportion of plant species i in the diet, and S is the total number of species.
2.6. Dietary Selectivity Index
Dietary selectivity was quantified using Jacobs’ index (D). This index was chosen for its ability to account for both plant availability and proportional dietary intake, offering a robust measure of preference independent of overall plant abundance [18]:
where r is the relative proportion of a plant species in the diet and p is its relative proportion in the available vegetation (importance value). Given the constraints of early-season vegetation surveys, July importance values (IV) were used as the proxy for p in both April and July calculations. For each season, basin-wide priority forage taxa were identified using a cross-subpopulation pairwise comparison ranking approach based on Jacobs’ electivity values. Within each subpopulation, a plant taxon received a “win” when its electivity exceeded that of another taxon; wins were summed across pairwise comparisons and subpopulations to generate seasonal rankings [31]. Median electivity values (median D) and the number of subpopulations with available estimates (n) were also reported to summarize preference strength and spatial consistency. To reduce instability associated with rare taxa and extreme electivity values, electivity analyses were restricted to forage taxa that reached ≥3% in dietary proportion in at least one subpopulation within a given season, and that also occurred in the vegetation dataset [32].
2.7. Statistical Analysis
Seasonal differences in dietary diversity indices (species richness, Shannon diversity, Pielou’s evenness, and Levins’ niche breadth) were evaluated using paired t-tests, treating each gazelle subpopulation as a matched replicate sampled in both April and July [14]. Prior to analysis, data distributions were checked for normality (Shapiro–Wilk test), and all indices met the assumptions required for parametric paired comparisons. Statistical significance was set at p < 0.05. All statistical analyses and figure preparation were conducted in OriginPro 2024, with additional visualization performed in R version 4.4.1.
3. Results
3.1. Seasonal Variation in Dietary Functional Groups
Przewalski’s gazelle exhibited distinct seasonal shifts in the composition of major plant functional groups (Figure 2). In April, the diet was dominated by graminoids (Poaceae and Cyperaceae) and Asteraceae across most subpopulations. By July, the contribution of Fabaceae and Rosaceae increased markedly, accompanied by a relative decline in Poaceae and Cyperaceae, reflecting a seasonal transition toward more diverse herbaceous forages during the growing season. At the subpopulation level, functional-group composition showed spatial variation. In April, Cyperaceae contributed more substantially to the diet in TL (20.5%), GS (19.0%), and WY (18.1%) than in other subpopulations, whereas Asteraceae accounted for a higher proportion in GN (25.9%), SI (23.4%), and HS (20.3%). In July, Fabaceae became more prominent in TL (20.9%), while Rosaceae was relatively more important in WY (16.5%) and GN (15.6%). These results demonstrate both a general increase in dietary diversity during summer and marked spatial heterogeneity in foraging strategies among fragmented subpopulations (Table S2).
3.2. Seasonal Variation in Dietary Diversity and Niche Breadth
Dietary diversity and niche breadth varied significantly between seasons (Figure 3). Species richness (S), Shannon diversity (H), and Levins’ niche breadth (B) were all significantly higher in July compared to April (Paired t-test, p < 0.001). This increase is consistent with a broader and more diverse diet during the growing season. In contrast, Pielou’s evenness (J) showed no significant seasonal difference (p > 0.05), indicating that although more plant species were consumed in summer, the proportional distribution among them remained relatively stable. These patterns reflect seasonal constraints on forage availability, where the spring bottleneck restricts diets to fewer dominant taxa compared to the broader niche supported by the summer growing season.
3.3. Vegetation Composition and Availability
Key forage resources were identified by including all plant species with an importance value (IV) > 0.05 in at least one subpopulation, together with several habitat-specialist shrubs of ecological relevance, such as Hippophae tibetana and Ephedra monosperma. Based on July vegetation surveys, regional forage availability was dominated by graminoids, particularly Poaceae and Cyperaceae, with additional contributions from forbs in Asteraceae and Rosaceae (Table 1). These major functional groups were present across all sites, but the relative importance of individual species differed among subpopulations. For example, within Poaceae, Poa pratensis showed relatively high importance values in QF (IV = 0.179), HN (0.133), and GN (0.125), whereas Orinus kokonorica was particularly important in HD (0.311), SI (0.250), and WY (0.110). Among Cyperaceae, Carex arcatica had comparatively high values in WY (0.212) and TL (0.153). In Asteraceae, Artemisia frigida was more important in GS (0.095), TL (0.090), and HD (0.076). These patterns reveal clear spatial heterogeneity in species-level forage composition among fragmented subpopulations (Table S3).
3.4. Dietary Selectivity and Preference Patterns
Dietary selectivity quantified using Jacobs’ electivity index (D) indicated that gazelle foraging was frequently decoupled from local plant availability (Figure 4). Across subpopulations, several locally abundant taxa (large bubbles) showed neutral-to-negative electivity, indicating that high availability did not necessarily translate into high use (e.g., Carex spp. in multiple sites). Conversely, a number of preferred taxa exhibited relatively low availability (small bubbles) yet positive electivity, suggesting active selection for high-quality resources despite their scarcity.
Selectivity patterns also differed among functional groups and seasons. Graminoids such as Agropyron cristatum showed consistently positive electivity across most subpopulations, indicating stable year-round preference. Artemisia frigida (Asteraceae) likewise exhibited broadly positive electivity in both seasons. In contrast, Fabaceae displayed a marked seasonal signal, shifting from neutral or negative electivity in April to strong positive selection in July.
3.5. Seasonal Priority Forage Taxa Across the Basin
The ranking analysis revealed distinct seasonal turnover between April and July (Table 2). A subset of taxa consistently ranked highly in both seasons and were therefore classified as core forage resources supported across subpopulations, most notably Agropyron cristatum and Artemisia frigida. In addition, several taxa emerged as strongly season-specific priorities: Kobresia humilis was characteristic of the spring bottleneck selection, whereas Thermopsis lanceolata and Astragalus polycladus were preferred primarily during the summer growing season. A small number of taxa, such as Hippophae tibetana, exhibited high electivity but occurred in relatively few subpopulations, indicating that they function as locally important resources. Together, these seasonal rankings provide a basin-wide, seasonally resolved priority list for guiding forage restoration and species selection in Qinghai Lake National Park (Table S4).
4. Discussion
Our study moves beyond descriptive diet lists and biomass-centered assessments by explicitly linking gazelle foraging decisions to resource availability across spatially isolated subpopulations. By integrating diet composition with availability-corrected selectivity across nine subpopulations, we demonstrate that Przewalski’s gazelle foraging is structured around a hierarchical set of consistently selected core taxa, supplemented by season-specific resources. This framework provides both mechanistic insight into dietary strategies and an applied basis for identifying priority forage taxa relevant to management in the Qinghai Lake Basin.
4.1. Regional-Scale Patterns of Foraging Strategies
Przewalski’s gazelle exhibited pronounced seasonal shifts in dietary composition and niche breadth, reflecting an intermediate mixed feeder strategy that enables flexible resource use in highly seasonal alpine environments [33,34]. Such seasonal shifts represent a form of phenological tracking, whereby foraging decisions align with temporal changes in plant availability and nutritional quality [35,36]. Importantly, these seasonal patterns varied spatially, with some subpopulations relying heavily on sedges while others shifted more strongly toward forbs, reflecting substantial dietary plasticity in response to local plant assemblages. We interpret this dietary strategy as an adaptive mechanism that may contribute to population persistence in the fragmented and heterogeneous landscapes of the Qinghai Lake Basin.
4.2. Core Foundation Taxa and Season-Specific Priority Resources
Availability-corrected selectivity analyses further revealed that gazelle foraging is not structured around a fixed species list, but rather around a hierarchical spectrum of preferences. A shared set of core taxa, primarily dominant graminoids together with widespread forbs such as Artemisia, were consistently preferred across subpopulations and seasons, forming a basin-wide forage foundation. These core species are likely favored because they provide reliable baseline energy intake under variable conditions: dominant Poaceae combine high standing biomass and broad spatial coverage with relatively high digestible carbohydrate supply, making them predictable forage resources supporting daily maintenance [37].
Beyond this core set, several taxa emerged as strongly season-specific. Spring priority species captured resources repeatedly selected during the bottleneck period, when regreening is delayed and accessible forage is limited. Increased reliance on sedges and cushion plants such as Kobresia spp. likely reflects their persistence as available biomass when other herbaceous resources are scarce [38]. In contrast, summer priority rankings were dominated by legumes and diverse forbs, consistent with selection for nutrient-rich forage during the growing season when plant diversity peaks [13,14].
The observed preference hierarchy further suggests that gazelles balance multiple nutritional objectives beyond energy acquisition alone. Legumes such as Astragalus spp. and Thermopsis lanceolata may provide critical protein supplementation [39,40], while consistent selection of aromatic taxa such as Artemisia frigida and Allium przewalskianum is consistent with the use of plants rich in secondary metabolites that could influence gut function or parasite regulation [41,42]. However, high electivity does not necessarily indicate desirable restoration targets. For example, positive electivity for Stellera chamaejasme in the HS, GS, QF, and WY subpopulations likely reflects context-dependent foraging under extreme resource scarcity, given its known toxicity [43]. Together, these patterns indicate that gazelle persistence depends jointly on forage quantity, composition, and functional diversity, rather than on biomass availability alone.
4.3. Implications for Restoration and Management
Identifying basin-wide core taxa together with season-specific priority resources provides an operational framework for forage restoration in Qinghai Lake National Park. Habitat restoration for Przewalski’s gazelle should prioritize the re-establishment of functionally diverse perennial graminoid communities dominated by Agropyron, Leymus, and Poa, rather than focusing solely on increasing forage biomass [38]. Emphasizing this diverse guild of grasses promotes community stability while providing a reliable energetic baseline across seasons [44,45].
Supplementary incorporation of native legumes and high-quality forbs (e.g., Allium przewalskianum) can further enhance seasonal nutritional carrying capacity by increasing protein availability during the growing season [39,46,47]. Locally selected shrubs such as Ephedra monosperma and Hippophae spp. are consistently chosen by specific subpopulations, underscoring their conservation value and the need to preserve them where they naturally occur [48,49]. For spring bottleneck habitats, particularly Kobresia-dominated meadows, management should emphasize reducing livestock competition [21] and protecting turf integrity during early spring rather than relying on active planting, given the difficulty of restoring turf-forming sedges [50]. Finally, elevated reliance on toxic or unpalatable plants should be interpreted as a warning signal of habitat degradation rather than a conservation objective [51]; high selectivity for species such as Stellera chamaejasme reflects constrained forage choice under scarcity and underscores the need for broader range rehabilitation to restore a balanced graminoid–forb community.
Several limitations should be acknowledged. Vegetation availability was quantified in July, whereas early-spring plant availability could not be fully surveyed due to widespread senescence, requiring spring selectivity to be interpreted relative to the broader plant pool. In addition, dietary estimates were derived from composite fecal samples per subpopulation and season, providing robust subpopulation-level patterns but limiting inference on individual variation. Future work combining multi-season vegetation monitoring, individual-based sampling, and complementary approaches such as DNA metabarcoding or tracking could further refine our understanding of dietary adaptation in alpine herbivores.
5. Conclusions
This study demonstrates that Przewalski’s gazelle adopts a flexible and seasonally structured foraging strategy to cope with the strong seasonal constraints of the Qinghai–Tibet Plateau. By accounting for plant availability, we showed that graminoids form the dietary foundation throughout the year, whereas the selective use of forbs, particularly Fabaceae and Rosaceae, becomes increasingly important during summer. This seasonal shift likely supports recovery from the severe nutritional limitation experienced in early spring, which remains a key bottleneck for population persistence in fragmented habitats.
Our results provide practical guidance for habitat restoration within Qinghai Lake National Park. Rather than relying on generalized re-greening efforts, restoration strategies should focus on functional forage by prioritizing the reseeding of identified core and seasonally important plant taxa, especially those that alleviate spring food shortages. This targeted approach offers a transferable framework for restoring habitats and supporting population recovery of endangered large herbivores facing strong seasonality and habitat fragmentation in alpine ecosystems.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Atwood T.B. Valentine S.A. Hammill E. Mc Cauley D.J. Madin E.M.P. Beard K.H. Pearse W.D. Herbivores at the Highest Risk of Extinction among Mammals, Birds, and Reptiles Sci. Adv.20206 eabb 845810.1126/sciadv.abb 845832923612 PMC 7457337 · doi ↗ · pubmed ↗
- 2Pringle R.M. Abraham J.O. Anderson T.M. Coverdale T.C. Davies A.B. Dutton C.L. Gaylard A. Goheen J.R. Holdo R.M. Hutchinson M.C. Impacts of Large Herbivores on Terrestrial Ecosystems Curr. Biol.20233358461010.1016/j.cub.2023.04.02437279691 · doi ↗ · pubmed ↗
- 3Villar N. Medici E.P. Large Wild Herbivores Slow down the Rapid Decline of Plant Diversity in a Tropical Forest Biodiversity Hotspot J. Appl. Ecol.2021582361237010.1111/1365-2664.14054 · doi ↗
- 4Ripple W.J. Newsome T.M. Wolf C. Dirzo R. Everatt K.T. Galetti M. Hayward M.W. Kerley G.I.H. Levi T. Lindsey P.A. Collapse of the World’s Largest Herbivores Sci. Adv.20151 e 140010310.1126/sciadv.140010326601172 PMC 4640652 · doi ↗ · pubmed ↗
- 5Yang G. Peng C. Chen H. Dong F. Wu N. Yang Y. Zhang Y. Zhu D. He Y. Shi S. Qinghai–Tibetan Plateau Peatland Sustainable Utilization under Anthropogenic Disturbances and Climate Change Ecosyst. Health Sustain.20173 e 0126310.1002/ehs 2.1263 · doi ↗
- 6Liang D. Liu Y. Ping X. Jiang Z. Li C. Ensuring Recovery for the Przewalski’s Gazelle Science 202137416310.1126/science.abm 286534618593 · doi ↗ · pubmed ↗
- 7Liu T. Jiang Z. Wang W. Wang G. Song X. Xu A. Li C. Changes in Habitat Suitability and Population Size of the Endangered Przewalski’s Gazelle Glob. Ecol. Conserv.202343 e 0246510.1016/j.gecco.2023.e 02465 · doi ↗
- 8Liang D. Li C. Habitat Suitability, Distribution Modelling and GAP Analysis of Przewalski’s Gazelle Conservation Animals 20241414910.3390/ani 1401014938200880 PMC 10778258 · doi ↗ · pubmed ↗