Revision of powdery mildews (Ascomycota, Erysiphaceae) on Rosa in China: unexpected taxonomic complexity with phytopathological implications

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
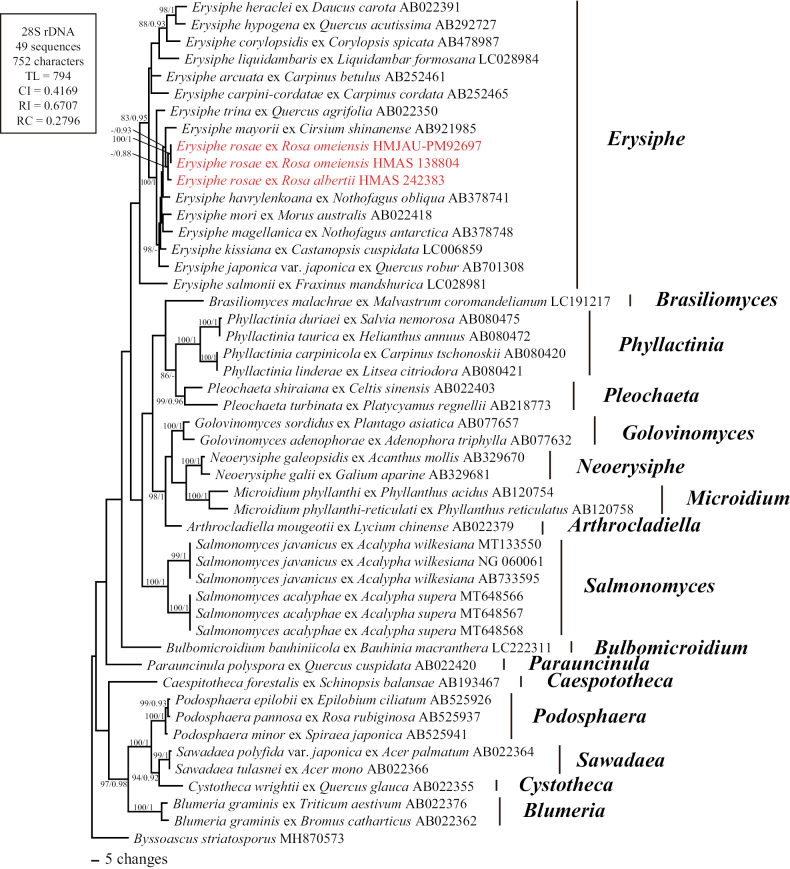 Figure 1
Figure 1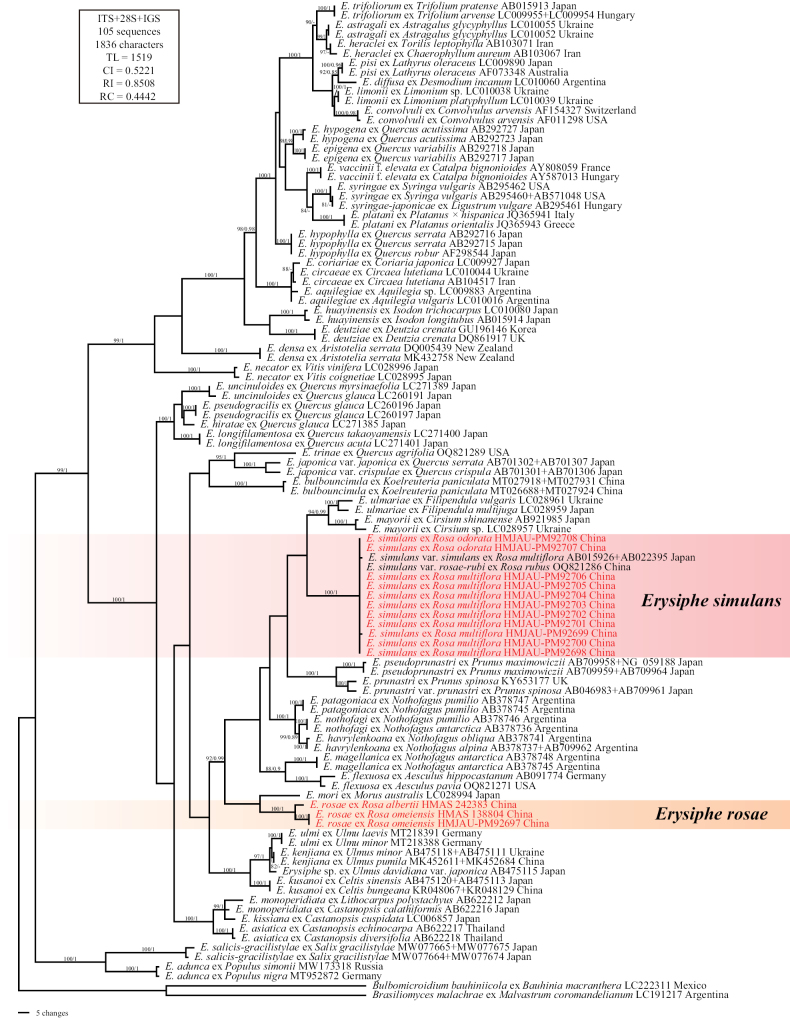 Figure 2
Figure 2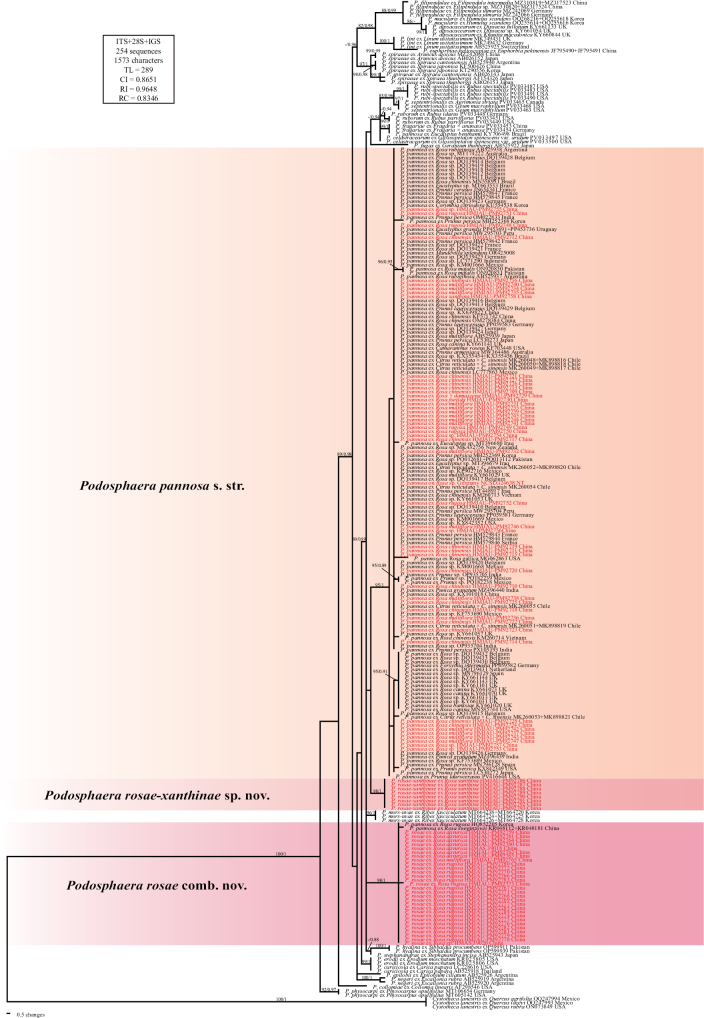 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6 Figure 7
Figure 7 Figure 8
Figure 8 Figure 9
Figure 9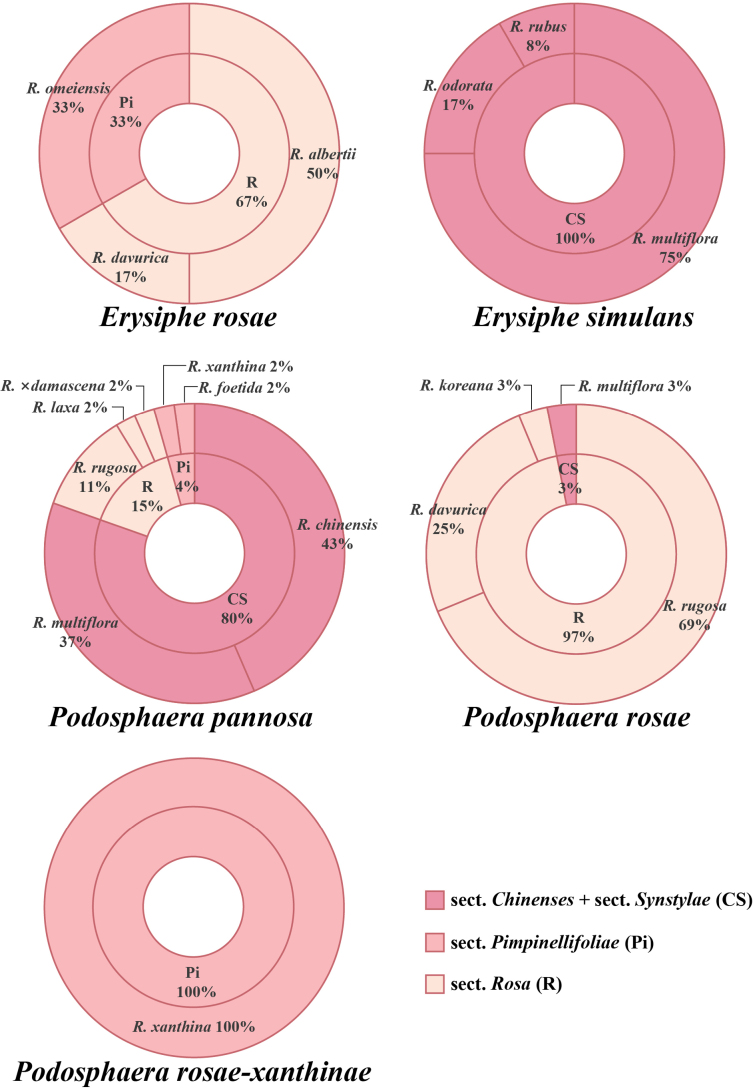 Figure 10
Figure 10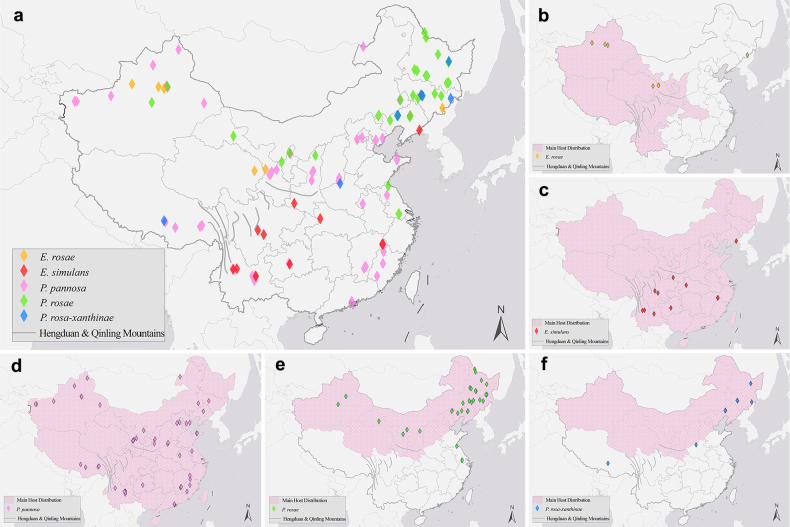 Figure 11
Figure 11| Locus | Primer pair (forward/reverse) | Sequence (5’–3’) | Annealing temperature (°C) | Reference |
|---|---|---|---|---|
| ITS | ITS5/ITS4 | GGAAGTAAAAGTCGTAACAAGG/ TCCTCCGCTTATTGATATGC | 56 |
|
| PM1/PM2 | TCGGACTGGCCYAGGGAGA/ TCACTCGCCGTTACTGAGGT | 55 |
| |
| PM10/PM11 | GGCCGGAAAGTTGTCCAAAC/ TACCGCTTCACTCGCCGTTA | 56 |
| |
| 28S | PM3/TW14 | GKGCTYTMCGCGTAGT/ GCTATCCTGAGGGAAACTTC | 52 |
|
| LSU1/NLP2 | ACCCGCTGAACTTAAGCATA/ GGTCCCAACAGCTATGCTCT | 52 | ||
|
| AGTCTGTGGATTAGTGGCCG/ GAGACAAGCATATGACTAC | 58 |
|
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPowdery Mildew Fungal Diseases · Plant Pathogens and Fungal Diseases · Plant Pathogens and Resistance
Introduction
Rosa (Rosaceae) comprises one of the most economically significant horticultural plant genera worldwide. It is native to the Northern Hemisphere, widely distributed in subtropical to cold-temperate regions (Gu and Robertson 2003), and has been extensively cultivated due to its horticultural appeal, culinary uses, and medicinal properties (Božanić Tanjga et al. 2022; Cui et al. 2022; Hegde et al. 2022; Wang et al. 2022; Zhou et al. 2023; Fayaz et al. 2024; Zhang et al. 2024a; Prasad et al. 2025).
China represents a major center of diversity for Rosa, with 86 species recorded from the country (Luo et al. 2024). Native Chinese roses, such as R. chinensis and R. odorata, have provided many important agronomic and ornamental traits for the breeding of modern rose cultivars (Raymond et al. 2018; Yang et al. 2020; Zhou et al. 2020; Zhang et al. 2024b). The cultivation of edible roses in China dates back more than 2,000 years and is concentrated in three major regions: Shandong, Gansu, and Yunnan provinces. Many species have been widely cultivated for edible products, such as rose tea, rose flower cake, rose paste, and rose wine (Jiang 2024). Rosa species, such as R. chinensis, R. laevigata, R. roxburghii, and R. rugosa, are also used for medicinal purposes in China (Chinese Pharmacopoeia Commission 2020). However, the sustainable production of roses is persistently challenged by powdery mildew. This disease is particularly severe in the cut-flower sector, which constitutes over 80% of China’s floricultural output. Incidences of powdery mildew disease in greenhouses reach 60–90%, with yield losses exceeding 30% (Wang et al. 2018). Furthermore, powdery mildew also inflicts significant damage in the production of edible and medicinal roses, where it causes economic losses of 30–40% (Zuo et al. 2014; Li et al. 2021).
Powdery mildews, a group of obligate biotrophic pathogens within Erysiphaceae, encompass approximately 1,000 species across 17 genera and infect around 10,000 angiosperm taxa worldwide (Braun and Cook 2012; Marmolejo et al. 2018; Kiss et al. 2020; Bradshaw et al. 2024, 2025b). China has reported over 300 species of powdery mildew fungi (Zheng and Yu 1987; Tang et al. 2017; Tang et al. 2018; Qiu et al. 2019; Jin et al. 2021; Feng et al. 2022, 2025a; Liu et al. 2022; Zhang et al. 2025), among which four species and one variety across three genera have been reported to infect roses, including Medusosphaera rosae (now known as Erysiphe rosae), Sphaerotheca pannosa (now known as Podosphaera pannosa), S. rosae (currently considered a synonym of P. pannosa), and Uncinuliella simulans var. rosae-rubi (now known as Erysiphe simulans var. rosae-rubi) (Zheng and Yu 1987; Braun and Cook 2012). This assemblage encompassed nearly all powdery mildew species known globally on Rosa. However, the name Podosphaera pannosa has become a catch-all designation for powdery mildew on roses during the past century. The traditional focus on P. pannosa has contributed to persistent taxonomic confusion and limited recognition of diversity within rose powdery mildews. The genus Medusosphaera, with M. rosae as its type species (Golovin and Gamalitskaya 1962), was previously reduced to synonymy with Erysiphe, as the differences in the morphology of the appendages were regarded as insufficient to maintain it as an independent genus (Braun and Takamatsu 2000). However, so far, this treatment has not been supported by molecular data. In addition, critical species-level ambiguities persist. For example, the recognition of two Sphaerotheca species associated with rose powdery mildew prevailed in previous taxonomic concepts (Salmon 1900; Jaczewski 1927; Blumer 1933, 1967). One is the well-known, previously mentioned species, Podosphaera pannosa, whereas the other species, Sphaerotheca rosae, is a combination introduced by Zhao (1981) based on Jaczewski’s (1927) S. macularis f. rosae. Zheng and Yu (1987) accepted Zhao’s treatment, whereas most subsequent authors considered this name a synonym of P. pannosa (Braun 1987, 1995; Shin 2000; Liu 2010; Braun and Cook 2012). The relationship between the commonly reported P. pannosa and the previously proposed S. rosae has remained unclear until recently due to the lack of detailed morphological and molecular comparison of type specimens. Additionally, Bradshaw et al. (2023) regarded the varieties of E. simulans as dubious due to indistinct morphological boundaries and the lack of genetic differences between E. simulans var. rosae-rubi and E. simulansvar. tandae and emphasized the need for analyses of additional specimens and loci for this species. Consequently, the narrow emphasis on P. pannosa has resulted in an incomplete taxonomic framework for rose powdery mildews, constraining analyses of host range, geographic distribution, and genetic diversity.
These gaps hinder the evaluation of host-associated diversification of powdery mildew fungi on Rosa. As obligate fungi, powdery mildews typically share a close relationship with their host plants. For instance, co-evolutionary relationships have been reported in both the Golovinomyces–Asteraceae and Podosphaera–Rosaceae systems (Matsuda and Takamatsu 2003; Takamatsu et al. 2010). However, it remains unclear whether and how Rosa, a genus with high diversity at the species level, affects the species diversity and distribution patterns of powdery mildews. This lack of knowledge consequently impedes the development of targeted control measures against powdery mildew on Rosa.
Therefore, we sought to establish a comprehensive taxonomic framework for powdery mildews on Rosa. Using nationwide sampling in China and an integrative approach combining morphology with phylogenetic analyses of the ITS region (including 5.8S rDNA), partial 28S rDNA (D1–D2 domains), and the intergenic spacer (IGS), we (1) resolved longstanding taxonomic confusion and clarified species diversity of rose powdery mildews in China and globally; (2) characterized host associations and distribution patterns; and (3) evaluated relationships between these fungi and their Rosa hosts.
Materials and methods
Specimen collection
A comprehensive investigation was conducted across 30 provinces in China. A total of 97 rose powdery mildew specimens from 23 provinces were obtained and deposited in the Herbarium of Mycology of Jilin Agricultural University (HMJAU). An additional 15 specimens were analyzed, of which 14 were type or representative specimens of Medusosphaera rosae (5), Uncinuliella simulans var. simulans (1), Un. simulans var. rosae-rubi (1), Sphaerotheca rosae (4), and Podosphaera pannosa (3), which were borrowed from the Herbarium Mycologicum Academiae Sinicae (HMAS), and one German specimen from the Martin-Luther-Universität Herbarium (HAL), which is designated as the neotype for P. pannosa. In total, 112 dried herbarium specimens were examined in this study. Detailed information about these specimens is shown in Suppl. material 1.
Morphological examination
The sample preparation procedure followed Feng et al. (2025b). For anamorph examinations of dried specimens, the parts of the plants with the most abundant powdery mildew colonies were cut off using a sterile surgical blade and immersed in lactic acid, with the powder layer facing downward on the slide. For teleomorph examinations of dried specimens, chasmothecia were placed onto a drop of lactic acid on the slide. The slide was then gently heated, and the plant tissue was removed before microscopic examination with a phase-contrast light microscope (ZEISS Scope A1, Germany).
DNA extraction and PCR amplification
Genomic DNA was extracted from conidia, mycelia, or chasmothecia using the Chelex-100 method (Walsh et al. 1991; Hirata and Takamatsu 1996). The ITS, 28S, and IGS rDNA regions were generated using polymerase chain reaction (PCR) for molecular analyses. Primers used for amplification are listed in Table 1. The amplification reactions were performed in a total volume of 25 μL reaction mixture, including 2 μL template DNA, 12.5 μL Premix Taq [TaKaRa Taq 1.25 U/25 μL, 2× dNTP Mixture (0.4 mM each), 2× Taq Buffer (3 mM Mg^2+^)] (TaKaRa, Tokyo, Japan), 1 μL of each primer (10 μM), and 8.5 μL ddH_2_O. The PCR reactions were conducted under the following thermal cycling conditions: an initial predenaturation step of 5 min at 95 °C, followed by 35 cycles of 30 s at 94 °C for denaturation, 30 s at 52–58 °C for annealing (temperatures for different primer pairs are listed in Table 1), and 1 min at 72 °C for extension, and a final extension step of 10 min at 72 °C. The PCR products were subjected to electrophoresis in a 1.2% agarose gel in 0.5× TBE buffer. The amplicons were sent to Sangon Biotech (Shanghai, China) for direct sequencing of both strands using the same primers as for the PCR. The assembled sequences were deposited in the National Center for Biotechnology Information (NCBI) GenBank nucleotide database, and the accession numbers are listed in Suppl. material 1.
Phylogenetic analyses
Sequences of ITS, 28S, and IGS newly generated in this study, along with related sequences from GenBank, were automatically aligned using MUSCLE in MEGA X separately (Kumar et al. 2018) and then manually adjusted. Sequence alignments from different fragments were concatenated using PhyloSuite v1.2.3 (Zhang et al. 2019; Xiang et al. 2023) to construct a concatenated matrix.
Three datasets were then constructed: a 28S rDNA dataset of Erysiphaceae for clarifying the phylogenetic position of Medusosphaera in Erysiphaceae, with Byssoascus striatosporus as the outgroup; an ITS+28S+IGS dataset of the genus Erysiphe for clarifying the phylogenetic position of Erysiphe rosae and exploring whether there are genetic differences among varieties of E. simulans, with both Bulbomicroidium bauhiniicola and Brasiliomyces malachrae as outgroups; and an ITS+28S+IGS dataset of Podosphaera sect. Sphaerotheca with all credible sequences of P. pannosa from GenBank for clarifying the phylogenetic relationship of P. pannosa and Sphaerotheca rosae, with Cystotheca lanestris as the outgroup. The accession numbers for the sequences from GenBank are indicated on the corresponding phylogenetic trees. Alignments were submitted as Suppl. materials 2–4.
Phylogenetic analyses were conducted using the maximum parsimony (MP), maximum likelihood (ML), and Bayesian inference (BI) methods. MP analyses were conducted in PAUP* 4.0 (Swofford 2002), identifying optimal trees through a heuristic search with tree-bisection-reconnection (TBR) branch swapping and 1,000 random sequence additions to thoroughly explore tree space. All gaps were treated as missing data, and sites were treated as unordered and unweighted. The ML analyses were performed in raxmlGUI v2.0-beta (Edler et al. 2021) using the GTRGAMMA evolutionary model. Bootstrap supports and trees were obtained by running a rapid bootstrap analysis of 1,000 replicates. For BI analyses, the best-fit substitution models were estimated using MrModeltest v2.3 (Nylander 2004) based on the implementation of the Akaike information criterion (AIC). Four Markov chain Monte Carlo (MCMC) chains were run using MrBayes v3.2.7 (Ronquist and Huelsenbeck 2003). The analysis ran for 3 M generations, and trees were sampled every 100 generations. The first 25% of trees were discarded as burn-in, and a majority-rule consensus tree of all remaining trees was calculated to determine the posterior probabilities for individual branches. The runs were stopped when the standard deviation of split frequencies reached below 0.01. All methods produced congruent topologies, with horizontal branch lengths in the selected MP tree reflecting inferred substitutions. The best-scoring tree from the MP analysis was further edited using Adobe Illustrator 2024. Bootstrap values from BI and ML analyses were superimposed on the MP framework.
Host preference
To elucidate the association between powdery mildews and their Rosa hosts, the relative frequency (RF) was calculated to present the host frequency of rose powdery mildews. For each rose powdery mildew species, the respective distribution among different Rosa sections and species was analyzed. The frequency of individual powdery mildew species on a particular host section or species was calculated as RF (%) = (f / N) × 100. Here, f represents the number of specimens of a powdery mildew species from a specific Rosa section or species, and N is the total number of specimens identified as the powdery mildew species concerned. This study involved 13 Rosa species across four sections: sect. Chinenses (C), sect. Pimpinellifoliae (Pi), sect. Rosa (R), and sect. Synstylae (S). Because sections Chinenses and Synstylae are not reciprocally monophyletic and form a nested (paraphyletic) relationship (Cheng et al. 2025), we treated them as a single group (Chinenses + Synstylae, CS) in this analysis.
Distribution patterns and co-distribution analysis with primary hosts
The accurate locations of 110 Chinese specimens (Suppl. material 1) were used in the distribution analysis. The distribution maps were drawn using ArcGIS 10.8 (Esri, Redlands, CA, USA). To clarify the distribution patterns of rose powdery mildews in China, the coordinates of all 110 specimens were inserted into a map as point features, with different species labeled in different colors. To explore whether there are co-distributions of rose powdery mildews and their hosts, separate maps were generated for each powdery mildew species, overlaid with the distribution range of its primary host. The distribution areas in China of the corresponding primary hosts are based on the Rosa monograph of Luo et al. (2024) and are marked in pink on the maps.
Results
Phylogenetic analyses
A total of 292 sequences were obtained in this study, comprising 102 ITS, 101 28S, and 89 IGS sequences (Suppl. material 1). Four phylogenetic trees were constructed based on these newly generated sequences and those retrieved from GenBank.
The 28S sequence alignment matrix of Erysiphaceae totaled 752 characters and consisted of 49 sequences, including three newly generated and 45 retrieved from GenBank. The maximum likelihood (ML) bootstrap supports greater than 80%, and Bayesian posterior probabilities (BPP) over 0.8 are shown in Fig. 1. In this phylogenetic tree, sequences of Erysiphe rosae cluster within the Erysiphe clade, confirming the previously proposed synonymy of Medusosphaera and Erysiphe.
Phylogenetic analysis of Erysiphaceae based on 28S rDNA sequences. Bootstrap values larger than 80% for the maximum likelihood (ML) analyses conducted are displayed, followed by posterior probabilities ≥80. The newly generated sequences in this study are highlighted in red. Byssoascus striatosporus is used as the outgroup taxon.
The ITS+28S+IGS sequence alignment matrix of Erysiphe comprised 1,836 characters and consisted of 105 sequences, including 14 newly generated and 91 retrieved from GenBank. The maximum likelihood (ML) bootstrap supports greater than 80%, and Bayesian posterior probabilities (BPP) over 0.8 are shown in Fig. 2. Sequences of E. simulans from this study and GenBank form a highly supported monophyletic group. Sequences of E. rosae vary across different hosts, which may indicate cryptic speciation (Fig. 2).
Phylogenetic analysis of related Erysiphe species based on ITS + 28S + IGS sequences. Bootstrap values larger than 80% for the maximum likelihood (ML) analyses conducted are displayed, followed by posterior probabilities ≥80. The newly generated sequences in this study are highlighted in red. Bulbomicroidium bauhiniicola and Brasiliomyces malachrae are used as outgroup taxa.
The ITS+28S+IGS sequence alignment matrix of Podosphaera sect. Sphaerotheca totaled 1,573 characters and consisted of 254 sequences, including 88 newly generated and 166 retrieved from GenBank. The maximum likelihood (ML) bootstrap supports greater than 80%, and Bayesian posterior probabilities (BPP) over 0.8 are shown in Fig. 3. The phylogenetic tree showed that sequences of Podosphaera on Rosa hosts cluster into three distinct clades. The ex-neotype sequences of P. pannosa (PV581762 + PV584310) and ex-type sequences of Sphaerotheca rosae (PX239400 + PX239279) fall into separate clades, providing definitive molecular evidence that S. rosae is a species distinct from P. pannosa. Moreover, sequences retrieved from powdery mildew on Rosa xanthina form a well-supported clade separate from P. pannosa, suggesting the presence of a novel Podosphaera species on Rosa.
Phylogenetic analysis of related Podosphaera sect. Sphaerotheca species based on ITS + 28S + IGS sequences. Bootstrap values larger than 80% for the maximum likelihood (ML) analyses conducted are displayed, followed by posterior probabilities ≥80. The newly generated sequences in this study are highlighted in red. Cystotheca lanestris is used as the outgroup taxon.
Taxonomy
Integrated morphological and phylogenetic analyses led to the following taxonomic conclusions: 1) confirmation of Medusosphaera as a synonym of Erysiphe; 2) confirmation that the previously described varieties of E. simulans do not have taxonomic value and can be dropped; 3) clarification that Podosphaera rosae comb. nov. is an independent species distinct from P. pannosa s. str.; and 4) description of a new species, P. rosae-xanthinae sp. nov. Detailed descriptions and notes are provided below.
Erysiphe
rosae
Taxon classificationFungiHelotialesErysiphaceae
(Golovin & Gamalitsk.) U. Braun & S. Takam., Schlechtendalia 4: 4, 2000
CECF8F03-0AD5-568B-AE7E-9110C260BDB5
≡ Medusosphaera rosae Golovin & Gamalitsk., Bot. Mater. Otd. Sporov. Rast. Bot. Inst. Komarova Akad. Nauk S.S.S.R. 15: 92, 1962.
Type.
Holotype: Kyrgyzstan • Tjan-Shan, Kavak-Tau, Tabylgyty, on Rosa albertii, 14 Sep. 1959, Gamalitskaya (LE 193792). Reference collection: China, Xinjiang, Urumqi, on Rosa albertii, 3 Aug. 2009, Z.Y. Zhao & B. Xu (HMAS 242383). Reference sequences: PX506005 (ITS), PX506007 (28S), PX570626 (IGS).
Symptoms and morphology of Erysiphe rosae on Rosa omeiensis (HMJAU-PM92697). a Symptoms. b1, b2 Hyphal appressoria. c1, c2 Conidiophores with conidia formed singly. d1, d2 Conidia. e Chasmothecium. f Peridium cells. g Short appendages. h1, h2 Long appendages. i1–i3 Asci. j1, j2 Ascospores. Scale bars: 10 μm (b1, d2, f, j1, j2); 20 μm (e, g–i3).
Specimens examined.
• HMJAU-PM92697, HMAS 36517, HMAS 36518, HMAS 44287, and HMAS 138804. Detailed specimen information is presented in Suppl. material 1.
Morphological description.
Braun and Cook (2012: 501–502).
Host range and distribution.
on Rosa (albertii, amblyotis, davurica, omeiensis, webbiana), Rosaceae; Asia (China, India, Kyrgyzstan, Russia, Far East).
Notes.
Erysiphe rosae is characterized by sinuous long appendages and bristle-like short appendages. This taxon has been controversially treated at the generic level. It was first introduced as the type species of the genus Medusosphaera and later transferred to Erysiphe because the morphology of the chasmothecial appendages was no longer recognized as suitable for differentiation at the generic level (Golovin and Gamalitskaya 1962; Braun and Takamatsu 2000). The correct allocation of M. rosae to Erysiphe was confirmed in the present study based on lobed hyphal appressoria and conidiophores with singly formed conidia (Fig. 4), as well as sequences successfully retrieved from specimens of this species, which form a highly supported species clade within Erysiphe (Fig. 1). It should be noted that the sequences of the three examined specimens clustered together according to their host plants, which indicates previously unrecognized diversity within E. rosae, possibly suggesting cryptic formae, a taxonomic unit recently reapplied in Erysiphaceae (Bradshaw et al. 2025c; Feng et al. 2025a). However, due to the limited number of specimens available, it was impossible to obtain additional molecular data or to observe potential morphological differences. Therefore, following the species concept proposed by Bradshaw et al. (2022), we provisionally treated these specimens as E. rosae s. lat. Since the holotype of this species from Kyrgyzstan has not yet been sequenced, the Chinese specimen HMAS 242383 on the type host Rosa albertii is selected as the reference collection with reference sequences for the interim.
Erysiphe
simulans
Taxon classificationFungiHelotialesErysiphaceae
(E.S. Salmon) U. Braun & S. Takam., Schlechtendalia 4: 23, 2000
A58C0F16-B9A6-5965-8855-B877253C0723
≡ Uncinula simulans E.S. Salmon, Ann. Mycol. 6: 2, 1908. ≡ Uncinuliella simulans (E.S. Salmon) R.Y. Zheng & G.Q. Chen, Acta Microbiol. Sin. 19(3): 286, 1979. = Uncinuliella simulans var. rosae-rubi R.Y. Zheng & G.Q. Chen, Acta Microbiol. Sin. 19(3): 288, 1979. ≡ Erysiphe simulans var. rosae-rubi (R.Y. Zheng & G.Q. Chen) U. Braun & S. Takam., Schlechtendalia 4: 23, 2000. = Uncinuliella simulans var. tandae U. Braun, Mycotaxon 22(1): 92, 1985. ≡ Erysiphe simulans var. tandae (U. Braun) U. Braun & S. Takam., Schlechtendalia 4: 24, 2000.
Type.
Lectotype (designated by Braun 1987): Japan • Morioka, on Rosa multiflora, 21 Oct. 1906, Sawada (TNS-F-214613). Reference sequences (designated by Bradshaw et al. 2023): AB015926 (ITS), AB022395 (28S) [in Takamatsu et al. (1999) and Mori et al. (2000)].
Symptoms and morphology of Erysiphe simulans on the type host, Rosa multiflora (HMAS 13631). a1–a3 Symptoms. b Chasmothecium. c Peridium cells. d1–d4 Long appendages. e1–e3 Short appendages. f1–f6 Asci. g1, g2 Ascospores. Scale bars: 50 μm (b); 20 μm (c–d4, f1–f6); 10 μm (e1–e3, g1, g2).
Symptoms and morphology of Erysiphe simulans “var. rosae-rubi” on Rosa rubus (HMAS 11418). a1–a4 Symptoms. b Chasmothecium. c Peridium cells. d1–d4 Long appendages. e1–e3 Short appendages. f1–f5 Asci. g1, g2 Ascospores. Scale bars: 50 μm (b); 20 μm (c–d4, f1–f5); 10 μm (e1–e3, g1, g2).
Specimens examined.
• A total of 13 specimens were examined, including HMJAU-PM92698 to HMJAU-PM92708, and two specimens from the Herbarium Mycologicum Academiae Sinicae, HMAS 11418 and HMAS 13631. Detailed specimen information is presented in Suppl. material 1.
Morphological description.
Braun and Cook (2012: 586).
Host range and distribution.
on Rosa (multiflora, odorata, rubus), Rosaceae; Asia (China, Japan, Korea).
Notes.
Bradshaw et al. (2023) found that the sequences of E. simulans var. rosae-rubi and E. simulans var. tandae grouped together with high bootstrap values. The authors reported that the morphological differences were only gradual or had overlapping quantitative features and, as such, concluded that varieties under E. simulans should be discarded. The morphology of a reference specimen of E. simulans from Japan (HMAS 13631) and the holotype of E. simulans var. rosae-rubi from China (HMAS 11418) were re-examined in this study (Figs 5, 6). We confirmed that the two are highly similar in all characteristics. The differences in the number of appendages described by Zheng and Chen (1979) are also indistinct and overlap with each other. Although we failed to amplify sequences from those two specimens, other sequences of E. simulans were obtained from eleven newly collected specimens. The present phylogenetic analysis showed that there are minimal genetic differences within this species. Therefore, it is reasonable and justified to discard these varieties of E. simulans.
Podosphaera
pannosa
Taxon classificationFungiHelotialesErysiphaceae
(Wallr.) de Bary, Abh. Senkenb. Naturf. Ges. 7: 408, 1870. s. str. [emend]
17EF615C-5662-5F30-A2A4-2B9457C570C7
≡ Alphitomorpha pannosa Wallr., Verh. Ges. Naturf. Freunde Berlin 1: 43, 1819. ≡ Erysiphe pannosa (Wallr.) Link (as “Erysibe”), in Willdenow, Sp. pl., Edn 4, 6(1): 104, 1824, nom. sanct. (Fr., Syst mycol. 3(1): 236, 1829). ≡ Sphaerotheca pannosa (Wallr.) Lév., Ann. Sci. Nat., Bot., 3 Sér., 15: 138, 1851.
Type.
Neotype (designated here, MBT10030088): Germany • Lower Saxony, Hannover-Ahlem, experimental section “cemetery planting,” on Rosa sp. cult. (hybrid), 27 Sep. 2018, P. Houska (NCSLG24638). Isoneotype: HAL 3600 F. Ex-neotype sequences: PV581762 (ITS+28S), PV584310 (IGS), PV584347 (TUB).
Symptoms and morphology of Podosphaera pannosa on Rosa chinensis (HMJAU-PM92727). a1–a7 Symptoms. b Secondary hyphae. c Hyphal appressoria. d1–d3 Conidiophores with catenescent conidia. e1–e5 Conidia. f1, f2 Germ tubes. g Chasmothecium. h Peridium cells. i1–i3 Appendages. j1–j4 Asci. k1, k2 Ascospores. Scale bars: 20 μm (b–k2).
Description.
Mycelium on leaves, amphigenous, also on stems and petals, persistent, infected shoots often curled and distorted, forming irregular patches or effuse; primary hyphae hyaline, thin-walled, straight or sometimes flexuous, (2.5–)3–5.5 μm wide; secondary hyphae hyaline, thick-walled, 4–6 μm wide, colonies on twigs only developed in the later stages of the disease; hyphal appressoria indistinct to nipple-shaped, solitary or side by side; conidiophores arising from the upper surface of mother cells, erect, (52–)60–97(–99) × (6.5–)7.5–9.5 μm, foot cells straight, cylindrical, 33–59 × 7–9.5 μm, followed by (1–)2–3 shorter cells, forming catenescent conidia; conidia variable, primary conidia obovoid, secondary conidia doliiform to cylindrical, 20–26 × 11.5–15 μm, length/width ratio 1.5–2.1; germ tubes almost terminal. Chasmothecia gregarious, subglobose to globose, immersed in the secondary mycelial patches, mostly on stems, but also produced on leaves when the disease is severe, (74–)88–113 μm diam.; peridium cells round to irregularly polygonal, 4.0–16 μm diam.; appendages few, in the lower half of chasmothecia, mycelioid, short, usually shorter than chasmothecial diam., rarely up to 1.5 times diam., occasionally branching, brownish at the basal part or brown throughout, septate, thin-walled, smooth, coarse; ascus ovoid to fusiform, colorless, thick-walled, sessile or almost sessile, 87–140 × 63–84 μm, length/width ratio 1.2–1.9, terminal oculus obvious, large, 13.5–21 μm, 8-spored; ascospores ellipsoid to round, colorless, (16.5–)18–25 × 12.5–17.5 μm, length/width ratio 1.1–1.9.
Additional specimens examined.
• A total of 51 specimens were examined, including 50 specimens with voucher numbers HMJAU-PM92709 to HMJAU-PM92758 and one specimen of HMAS 38723. Detailed specimen information is presented in Suppl. material 1.
Host range and distribution.
(phylogenetically confirmed hosts and distribution). Rosa (canina, chinensis, × damascena, foetida, gallica, laxa, majalis, multiflora, albertiirubiginosa, rugosa, xanthina), Prunus (armeniaca, cerasus, laurocerasus, persica, granatum), Rosaceae; Catharanthus roseus, Apocynaceae; Citrus reticulata × C. sinensis, Rutaceae; Corymbia citriodora, Eucalyptus (benthamii, grandis, sp.), Myrtaceae; and Forsythia × intermedia, Oleaceae. Asia (China, India, Iraq, Israel, Japan, Pakistan, South Korea, Vietnam), Europe (Belgium, France, Germany, Netherlands, Serbia, Spain, United Kingdom), North America (Mexico, United States), Oceania (Australia, New Zealand), and South America (Argentina, Brazil, Chile, Peru, Uruguay).
Notes.
Podosphaera pannosa is the most widely distributed species on Rosa spp. Alphitomorpha pannosa, the basionym, was described from Germany, but type material is not preserved (Braun and Cook 2012). Therefore, a German specimen is designated as the neotype with ex-neotype ITS, 28S, IGS, and TUB sequences. Based on the current morphological and phylogenetic results, it is clear that P. rosae is genetically distinguishable from P. pannosa. The records of P. pannosa on R. rugosa (HMAS 01630, 14132) in Zheng and Yu (1987) from China should probably be revised to P. rosae after re-examining the specimens. The description and illustration of P. pannosa in Shin (2000) from Korea are also more in line with P. rosae and in need of revision. There are numerous morphological descriptions of P. pannosa that appear to be composed of characteristics of both P. pannosa and P. rosae (Braun 1987, 1995; Liu 2010; Braun and Cook 2012).
The confusions and misidentifications were documented as early as Salmon’s monograph (1900). He postulated that the primary reason for this confusion was the erroneous assumption that the species of fungus that occurs on a certain host plant in one part of the world will be the same as that growing on the same host in other parts of the world. As such, many powdery mildews on Rosa spp. were identified as P. pannosa, and the other possible species were ignored. In addition, the chasmothecia of P. pannosa are rarely formed (Salmon 1900; Blumer 1933; Homma 1937; Junell 1967; Nomura 1997), so the fact that chasmothecia of P. rosae appear to be more common than those of P. pannosa may be another reason for the confusion. To our knowledge, P. rosae often forms chasmothecia in China in late autumn when the temperature decreases. Chasmothecia of P. pannosa are rarely formed, even when chasmothecia of P. rosae are formed at the same time and place. The chasmothecia of P. pannosa can be found in winter or even the following spring, and they are primarily formed on stems, rarely on leaves, with abundant secondary mycelium. The more frequently produced chasmothecia of P. rosae are undoubtedly the reason that they have often erroneously been assigned to P. pannosa, at least before these two species were separated. Consequently, on account of the persistent taxonomic confusion between these two species, the distribution and host range require further clarification. In this study, host range and distribution are listed based on the results of the phylogenetic analysis (see Fig. 3).
Podosphaera
rosae
Taxon classificationFungiHelotialesErysiphaceae
(Jacz.) D.-N. Jin & S.-Y. Liu comb. nov.
939B626C-C809-5AA2-B5B5-8DC411D12293
861536
≡ Sphaerotheca rosae (Jacz.) Z. -Y. Zhao, Acta Microbiol. Sin. 4: 439, 1981.
Basionym.
Sphaerotheca macularis f. rosae Jacz., Karmannyi opredelitel’ gribov II. Muchnistorosyanye griby: 76. 1927.
Symptoms and morphology of Podosphaera rosae on Rosa rugosa (HMJAU-PM92763). a1–a4 Symptoms. b Secondary hyphae. c1, c2 Hyphal appressoria. d1, d2 Conidiophores with catenescent conidia. e1–e4 Conidia. f1–f4 Germ tubes. g Chasmothecium. h Peridium cells. i1–i3 Appendages. j1, j2 Asci. k1–k3 Ascospores. Scale bars: 20 μm (b–k3).
Type.
Lectotype (designated here, MBT10029826), • original illustration in Jaczewski (1927: 77, fig. 19). Epitype (designated here, MBT10030086): China • Shaanxi, Yulin, on Rosa davurica, 18 Oct. 2023, L. Liu & Z.-Y. Zhang (HMJAU-PM92763). Ex-epitype sequences: PX239405 (ITS), PX239284 (28S), PX239896 (IGS).
Mycelium on stems and both sides of leaves, more abundant on the lower surfaces of the leaves, infected shoots often curled and distorted, forming irregular patches or effuse, persistent; primary hyphae hyaline, thin-walled, 3.0–4.5 μm wide, secondary hyphae hyaline, thick-walled, 3.5–6 μm wide, sparsely branched, often intertwined with each other on the lower surface of the leaves; hyphal appressoria indistinct to nipple-shaped, solitary or side by side; conidiophores arising from the upper surface of mother cells, erect, very long, 65–240 (–342) × 7–10.5 μm, foot cells straight, cylindrical, long, 31–195(–302) × 7–10.5 μm, followed by 2–3 shorter cells, forming catenescent conidia; conidia doliiform to ellipsoid, 22–29(–31) × 11.5–15.5(–16.5) μm, length/width ratio 1.5–2.3; germ tubes almost terminal or terminal, short or long. Chasmothecia scattered, subglobose to globose, amphigenous, mostly on the lower surface of leaves, (74–)77–101(–106) μm diam.; peridium cells round to irregularly polygonal, 6.5–17.5 μm diam.; appendages mycelioid, some of them slightly spirally twisted at the base, arising from the equator of the chasmothecia, sometimes curved, wall thick and smooth, light brown below, paler towards the apex, numerous, various in length, (59–)80–405(–483) μm, 4–6 μm in width, septate; ascus ovoid to broad fusiform, colorless, thick-walled, sessile, 84–116(–121) × 56–79 μm, length/width ratio 1.3–1.6, terminal oculus obvious, large, 17–27 μm diam., 8-spored; ascospores ellipsoid–ovoid, colorless, 20–26 × 10–15 μm, length/width ratio 1.5–2.2.
Additional specimens examined.
• A total of 33 specimens were examined, including 27 specimens with voucher numbers HMJAU-PM92759 to HMJAU-PM92785 and six specimens from Herbarium Mycologicum Academiae Sinicae, HMAS 01630, HMAS 14132, HMAS 39017, HMAS 39018, HMAS 39019, and HMAS 39020. Detailed specimen information is presented in Suppl. material 1.
Host range and distribution.
On Rosa (davurica, koreana, multiflora, rugosa, sp.), Rosaceae; Asia (China, Korea).
Notes.
This species was once proposed as Sphaerotheca rosae by Zhao (1981) and accepted by Zheng and Yu (1987), whereas most other authors considered this name a synonym of P. pannosa (Braun 1987, 1995; Shin 2000; Liu 2010; Braun and Cook 2012). After our detailed examinations, including the results of sequence analyses, Zhao’s (1981) taxonomic treatment can be confirmed, i.e., S. rosae can be confirmed as a species. P. rosae is different from P. pannosa in (1) forming chasmothecia mostly on the lower surfaces of the leaves; (2) having longer conidiophores and foot cells; (3) smaller chasmothecia and asci with larger terminal oculi; and (4) longer chasmothecial appendages. Sequences of S. rosae, including ex-type sequences (PX239400 and PX239279 from HMAS 39018; PX239428 and PX239307 from HMAS 39020), grouped together in a monophyletic, highly supported clade separate from the P. pannosa clade. As a result, S. rosae should be reinstated as the new combination Podosphaera rosae. P. pannosa reported by Lee et al. (2011) on Rosa rugosa in Korea is morphologically and phylogenetically consistent with P. rosae.
Podosphaera
rosae-xanthinae
Taxon classificationFungiHelotialesErysiphaceae
D.-N. Jin & S.-Y. Liu sp. nov.
7C92D18C-B9D5-56D4-AB37-B080690134EE
861535
Etymology.
Epithet derived from the name of the type host species, Rosa xanthina.
Symptoms and morphology of Podosphaera rosae-xanthinae on Rosa xanthina (HMJAU-PM92790). a1–a5 Symptoms. b1, b2 Secondary hyphae. c Hyphal appressoria. d1, d2 Conidiophores with catenescent conidia. e1–e4 Conidia. f1, f2 Germ tubes. g1, g2 Chasmothecia. h Peridium cells. i1–i3 Appendages. j1–j3 Asci. k1, k2 Ascospores. Scale bars: 20 μm (b1–k2).
Type.
Holotype: China • Jilin, Changchun, on Rosa xanthina, 8 Aug. 2023, D.N. Jin & X.L. Wu (HMJAU-PM92790). Ex-holotype sequences: PX239433 (ITS), PX239312 (28S), PX239920 (IGS). Isotype: HMAS 354228.
Diagnosis.
Close to Podosphaera pannosa, but differing in having shorter conidiophores and smaller asci, and forming a separate well-supported species clade in the phylogenetic analysis.
Description.
Mycelium on stems and both sides of leaves, more abundant on the lower surface of the leaves, forming irregular patches or effuse, persistent, infected shoots often curled and distorted, primary mycelium white, secondary mycelium in dense, pannose patches on the stems; primary hyphae hyaline, thin-walled, 2.5–5 μm wide, secondary hyphae hyaline, thick-walled, 3.5–5.5 μm wide, sparsely branched; hyphal appressoria indistinct to nipple-shaped, solitary or side by side; conidiophores arising from the upper surface of mother cells, erect, 48–91(–96) × 7–9.5 μm, foot cells straight, cylindrical, (24–)26–56(–61) × 6.5–9.5 μm, followed by 2–3 shorter cells, forming catenescent conidia; primary conidia obovoid, secondary conidia doliiform to ellipsoid, 18.5–25 × 11–14.5 μm, length/width ratio 1.4–1.9; germ tubes almost terminal to almost lateral, short. Chasmothecia gregarious, subglobose to globose, immersed in the secondary mycelial patches, mostly on the stems, (71–)73–96 μm diam.; peridium cells round to irregularly shaped, 5–21(–26) μm diam.; appendages mycelioid, simple, short, arising from the lower half of chasmothecia, usually curved, hyaline throughout or light brown below, paler towards the apex, septate, variable in length, usually 0.2–1 times as long as the chasmothecial diam.; ascus ovoid to ellipsoid, colorless, thick-walled, sessile or short-stalked, 73–102 × 57–71 (–75) μm, length/width ratio 1.2–1.6, terminal oculus inconspicuous to distinct, (11–)14–26 μm, 8-spored; ascospores ellipsoid–ovoid, colorless, 23–28 × 15.5–18.5 μm, length/width ratio 1.4–1.7.
Additional specimens examined.
• A total of eight specimens were examined under the vouchers from HMJAU-PM92786 to HMJAU-PM92793. Detailed specimen information is presented in Suppl. material 1.
Host range and distribution.
on Rosa (xanthina), Rosaceae; Asia (China).
Notes.
Asexual powdery mildew morphs are common on Rosa xanthina. The conidiophores and foot cells are usually shorter than those in Podosphaera pannosa. Conidia are also smaller, and the length/width ratio is always less than 2. Chasmothecia are usually formed and are immersed in secondary mycelial patches on twigs in late autumn or early winter. The teleomorph of powdery mildew on Rosa xanthina is similar to P. pannosa, but differs in having broader asci, sometimes with stalks. Sequences of the powdery mildew on R. xanthina form a well-supported, separate species clade in phylogenetic analyses. As a result, a new species, P. rosae-xanthinae, is described, which is so far known only on R. xanthina.
Host preference
A total of 104 Chinese specimens with unequivocal host identifications were used for relative frequency (RF) calculations. Host frequency of powdery mildew is shown in Fig. 10. Species within the same powdery mildew genus display host ranges that are both partially specialized and mutually complementary. Among Erysiphe species, E. simulans is commonly associated with sect. Chinenses + sect. Synstylae (with R. multiflora as the primary host), while E. rosae is more prevalent on hosts of sect. Pimpinellifoliae and sect. Rosa (with R. omeiensis and R. albertii as the primary hosts). In Podosphaera, P. pannosa has the broadest host range, infecting seven Rosa species, with primary hosts belonging to sect. Chinenses + sect. Synstylae (with R. chinensis and R. multiflora as the primary hosts). P. rosae, with the second broadest host range, mainly infects hosts within sect. Rosa (with R. rugosa and R. davurica as the primary hosts). Another Podosphaera species, P. rosae-xanthinae, is so far known only on R. xanthina, a species belonging to sect. Pimpinellifoliae.
Host frequency of each rose powdery mildew species on Rosa. CS = sect. Chinenses + sect. Synstylae, Pi = sect. Pimpinellifoliae, R = sect. Rosa.
Distribution patterns of powdery mildews on Rosa spp. in China
The five rose powdery mildew species identified in this study are distributed across 23 provinces in China, and their overall distribution is summarized in Fig. 11a (Erysiphe rosae, E. simulans, Podosphaera pannosa, P. rosae, and P. rosae-xanthinae are represented by yellow, red, pink, green, and blue points, respectively). The distribution patterns of each species are shown separately in Fig. 11b–f. Additionally, to investigate whether co-distribution exists between each powdery mildew species and its Rosa hosts, the main distribution ranges (including both wild and cultivated) of their primary hosts (according to the results of the Host preference part), based on the Rosa monograph of Luo et al. (2024), are marked in pink for each powdery mildew species on the maps (Fig. 11b–f).
Distribution patterns of rose powdery mildews and the co-distribution with their primary hosts in China. a Distribution of five rose powdery mildews in China. b–f Co-distribution of rose powdery mildews with their primary hosts: bErysiphe rosae, cE. simulans, dPodosphaera pannosa, eP. rosae, fP. rosae-xanthinae (distribution ranges of primary hosts are colored in pink on the maps).
The results show that powdery mildew on Rosa occurs almost nationwide, but different species exhibit distinct distribution patterns (Fig. 11). Erysiphe rosae is distributed in the Xinjiang Autonomous Region, Qinghai Province, and Jilin Province. The distribution of E. simulans is mainly concentrated in southwestern China and is surrounded by the Hengduan Mountains and Qinling Mountains. Podosphaera pannosa is extensively spread across the country, likely due to its wide host range. P. rosae is mainly concentrated in northern China. P. rosae-xanthinae is mainly found in northeast China (Fig. 11f).
Co-distribution analysis, based on primary Rosa hosts, indicates a host-driven distribution of these fungi (Fig. 11b–f). The distributions of P. pannosa and P. rosae are congruent with the ranges of their primary hosts (Fig. 11d, e). Because of the small sample size of E. rosae and P. rosae-xanthinae, meaningful predictions regarding co-distribution phenomena are not yet possible (Fig. 11b, f). However, since almost all sampling points fell within the distribution range of the main hosts, it can be assumed that these species follow the same co-distribution trend. E. simulans has a narrower distribution range than its primary host (Fig. 11c). It is speculated that, in addition to following co-distribution with its host, its distribution pattern is also influenced by geographical barriers, such as the Hengduan and Qinling Mountains.
Discussion
Species diversity of rose powdery mildews in China
Resolving longstanding taxonomic issues clarified and expanded the recognized species diversity of rose powdery mildews. Five species across two genera were identified on Rosa in China. Regarding the genus Erysiphe, sequences from E. rosae provided the first confirmation of this species and the first phylogenetic confirmation that Medusosphaera is a synonym of the genus Erysiphe. Concurrently, this study confirms E. simulans as a single, well-defined species under which the varieties can be removed due to the lack of stable morphological and genetic differentiation. Within the genus Podosphaera, we established a taxonomic benchmark for P. pannosa s. str. through neotypification and reinstated the independent species status of P. rosae, long treated as a synonym of P. pannosa. Moreover, the new species P. rosae-xanthinae sp. nov. is described. These taxonomic results validate the foresight of early mycologists who recognized the species now known as P. rosae (Salmon 1900; Jaczewski 1927; Blumer 1933, 1967; Zheng and Yu 1987), as well as later authors who refined genus- and species-level concepts (Braun and Takamatsu 2000; Bradshaw et al. 2022), while also correcting more than a century of taxonomic confusion largely driven by an overreliance on host identity as a basic diagnostic criterion. More importantly, this discovery clearly indicates that across the vast distribution area of Rosa species in China, the diversity of powdery mildew pathogens cannot be represented by a single species. Therefore, future research and management of rose powdery mildew must extend beyond P. pannosa. When investigating pathogenic mechanisms, screening for disease resistance genes, developing control strategies, and assessing pathogen population dynamics, it is essential to consider the diverse range of pathogenic species, including P. rosae, P. rosae-xanthinae, E. simulans, and E. rosae.
Although Podosphaera rosae and P. rosae-xanthinae have been segregated from the broadly defined P. pannosa species complex, several subclades persist within the P. pannosa clade in the ITS+28S+IGS phylogenetic tree (Fig. 3). This suggests the possible existence of distinct formae or even cryptic species within this taxon. However, relying solely on rDNA sequences is insufficient to explore these questions in depth. While multi-gene sequence analyses are increasingly employed in the taxonomy of powdery mildews and have successfully resolved various taxonomic challenges (Qiu et al. 2020; Bradshaw et al. 2022, 2023, 2025b, 2025c; Zhang et al. 2025), such studies remain limited for the genus Podosphaera (Bradshaw et al. 2025a). Therefore, future multi-gene phylogenetic analyses based on global samples of powdery mildews on Rosa are necessary to clarify this issue.
Evolutionary patterns of rose powdery mildews
This study reveals that the species diversity, host preferences, and geographical distribution of rose powdery mildews are closely correlated with the phylogenetic lineages of their host plants, indicating a co-evolutionary history with their Rosa hosts. Each powdery mildew species exhibited a distinct primary host group, and these host associations were non-random and correlated with the phylogeny of Rosa. For example, within Podosphaera, P. pannosa, P. rosae, and P. rosae-xanthinae primarily infect different Rosa sections (Chinenses + Synstylae, Rosa, and Pimpinellifoliae, respectively). This situation also occurs within Erysiphe. This species–host section correspondence strongly suggests that co-evolution with Rosa hosts has been a primary driver of speciation. These findings are consistent with co-evolutionary patterns documented in other powdery mildew and host systems (Matsuda and Takamatsu 2003; Takamatsu et al. 2010; Guan et al. 2022). Species appear to achieve niche differentiation through specialization on distinct host lineages, thereby reducing interspecific competition. Species within the genus Erysiphe exhibit intergeneric niche overlap with Podosphaera species on some hosts (e.g., sect. Synstylae), indicating that pathogens from different genera may employ different molecular mechanisms to infect the same host group. Alternatively, these pathogens may have co-evolved on a different host genus or species and then jumped to these other hosts.
The distribution analysis further reinforces the co-evolutionary pattern, revealing a host-driven distribution. The distribution of P. pannosa and P. rosae closely matches the ranges of their respective primary host groups. An interesting exception is E. simulans, whose current distribution is much smaller than the modern distribution range of its primary host in sect. Synstylae. This suggests that the origin of E. simulans may coincide with the center of origin for sect. Synstylae (Cheng et al. 2025), but its subsequent dispersal failed to fully keep pace with the radiative evolution of its hosts, potentially limited by geographical barriers such as the Qinling and Hengduan Mountains. However, the distribution of E. simulans in Hubei and Liaoning Provinces indicates that the fungus may be influenced by the expansion of its hosts, gradually expanding its distribution to the east. As a major center of origin and diversity for Rosa, China contains substantial species richness (Luo et al. 2024). The highly diverse powdery mildew taxa revealed in this study, encompassing almost all currently known powdery mildew species infecting Rosa globally (with the exception of E. karisiana, which is known only from its type specimen), further support the notion that host diversity is a key driver of powdery mildew diversity.
However, co-evolution with Rosa is likely not the sole evolutionary force shaping the history of these pathogens. Although diversification appears to track host radiation within Rosa, host jumps at the genus and family levels must also be considered for powdery mildews. In addition to the Rosa hosts, Podosphaera pannosa infects plants from six other genera in five families. These non-Rosa hosts exhibit a phylogenetically discontinuous distribution, a pattern that strongly implies repeated host jumps in their evolutionary history. Notably, the Rosa hosts of P. pannosa are predominantly commercially cultivated ornamental species and cut-flower cultivars that are extensively traded worldwide. As Thines et al. (2023) emphasized, host jumps and geographic expansions have been dramatically accelerated in the Anthropocene by global trade. Therefore, the exceptionally broad host range of P. pannosa likely represents a recent, human-mediated host range expansion. Taken together, the evolutionary history of rose powdery mildews likely exhibits a bimodal pattern. Host–pathogen correspondence at lower taxonomic levels primarily reflects ancient co-speciation, whereas broad and phylogenetically discontinuous host ranges reflect recent, globalization-driven host jumps.
Conclusion
We successfully resolved longstanding taxonomic confusions, revealing that Rosa species host at least five different powdery mildew species across the genera Erysiphe (E. rosae s. lat. and E. simulans) and Podosphaera (P. pannosa s. str., P. rosae comb. nov., and P. rosae-xanthinae sp. nov.) in China. Host preferences and distribution patterns of these fungi indicate that the long-term co-evolutionary relationship between Rosa and their powdery mildews serves as the core driver of pathogen speciation and niche specialization, highlighting the importance of Rosa hosts to the evolutionary history of powdery mildew. Future disease-resistance breeding efforts must account for pathogen diversity by targeting the locally dominant powdery mildew species and tailoring resistant cultivars to specific ecological regions and Rosa species. Furthermore, multi-gene and genomic analyses are needed to more deeply resolve genetic and population-level diversity and to elucidate the co-evolutionary mechanisms underlying host–pathogen interactions.
Supplementary Material
XML Treatment for Erysiphe rosae
XML Treatment for Erysiphe simulans
XML Treatment for Podosphaera pannosa
XML Treatment for Podosphaera rosae
XML Treatment for Podosphaera rosae-xanthinae
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Blumer S (1967) Echte Mehltaupilze (Erysiphaceae) Ein Bestimmungsbuch für die in Europa Vorkommenden Arten. Gustav Fischer Verlag, Jena, Germany, 436 pp.
- 2Božanić Tanjga B, Ljubojević M, Ðukić A et al. (2022) Selection of garden roses to improve the ecosystem services they provide. Horticulturae 8: 883. 10.3390/horticulturae 8100883 · doi ↗
- 3Bradshaw M, Boufford D, Braun U et al. (2024) An in-depth evaluation of powdery mildew hosts reveals one of the world’s most common and widespread groups of fungal plant pathogens. Plant Disease 108: 576–581. 10.1094/PDIS-07-23-1471-RE 37755416 · doi ↗ · pubmed ↗
- 4Bradshaw M, Braun U, Crouch U et al. (2025 a) Phylogeny and taxonomy of the genera of Erysiphaceae, part 8: Podosphaera sect. Tridactyla. Mycologia 118: 116–129. 10.1080/00275514.2025.255455841191582 · doi ↗ · pubmed ↗
- 5Bradshaw M, Braun U, Khodaparast SA et al. (2025 b) Phylogeny and taxonomy of the genera of Erysiphaceae, part 7: Phyllactinieae. Mycologia 117: 640–700. 10.1080/00275514.2025.247637540258175 · doi ↗ · pubmed ↗
- 6Bradshaw M, Braun U, Mitchell JK et al. (2025 c) Phylogeny and taxonomy of the genera of Erysiphaceae, part 6: Erysiphe (the “Microsphaera lineage” part 2). Mycologia 117: 110–165. 10.1080/00275514.2024.238623039495585 · doi ↗ · pubmed ↗
- 7Bradshaw M, Braun U, Pfister DH (2022) Phylogeny and taxonomy of the genera of Erysiphaceae, part 1: Golovinomyces. Mycologia 114: 964–993. 10.1080/00275514.2022.211541936223598 · doi ↗ · pubmed ↗
- 8Bradshaw M, Braun U, Pfister DH (2023) Phylogeny and taxonomy of the genera of Erysiphaceae, part 4: Erysiphe (the “Uncinula lineage”). Mycologia 115: 871–903. 10.1080/00275514.2023.223085337676759 · doi ↗ · pubmed ↗