Integrated bulk and single-cell transcriptomic profiling reveals bromocriptine sensitivity genes and cellular targets in adenomyosis
Yuqiang Zhang, Danfen Luo, Zhaomei Zhong, Juan Chen

TL;DR
This study identifies genes linked to bromocriptine sensitivity in adenomyosis using bulk and single-cell RNA sequencing, revealing potential biomarkers and treatment targets.
Contribution
The study introduces a novel integrative approach combining bulk and single-cell transcriptomics to identify bromocriptine sensitivity genes in adenomyosis.
Findings
Nine candidate genes were identified as bromocriptine sensitivity markers through cross-validation with external datasets.
Single-cell analysis showed distinct cellular origins for these genes, with ADAM12 and HOXA11 in smooth muscle and epithelial cells.
Genes like SFRP1 and SESN3 were predominantly expressed in fibroblasts and showed increased expression in ectopic lesions.
Abstract
Bromocriptine has emerged as a potential treatment for adenomyosis. This study aimed to identify genes associated with Bromocriptine sensitivity and explore the biological processes and cell-type-specific expression patterns involved. A cross-sectional bioinformatics study was carried out at Shenzhen Baoan Shiyan People’s Hospital from August 2023 to January 2025. We performed differential expression analysis on endometrial RNA-seq data from patients with adenomyosis before and after Bromocriptine treatment. Functional enrichment analysis was conducted to identify pathways influenced by treatment. Candidate genes were validated in two datasets by comparing normal and adenomyotic endometrial tissues. Single-cell RNA sequencing analysis on eutopic, ectopic and control endometrial samples was conducted to further explore their cellular and molecular regulation pathways. Differential…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
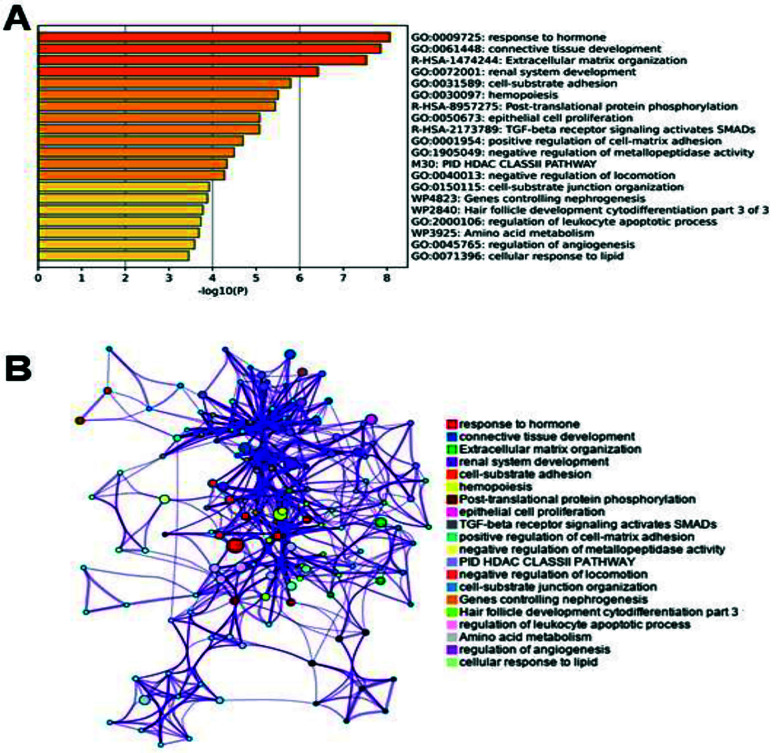 Fig. 1
Fig. 1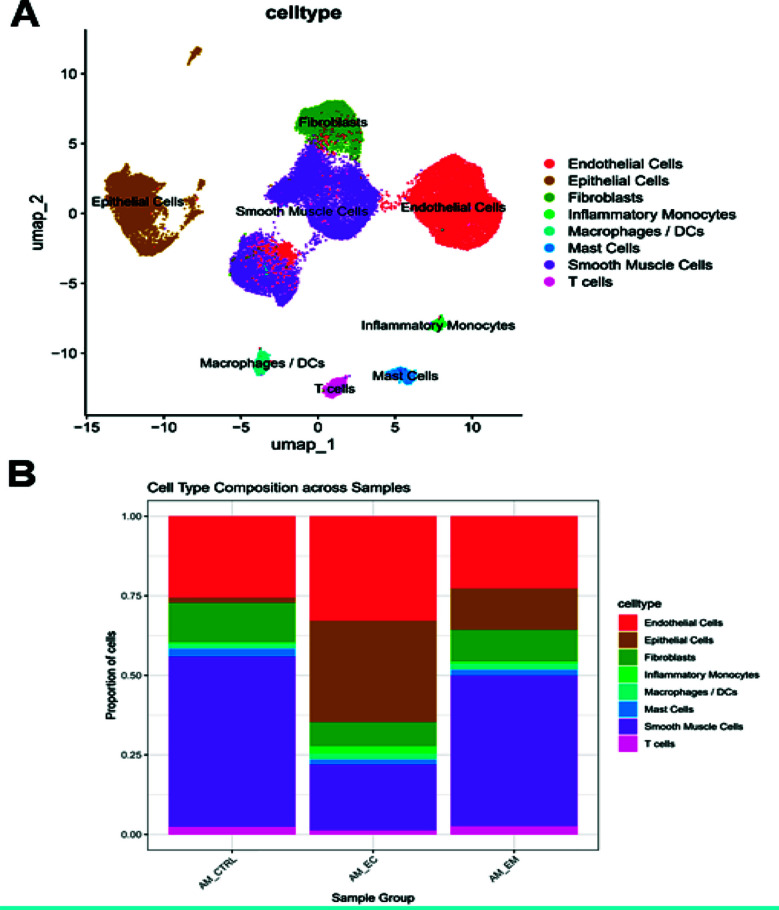 Fig. 2
Fig. 2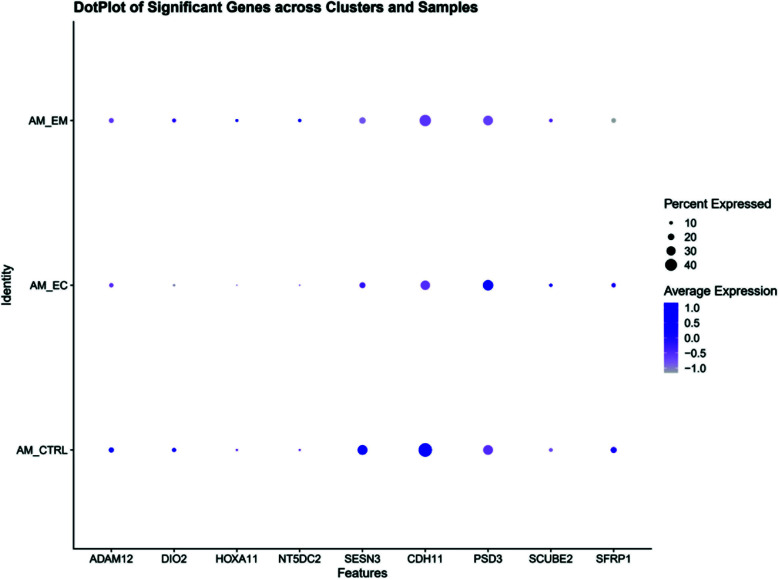 Fig. 3
Fig. 3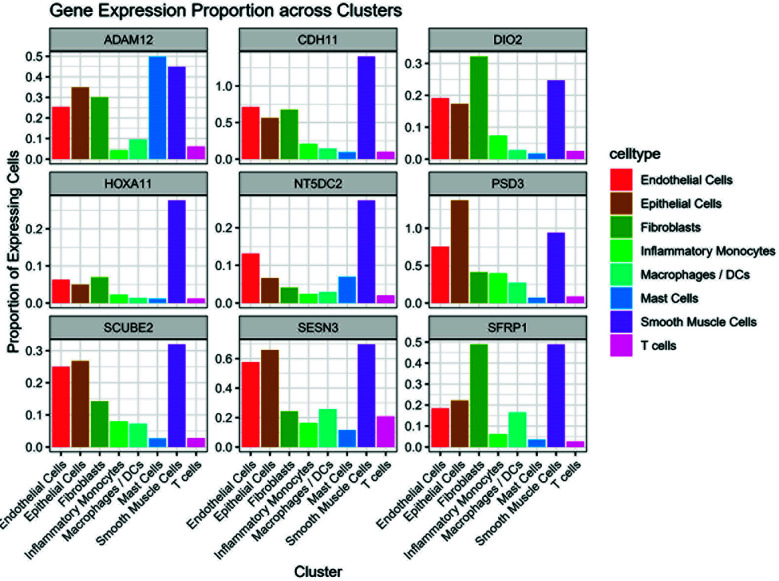 Fig. 4
Fig. 4| Symbol | ||||||||
|---|---|---|---|---|---|---|---|---|
| post-therapy vs. prior therapy | normal vs. lesioned tissue | normal vs. lesioned tissue | ||||||
| log2FC | p value | log2FC | p value | FDR | log2FC | p value | FDR | |
| ADAM12 | 1.01 | 0.0377 | 3.80 | 1.23E-10 | 6.35E-09 | 3.07 | 2.58E-07 | 4.39E-06 |
| CDH11 | 0.74 | 0.0402 | 0.68 | 0.0029 | 0.0186 | 1.83 | 4.27E-05 | 0.0002 |
| DIO2 | 0.84 | 0.0328 | 2.35 | 1.27E-05 | 0.0002 | 2.51 | 9.17E-06 | 5.20E-05 |
| HOXA11 | 0.81 | 0.0368 | 0.73 | 6.27E-06 | 0.0001 | 2.83 | 0.0001 | 0.0003 |
| NT5DC2 | 0.93 | 0.0422 | 0.65 | 0.0042 | 0.0240 | 1.07 | 0.0023 | 0.0043 |
| PSD3 | 0.68 | 0.0292 | 0.73 | 0.0108 | 0.0480 | 0.74 | 0.0023 | 0.0043 |
| SCUBE2 | 1.09 | 0.0021 | 0.86 | 0.0070 | 0.0338 | 1.26 | 0.0007 | 0.0020 |
| SESN3 | 0.72 | 0.0043 | 0.81 | 0.0086 | 0.0390 | 1.71 | 0.0009 | 0.0022 |
| SFRP1 | 0.93 | 0.0374 | 1.49 | 0.0002 | 0.0018 | 2.50 | 3.38E-06 | 2.87E-05 |
| GeneSymbol | Description | log2FC | SE | p-value |
|---|---|---|---|---|
| AQP3 | aquaporin 3 (Gill blood group) | -2.36 | 0.66 | 0.0004 |
| SCGB2A1 | secretoglobin family 2A member 1 | -2.18 | 0.75 | 0.0037 |
| NAPSB | napsin B aspartic peptidase, pseudogene | -2.05 | 0.62 | 0.0009 |
| PDK4 | pyruvate dehydrogenase kinase 4 | -1.83 | 0.70 | 0.0095 |
| LINC02888 | long intergenic non-protein coding RNA 2888 | -1.79 | 0.52 | 0.0006 |
| HGD | homogentisate 1,2-dioxygenase | -1.78 | 0.79 | 0.0237 |
| PTGS1 | prostaglandin-endoperoxide synthase 1 | -1.62 | 0.55 | 0.0034 |
| CFD | complement factor D | -1.54 | 0.71 | 0.0302 |
| RXFP1 | relaxin family peptide receptor 1 | -1.53 | 0.74 | 0.0395 |
| IDO1 | indoleamine 2,3-dioxygenase 1 | -1.50 | 0.65 | 0.0207 |
| LGR5 | leucine rich repeat containing G protein-coupled receptor 5 | -1.49 | 0.46 | 0.0013 |
| CFAP300 | cilia and flagella associated protein 300 | -1.48 | 0.39 | 0.0001 |
| SLC3A1 | solute carrier family 3 member 1 | -1.45 | 0.62 | 0.0196 |
| CTNNA2 | catenin alpha 2 | -1.43 | 0.62 | 0.0202 |
| SYT14 | synaptotagmin 14 | -1.36 | 0.55 | 0.0127 |
| TNFSF10 | TNF superfamily member 10 | -1.35 | 0.45 | 0.0028 |
| MGST1 | microsomal glutathione S-transferase 1 | -1.34 | 0.51 | 0.0082 |
| ALDH3B1 | aldehyde dehydrogenase 3 family member B1 | -1.34 | 0.51 | 0.0083 |
| GMPR | guanosine monophosphate reductase | -1.33 | 0.47 | 0.0045 |
| TNFRSF10A | TNF receptor superfamily member 10a | -1.33 | 0.45 | 0.0032 |
| PRUNE2 | prune homolog 2 with BCH domain | -1.31 | 0.59 | 0.0265 |
| SLC26A2 | solute carrier family 26 member 2 | -1.31 | 0.52 | 0.0116 |
| LOC112268284 | -1.31 | 0.62 | 0.0364 | |
| PSORS1C3 | psoriasis susceptibility 1 candidate 3 | -1.28 | 0.62 | 0.0372 |
| H2AC6 | H2A clustered histone 6 | -1.27 | 0.62 | 0.0402 |
| PRKX | protein kinase cAMP-dependent X-linked catalytic subunit | -1.26 | 0.47 | 0.0073 |
| FBXO6 | F-box protein 6 | -1.22 | 0.45 | 0.0064 |
| RGS2 | regulator of G protein signaling 2 | -1.20 | 0.47 | 0.0106 |
| LPIN3 | lipin 3 | -1.20 | 0.58 | 0.0373 |
| DRC1 | dynein regulatory complex subunit 1 | -1.19 | 0.57 | 0.0383 |
| THEM4 | thioesterase superfamily member 4 | -1.18 | 0.53 | 0.0253 |
| TUBA4A | tubulin alpha 4a | -1.15 | 0.49 | 0.0201 |
| SLC34A2 | solute carrier family 34 member 2 | -1.15 | 0.41 | 0.0055 |
| CFAP45 | cilia and flagella associated protein 45 | -1.14 | 0.55 | 0.0368 |
| ANXA2P1 | annexin A2 pseudogene 1 | -1.14 | 0.55 | 0.0398 |
| FTH1P3 | ferritin heavy chain 1 pseudogene 3 | -1.12 | 0.44 | 0.0108 |
| MT2A | metallothionein 2A | -1.11 | 0.56 | 0.0473 |
| GBP2 | guanylate binding protein 2 | -1.11 | 0.55 | 0.0438 |
| PPM1H | protein phosphatase, Mg2+/Mn2+ dependent 1H | -1.10 | 0.49 | 0.0260 |
| TMEM63A | transmembrane protein 63A | -1.09 | 0.41 | 0.0072 |
| MFHAS1 | multifunctional ROCO family signaling regulator 1 | -1.07 | 0.49 | 0.0286 |
| JUNB | JunB proto-oncogene, AP-1 transcription factor subunit | -1.07 | 0.53 | 0.0421 |
| GPT2 | glutamic--pyruvic transaminase 2 | -1.06 | 0.46 | 0.0206 |
| TPD52L1 | TPD52 like 1 | -1.06 | 0.50 | 0.0344 |
| RORC | RAR related orphan receptor C | -1.06 | 0.49 | 0.0322 |
| ITGB8 | integrin subunit beta 8 | -1.05 | 0.47 | 0.0264 |
| ARG2 | arginase 2 | -1.05 | 0.46 | 0.0216 |
| DAPK1 | death associated protein kinase 1 | -1.05 | 0.43 | 0.0160 |
| ZFP36 | ZFP36 ring finger protein | -1.05 | 0.53 | 0.0491 |
| AK7 | adenylate kinase 7 | -1.04 | 0.44 | 0.0183 |
| GABARAPL1 | GABA type A receptor associated protein like 1 | -1.03 | 0.42 | 0.0140 |
| BEST1 | bestrophin 1 | -1.01 | 0.37 | 0.0059 |
| FTH1 | ferritin heavy chain 1 | -0.98 | 0.36 | 0.0067 |
| ARL4C | ADP ribosylation factor like GTPase 4C | -0.94 | 0.47 | 0.0463 |
| SLC48A1 | solute carrier family 48 member 1 | -0.94 | 0.41 | 0.0225 |
| INO80E | INO80 complex subunit E | -0.93 | 0.44 | 0.0357 |
| ST6GAL1 | ST6 beta-galactoside alpha-2,6-sialyltransferase 1 | -0.92 | 0.40 | 0.0225 |
| C4A | complement C4A (Rodgers blood group) | -0.92 | 0.43 | 0.0336 |
| C4B | complement C4B (Chido blood group) | -0.92 | 0.43 | 0.0316 |
| HOOK1 | hook microtubule tethering protein 1 | -0.92 | 0.46 | 0.0447 |
| TRIB1 | tribbles pseudokinase 1 | -0.91 | 0.35 | 0.0090 |
| LGMN | legumain | -0.90 | 0.39 | 0.0206 |
| CMBL | carboxymethylenebutenolidase homolog | -0.90 | 0.45 | 0.0468 |
| VTCN1 | V-set domain containing T cell activation inhibitor 1 | -0.90 | 0.43 | 0.0377 |
| JUP | junction plakoglobin | -0.88 | 0.35 | 0.0113 |
| BTG2 | BTG anti-proliferation factor 2 | -0.87 | 0.37 | 0.0190 |
| HOOK2 | hook microtubule tethering protein 2 | -0.86 | 0.39 | 0.0268 |
| KCNK1 | potassium two pore domain channel subfamily K member 1 | -0.86 | 0.38 | 0.0235 |
| UCA1 | urothelial cancer associated 1 | -0.85 | 0.37 | 0.0213 |
| ERRFI1 | ERBB receptor feedback inhibitor 1 | -0.84 | 0.35 | 0.0145 |
| STRADB | STE20 related adaptor beta | -0.83 | 0.39 | 0.0306 |
| NEDD4L | NEDD4 like E3 ubiquitin protein ligase | -0.83 | 0.41 | 0.0435 |
| SECISBP2L | SECIS binding protein 2 like | -0.81 | 0.41 | 0.0467 |
| ABHD12 | abhydrolase domain containing 12, lysophospholipase | -0.80 | 0.33 | 0.0146 |
| PNP | purine nucleoside phosphorylase | -0.80 | 0.39 | 0.0423 |
| TSPAN1 | tetraspanin 1 | -0.79 | 0.40 | 0.0484 |
| CNPPD1 | cyclin Pas1/PHO80 domain containing 1 | -0.78 | 0.39 | 0.0439 |
| PLIN2 | perilipin 2 | -0.75 | 0.37 | 0.0439 |
| BCAT1 | branched chain amino acid transaminase 1 | -0.74 | 0.32 | 0.0194 |
| LIMS3 | LIM zinc finger domain containing 3 | -0.74 | 0.36 | 0.0387 |
| ALDH2 | aldehyde dehydrogenase 2 family member | -0.74 | 0.28 | 0.0088 |
| WARS1 | tryptophanyl-tRNA synthetase 1 | -0.69 | 0.31 | 0.0277 |
| MTMR14 | myotubularin related protein 14 | -0.67 | 0.34 | 0.0481 |
| GPR108 | G protein-coupled receptor 108 | -0.66 | 0.32 | 0.0381 |
| TAFAZZIN | tafazzin, phospholipid-lysophospholipid transacylase | -0.65 | 0.33 | 0.0472 |
| LIMS4 | LIM zinc finger domain containing 4 | -0.65 | 0.33 | 0.0486 |
| CCDC97 | coiled-coil domain containing 97 | -0.65 | 0.31 | 0.0379 |
| COX2 | cytochrome c oxidase subunit II | -0.61 | 0.29 | 0.0374 |
| TALDO1 | transaldolase 1 | -0.60 | 0.26 | 0.0196 |
| ATXN1L | ataxin 1 like | -0.59 | 0.30 | 0.0448 |
| FKTN | fukutin | 0.61 | 0.29 | 0.0345 |
| ANGEL2 | angel homolog 2 | 0.62 | 0.31 | 0.0433 |
| EMP1 | epithelial membrane protein 1 | 0.63 | 0.31 | 0.0452 |
| SGCE | sarcoglycan epsilon | 0.65 | 0.32 | 0.0449 |
| USP46 | ubiquitin specific peptidase 46 | 0.66 | 0.30 | 0.0248 |
| VCPIP1 | valosin containing protein interacting protein 1 | 0.66 | 0.27 | 0.0131 |
| COPS9 | COP9 signalosome subunit 9 | 0.67 | 0.31 | 0.0297 |
| PSD3 | pleckstrin and Sec7 domain containing 3 | 0.68 | 0.31 | 0.0292 |
| SACS | sacsin molecular chaperone | 0.71 | 0.34 | 0.0359 |
| PEAK1 | pseudopodium enriched atypical kinase 1 | 0.72 | 0.36 | 0.0456 |
| SESN3 | sestrin 3 | 0.72 | 0.25 | 0.0043 |
| DPY19L1 | dpy-19 like C-mannosyltransferase 1 | 0.73 | 0.32 | 0.0229 |
| MZT2B | mitotic spindle organizing protein 2B | 0.73 | 0.36 | 0.0420 |
| CDH11 | cadherin 11 | 0.74 | 0.36 | 0.0402 |
| CD34 | CD34 molecule | 0.75 | 0.37 | 0.0443 |
| KDR | kinase insert domain receptor | 0.76 | 0.37 | 0.0412 |
| NR3C1 | nuclear receptor subfamily 3 group C member 1 | 0.76 | 0.37 | 0.0412 |
| MEF2C | myocyte enhancer factor 2C | 0.76 | 0.36 | 0.0369 |
| STIL | STIL centriolar assembly protein | 0.76 | 0.33 | 0.0209 |
| CAVIN3 | caveolae associated protein 3 | 0.76 | 0.38 | 0.0470 |
| PURA | purine rich element binding protein A | 0.77 | 0.38 | 0.0410 |
| RAB8B | RAB8B, member RAS oncogene family | 0.78 | 0.23 | 0.0006 |
| SLC12A2 | solute carrier family 12 member 2 | 0.80 | 0.40 | 0.0437 |
| LAMA4 | laminin subunit alpha 4 | 0.80 | 0.37 | 0.0315 |
| GLT8D2 | glycosyltransferase 8 domain containing 2 | 0.81 | 0.37 | 0.0278 |
| HOXA11 | homeobox A11 | 0.81 | 0.39 | 0.0368 |
| AFG3L1P | AFG3 like matrix AAA peptidase subunit 1, pseudogene | 0.82 | 0.42 | 0.0491 |
| GUCY1A2 | guanylate cyclase 1 soluble subunit alpha 2 | 0.83 | 0.39 | 0.0345 |
| DIO2 | iodothyronine deiodinase 2 | 0.84 | 0.39 | 0.0328 |
| NAP1L3 | nucleosome assembly protein 1 like 3 | 0.84 | 0.40 | 0.0353 |
| TBC1D2B | TBC1 domain family member 2B | 0.84 | 0.39 | 0.0288 |
| CCDC171 | coiled-coil domain containing 171 | 0.85 | 0.43 | 0.0496 |
| UNC5C | unc-5 netrin receptor C | 0.85 | 0.43 | 0.0493 |
| NADK2 | NAD kinase 2, mitochondrial | 0.86 | 0.42 | 0.0397 |
| BAZ1B | bromodomain adjacent to zinc finger domain 1B | 0.87 | 0.41 | 0.0346 |
| PYGO1 | pygopus family PHD finger 1 | 0.87 | 0.43 | 0.0451 |
| ZNF334 | zinc finger protein 334 | 0.87 | 0.42 | 0.0363 |
| TIMP1 | TIMP metallopeptidase inhibitor 1 | 0.88 | 0.44 | 0.0463 |
| COL21A1 | collagen type XXI alpha 1 chain | 0.88 | 0.41 | 0.0297 |
| RAB30 | RAB30, member RAS oncogene family | 0.89 | 0.38 | 0.0195 |
| TIMP2 | TIMP metallopeptidase inhibitor 2 | 0.89 | 0.44 | 0.0437 |
| ARL15 | ADP ribosylation factor like GTPase 15 | 0.89 | 0.32 | 0.0047 |
| PABIR2 | PABIR family member 2 | 0.90 | 0.38 | 0.0172 |
| FBN1 | fibrillin 1 | 0.91 | 0.32 | 0.0042 |
| EMX2 | empty spiracles homeobox 2 | 0.92 | 0.45 | 0.0428 |
| NT5DC2 | 5’-nucleotidase domain containing 2 | 0.93 | 0.46 | 0.0422 |
| SFRP1 | secreted frizzled related protein 1 | 0.93 | 0.45 | 0.0374 |
| MXRA8 | matrix remodeling associated 8 | 0.94 | 0.41 | 0.0214 |
| Gse1 coiled-coil protein | 0.95 | 0.45 | 0.0366 | |
| DST | dystonin | 0.95 | 0.32 | 0.0031 |
| SLC25A37 | solute carrier family 25 member 37 | 0.95 | 0.46 | 0.0409 |
| SH3PXD2B | SH3 and PX domains 2B | 0.96 | 0.39 | 0.0147 |
| TTC39C | tetratricopeptide repeat domain 39C | 0.97 | 0.48 | 0.0459 |
| RECK | reversion inducing cysteine rich protein with kazal motifs | 0.99 | 0.48 | 0.0382 |
| CLEC11A | C-type lectin domain containing 11A | 0.99 | 0.41 | 0.0166 |
| ECM2 | extracellular matrix protein 2 | 0.99 | 0.37 | 0.0066 |
| TUNAR | TCL1 upstream neural differentiation-associated RNA | 1.01 | 0.47 | 0.0321 |
| PRICKLE2-AS1 | PRICKLE2 antisense RNA 1 | 1.01 | 0.39 | 0.0109 |
| ADAM12 | ADAM metallopeptidase domain 12 | 1.01 | 0.49 | 0.0377 |
| PTPRD | protein tyrosine phosphatase receptor type D | 1.01 | 0.44 | 0.0225 |
| ZNF587 | zinc finger protein 587 | 1.02 | 0.47 | 0.0306 |
| LTBP1 | latent transforming growth factor beta binding protein 1 | 1.02 | 0.40 | 0.0114 |
| MMD | monocyte to macrophage differentiation associated | 1.04 | 0.46 | 0.0244 |
| HDAC4 | histone deacetylase 4 | 1.05 | 0.50 | 0.0360 |
| ZNF264 | zinc finger protein 264 | 1.08 | 0.51 | 0.0358 |
| SCUBE2 | signal peptide, CUB domain and EGF like domain containing 2 | 1.09 | 0.35 | 0.0021 |
| CAMK4 | calcium/calmodulin dependent protein kinase IV | 1.09 | 0.43 | 0.0118 |
| TMEM120B | transmembrane protein 120B | 1.10 | 0.55 | 0.0439 |
| PMEPA1 | prostate transmembrane protein, androgen induced 1 | 1.15 | 0.52 | 0.0276 |
| KCNIP4 | potassium voltage-gated channel interacting protein 4 | 1.16 | 0.50 | 0.0200 |
| MPRIP | myosin phosphatase Rho interacting protein | 1.17 | 0.48 | 0.0147 |
| TYROBP | transmembrane immune signaling adaptor TYROBP | 1.19 | 0.53 | 0.0256 |
| BHLHE41 | basic helix-loop-helix family member e41 | 1.23 | 0.61 | 0.0428 |
| BGN | biglycan | 1.24 | 0.59 | 0.0353 |
| ERICH1 | glutamate rich 1 | 1.25 | 0.56 | 0.0242 |
| FMN1 | formin 1 | 1.27 | 0.51 | 0.0133 |
| STARD10 | StAR related lipid transfer domain containing 10 | 1.28 | 0.61 | 0.0358 |
| H1-4 | H1.4 linker histone, cluster member | 1.30 | 0.56 | 0.0197 |
| GABPB2 | GA binding protein transcription factor subunit beta 2 | 1.38 | 0.52 | 0.0080 |
| DYNC2I1 | dynein 2 intermediate chain 1 | 1.39 | 0.58 | 0.0178 |
| LOC112268323 | 1.39 | 0.56 | 0.0124 | |
| LINC01480 | long intergenic non-protein coding RNA 1480 | 1.42 | 0.43 | 0.0010 |
| ANK2 | ankyrin 2 | 1.45 | 0.51 | 0.0046 |
| COL27A1 | collagen type XXVII alpha 1 chain | 1.49 | 0.75 | 0.0486 |
| IGFBP5 | insulin like growth factor binding protein 5 | 1.55 | 0.55 | 0.0050 |
| TNXB | tenascin XB | 1.69 | 0.61 | 0.0053 |
| SLIT2 | slit guidance ligand 2 | 1.86 | 0.69 | 0.0068 |
| MUC5B | mucin 5B, oligomeric mucus/gel-forming | 1.93 | 0.94 | 0.0392 |
| TUBD1 | tubulin delta 1 | 1.93 | 0.61 | 0.0014 |
| CCL21 | C-C motif chemokine ligand 21 | 2.06 | 0.91 | 0.0233 |
| Cluster | Cell Type Annotation | Most Characteristic Marker Genes |
|---|---|---|
| 0 | Endothelial Cells | SELE, VCAM1, ACKR1 |
| 1 | Smooth Muscle Cells | RGS5, APOE, ABCC9 |
| 2 | Epithelial Cells | KRT7, MAL2, LAMB3 |
| 3 | Smooth Muscle Cells | DES, ACTG2, GREM1 |
| 4 | Endothelial Cells | KDR, CD93, ESM1 |
| 5 | Fibroblasts | LUM, DCN, APOD, CTSK |
| 6 | Smooth Muscle Cells | MYOM2, FOXC2, CDH6 |
| 7 | Epithelial Cells | PAEP, SCGB2A1, SLC34A2 |
| 8 | T cells | CD3D, CD3E, CD3G, TRBC1, TRBC2 |
| 9 | Mast Cells | TPSAB1, TPSB2, CPA3, MS4A2 |
| 10 | Macrophages / DCs | MRC1, CD36, CCL21 |
| 11 | Inflammatory Monocytes | IL1B, CCL3, LYZ, CD86 |
| 12 | Epithelial Cells | KRT6A, KRT6B, S100A8, S100A9 |
| 13 | Epithelial Cells | S100A7, S100A8, S100A9, CSTA |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEndometriosis Research and Treatment · Reproductive System and Pregnancy · Uterine Myomas and Treatments
INTRODUCTION
Adenomyosis is a common gynecological disorder characterized by the presence of ectopic endometrial tissue within the myometrium, leading to severe dysmenorrhea, abnormal uterine bleeding, and infertility.1,2 It is estimated to affect approximately 10% of reproductive-aged women, with prevalence rates reaching 20–35% among patients undergoing hysterectomy.3,4 Despite this considerable burden, the pathogenesis of adenomyosis remains poorly understood, which complicates the development of effective, mechanism-based therapeutic strategies. Current treatments, including hormonal therapies and surgical interventions, mainly provide symptomatic relief rather than targeting the underlying disease processes.5
Bromocriptine is a dopamine agonist primarily used to treat hyperprolactinemia and Parkinson’s disease. Its therapeutic effects are thought to be mediated mainly through inhibition of prolactin secretion. Evidence from endometriosis models suggests it may also inhibit angiogenesis and fibrosis, supporting the hypothesis of a broader mechanism in adenomyosis. Recently, bromocriptine has emerged as a potential therapeutic agent for adenomyosis,6 and clinical studies have reported encouraging evidence supporting its efficacy.7,8 However, the precise molecular mechanisms by which bromocriptine alleviates adenomyosis-related symptoms remain largely unexplored. Elucidating these mechanisms is essential for optimizing its clinical use and identifying biomarkers predictive of treatment response.
The pathophysiology of adenomyosis involves complex molecular and cellular alterations, including dysregulated gene expression, inflammatory responses, and aberrant tissue remodeling. Previous studies have highlighted several genes implicated in the disease, such as MMP2, which is associated with extracellular matrix remodeling, and FSHR, the receptor for follicle-stimulating hormone.9,10 These gene alterations contribute to the invasive growth and impaired function of ectopic endometrial tissue. An in vitro study has further suggested that bromocriptine may exert therapeutic effects in adenomyosis by modulating signaling pathways related to cell proliferation and apoptosis11. Genes that change significantly after treatment may therefore serve as biomarkers of therapeutic response or potential targets for new interventions.
This study aimed to identify genes associated with bromocriptine sensitivity in adenomyosis through comprehensive transcriptomic analysis. Using three datasets from the GEO database, we performed differential expression analysis to identify differentially expressed genes (DEGs) in endometrial tissues from adenomyosis patients before and after bromocriptine treatment. We then conducted functional enrichment analyses to elucidate the biological processes and pathways significantly affected by bromocriptine and constructed a Gene Ontology (GO) enrichment network to map the key pathways involved.
METHODOLOGY
This was a retrospective bioinformatics study based on publicly available transcriptomic datasets obtained from the GEO and SRA databases. All analyses were performed at Shenzhen Baoan Shiyan People’s Hospital between August 2023 and January 2025, and the study protocol was approved by the institutional ethics committee (2024SL006; dated January 24, 2024). Three bulk RNA-sequencing (RNA-seq) datasets (GSE17165311, GSE22800512 and GSE7885113) were retrieved from the GEO database. The GSE171653 dataset comprises RNA-seq data from endometrial tissues of 11 patients with diffuse adenomyosis, including 11 pre-treatment samples and 10 post-treatment samples after six months of bromocriptine therapy. The GSE228005 dataset provides RNA-seq data from normal and ectopic endometrial tissues of five adenomyosis patients. The GSE78851 dataset includes microarray expression data from normal endometrial tissues of three adenomyosis patients and ectopic endometrial tissues from five controls. In addition, a single-cell RNA-seq (scRNA-seq) dataset containing eutopic endometrium (AM_EM, SRR12791872), ectopic endometrium (AM_EC, SRR12791871) and control endometrium (AM_CTRL, SRR12791873) was obtained from the Sequence Read Archive (SRA).14
This study aimed to identify genes associated with bromocriptine sensitivity in adenomyosis by integrating bulk RNA-seq and scRNA-seq analyses. First, differentially expressed genes (DEGs) before and after bromocriptine treatment were identified in the GSE171653 dataset. Functional enrichment analysis was then performed to characterize the biological pathways modulated by treatment. To validate the potential therapeutic relevance of these DEGs, their expression patterns were further examined in normal and ectopic endometrial tissues from the GSE228005 and GSE78851 datasets. Finally, the scRNA-seq dataset was used to assess the cell-type specificity of candidate genes and to compare cell-type composition and gene expression across control, eutopic and ectopic endometrium.
Identification of adenomyosis DEGs associated with Bromocriptine therapy:
We first applied a pre-filtering step to eliminate genes with low counts, ensuring robust downstream analyses. The filtering retained genes showing a minimum expression level across samples, specifically those with counts of 10 or higher in at least as many samples as the smallest group in our study. Next, we used the DESeq2 package in R (v4.3.1) for differential gene analysis, grouping samples by pre-treatment and post-treatment conditions. We identified candidate differentially expressed genes (DEGs) based on a log2-transformed fold-change (lg2FC) greater than 0.585 (fold change > 1.5 times) and a p-value < 0.05.
Functional enrichment analyses on Bromocriptine related DEGs:
For the Bromocriptine therapy related DEGs, we used the online platform Metascape (http://metascape.org) to perform enrichment analysis. Metascape performed enrichment analysis based on multiple databases, including KEGG Pathway, GO Biological Processes, Reactome Gene Sets, Canonical Pathways, CORUM, WikiPathways and PANTHER Pathway. The enriched function terms were clustered according to their similarity (kappa score), with function terms having a kappa score > 0.3 being grouped together. Clusters were filtered to retain those with function terms meeting the criteria of p < 0.01, enrichment factor > 1.5 and count > 3. The adjusted p-values were calculated using the Benjamini-Hochberg procedure. The most statistically significant term within each cluster was selected to represent the cluster. This analysis focused on identifying the primary enriched biological processes, cellular components, molecular functions and other biological pathways. We then utilized online biological tools to visualize the results.
Identification of Bromocriptine sensitive DEGs for adenomyosis:
We further explored the expression differences of these Bromocriptine therapy related DEGs in normal versus adenomyosis-affected tissues using the GSE228005 and the GSE78851 datasets. RNA-seq data of GSE228005 was analyzed using the DEseq2 package and microarray profiling data of GSE78851 was analyzed using the ‘limma’ package. This analysis aimed to identify Bromocriptine-sensitive genes by using the diseased group as a reference, while the expression levels in the normal group were considered the ideal state of treatment. Bromocriptine-sensitive DEGs were defined by the criteria log2FC > 0.585 and false discovery rate (FDR) adjusted p < 0.05.
Single-cell RNA-seq data processing and clustering:
scRNA-seq data from eutopic and ectopic endometrium tissues (AM_CTRL, AM_EM, AM_EC) were processed using Cell Ranger and analyzed with Seurat (v5.0). After quality control filtering (mitochondrial genes >20%, hemoglobin genes >1%, ribosomal genes <1%), normalization and identification of highly variable genes, datasets were integrated using the Harmony algorithm to correct for batch effects. Cells were clustered (resolution = 0.4) and visualized via UMAP. Marker genes were identified using Wilcoxon rank-sum test.
Cell type annotation and focus gene analysis:
Clusters were annotated manually based on canonical cell-type markers, identifying major populations such as epithelial cells, smooth muscle cells, endothelial cells, fibroblasts, T cells, macrophages, mast cells and inflammatory monocytes. To investigate the cellular origin and expression patterns of the candidate genes, we visualized their distribution using Feature Plot, Dot Plot and violin plots. Gene expression proportions within each cluster and between experimental groups (AM_CTRL, AM_EM, AM_EC) were statistically compared using the Wilcoxon test with FDR correction.
RESULTS
After data preprocessing and filtering out low-expression genes, a total of 10,347 genes were retained for differential expression analysis. Using the DESeq2 package, we identified 180 genes that met the predefined significance thresholds (log2FC > 0.585, p < 0.05) and were differentially expressed before and after Bromocriptine treatment. Of these, 90 genes were upregulated and 90 genes were downregulated after Bromocriptine treatment compared to pre-treatment levels. The DEGs were represented in supplemental Table-SI.
Enriched functional process of Bromocriptine therapy-associated DEGs:
The Metascape functional enrichment analysis, based on multiple databases, identified 13 function terms with an adjusted p-value < 0.05. According to statistical significance, the top three terms were response to hormone (GO:0009725), connective tissue development (GO:0061448) and extracellular matrix organization (R-HSA-1474244). Fig.1A displays the top 20 function terms ranked by p-value. To further capture the relationships among the enriched function terms, the enriched network was constructed by the enrichment clusters, where terms with a Kappa score > 0.3 were connected by edges (Fig.1B).
Enrichment analysis on DEGs. (A) Top 20 enriched biological pathways of DEGs associated with Bromocriptine treatment. Bar plot showing the -log10(p-value) of the top 20 functional terms significantly enriched among the DEGs. The most enriched terms include “response to hormone,” “connective tissue development,” and “extracellular matrix organization.” (B) Enrichment network of DEGs before and after Bromocriptine treatment. The network was generated using Metascape based on kappa-statistical similarities among enriched terms (kappa score > 0.3). Nodes represent enriched biological terms, and edges indicate term-term similarity. The network reveals clusters of interconnected terms, highlighting coordinated functional responses, including hormone signaling and tissue remodeling.
Bromocriptine sensitive DEGs for adenomyosis:
For the DEGs before and after Bromocriptine treatment, we further determined treatment sensitivity genes based on their expression differences in normal versus abnormal endometrial tissues. Out of the 180 candidate genes, 161 were found to overlap in the GSE228005 dataset. Differential expression analysis showed that 35 genes were differentially expressed (log2FC > 0.585, FDR < 0.05) in abnormal endometrial tissues compared to normal endometrial tissues. However, only those genes showing consistent differential expression trends before and after Bromocriptine treatment and also exhibited similar differential expression when comparing normal to abnormal endometrial tissues, were identified as Bromocriptine treatment sensitivity genes. In normal endometrial tissues of the GSE228005, 11 genes were upregulated compared to abnormal endometrial tissues and these genes were also highly expressed in endometrial tissues post Bromocriptine treatment. Additionally, six other genes were downregulated in abnormal endometrial tissues and also exhibited low expression levels before Bromocriptine treatment. These genes were further confirmed using the GSE78851, nine genes upregulated in the normal tissue, showed consistent trend in with the GSE228005 Cell clustering and annotation.
After quality control and batch correction, a total of 42,291 cells were retained for downstream analysis. UMAP visualization revealed clear separation of major cell types across samples (Fig.2A). Fourteen clusters were identified and annotated into eight major cell types based on the expression of canonical markers (Table-SII), including endothelial cells, smooth muscle cells, epithelial cells, fibroblasts, T cells, mast cells, macrophages/DCs and inflammatory monocytes. The proportion of each cell type varied between AM_CTRL, AM_EM and AM_EC groups (Fig.2B). Notably, the proportion of Epithelial Cells and Smooth Muscle Cells was markedly increased in the AM_EC group, reflecting potential pathological changes associated with adenomyosis.
Single cell RNA-sequencing analysis. (A) UMAP visualization of cell type annotations across all samples. Uniform Manifold Approximation and Projection (UMAP) plot showing the distribution of 42,291 single cells across 14 clusters. Cells were annotated into eight major cell types based on canonical markers, including endothelial cells, smooth muscle cells, epithelial cells, fibroblasts, T cells, mast cells, macrophages/DCs, and inflammatory monocytes. Each color represents a distinct cell type. Cells from AM_CTRL, AM_EM, and AM_EC samples are integrated using Harmony batch correction. (B) Barplot showing cell type composition across different sample groups. Stacked bar plot illustrating the proportion of each annotated cell type in the three sample groups: AM_CTRL (control endometrium), AM_EM (eutopic endometrium from adenomyosis), and AM_EC (ectopic endometrium). Notably, epithelial cells and smooth muscle cells were increased in the AM_EC group, suggesting tissue remodeling and invasion in adenomyotic lesions.
Cell-Type-Specific Expression of Focus Genes:
The nine candidate Bromocriptine-sensitive genes displayed distinct expression patterns across clusters. For instance: ADAM12 and DIO2 were specifically enriched in smooth muscle cells, implicating their role in matrix remodeling and muscle regeneration. SFRP1 and SESN3 showed strong expression in fibroblasts, suggesting involvement in Wnt signaling and oxidative stress response. HOXA11, a gene essential for endometrial development, was enriched in epithelial cells and upregulated in AM_EC samples. These results were confirmed by DotPlot visualizations (Fig.3).
DotPlot of focus genes across cell clusters and groups. (A) DotPlot visualizing the expression and proportion of expressing cells for nine focus genes (e.g., ADAM12, HOXA11, SCUBE2, SFRP1, SESN3) across all 14 clusters. Dot size represents the percentage of cells expressing each gene, while color intensity indicates average expression level. These genes showed distinct cluster-specific expression, highlighting cell-type specificity in Bromocriptine response. (B) DotPlot displaying the expression of the nine candidate genes across different sample groups (AM_CTRL, AM_EM, and AM_EC). Differential expression was evident for multiple genes, such as ADAM12 and HOXA11, particularly in the AM_EC group, indicating their potential role in disease progression and treatment response.
Differential expression of focus genes between groups:
Differential expression analysis revealed that several focus genes showed significant differences between AM_CTRL and AM_EC/AM_EM groups. Particularly, ADAM12 and HOXA11 were significantly upregulated in the AM_EC group compared to controls (FDR < 0.05). These changes were mainly observed in specific cell types, including Smooth Muscle Cells and Epithelial Cells (Fig.4).
Expression levels of focus genes across cell types and sample groups. Bar plots summarizing the expression proportions of the nine focus genes across annotated cell types within AM_CTRL, AM_EM, and AM_EC groups. Genes such as ADAM12 and HOXA11 showed elevated expression in smooth muscle and epithelial cells in AM_EC tissues, while SFRP1 and SESN3 were enriched in fibroblasts, supporting their role in tissue remodeling and cell-type-specific Bromocriptine sensitivity.
DISCUSSION
Our study is the multi-tiered validation strategy integrating differential expression, independent cohorts, and single cell resolution which robustly identifies cell type specific candidate genes for Bromocriptine response. In this study, we integrated bulk and single-cell transcriptomic data to characterize the molecular response to bromocriptine in adenomyosis. By combining paired endometrial samples before and after bromocriptine treatment with independent datasets of normal and adenomyotic endometrium, we identified nine candidate genes whose expression was consistently dysregulated in disease and shifted toward normal levels following therapy. These bromocriptine-sensitive genes, together with the associated pathways and cell types, provide new insights into the potential mechanisms of bromocriptine action and suggest candidate biomarkers for treatment response.
Pathway enrichment analysis showed that bromocriptine-associated DEGs were significantly enriched in the “response to hormone” pathway. This finding is consistent with previous reports that adenomyosis is characterized by endocrine dysregulation, including disturbances of the hypothalamic pituitary ovarian axis, hyperprolactinemia, and hyperestrogenism.15 As a dopamine agonist, bromocriptine primarily inhibits prolactin secretion and may thereby help restore hormonal homeostasis. In addition, membrane progesterone receptors (mPRα, mPRβ) and estrogen receptor β (ERβ) are upregulated in adenomyotic tissues, and estradiol has been shown to markedly enhance the proliferation of adenomyotic cells.16 Together, these observations suggest that the heightened hormonal responsiveness of adenomyotic tissue plays a central role in disease pathogenesis and that bromocriptine may exert therapeutic effects, in part, by modulating this axis.
Beyond endocrine regulation, our enrichment results also implicated pathways related to connective tissue development and extracellular matrix (ECM) organization, indicating that bromocriptine may influence tissue remodeling another key pathological feature of adenomyosis. Previous animal studies have demonstrated that bromocriptine ameliorates vascular pathology and metabolic syndrome in rats by modulating connective tissue integrity and fibrosis.17,18 Consistent with this, several of the bromocriptine-sensitive genes identified in our analysis are functionally linked to ECM dynamics and fibrotic remodeling. For example, ADAM12, which encodes a metalloprotease involved in cell–matrix interactions and muscle regeneration, has been associated with tissue fibrosis and enhanced cell migration.19,20 CDH11, a cadherin mediating cell–cell adhesion, is associated with multiple fibrotic disorders,21 whereas NT5DC2 has been implicated in the regulation of cellular stress and metabolic responses.22 These findings support a model in which bromocriptine may modulate ECM composition and fibroblast activity, contributing to structural remodeling of eutopic and ectopic endometrium.
Several other candidate genes point to dysregulation of canonical signaling pathways relevant to cell survival and inflammation. SFRP1, SCUBE2, and SESN3 are known regulators of Wnt,23 TGF-β,24 and mTOR25 signaling, respectively. These pathways control key processes such as proliferation, apoptosis, and inflammatory responses and have been implicated in gynecologic diseases. The observation that expression of these signaling-related genes shifts toward a more normal pattern after bromocriptine treatment suggests that the drug may promote favorable molecular remodeling not only at the level of ECM and connective tissue but also through rebalancing core regulatory pathways in endometrial cells.
To dissect these tissue-level changes at cellular resolution, we leveraged single-cell RNA-sequencing data. Our scRNA-seq analysis revealed substantial heterogeneity in cellular composition across control and adenomyosis samples, with a notable expansion of epithelial and smooth muscle cells in AM_EC tissues, reflecting the pathological hallmarks of invasive lesion growth and tissue remodeling in adenomyosis.26 Mapping the nine candidate genes onto specific cell types further refined their potential roles. ADAM12 and HOXA11 were predominantly expressed in smooth muscle and epithelial cells, suggesting that these populations are primary effectors in tissue invasion and fibrotic remodeling. In contrast, SFRP1 and SESN3 were enriched in fibroblasts, supporting their contribution to ECM turnover and fibroblast proliferation.14 These cell type–specific expression patterns provide a more nuanced understanding of how bromocriptine may act through distinct cellular compartments within the endometrium.
Moreover, differential expression analyses demonstrated that ADAM12 and HOXA11 were significantly upregulated in the AM_EC group compared with AM_CTRL, particularly within smooth muscle and epithelial compartments, supporting a potential dual role as biomarkers of disease severity and predictors of therapeutic response.27,28 Collectively, our findings highlight the added value of combining bulk RNA-seq screening with scRNA-seq validation. This integrative approach allows complex tissue-level changes to be decomposed into specific cellular programs and signaling pathways and may inform more precise, cell type–targeted treatment strategies. In particular, therapeutic interventions directed at smooth muscle cells and fibroblast subsets may hold promise as disease-modifying approaches in adenomyosis.29
Limitations:
Nevertheless, several limitations must be acknowledged. First, our analysis was based on publicly available datasets that lacked detailed clinical annotations (e.g., patient age, disease stage, and specific Bromocriptine treatment regimens). These datasets may also harbor inherent batch effects and technical variability, despite our robust normalization efforts. Second, the sample size in the included datasets was modest, limiting statistical power and generalizability. Third, transcriptomic data alone cannot fully capture the complexity of functional outcomes; gene expression changes do not always correlate with protein or metabolite levels.
CONCLUSION
This study identified nine genes potentially sensitive to Bromocriptine treatment in adenomyosis patients by integrating bulk RNA-seq and single-cell transcriptomic data. These genes are involved in key biological processes such as cell adhesion, ECM remodeling, signaling and stress response, all relevant to adenomyosis pathophysiology. Our findings suggest that Bromocriptine may exert cell type-specific effects by modulating pathways involved in hormonal regulation and tissue remodeling. These insights provide a foundation for developing biomarkers of therapeutic response and guide future precision therapies. However, further functional validation and clinical correlation studies are warranted to confirm their utility in clinical practice.
Recommendations:
Therefore, future research should integrate proteomic and metabolomic datasets for a more comprehensive view. Finally, while we identified key genes and pathways, functional validation in vitro or in vivo remains essential to confirm their roles and potential as therapeutic targets.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Vannuccini S Petraglia F Recent advances in understanding and managing adenomyosis F 1000 Res 20198 F 1000 Faculty Rev-283. doi: 10.12688/f 1000 research.17242.1
- 2Javed L Ashraf N Sabqat M Zareen A Atypical Polypoid Adenomyoma (APAM)J Coll Physicians Surg Pak 202131719721 doi: 10.29271/jcpsp.2021.06.7193410278810.29271/jcpsp.2021.06.719 · doi ↗ · pubmed ↗
- 3Upson K Missmer AS Epidemiology of Adenomyosis Semin Reprod Med 20203802-0389107 doi: 10.1055/s-0040-17189203310550910.1055/s-0040-1718920 PMC 7927213 · doi ↗ · pubmed ↗
- 4Gordts S Brosens JJ Fusi L Benagiano G Brosens I Uterine adenomyosis: a need for uniform terminology and consensus classification Reprod Biomed Online 200817244248 doi: 10.1016/s 1472-6483(10)60201-51868199910.1016/s 1472-6483(10)60201-5 · doi ↗ · pubmed ↗
- 5Donnez J Stratopoulou CA Dolmans MM Endometriosis and adenomyosis: Similarities and differences Best Pract Res Clin Obstet Gynaecol 202492102432 doi: 10.1016/j.bpobgyn.2023.1024323810350910.1016/j.bpobgyn.2023.102432 · doi ↗ · pubmed ↗
- 6Naz F Malik A Riaz M Mahmood Q Mehmood MH Rasool G Bromocriptine therapy: Review of mechanism of action, safety and tolerability Clin Exp Pharmacol Physiol 202249903922 doi: 10.1111/1440-1681.136783563503510.1111/1440-1681.13678 · doi ↗ · pubmed ↗
- 7Andersson JK Khan Z Weaver AL Vaughan LE Gemzell-Danielsson K Stewart EA Vaginal bromocriptine improves pain, menstrual bleeding and quality of life in women with adenomyosis: A pilot study Acta Obstet Gynecol Scand 2019981013411350 doi: 10.1111/aogs.136323102531310.1111/aogs.13632 · doi ↗ · pubmed ↗
- 8Andersson JK Pozzi Mucelli R Epstein E Stewart EA Gemzell-Danielsson K Vaginal bromocriptine for treatment of adenomyosis: Impact on magnetic resonance imaging and transvaginal ultrasound Eur J Obstet Gynecol Reprod Biol 20202543843 doi: 10.1016/j.ejogrb.2020.08.0403292042110.1016/j.ejogrb.2020.08.040 · doi ↗ · pubmed ↗